
Abstract
Mushrooms are important organisms because of their human nutritional and medicinal value. With the expansion of the cultivation of edible mushrooms, fungal diseases have become a major problem in limiting their production. Numerous fungi can cause mushroom deformation or rots. In this publication we report on fungal diseases found during Morchella cultivation in China, with emphasis on morphology and phylogeny to characterise species. The key findings include 1) establishment of a new family Albomorchellophilaceae in Hypocreales, and a novel monotypic genus Albomorchellophila with the type species A. morchellae. Divergence time estimates indicate that Albomorchellophilaceae diverged from its sister family Calcarisporiaceae at ca. 105 (92–120) MYA; 2) the phylogeny and morphology of the family Pseudodiploosporeaceae (Hypocreales) is revised. The family contains a single genus Pseudodiploospora. Intraspecific genetic analyses of Pseudodiploospora longispora reveals significant base differences within strains, especially in the regions of protein-coding genes RPB 2 and TEF; 3) four fungicolous taxa, i.e., Cylindrodendrum alicantinum, Hypomyces aurantius, Hypomyces rosellus, and Trichothecium roseum, are reported as putative pathogens on cultivated morels for the first time. In addition, the previously reported pathogens of morels, Clonostachys rosea, Clonostachys solani, Hypomyces odoratus, and Pseudodiploospora longispora are also detailed in their symptoms and morphology; 4) the phylogeny and morphology of “Zelopaecilomyces” previously placed within Pseudodiploosporeaceae are re-assessed. “Zelopaecilomyces” is proved to be introduced through a chimerism of gene fragments sourced from two distinct organisms. Consequently, it is recommended that “Zelopaecilomyces” should not be recognised due to the mixed up molecular data in phylogeny and a lack of support from morphological evidence. Furthermore, this study discusses the voucher specimen Paecilomyces penicillatus (CBS 448.69), which may contain two mixed taxa, i.e., Pseudodiploospora longispora and a member of Penicillium. Publications on pathogenic fungi of cultivated mushrooms is sporadically, which leads to a lack of understanding of causal agents. As a follow up to the diseases of morel cultivation, we also review the fungal diseases of cultivated mushrooms reported over the last four decades. More than 130 pathogens affect the growth and development of the main cultivated mushrooms. The taxonomic diversity of these pathogens is high, distributed in 58 genera, 40 families, 20 orders, 12 classes and six phyla. The host infected are from Ascomycota to Basidiomycota, mainly being reported from Agaricus bisporus, Cordyceps militaris, Morchella spp., and Pleurotus spp. This study not only enriches our current knowledge on the diversity of pathogens of cultivated mushrooms, especially morels, but also recognizes the importance of some taxa as potential pathogens. Taxonomic investigation and accurate identification are initial and key steps to understanding pathogen-mushroom interactions, and will result in better disease management strategies in the mushroom industry.
Similar content being viewed by others


Introduction
Fungicolous fungi are a large and diverse ecological group which coexist with other fungi (Gams et al. 2004; Põldmaa 2011; Sun et al. 2019a). They currently contain more than 1,500 taxa distributed in many lineages across the fungal kingdom (Sun et al. 2019a). Fungicolous fungi growing consistently with or on other fungi can have many relationships, e.g., mutualistic, commensal, parasitic, and saprotrophic (Gams et al. 2004; Sun et al. 2019a). They are recognized as mycoparasites, saprotrophs, symbionts, and even neutrals (Sun et al. 2019a). This group of fungi has been widely documented and has potential economic and scientific value. Some fungicolous fungi, such as Naematelia aurantialba (a parasite of Stereum hirsutum) and Tremella fuciformis (a parasite of Hypoxylon species) are food sources (Wu et al. 2019). The lobster mushrooms formed after the infection of Hypomyces lactifluorum on mushrooms such as Lactifluus piperatu and Russula brevipes are also consumed as food (Bandoni and Zang 1990; Bandoni and Boekhout 2011; Laperriere et al. 2018). Some fungicolous species have pharmacological effects, and play an invaluable role in drug discovery research; for example, the cosmopolitan parasite Hypomyces chrysospermus was reported to have anticancer and antimetastatic activity (Dikmen et al. 2020). Conversely, certain fungicolous species can be pathogenic to mushrooms and cause diseases on cultivable mushrooms resulting in loss of quality and quantity, which leads to economic losses in commercial mushroom cultivation and indirectly affects food safety (Sun et al. 2019a, b, 2022).
Pathogens on cultivated mushrooms have been studied for over two centuries (Gams et al. 2004; Sun et al. 2019b). They significantly compromise the quality and yield of cultivated mushrooms (Fletcher and Gaze 2008; Sun et al. 2019b). Fungal pathogens affecting mushrooms exhibit their highest diversity within Hypocreales (Sordariomycetes), with Hypomyces (Hypocreaceae) being the largest genus (Põldmaa 2000, 2011; Zhu and Zhuang 2013; Sun et al. 2019a, 2019b). Among these pathogens, some of the most severe diseases in mushroom cultivation include cobweb disease (caused by Hypomyces/Cladobotryum spp.), dry bubble disease (caused by Lecanicillium fungicola), wet bubble disease (caused by Hypomyces perniciosus and Mycogone spp.), and green mold (caused by Trichoderma spp.) (Fletcher and Gaze 2008; Gea et al. 2021). These taxa have been documented to infect Agaricus bisporus, Cordyceps spp., Flammulina velutipes, Hypsizygus marmoreus, Morchella spp., and Pleurotus spp., resulting in the deformation and necrosis of their fruiting bodies (Fletcher and Gaze 2008; Zhang and Tang 2015; Gea et al. 2021; Liu et al. 2021a; Sun et al. 2019b, 2022; Xu et al. 2022). Nonetheless, research on fungal disease related to mushroom cultivation is still in its infancy, and most have focused on the identification of pathogens. However, fungal pathogen identification is mainly based on sequence similarity of ITS and lacks convincing morphological evidence (Sun et al. 2022). Previous phylogenetic studies have verified that SSU and ITS datasets alone were insufficient to provide ideal resolution in some sordariomycetous groups (Hyde et al. 2020; Chethana et al. 2021). This led to some misidentification of pathogens, such as Pseudodiploospora longispora (previously known as Diploospora longispora) and Zelopaecilomyces penicillatus (previously known as Paecilomyces penicillatus) (He et al. 2017, 2018; Sun et al. 2022).
Many studies have been conducted on various fungal diseases on cultivated mushrooms. More than 70 and 20 types of pathogens can infect Agaricus bisporus and Pleurotus spp., respectively, and cause different degrees of damage (summarized by this study, Table 7). In the Morchella cultivation industry, which has only emerged in recent years, some fungal diseases have been reported intermittently since 2016 (Guo et al. 2016; He et al. 2017, 2018; Masaphy 2022; Sun et al. 2022; Liu et al. 2023; Fu et al. 2023). Although some research has been carried out on pathogens, some reports lack good morphological descriptions and phylogenetic support, which undoubtedly increases the difficulty in understanding fungal disease on cultivated mushrooms (Sun et al. 2022). In order to better understand the relationship between pathogens and their hosts, this study uses Morchella as a case study to investigate the species diversity of “molds” on its fruiting bodies, and also summarizes the reported fungal diseases and their causal agents on cultivated mushrooms.
Case study: “Molds” on cultivated morels (Morchella, Pezizales)
True morels (Morchella spp., Morchellaceae, Pezizales) are commercially important edible mushrooms with aromatic and gustatory qualities (Du and Yang 2021). Coupled with their short fruiting season in nature, morel cultivation has become a hot research topic in recent years (Du and Yang 2021). In China, the morel cultivation area reached approximately 16,000 ha in the production season of 2021–2022, and is still expanding (Fig. 1, 2a). With the expanding cultivation scale and density of production, fungal disease has become a major factor limiting the morel yield (Fig. 2b). Typical fungal diseases include cobweb disease caused by Cladobotryum protrusum and C. mycophilum (Lan et al. 2020; Liu et al. 2023), pileus rot disease/white mold disease caused by Clonostachys rosea, Pseudodiploospora longispora and Zelopaecilomyces penicillatus (He et al. 2017, 2018; Sun et al. 2022; Fu et al. 2023), and stipe rot disease caused by the Fusarium incarnatum—F. equiseti species complex (Guo et al. 2016), F. nematophilum (Liu et al. 2021b), and Purpureocillium lilacinum (Masaphy 2022) (Fig. 2b). These fungal diseases occur in most cultivation sites, threatening morel production and causing economic losses.The classification of some fungal pathogens on cultivated morels is confusing, especially the causal agent of “white mold” disease. Diploospora longispora (in Sordariomycetes) and Paecilomyces penicillatus (in Eurotiomycetes) were once considered as serious pathogens that caused pileus rot / “white mold” disease of cultivated morel (He et al. 2017, 2018). These diseases developed similar white mold-like symptoms. Sun et al. (2022) clarified the phylogenetic position of Diploospora longispora and Paecilomyces penicillatus, and the results showed both were affiliated in Hypocreales in Sordariomycetes. Based on morphology and phylogeny of Diploospora longispora and Paecilomyces penicillatus respectively, Sun et al. (2022) introduced a novel family Pseudodiploosporeaceae (Hypocreales) to accommodate them. Although much research has been carried out on the fungal diseases of cultivated morels, the species diversity, origin, transmission, symptoms and pathogenesis of these pathogens have not been confirmed. Additionally, the existing research on fungal diseases of cultivated morels is in infant stage and unsystematic, which undoubtedly increases the difficulty in understanding the pathogen–Morchella interactions. In this study, we investigated the diversity of “mold” diseases caused by fungi on cultivated morels in China, in order to identify the caused agents, find their correct taxonomic placement, and carefully detect the puzzling or erroneous tags associated with sequences in GenBank. We try to answer the following questions: how many fungal diseases are there on cultivated morel? How do fungal pathogens live in association with morels? And are they obligate or facultative?

Healthy fruiting bodies in cultivated morels’ farm in Yunnan, China. a Cultivated Morchella sextelata, b Cultivated Morchella eximia

a Morel cultivation scale in China from 2012 to 2022. b Research history of fungal disease related to cultivated morels
Materials & methods
Sample collections
Infected fruiting bodies of Morchella spp. were collected from China (Guizhou, Shaanxi, Sichuan, and Yunnan Provinces). In total, 38 fungal pathogens from the infected Morchella, and one pathogen from Peziza varia in morels’ farm were isolated successfully (Table 1). Single spore isolations or tissue isolations were carried out following the method described by Senanayake et al. (2020). All materials were deposited in the Cryptogamic Herbarium (KUN-HKAS) of the Kunming Institute of Botany, Chinese Academy of Sciences, and all living cultures were deposited at Kunming Institute of Botany Culture Collection (KUNCC), Yunnan, China. New taxa were established following the guidelines of Chethana et al. (2021) and Maharachchikumbura et al. (2021). Index Fungorum numbers of new taxa were obtained following Index Fungorum (http://www.indexfungorum.org/, 2023). Facesoffungi (FoF) numbers were acquired according to Jayasiri et al. (2015).
Morphological studies
Macro-morphological characteristics of fresh or dried samples were recorded and photographed with a Canon EOS M100 camera. External examinations and free-hand sections were made using a Nikon SMZ 745 T dissecting microscope. Micro-morphological characteristics such as conidiophores, conidiogenous cells/phialides, and conidia were investigated. Hand-sections of mycelium with conidiogenous structure were mounted on slides with sterile water for light microscopy examination using a charge-coupled device SC 2000C attached to a Nikon ECLIPSE Ni-U compound microscope (Model Eclipse Ni-U Nikon Corporation Tokyo, Japan). All measurements were obtained using the Tarosoft (R) Image Framework program (IFW). The photographs were edited in Adobe Photoshop 2018 (Adobe system, USA).
DNA extraction, PCR amplification, and sequencing
Genomic DNA was extracted from fungal mycelium using an Ezup column fungi genomic DNA purification Kit (Sangon Biotech, Shanghai, China) according to the manufacturer’s instructions. Amplification of five selected genes were performed by polymerase chain reaction (PCR). The regions of internal transcribed spacer (ITS), the partial large-subunit ribosomal RNA (LSU), the partial small-subunit ribosomal RNA (SSU), the second largest subunit of RNA polymerase II (RPB 2), and the translation elongation factor 1-alpha (TEF) were amplified with the primer pairs of ITS1-F/ITS4 (White et al. 1990; Gardes and Bruns 1993), LR0R/LR5 (Vilgalys and Hester 1990), PNS1/NS41 (O’Donnell et al. 1997, 1998), RPB 2-5F/RPB 2-7cR (Liu et al. 2000), and EF1-983F/EF1-2218R (Rehner and Buckley 2005, D'Alessandro et al. 2014). Total reaction mixtures (25 μl) contained 21 μl of 1.1 × T3 Super PCR master mix (Tsingke Biotechnology Co., Ltd., Beijing, China), 2 μl DNA template, and 1 μl of each primer. Reactions included an initial denaturation condition were 98 °C for 5 min, followed by 35 cycles of denaturation at 98 °C for 20 s, annealing at 53 °C for 30 s for ITS, SSU, LSU, and RPB 2 genes, 64 °C for 40 s for TEF, followed by extension at 72 °C for 45 s, and a final extension at 72 °C for 10 min. PCR products were sequenced by Sangon Biotech (Shanghai) Co., Ltd. (China). Newly generated sequences were deposited in NCBI GenBank with the corresponding unique accession numbers (Supplemental Table 1).
Phylogenetic analyses
Phylogenetic trees were constructed using our sequencing data and their respective allied reference sequences downloaded from the GenBank (Supplemental Table 1). Individual data sets of ITS, LSU, SSU, TEF and RPB 2 were assembled, and aligned using the default settings of MAFFT v. 7 online Server (https://mafft.cbrc.jp/alignment/server/) (Kuraku et al. 2013; Katoh et al. 2019), and manually edited where necessary in BioEdit version 7.0.9 (Hall 1999). Ambiguous regions were excluded from the analyses, and gaps were treated as missing data. The maximum likelihood (ML) analysis was performed at the IQ-Tree web portal (http://iqtree.cibiv.univie.ac.at/). The substitution model options for each gene were auto-evaluated according to the provided partition file. Clade support for the ML analysis was assessed using an SH-aLRT test with 1,000 replicates and the ultrafast bootstrap (UFB) (Guindon et al. 2010; Hoang et al. 2018). Nodes with support values of both SH-aLRT ≥ 80 and UFB ≥ 95 were considered well-supported, nodes with one of SH-aLRT < 80 or UFB < 95 were weakly supported, and nodes with both SH-aLRT < 80 and UFB < 95 were considered unsupported (Guindon et al. 2010).
Bayesian inference (BI) analysis was calculated in MrBayes v. 3.2.2 (Ronquist et al. 2012). The appropriate nucleotide substitution model for LSU, ITS, TEF, and RPB 2 were tested via the Akaike information criterion (AIC) with jModelTest2 on XSEDE in the CIPRES Science Gateway web server (Posada 2008). Four simultaneous Markov Chains were run for 2,000,000 generations. Markov Chain Monte Carlo sampling (MCMC) analysis started from a random tree that was sampled every 1,000 generations. The average standard deviation < 0.01 for split frequencies was used to suggest a convergence between parallel runs. The first 25% of total trees were discarded as burn-in, and the remaining trees in each analysis were utilized to suggest calculating posterior probabilities (PP) in the majority-rule consensus tree. PP ≥ 0.95 is considered indicative of strong support.
A neighbor joining (NJ) phylogenetic tree was constructed based on genetic distances, which were calculated using Kimura 2-parameter model (K2P) (Kimura 1980) in MEGA-X (Kumar et al. 2018). Phylogenetic trees were viewed in FigTree v.1.4.4. Genetic distances of SSU, ITS, LSU, TEF, and RPB 2 sequences of Pseudodiploospora longispora were calculated with the K2P model using MEGA-X software (Kumar et al. 2018).
Species delimitation analyses
Genealogical concordance phylogenetic species recognition (GCPSR) was employed to investigate species limits in Pseudodiploospora longispora. To identify independent evolutionary lineages under the GCPSR, we followed two criteria based on Dettman et al. (2003): a) genealogical concordance, the clade was present in the majority (75%) of the single-locus genealogies, and b) genealogical non-discordance, the clade was well supported in at least one single-locus genealogy, as judged by NJ-BP ≥ 0.75 for NJ, and was not contradicted in any other single-locus genealogy at the same support level. The ITS, LSU, TEF and RPB 2 genealogies were visually compared to find concordance. Exhaustive subdivision was used to decide which independent evolutionary lineages represented phylogenetic species, which means that all individuals had to be placed within a phylogenetic species (Dettman et al. 2003). If an individual was not included in one of the lineages, the nodes of the tree were traced down from that the individual who is not surrounded by thick branches until all individuals were included in a clade surrounded by a thick branch, and recognized such clades as phylogenetic species (Dettman et al. 2003; Wang et al. 2016b).
Calibration, divergence time, and evolutionary rate estimations
One fossil and a secondary calibration were used for the divergence time estimates of new taxa in the Hypocreales. The fossil Paleoophiocordyceps coccophagus is nested in Ophiocordyceps (Ophiocordycipitaceae, Hypocreales) due to its morphology resembles the asexual states of Hirsutella and Hymenostilbe (Cruickshank and Ko 2003). The age of the fossil has been calculated around 99–105 million years ago (MYA) based on the geological timescale (Cruickshank and Ko 2003). In this study, the fossil P. coccophagus evidence was used for the calibration of the crown node of Ophiocordyceps with an exponential distribution, offset = 100, mean = 27.5, and 95% credibility interval = 182.4 MYA (Sung et al. 2008). The crown age of Xylariomycetidae with a normal distribution (mean = 168 MYA, and SD = 16) was used as the secondary calibration point (Hongsanan et al. 2016; Samarakoon et al. 2016, 2022).
Divergence time estimates were carried out by BEAST v. 2.7.0 (Bouckaert et al. 2014, 2019). Aligned sequence data were partitioned separately for LSU, ITS, TEF, and RPB 2 datasets, and loaded to prepare an XML file constructed in BEAUti v. 2.7.0. The substitution model selected GTR, which was applicable to all gene segments. The strict clock model and the Calibrated Yule tree were used for the analyses. The analysis was performed for 80,000,000 generations using BEAST v. 2.7.0, obtaining logging parameters and trees for every 1,000 generations. The effective sample sizes (ESS) of parameters were checked in Tracer v.1.6 (Rambaut et al. 2013), and the acceptable values were greater than 200. The first 25% of trees were discarded based on the ESS values, and the remaining trees were used to generate a maximum clade credibility tree by using TreeAnnotator v1.10.4. The geographical timescale was followed as in Walker (2019).
Results
Phylogenetic analyses
Newly generated fungal pathogens on cultivated morels
The combined LSU + ITS + TEF + RPB 2 dataset consists of 271 taxa, representing 25 families in Hypocreales, two families of Sordariales, and three orders in Xylariomycetidae (Fig. 3). Of these, taxa of Sordariales and Xylariomycetidae were selected as outgroup. The final alignment comprised 4,148 characters (LSU: 1–921, ITS: 922–2,132, TEF: 2,133–3,044, and RPB 2: 3,045–4,148) including alignment gaps. The best-fit model according to AIC: GTR + I + G was for LSU, ITS, TEF and RPB 2. The combined alignment contained 1,928 parsimony-informative characters, 1,844 constant characters, and 376 singleton characters. Phylogenetic analyses obtained from ML and BI analyses resulted in trees with similar topologies. The ML tree with a final log-likelihood of -112,303.999 was shown in Fig. 3.


Phylogram generated from maximum likelihood analysis based on combined LSU, ITS, TEF and RPB 2 sequence data. Bootstrap support values for ML ≥ 80 of SH-aLRT or 95 of UFB and posterior probability for BI ≥ 0.95 are indicated above the nodes and separated by “–/–/–” (SH-aLRT/UFB/PP). Newly collected samples in this study are given in red
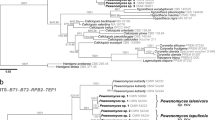
In phylogenetic tree (Fig. 3), our newly generated sequences were distributed in six lineages, i.e., Albomorchellophilaceae (new family), Bionectriaceae, Hypocreaceae, Myrotheciomycetaceae, Nectriaceae, and Pseudodiploosporeaceae. Stains KUNCC21-10005 and KUNCC21-10100 formed a sister lineage with Calcarisporiaceae (SH-aLRT = 100, UFB = 100, PP = 1). A new family Albomorchellophilaceae, with the type species Albomorchellophila morchellae gen. et sp. nov. is erected here. In Bionectriaceae, two known species in Clonostachys were identified successfully, there are C. rosea (KUNCC21-10000, KUNCC21-10001, and KUNCC21-10002) and C. solani (KUNCC21-10003 and KUNCC21-10004). Eight strains were placed in Hypocreaceae, and belonged to three known species of Hypomyces, i.e., H. aurantius (KUNCC21-10012, KUNCC21-10013, and KUNCC21-10014), H. odoratus (KUNCC21-10006 and KUNCC21-10007), and H. rosellus (KUNCC21-10009, KUNCC21-10010 and KUNCC21-10011). The strain KUNCC21-10015 clustered with the common pathogen Trichothecium roseum in Myrotheciomycetaceae with a SH-aLRT = 100, UFB = 99 and PP = 1 support value. In Nectriaceae, three strains KUNCC21-10016, KUNCC21-10017, and KUNCC21-10018 clustered together, and identified as Cylindrodendrum alicantinum with a SH-aLRT = 96.8, UFB = 100 and PP = 1 support value. In addition, 20 strains were placed in Pseudodiploosporeaceae, which divided into five distinguishable groups (groups I–V) in Pseudodiploospora. However, five evolutionary lineages were not supported by GCPSR, exhaustive subdivision, and morphology (see the next analyses of P. longispora), so they were considered to be the same species, and consistent with Pseudodiploospora longispora.
The genetic divergence comparisons in Pseudodiploospora longispora
The 20 strains isolated in this study, as well as three Paecilomyces penicillatus samples (BH, BHJ, and mp-9) in He et al. (2017), two Diploospora longispora samples (60319 and 60320) in He et al. (2018), three Pseudodiploospora longispora samples (CGMCC 3.23768, CGMCC 3.23769 and CGMCC 3.23771) in Sun et al. (2022), and the sample CBS 448.69 named Zelopaecilomyces penicillatus (Sun et al. 2022), constituted a monophyletic lineage but with obvious base differences (Fig. 3). In Fig. 3, these 29 strains were divided into five distinguishable subgroups, i.e., group I–V. Their evolutional lineage boundaries were evaluated by a (phylo-) genetic distance matrix calculation for the markers ITS, LSU, SSU, TEF and RPB 2, respectively. The barcoding gap between interspecific and intraspecific distances is graphed based on the K2P model for each individual gene. The results indicated that the gene RPB 2 showed the highest intraspecific genetic distances, while no intraspecific variability is found in SSU region (Table 2). In addition, there was an obvious barcoding gap between the genetic distances within and between groups of RPB 2, followed by sequence data for the ITS, TEF, and LSU (Table 2, 3).
Examination of the RPB 2 sequence data in 23 samples [20 strains isolated from this study and three Pseudodiploospora longispora samples CGMCC 3.23768, CGMCC 3.23769 and CGMCC 3.23771 in Sun et al. (2022)] showed that 9% (8/89) of the base mutations in the open reading frame region (ORF) were located in the first position of codon base, while 91% (81/89) were located at the third position of codon base. These base differences lead to synonymous mutations, except for the 601st base mutation, which leads to amino acid mutation from I to V or from V to I. In TEF sequence data, there were 31 base mutations in the ORF region, of which six were located in the first position of codon base, three in the second position of codon base, and 22 in the third position of codon base. These mutations resulted in eight amino acid changes, all caused by the mutations of the first two types of bases.
Intrageneric relationship of Pseudodiploospora longispora
Supported conflict were detected between the individual gene phylogenies (ITS, LSU, TEF and RPB 2) for combined analyses in terms of species discrimination of Pseudodiploospora longispora (Fig. 4, Table 4). In the Neighbor-joining (NJ) analyses of ITS, LSU and RPB 2 datasets, KUNCC21-10028 and KUNCC21-10029 of group II formed a moderately to strongly supported monophyletic group (ITS: NJBP = 0.618, LSU: NJBP = 0.88, RPB 2: NJBP = 1). However, KUNCC21-10028 and KUNCC21-10029 were non-monophyletic in TEF phylogeny. In group III, three strains KUNCC21-10030, KUNCC21-10031, and KUNCC21-10032 constituted a monophyletic clade that was strongly supported by ITS (NJBP = 0.98), TEF (NJBP = 1), and RPB 2 (NJBP = 1). In group IV, strains KUNCC21-10033, KUNCC21-10034, KUNCC21-10035, and KUNCC21-10036 were grouped as an independent lineage, and strongly supported by LSU (NJBP = 0.90) and RPB 2 (NJBP = 1), but the placement of these four samples were in conflict in the ITS and TEF phylogenies. In group V, independent branch containing KUNCC21-10037, KUNCC21-10038, and KUNCC21-10039 was strongly supported by ITS (NJBP = 0.84), TEF (NJBP = 1), and RPB 2 (NJBP = 1), but it showed some conflicts in LSU phylogeny.

Neighbor-joining (NJ) phylograms of ITS, LSU, TEF, and RPB 2, of Pseudodiploospora longispora taxa, with the tree midpoint rooted. Distances are based on the K2P model. Estimates of topology stability (bootstrap test; 1,000 replicas) are given above the nodes
Species recognition of Pseudodiploospora longispora using GCPSR
Seven independent lineages were determined for the intraspecific species in Pseudodiploospora by GCPSR (thick branches in Fig. 5). Three of them (indicated by triangles) met the criteria of GCPSR, i.e., group II (KUNCC21-10028 and KUNCC21-10029), group III (KUNCC21-10030, KUNCC21-10031 and KUNCC21-10032) and group V (KUNCC21-10037, KUNCC21-10038 and KUNCC21-10039) (Table 4). However, combined with the exhaustive subdivision, none of the five groups met the conditions of species delimitation. Considering GCPSR and exhaustive subdivision, these 23 strains belong to the same species, although there are obvious base differences, and all of them are identified as Pseudodiploospora longispora.

Neighbor-joining (NJ) phylograms of combined ITS + LSU + TEF + RPB 2 of Pseudodiploospora longispora taxa, with the tree midpoint rooted. Distances are based on the K2P model. Estimates of topology stability (bootstrap test; 1,000 replicas) are given in nodes. The thick branches indicate seven independent lineages of P. longispora, and the triangles at the nodes are three evolutional lineages recognized by genealogical concordance phylogenetic species recognition
Divergence time estimation
In the divergence time analysis (Fig. 6), the stem age estimate is in Carboniferous (323 MYA with 95% CI of 297–352), and its crown age was in Permian (263 MYA with 95% CI of 241–287) in Hypocreales. The family Albomorchellophilaceae diverged from Calcarisporiaceae at 105 (92–120) MYA. According to the recommendations provided by Hyde et al. (2017), the stem age for a family fall within 50–130 MYA. The divergence times of Albomorchellophilaceae falls within the family range, and thus it’s introduced as a new family.

Maximum clade credibility (MCC) tree with divergence time estimates for Hypocreales. Divergence time (MYA) and posterior probabilities (PP) are indicated above and below the nodes, respectively. Bars correspond to the 95% highest posterior density (HPD) intervals. An asterisk (*) in the red indicates the fossil calibration Paleoophiocordyceps coccophagus that nested in Ophiocordyceps (Ophiocordycipitaceae), and blue asterisk indicates the secondary calibration point at the crown node of Xylariomycetidae
Taxonomy
Hypocreales Lindau.
Albomorchellophilaceae F.M. Yu, K.D. Hyde & Q. Zhao, fam. nov.
Index Fungorum number: IF901520; Facesoffungi: FoF 15281.
Etymology: Named after the type genus, Albomorchellophila.
Diagnosis: in addition to the phylogenetic distinctions (Fig. 3), Albomorchellophilaceae differs from other families in Hypocreales by its degenerated conidiophores, and the way of sporulation.
Fungicolous or saprobic. Asexual morph: Sporulation in the aerial mycelium, flask-shaped conidiogenous cells, sometimes conidiogenous cells inconspicuous, with narrow, inconspicuous scars on the surface of mycelium. Conidia ovoid, hyaline, smooth walled. Sexual morph: not observed.
Type genus: Albomorchellophila F.M. Yu, K.D. Hyde & Q. Zhao.
Distribution and habitat: China, Yunnan Province, on the fruiting bodies of cultivated Morchella.
Albomorchellophila F.M. Yu, K.D. Hyde & Q. Zhao, gen. nov.
Index Fungorum number: IF901517; Facesoffungi: FoF 15282.
Etymology: Name composed of Latin “albo-” (white) + the favorite natural substrate genus Morchella + Latin “-phile” (loving).
Diagnosis: in addition to the phylogenetic distinctions (Fig. 3), Albomorchellophila differs from Calcarisporium (Calcarisporiaceae) by conidiophores degenerating into conidiogenous cells, the presence of phialidic conidiogenous cells, and the ellipsoidal, oval to subglobose conidia.
Fungicolous or saprobic. Colonies fluffy, whitish. Mycelia partially superficial, composed of branched, septate, and hyaline hyphae. Asexual morph: Conidiophores micronematous, usually reduced to conidiogenous cells, arising terminally or laterally from hypha, hyaline, smooth-walled. Conidiogenous cells holoblastic, polyblastic, terminal and intercalary in hypha, hyaline, phialidic. Conidia aseptate, smooth-walled, ellipsoidal, oval, subglobose. Sexual morph: not observed.
Type species: Albomorchellophila morchellae F.M. Yu, K. D. Hyde & Q. Zhao.
Distribution and habitat: China, Yunnan Province, on the fruiting bodies of cultivated morels.
Notes: Albomorchellophila is introduced here based on phylogenetic and morphological evidence. Albomorchellophila is sister to Calcarisporium (Calcarisporiaceae) with high support values (SH-aLRT = 99, UFB = 100, PP = 1) in our phylogeny (Fig. 3). It can be distinguished from Calcarisporium by the presence of phialidic conidiogenous cells, and conidiophores degenerated into conidiogenous cells.
Albomorchellophila morchellae F.M. Yu, K.D. Hyde & Q. Zhao sp. nov. ……………….………Fig. 7

Albomorchellophila morchellae (KUN-HKAS 129638). a, b White colony on dried fruiting body of Morchella sp. c, d Mycelia on the surface of morel’s cap. e Hyphae with conidia. f Conidia. g–i Conidiogenous structure and conidia. l Colony of A. morchellae (KUNCC21-10005) on PDA at room temperature after 15 days. Scale bars: e = 100 μm; g, h = 50 μm; f, i = 10 μm; l = 30 mm
Index Fungorum number: IF901516; Facesoffungi: FoF 15283.
Etymology: Name bases on the host Morchella.
Diagnosis: the main diagnostic criteria of Albomorchellophila morchella are abundant ellipsoidal, oval, subglobose conidia, lack of conidiophores, and conidiogenous cells phialidic, verticillate or sympodial, most solitary or 2 to 3, and absent pigment.
Holotype: KUN-HKAS 129638.
Fungicolous, growing on the fruiting bodies of cultivated morels. Asexual morph: Hyphomycetous. Colonies on natural substrate loose floccose, white, often confluent and forming irregular patches. Mycelium partly superficial, white, composed of septate, branched, floccose masses of hyaline hyphae, with yellow or brownish reverse. Hyphae colourless, separate, with abundant conidiogenous loci, up to 4 μm diam. Conidiophores micronematous, usually reduced to conidiogenous cells directly, arising terminally or laterally from hypha. Conidiogenous cells, phialidic, hyaline, smooth-walled, verticillate or sympodial, most solitary or 2 to 3, 4–10 × 1–2.5 μm (n = 30). Conidia borne on hyphae, phialide, and side branches, one-celled, hyaline, smooth-walled, ellipsoidal, oval, subglobose, 3.5–8.5 × 3–5 μm (n = 30). Sexual morph: not observed.
Material examined: China, Yunnan Province, Kunming City, on the fruiting bodies of Morchella sp. (Morchellaceae, Pezizales), 12 March 2022, Qi Zhao, ZHAO3647 (KUN-HKAS 129638, holotype, ex-type living-culture: KUNCC21-10005); Yunnan Province, Kunming City, on the fruiting bodies of Morchella sp. (Morchellaceae, Pezizales), 16 May 2023, Feng-Ming Yu, 23MP-13 (KUN-HKAS 129639, living-culture: KUNCC21-10100).
Notes: Albomorchellophila morchellae was isolated from infected fruiting bodies of cultivated morels. It caused white spots on the ridges and pits, forming “white mold” like symptom. In the phylogeny (Fig. 3), A. morchellae forms a separate clade and is sister to members of Calcarisporium in Calcarisporiaceae. The nucleotide comparison (including gaps) between Albomorchellophila morchellae (KUN-HKAS 129638) and the type species Calcarisporium arbuscula (CBS 900.68) showed 50 bp (9.56%) differences across 523 bp ITS, 24 bp (2.88%) differences across 833 bp LSU, 3 bp (0.29%) differences across 1,018 bp SSU, 79 bp (8.64%) differences across 914 bp TEF, and 146 bp (17.51%) differences across 834 bp RPB 2, respectively. Albomorchellophila morchellae can be distinguished from species of Calcarisporium by its conidiophores degenerating into conidiogenous cells, phialidic conidiogenous cells, and ellipsoidal, oval to subglobose conidia (Sun et al. 2017). Most Calcarisporium species were reported as fungicolous, and can appear on the fruiting bodies of Cordyceps cordycipiticola, Cordyceps militaris, Hirsutella citriformis, rust fungi, and xylarialean taxa (Sun et al. 2016, 2019a; Liu et al. 2022). The related host/substrate of Albomorchellophila needs further study.
Bionectriaceae Samuels & Rossman, Stud. Mycol. 42: 15 (1999).
Clonostachys Corda, Pracht-Fl. Eur. Schimmelbild.: 31 (1839).
Clonostachys rosea (Link) Schroers, Samuels, Seifert & W. Gams, Mycologia 91(2): 369 (1999) ………………………………………………………………Fig. 8

Clonostachys rosea (KUN-HKAS 129648). a White colony on the fruiting body of Morchella eximia. b, c Mycelia on the surface of morel’s cap. d–k Conidiophores and conidiogenous cells. l, m Conidia. n Colony of C. rosea (KUNCC21-10000) on PDA at room temperature after 10 days. Scale bars: d = 100 μm; e–l = 50 μm; m = 20 μm; n = 50 mm
Index Fungorum number: IF 461067; Facesoffungi number: FoF 15284.
Fungicolous, growing on cotton layer on the fruiting bodies of Morchella sextelata. Asexual morph: Conidiophores verticillium-like, or solitary or aggregated, variable in length, 2.5–4 μm at the widest point, 1.5–3 μm at the tip. Penicillus divergent, in whorls of 2–5, straight or slightly curved, generally slightly tapering towards the tip, 15.5–46 μm long, 1.5–3.5 μm wide at the base, 1–2.5 μm at the tip. Conidia hyaline, minutely curved, distally broadly rounded or slightly tapering, 5.5–13.5 × 2.5–5 μm (n = 30). Sexual morph: Not observed.
Material examined: China, Shaanxi Province, Hanzhong City, on fruiting body of cultivated Morchella eximia, 04 March 2022, Qi Zhao, HZ-1 (KUN-HKAS 129648, living-culture: KUNCC21-10000); Shaanxi Province, Hanzhong City, on fruiting body of cultivated Morchella eximia, 04 March 2022, Qi Zhao, HZ-3 (KUN-HKAS 129649, living-culture: KUNCC 21–10001); Guizhou Province, Qianxi City, on a fruiting body of cultivated Morchella sextelata, 11 March 2022, Feng-Ming Yu, QX1-4 (KUN-HKAS 129650, living-culture: KUNCC 21–10002).
Notes: Clonostachys rosea is a commonly reported species, which exists in many habitats with the highest frequency in soil (Schroers 2001). As an excellent mycoparasite, C. rosea has a wide range of hosts, as plant pathogens and saprobes, and can kill hosts with its special enzymes and anti-fungal chemicals (Sun et al. 2019a). Its variant Clonostachys rosea f. catenulata was reported to parasitize the sclerotium of Ophiocordyceps sinensis (Sun et al. 2019b). Fu et al. (2023) reported C. rosea infected 30% fruiting bodies of Morchella sextaleta in Anhui Province, China, and formed a serious rot disease. In this paper, C. rosea was isolated from the fruiting bodies of cultivated morels in Guizhou and Shaanxi Provinces. It formed white or pale-yellow cotton-like mycelia on the ridges and pits of cultivated morels. The infection, colonization and expansion of C. rosea led to withering and decay of morel, which resulted in malformed fruiting bodies.
Clonostachys solani (Harting) Schroers & W. Gams, Stud. Mycol. 46: 111 (2001) ……….………Fig. 9

Clonostachys solani (KUN-HKAS 129651). a Colony on the stipe of Morchella sextelata. b, c Mycelia on the surface of morel’s stipe. d, e Conidiophores and conidiogenous cells. f Conidia. g, h Colony of C. solani (KUNCC21-10003) on PDA at room temperature after 10 days. Scale bars: d, e = 50 μm; f = 10 μm; g, h = 30 mm
Sexual morph: Bionectria solani (Reinke & Berthold) Schroers, Stud. Mycol. 46: 111 (2001).
Index Fungorum number: IF 456098; Facesoffungi number: FoF 15285.
Fungicolous, growing on the fruiting bodies of Morchella sextelata. Asexual morph: Conidiophores adpressed, solitary to aggregated and sporodochial, beanched, 15.5–33 μm long, 2–4 μm wide, variable in length; phialides in apical whorls of 2–5, loose or adpressed, straight to slightly curved, flask-shaped to cylindrical, generally slightly tapering towards the apex, without a visible collarette, 9.5–35 μm long, 2–3.5 μm wide at widest point, 1.5–2 μm at the tip. Conidia hyaline, straight to slightly curved, distally broadly rounded, with or without recognizable hilum, variable in shapes, reniform, allantoid, inequilateral, oval, and ellipsoid, 5–8 × 2.4–4 μm (n = 30). Sexual morph: not observed.
Material examined: China, Yunnan Province, Kunming City, Wuhua District, on the stipe of Morchella sextelata (Morchellaceae, Pezizales), 02 April 2021, Feng-Ming Yu, 402-1S (KUN-HKAS 129651, living-culture: KUNCC21-10003); Yunnan Province, Kunming City, Wuhua District, on the stipe of Morchella sextelata (Morchellaceae, Pezizales), 16 March 2021, Feng-Ming Yu, GK-2S (KUN-HKAS 129652, living-culture: KUNCC21-10004).
Notes: Clonostachys solani is frequently encountered in tropical and temperate regions, and mainly on bark of recently dead trees and on various other plant material, including different parts of potato plants or from soil (Schroers 2001). Through ITS amplicon sequencing on the morel ascocarp lesions, Shi et al. (2022) reported that C. solani was the second largest putative pathogen causing fungal diseases of Morchella, and its detection rate was 5.04%, second only to that of Pseudodiploospora longispora (75.48%). In this study, two samples of C. solani were isolated from the infected morels’ fruiting bodies in Yunnan Province, China. This species was found infecting the stipe of the morel, forming a spot, covered with white hyphae with a dark green center (Fig. 9).
Hypocreaceae De Not.
Hypomyces (Fr.) Tul. & C. Tul.
Hypomyces aurantius (Pers.) Fuckel, Jb. nassau. Ver. Naturk. 23–24: 183 (1870) [1869–70] ……Fig. 10

Hypomyces aurantius (KUN-HKAS 129646). a–d White colony on the fruiting body of Morchella sextelata. e–f Mycelia on the surface of morel’s cap. g–h Conidiophores, conidiogenous cells, and conidia. i Conidia. j Colony of H. aurantius (KUNCC21-10012) on PDA at room temperature after 10 days. Scale bars: g–i = 50 μm; j = 50 mm
Asexual morph: Cladobotryum varium Nees, Syst. Pilze (Würzburg): 56 (1816) [1816–17].
Index Fungorum number: IF 192954; Facesoffungi number: FoF 06013.
Fungicolous, colonizing on the fruiting body of Morchella sextalata. Asexual morph: Conidial fructifications develop as floccose masses of white to yellowish mycelium bearing conidiophores with abundant white to yellowish spore masses. Conidiophores arising from hyphae, micronematous, branched, septate, 1–3-verticillate, with terminal whorl of 2–6 phialides. Conidiogenous cells monoblastic, phialidic, subulate, tapering toward apex, smooth, 29–53 × 4–8.5 μm (n = 30). Conidia hyaline, broadly ellipsoid, with big truncate basal hilum, 0–1-spetate, constricted at the septa, smooth, 8.5–17.5 × 6–10.5 μm (n = 30). Sexual morph: not observed.
Material examined: China, Guizhou Province, Qianxi City, on fruiting bodies of Morchella sp. (Morchellaceae, Pezizales), 11 March 2022, Feng-Ming Yu, QX1-7 (KUN-HKAS 129646, living-culture: KUNCC 21–10012), QX2-8 (KUN-HKAS 129645, living-culture: KUNCC 21–10013). Yunnan Province, Kunming City, on fruiting bodies of Morchella sp. (Morchellaceae, Pezizales), 17 February 2022, Feng-Ming Yu, 22MP-42 (KUN-HKAS 129647, living-culture: KUNCC 21–10014).
Notes: Hypomyces is the largest fungicolous genus, and its members parasitize other fungi from temperate to tropical regions (Rogerson and Samuels 1989, 1993, 1994; Põldmaa 2000, 2011; Tamm and Põldmaa 2013; Sun et al. 2019b). Some Hypomyces species were reported as the causing-agent of cobweb disease of cultivated mushrooms. Hypomyces aurantius, commonly known as the orange polypore mold, was reported to be parasitic on a variety of basidiomycetes in nature, especially on polyporous fungi, such as Microporus sp., Polyporus sulphureus, and Trametes versicolor (Sun et al. 2019a). Sun et al. (2019b) provided a new record of Hypomyces aurantius growing on Panellus sp. in China. This species was also reported on cultivated mushrooms, such as, Agaricus bisporus, Flammulina velutipes, Hypsizigus marmoreus, and Pleurotus eryngii (Table 7). However, there was no record of H. aurantius parasitic on cultivatable ascomycetous mushrooms. In this study, the asexual morph of H. aurantius was isolated from infected morels. In morel cultivation bases, white to yellowish fluffy mycelia appeared on the surface of morels’ ascocarp and surrounding soils. The symptoms caused by H. aurantius were varied and depended upon the developmental stage of morel at the time of infection. At an early stage, the white hyphae of mycoparasite covered the morels’ primordium and young fruiting body. When affected in later stages, morels’ stipes were often imperfectly formed and became rotten. Usually, infection of the stipes was more serious than that of their caps, which indicated that this pathogen might originate from the soil.
Hypomyces odoratus G.R.W. Arnold, Česká Mykol. 18(3): 144 (1964) …..……………………..Fig. 11

Hypomyces odoratus (KUN-HKAS 129643). a White colony on the fruiting body of Morchella sextelata. b–d Mycelia on the surface of morel’s cap. e–i Conidiophores and conidiogenous cells. j Conidia. k Colony of H. odoratus (KUNCC21-10006) on PDA at room temperature after 10 days. Scale bars: e–j = 50 μm; k = 20 mm
Asexual morph: Cladobotryum mycophilum (Oudem.) W. Gams & Hooz., Persoonia 6(1): 102 (1970).
Index Fungorum number: IF 332444; Facesoffungi number: FoF 15286.
Fungicolous, colonizing on the fruiting body of Morchella sextelata. Asexual morph: Mycelium white to pinkish yellow on nature substance, partly superficial, developing as floccose masses of septate, branched, smooth and hyaline hyphae bearing robust conidiophores with white spore masses. Conidiophores hyaline, erect, with branches arranged in verticillate whorls, in pairs or/and singly, indefinite in length, 1–4-verticillate, with terminal whorl of 1–6 phialides. Conidiogenous cells polyblastic, phialidic, hyaline, subulate, tapering toward apex, smooth, 27–105.5 μm long, 4–6.5 μm at the base (n = 30). Conidia hyaline, smooth, 0–3-septate, ellipsoid, obovoid, ovoid, sometimes slightly curved, with a rounded tip and a basal hilum, 12.5–31.5 × 5.5–11 μm (n = 30). Sexual morph: not observed.
Material examined: China, Yunnan Province, Kunming City, on fruiting bodies of cultivated Morchella sextelata, 16 March 2021, Feng-Ming Yu, gk-3c (KUN-HKAS 129643, living-culture: KUNCC21-10006), gk-5 (KUN-HKAS 129644, living-culture: KUNCC21-10007).
Notes: The asexual morph of Hypomyces, previous known as Cladobotryum, as the causative agent of cobweb disease, is responsible for significant economic losses in the mushroom industry (Table 7). To date, several species of Hypomyces/Cladobotryum, H. aurantius (asexual morph: C. varium), H. rosellus (asexual morph: C. dendroides), C. protrusum (sexual morph: unknown), C. mycophilum (sexual morph: unknown), C. multiseptatum (sexual morph: unknown), and C. verticillatum (sexual morph: unknown) have been identified as fungal pathogens on cultivated mushrooms (Table 7). Their incidence is increasing at an alarming rate, causing serious damage. Among these pathogens, Hypomyces odoratus (asexual morph: C. mycophilum) has been found frequently on Agaricus bisporus and oyster mushrooms during the last two decades (Table 7). Lan et al. (2020) first reported that C. protrusum caused cobweb disease on cultivated Morchella importuna. Later, Liu et al. (2023) provided the genomic characterization of H. odoratus isolated from M. sextelata. In this study, Hypomyces odoratus was isolated and identified from infected fruiting bodies of morels in Yunnan Province, China. The symptoms were white or yellowish cobweb-like mycelia growing and covering the surface of morels’ stipe and surrounding soil, and quickly spread to the whole fruiting body, causing it to rot.
Hypomyces rosellus (Alb. & Schwein.) Tul. & C. Tul., Annls Sci. Nat., Bot., sér. 4 13: 12 (1860) ……………………………………………………………………………Fig. 12

Hypomyces rosellus (KUN-HKAS 129642). a, b White colony on the fruiting body of Morchella sextelata. c, d Mycelia on the surface of morel’s cap. e–k Conidiophores and conidiogenous cells. l Conidia. m Colony of H. rosellus (KUNCC21-10009) on PDA at room temperature after 10 days. Scale bars: e–g = 100 μm; h–l = 50 μm; m = 20 mm
Asexual morph: Cladobotryum dendroides (Bull.) W. Gams & Hooz., Persoonia 6(1): 103 (1970).
Index Fungorum number: IF 212420; Facesoffungi number: FoF 06015.
Fungicolous on the fruiting body of Morchella sextelata. Asexual morph: Conidial fructifications develop as loosely floccose masses of white to pinkish yellow mycelium bearing robust conidiophores with white spore masses. Conidiophores hyaline, erect, septate, branched, 1–4-verticillate, with terminal whorl of 2–6 phialides. Conidiogenous cells polyblastic, phialidic, hyaline, 21.5–54 × 4–7.5 μm (n = 30) wide basally, and tapering slightly to the tip. Conidia hyaline, subcylindrical, ellipsoid, rarely elongate ovoid, slightly curved at base each having a broad, blunt basal apiculus and appearing broadly rounded at the apex, commonly somewhat constricted at the septa, 0–3-septate, smooth-walled, 16.5–31.5 × 8–12.5 μm (n = 30). Sexual morph: not observed.
Material examined: China, Guizhou Province, Qianxi City, on fruiting bodies of cultivated Morchella sextelata, 11 March 2022, Feng-Ming Yu, QX1-2 (KUN-HKAS 129642, living-culture: KUNCC21-10009), QX2-1 (KUN-HKAS 129641, living-culture: KUNCC21-10010), QX2-2 (KUN-HKAS 129640, living-culture: KUNCC21-10011).
Notes: In the investigation of fungal diseases of cultivated morels in Guizhou Province, China, it was found that Hypomyces rosellus caused cobweb disease in cultivated Morchella sextelata. Typical symptoms were first, white to yellowish or pink cobweb-like mycelia appearing on the surface of the soil and the base of the stipe, spreading gradually to the morels’ pileus; later, the morels’ fruiting bodies became soft and rotten. To date, four species of Hypomyces/Cladobotryum were reported to cause cobweb disease on cultivated morels, i.e., C. protrusum (Lan et al. 2020), H. aurantius (this study), H. odoratus (Liu et al. 2023, this study), and H. rosellus (this study).
Myrotheciomycetaceae Crous
Trichothecium Link
Trichothecium roseum (Pers.) Link, Mag. Gesell. naturf. Freunde, Berlin 3(1-2): 18 (1809) ...……………………Fig. 13

Trichothecium roseum (KUN-HKAS 129653). a White colony on the fruiting body of morels. b–c Mycelia with abundant conidia on the surface of morel’s cap. d–j Conidiophores and conidia. k Conidia. l–m Germinating conidia. n Colony of T. roseum (KUNCC21-10015) on PDA at room temperature after 10 days. Scale bars: d–h = 100 μm; i–m = 50 μm; n = 20 mm
Index Fungorum number: IF 152448; Facesoffungi number: FoF 07768.
Fungicolous, colonizing on the fruiting body of cultured Morchella sextalata. Asexual morph: Mycelium partly superficial, composed of septate, loosely branched, floccose masses of white, hyaline hyphae bearing conidiophores and white spore masses. Conidiophores micronematous, sinuous or straight, erect, septate, sparingly branched and branches often more or less at right angles, originating from hyphae in the aerial mycelium. Conidiogenous cells monoblastic, terminal, hyaline, septate in the lower half, 2.5–4.5 μm wide. Conidia hyaline, pink in mass, initially aseptate, oval with distinct apiculus at the base, 1-septate cell when mature, ellipsoidal to pyriform, blunt basal apiculus and broadly rounded apex, with the curved basal cell, constricted at or near the septum, 10.5–25.5 × 7–15.5 μm (n = 30). Sexual morph: not observed.
Material examined: China, Yunnan Province, Kunming City, growing on the cultivated Morchella sextelata, Feng-Ming Yu, 08 April 2022, G8 (KUN-HKAS 129653, living-culture: KUNCC21-10015).
Notes: Trichothecium is generally considered as a contaminant. Members cause pink rot on various fruits and vegetables, and are a mycoparasite of other fungi (Summerbell et al. 2011). As mycoparasites, Trichothecium crotocinigenum, Trichothecium ovalisporum and Trichothecium roseum have been reported to cause fungal diseases in cultivated mushrooms, and cause economic losses to different degrees (Zaree et al. 2005; Sun et al. 2019a; Liu et al. 2021a). In this study, T. roseum is a new record of fungal agent that causing white mold disease on morel’s fruiting bodies in China. It appeared on the fresh fruiting bodies of Morchella sextelata, and the symptom was white hyphae with powdery conidia wrapping and covering the ridges and pits of cultivated morels.
Nectriaceae Tul. & C. Tul.
Cylindrodendrum Bonord.
Cylindrodendrum alicantinum Agustí-Bris., Armengol & A. Cabral, in Agustí-Brisach, Cabral, González-Domínguez, Pérez-Sierra, León, Abad-Campos, García-Jiménez, Oliveira & Armengol, Eur. J. Pl. Path. 145: 107 (2016) …………………………………………………………………………Fig. 14

Cylindrodendrum alicantinum (KUN-HKAS 129654). a, b White colony on the fruiting body of Morchella sextelata. c, d Mycelia with abundant conidia on the surface of morel’s cap. e–k Conidiophores and conidiogenous cells. l Conidia. m Colony of C. alicantinum (KUNCC21-10018) on PDA at room temperature after 10 days. Scale bars: e = 100 μm; f–l = 50 μm; m = 20 mm
Index Fungorum number: IF 811663; Facesoffungi number: FoF 15287.
Fungicolous, white, loose fluffy mycelium covering the ridges and pits of morels. Asexual morph: Conidiophores macronematous, micronematous, straight or slightly flexuous, branched or unbranched, septate, bearing up to five phialides, 21–56 μm long, 2.5–5 μm wide. Conidiogenous cells phialidic-shape, dimorphic; microphialides short, broad, flexuous, tapering markedly towards the apex, with a visible collarette, 9.5–17.5 μm long, 3 − 4 μm at widest point, 1 − 2 μm wide at the tip; macrophialides cylindrical and slightly subulate, 20.5–52.5 μm long, 2.5 − 4 μm at widest point, 1 − 2 μm wide at the tip. Conidia hyaline (0–)1–3 septate, cylindrical, long cylindrical, more or less straight, 10.5 − 24 × 2 − 3.5 μm (n = 30). Sexual morph: not observed.
Material examined: China, Yunnan Province, Kunming City, Panlong District, on fruiting bodies of cultivated Morchella sextelata, 15 February 2022, Feng-Ming Yu, 22mp-43 (KUN-HKAS 129656, living-culture: KUNCC21-10016), 22mp-44 (KUN-HKAS 129655, living-culture: KUNCC21-10017), and 22mp-45 (KUN-HKAS 129654, living-culture: KUNCC21-10018).
Notes: Cylindrodendrum species cause root rot diseases on a range of diverse hosts worldwide (Agustí-Brisach et al. 2016). However, there is no report on Cylindrodendrum species as fungal pathogens. In this study, we provide the first report of C. alicantinum growing on cultivated Morchella sextelata. Typical symptoms of this disease are white fluffy mycelia on the ridges and pits of morels, which colonize rapidly, and hyphae quickly cover the whole ascocarps. Cylindrodendrum alicantinum may lead morel to rot and decay, but its pathogenicity requires further testing.
Pseudodiploosporeaceae Jing Z. Sun, X.Z. Liu & H.W. Liu.
Pseudodiploospora Jing Z. Sun, X.Z. Liu & H.W. Liu.
Pseudodiploospora longispora (Matsush.) Jing Z. Sun, X.Z. Liu & H.W. Liu, in Sun, Yu, Lu, Liu & Liu, Mycology 14(1): 68 (2022) …………………………………………………………………..Fig. 15

Pseudodiploospora longispora (KUN-HKAS 129621). a, b White colony on the fruiting bodies of Morchella sextelata in morels’ farm. c Mycelia on the surface of morel’s cap. d Hyphae. e, f Racquet hyphae indicated by red arrows. g–l Conidiogenous structure and conidia. m, n Colony of P. longispora (KUNCC21-10023) on PDA at room temperature after 15 days. Scale bars: d = 200 μm; e–l = 50 μm; m, n = 50 mm
Index Fungorum number: IF 571283; Facesoffungi number: FoF 15288.
Fungicolous, growing on the fruiting bodies of cultivated morels and Peziza varia. Colonies on natural substrate loose floccose, whiteness, often confluent and forming irregular patches, sometimes effuse and hairy, with abundant powdery spores. Mycelium immersed and superficial, composed of septate, branched, floccose masses of white, hyaline hyphae. Hyphae 1.5–5.5 μm diam., abundant racquet hyphae, “racquet” 4–15.5 μm diam. Asexual morph: Hyphomycetous. Conidiophores macronematous and micronematous, mononematous, erect, simple, smooth, hyaline. Macronematous conidiophores straight or slightly flexuous, cylindrical, unbranched or sparingly branched, sometimes slightly attenuated towards the apex, pluriseptate, occasionally constricted at the septa. Micronematous conidiophores peg-like outgrowths with a single apical scar or somewhat longer, loci denticulate. Conidiogenous cells integrated, often seceding as ramoconidia, terminal and intercalary, straight or slightly geniculate, cylindrical, 28.5–101.5 × 2.5–7 μm, proliferation percurrent or sympodial, with one or several conidiogenous loci, protuberant, denticulate. Ramoconidia commonly formed, cylindrical to cylindrical-oblong, sometimes fusoid-ellipsoid, with a truncate base, sometimes attenuated towards the base, hyaline, 18–100 × 2–6.5 μm, 0–4-septata, with several conidiogenous loci. Conidia catenate, intercalary, lateral or terminal, hyaline, sometimes constricted in the middle or at the septa, smooth-walled; intercalary conidia cylindrical or ellipsoidal with apical hilum at both ends; lateral or terminal conidia, oval to limoniform, rounded towards the apex and with a single apical hilum or a truncated base, maturation acropetal and sometimes apparently basipetal, with immature septate conidia frequently occurring in the middle of the chain; variable in size, macroconidia 0–1-septata, rarely in 2–3-septata, 16.5–36 × 3–7.5 μm (n = 30); microconidia aseptate, variable in shape, lemon-like, droplet-like, ovoid, with a single apical hilum, sometimes constricted in the middle, 6–17 × 3–7.5 μm (n = 30). Sexual morph: not observed.
Material examined: China, Yunnan Province, Chuxiong City, on the fruiting bodies of Morchella sp. (Morchellaceae, Pezizales), 03 January 2022, Feng-Ming Yu, 22MP-1 (KUN-HKAS 129635, living-culture: KUNCC21-10037), 22MP-3 (KUN-HKAS 129637, living-culture: KUNCC21-10039), 22MP-14 (KUN-HKAS 129636, living-culture KUNCC21-10038). Yunnan Province, Kunming City, Wuhua District, on the fruiting bodies of Morchella sextelata (Morchellaceae, Pezizales), 17 February 2022, Feng-Ming Yu, 22MP-40 (KUN-HKAS 129621, living-culture: KUNCC21-10023), 22MP-41 (KUN-HKAS 129633, living-culture: KUNCC21-10035); 22 February 2022, Feng-Ming Yu, 22MP-48 (KUN-HKAS 129630, living-culture: KUNCC21-10032). Yunnan Province, Kunming City, Panlong District, on the fruiting bodies of Morchella sextelata (Morchellaceae, Pezizales), 22 February 2022, Feng-Ming Yu, 9S-1B (KUN-HKAS 129628, living-culture: KUNCC21-10031); 15 March 2022, Feng-Ming Yu, KIB315-10A (KUN-HKAS 129619, living-culture: KUNCC21-10021); 14 March 2022, Feng-Ming Yu, KIB314-3 (KUN-HKAS 129631, living-culture: KUNCC21-10033). Yunnan Province, Yuxi City, on the fruiting bodies of Morchella sp. (Morchellaceae, Pezizales), 06 January 2022, Feng-Ming Yu, 22MP-18 (KUN-HKAS 129623, living-culture: KUNCC21-10025), 22MP-20 (KUN-HKAS 129632, living-culture: KUNCC21-10034), 22MP-20A (KUN-HKAS 129629, living-culture: KUNCC21-10030), 22MP-22 (KUN-HKAS 129620, living-culture: KUNCC21-10020). Yunnan Province, Chuxiong City, on the fruiting bodies of Peziza varia (Pezizaceae, Pezizales), 26 February 2022, Feng-Ming Yu, 22MP-50 (KUN-HKAS 129622, living-culture: KUNCC21-10024); 03 January 2022, Feng-Ming Yu, 22MP-8 (KUN-HKAS 129627, living-culture: KUNCC21-10029), 22MP-9 (KUN-HKAS 129626, living-culture: KUNCC21-10028). Guizhou Province, Guiyang City, on the fruiting bodies of Morchella sp. (Morchellaceae, Pezizales), 16 January 2022, Feng-Ming Yu, 22MP-23 (KUN-HKAS 129634, living-culture: KUNCC21-10036); Qianxi City, on the fruiting bodies of Morchella sp. (Morchellaceae, Pezizales), 11 March 2022, Feng-Ming Yu, QX1-3 (KUN-HKAS 129618, living-culture: KUNCC21-10022). Hebei Province, on the fruiting bodies of Morchella sp. (Morchellaceae, Pezizales), 10 March 2022, Qi Zhao, HB-1(KUN-HKAS 129625, living-culture: KUNCC21-10027), HB-3 (KUN-HKAS 129624, living-culture: KUNCC21-10026).
Notes: Pseudodiploospora longispora can colonize the dead leaf of Colocasia esculenta var. antiquorum (Japan), human skin and feet (Canada), and the ascocarp of Morchella spp. (China) (Tanney et al. 2015; Sun et al. 2022). In this study, we isolated 19 strains of P. longispora from infected cultivated morels, and one strain from Peziza varia in morels’ farm. Although they have some base differences in gene fragments, there is no significant difference in morphology. In China, this species forms “white mold” symptom on morels, which leads to deformation and rot of fruiting bodies. Based on our field investigation, P. longispora accounts for the highest proportion of all pathogenic fungi (up to 80%) that cause “white mold” diseases, and it is speculated that P. longispora is a major epidemic factor (Shi et al. 2022).
Amendment of Pseudodiploosporeaceae and problems with Paecilomyces penicillatus (CBS 448.69)
Diploospora was established by Grove (1916) with D. rosea as the type species. The phylogenetic analyses of ITS and SSU + LSU revealed that D. rosea is an onygenalean taxon in Onygenales (Tanney et al. 2015). However, based on ITS BLAST query, specimens identified as Diploospora longispora (KT279806 – KT279808) and Diploospora longispora var. cubensis (KT279809) were most closely related to Paecilomyces penicillatus (CBS 448.69, AY624194) belonging to the order Hypocreales, and they reached affinity with Hypocreaceae (Luangsa-ard et al. 2004, 2005; Tanney et al. 2015). Paecilomyces was introduced by Bainier (1907a) with Paecilomyces variotii (CBS 102.74) as its type species (Samson 1974). Phylogeny based on SSU demonstrated that Paecilomyces was polyphyletic across two classes (Luangsa-Ard et al. 2004). Type species Pa. variotii and its thermophilic relatives were placed in Eurotiales (Eurotiomycetes), while the mesophilic species (e.g., Pa. penicillatus) were placed in Hypocreales (Sordariomycetes) (Luangsa-Ard et al. 2004, 2005). In Samson (1974), Paecilomyces penicillatus was recorded on dead moss, rotting wood, Arcyria punicea and rotting mushrooms, and it was placed in Eurotiales based on morphological characteristics. However, specimen Pa. penicillatus CBS 448.69 (AY526493) was not congeneric with Pa. variotii CBS 102.74 (AY526477) in the SSU phylogeny (Luangsa-ard et al. 2004). The former species was later transferred to Hypocreales, and showed a close relationship with Hypocreaceae (Luangsa-ard et al. 2004, 2005). Based on a combined SSU, ITS, LSU, TEF, and RPB 2 dataset, Sun et al. (2022) segregated a novel genus Pseudodiploospora from Diploospora to accommodate four combinations (i.e., Pseudodiploospora cubensis, Ps. fungicola, Ps. longispora, and Ps. zinniae), and erected the genus Zelopaecilomyces for the species Paecilomyces penicillatus, as well as introducing a new family Pseudodiploosporeaceae in Hypocreales to accommodate these two genera. In the phylogeny provided by Sun et al. (2022), Pseudodiploosporeaceae and Hypocreaceae formed a sister lineage with robust support, and Zelopaecilomyces diverged from Pseudodiploospora at ca. 14 MYA. However, the reference molecular data of Zelopaecilomyces for phylogenetic analyses were not specified in Sun et al. (2022).
In the phylogeny based on the LSU + ITS + TEF + RPB 2 sequence data, specimen Zelopaecilomyces penicillatus (CBS 448.69) unexpectedly nested within Pseudodiploospora longispora (Fig. 3), which was different from Sun et al. (2022). Molecular data for Paecilomyces penicillatus (CBS 448.69) in GenBank are AY526493 (SSU, Luangsa-ard et al. 2004), AY624194 (ITS, Luangsa-ard et al. 2005), AY624232 (TUB2, Luangsa-ard et al. 2005), JX012226 (ITS, submitted in 2012 without reference), MF416674 (RPB1, Kepler et al. 2017), and MH859348 (ITS, Vu et al. 2019). MH859348 and AY624194 are identical, and have 31.2% base pair (166/532 bp) differences from JX012226. Based on an ITS BLAST query, sequence JX012226 has an affinity with Penicillium (Eurotiales, Eurotiomycetes), while AY624194 and MH859348 have similarity with hypocrealean taxa in Sordariomycetes. We therefore re-analysed the data of Sun et al. (2022) with different sequence matrices (Table 5, Table S1, and S2). The combined SSU/ITS gene fragments used in the analyses of Sun et al. (2022), comprised two different sets of ITS data for Zelopaecilomyces, i.e., matrix I (SSU-AY526493 + ITS-AY624194) and matrix II (SSU-AY526493 + ITS-JX012226) (Table 5). In matrix III, two authentic Penicillium taxa (KUNCC21-10102 and KUNCC21-10101) isolated from infected morels and sequenced in this study, and the data of Penicillium chrysogenum CBS 306.48 (Schoch et al. 2014) downloaded from GenBank were used (Table 5). In matrix IV, three artificially designed chimeras, which were the set of SSU (AY526493) and ITS genes of three Penicillium (OR432197, OR432196 and NR_077145) respectively, were used to test the effect on the phylogenetic analyses as obtained in matrix II (Table 5).
The phylogeny revealed that the matrix I (SSU-AY526493 + ITS-AY624194) nested with Pseudodiploospora longispora, while the matrix II (SSU-AY526493 + ITS-JX012226) and three artificially designed chimeras of matrix IV clustered together and formed an independent lineage with high support (SH-aLRT = 100, UFB = 100, Fig. 16). The clade containing matrix II and IV was the sister of Pseudodiploospora in Pseudodiploosporeaceae, which was similar to the phylogenetic position of Zelopaecilomyces in Sun et al. (2022). A wider taxon sampling was used for phylogeny in Fig. 17, including sequences of Eurotiomycetes. The matrix III containing the sequences produced by strains KUNCC21-10102, KUNCC21-10101 and CBS 306.48 belonged to Penicillium in Eurotiomycetes, and the matrix I still belonged to Ps. longispora. The inconsistency of phylogenetic positions inferred by different ITS genes indicate that the sequences under the name of Zelopaecilomyces penicillatus (CBS 448.69) are problematic. The sequence AY526493 (SSU) and AY624194 (ITS) were derived from a sample of Pseudodiploospora longispora, while JX012226 (ITS) was generated from a taxon of Penicillium. Zelopaecilomyces was established by chimeric sequences in Sun et al. (2022), which contained a ITS gene region of Eurotiomycetes and an SSU gene region of Sordariomycetes. Therefore, this study suggests that Zelopaecilomyces should be rejected, and the family Pseudodiploosporeaceae has been amended here.

Maximum-Likelihood (IQ-TREE-ML) consensus tree inferred from the combined SSU, ITS, LSU, TEF and RPB 2 multiple sequence alignment of members of Sordariomycetes. Bootstrap support values for ML ≥ 80 of SH-aLRT or 95 of UFB are indicated above the nodes and separated by ‘–/–’ (SH-aLRT/UFB)


Maximum-Likelihood (IQ-TREE-ML) consensus tree inferred from the combined SSU, ITS, LSU, TEF, and RPB 2 multiple sequence alignment of members of Eurotiomycetes and Sordariomycetes. Bootstrap support values for ML ≥ 80 of SH-aLRT or 95 of UFB are indicated above the nodes and separated by ‘–/–’ (SH-aLRT/UFB)
Amended family Pseudodiploosporeaceae
Pseudodiploosporeaceae Jing Z. Sun, X.Z. Liu & H.W. Liu, in Sun, Yu, Lu, Liu & Liu, Mycology 14(1): 67 (2022), emended.
Index Fungorum number: IF 571193; Facesoffungi: FoF 15289.
Type genus: Pseudodiploospora Jing Z. Sun, X.Z. Liu & H.W. Liu.
Fungicolous or saprobic. Asexual morph: Colonies on natural substrate effuse, whitish. Mycelia superficial or immersed. Hyphae racquet hyphae present, branched, septate, hyaline. Conidiophores micronematous, mononematous, erect, simple, straight or slightly flexuous, unbranched or sparingly branched, smooth, aseptate to septate, hyaline. Conidiogenous cells holoblastic, polyblastic, sympodial, loci conspicuous, terminal and intercalary in conidiophores, hyaline, with denticles. Ramoconidia and secondary ramoconidia often formed, cylindrical or fusiform, aseptate or septate, truncate at the base, with terminal scars. Conidia subcylindrical-ellipsoidal, slightly pointed at both ends, sometimes constricted at the septa, smooth-walled, with a flat scar at each slightly denticulate end, aseptate or septate, in acroptetal chains, hyaline, dry, blastic, straight to slightly curved. Sexual morph: not observed.
Notes: Sun et al. (2022) established the family Pseudodiploosporeaceae in Hypocreales to accommodate two novel genera Pseudodiploospora and Zelopaecilomyces. Its original description is micronematous to macronematous, mononematous, and penicillate conidiophores (Sun et al. 2022). However, our phylogenetic analyses (Fig. 3, 16 and 17) indicate that Zelopaecilomyces is questionable as it was established by a chimerism of gene fragments from Pseudodiploospora longispora and Penicillium sp. in Sun et al. (2022). Hence, this study proposes that Zelopaecilomyces should be rejected, and the use of this name be discontinued. Thus, there is only a single genus Pseudodiploospora in Pseudodiploosporeaceae. Its characteristics are amended here.
Pseudodiploospora Jing Z. Sun, X.Z. Liu & H.W. Liu, in Sun, Yu, Lu, Liu & Liu, Mycology 14(1): 67 (2022).
Index Fungorum number: IF 571281; Facesoffungi: FoF 15290.
Colonies fluffy, whitish. Mycelia immersed or superficial, composed of branched, septate, hyaline hyphae with abundant racquet hyphae. Asexual morph: Conidiophores micronematous, mononematous, erect, simple, straight or slightly flexuous, unbranched or sparingly branched, smooth, aseptate, hyaline. Conidiogenous cells holoblastic, polyblastic, sympodial, acropetal, terminal and intercalary in conidiophores, hyaline, with denticles. Ramoconidia formed, cylindrical-oblong, 0–1-septate, rarely multi-septate, with usually more than one (mostly 2) conidial hilum, on which typically accumulate conidia and/or secondary ramoconidia at their tip, sometimes indistinguishable from conidiogenous cells. Conidia fusoid, ellipsoid, narrowly ellipsoid, subcylindrical, formed in branched chains, straight to slightly curved, septate, rarely aseptate, sometimes constricted at the septa, slightly pointed at both ends, smooth-walled, with a flat scar at each slightly denticulate end, 0–3-septate, in acroptetal chains, hyaline dry, blastic; small terminal conidia subglobose, ovoid to obovoid or somewhat limoniform, mostly aseptate. Secondary ramoconidia often formed, shorter but somewhat wider than ramoconidia, ellipsoid to subcylindrical or cylindrical to long-cylindrical. Sexual morph: not observed.
Type species: Pseudodiploospora longispora (Matsush.) Jing Z. Sun, X.Z. Liu & H.W. Liu.
Host and Habits: Found in fruiting bodies of other fungi, e.g., Auricularia spp., Morchella spp., Peziza spp.; on dead or fallen plant leaves and seeds, e.g., Colocasia esculenta var. antiquorum, Leguminosae spp., Zinnia elegans; on dung.
Distribution: Canada, China, Cuba, and Japan.
Notes: Pseudodiploospora consists of P. cubensis, P. fungicola, P. longispora, and P. zinniae. The main features are summarized in Table 6. They exist on humus, leaves and seeds of plants, and ascomata and basidiomata of other fungi (Sun et al. 2022).
Problems with Paecilomyces penicillatus (CBS 448.69)
All illustrations and descriptions of the conidiophores of Paecilomyces penicillatus are phialidic acremonium-like (Samson 1974; Sun et al. 2022), which is difficult to associate with that of in Pseudodiploospora. In Samson (1974), the description of Pa. penicillatus (CBS 448.69) is phialidic acremonium-like, which corresponds to ITS BLAST query of JX012226. However, the diploospora-like samples with sequences AY526493 and AY624194 have not been reported in published literatures. In our observation, diploospora-like and paecilomyces-like pathogens frequent occur on the fruiting bodies of cultivated morel (Fig. 18). This symptom is usually thought to be caused by a fungal pathogen, but in fact, it includes two pathogenic taxa. This study speculates that the original voucher labelled Paecilomyces penicillatus (CBS 448.69) contained two different species growing together, which generated two types of molecular information, one belonging to Pseudodiploospora in Hypocreales (Sordariomycetes) and the other belonging to Penicillium in Eurotiales (Eurotiomycetes). The description and illustration of Pa. penicillatus (CBS 448.69) in Samson (1974) and Sun et al. (2022) could be derived from the latter.

Pseudodiploospora longispora (indicated by a blue arrow) and Penicillium sp. (indicated by a red arrow) appeared on infected morels simultaneously. a, b KUNCC21-10101
Species diversity of fungal pathogens on common cultivated mushrooms
Research on fungal diseases of cultivated mushrooms is a hot topic (Fletcher and Gaze 2008; Sun et al. 2022). Cultivated mushrooms are excellent sources of proteins, minerals, vitamins, polysaccharides, and unsaturated fatty acids (Hyde et al. 2019; Mapook et al. 2022). For thousands of years, some taxa have been widely used as food, nutraceutical, and pharmaceutical products worldwide, and they are rich sources of bioactive compounds beneficial to human health (Anonymous 1955; Hyde et al. 2019; Wu et al. 2019; Ho et al. 2020; Mapook et al. 2022). It is estimated that there are 7,000 known mushroom species with different degrees of nutrient value, of which about 3,000 are considered as popular types of edible fungi, and more than 60 species could be cultivated commercially (Miles and Chang 2004; Fazenda et al. 2008; Elisashvili 2012; Hawksworth 2012; Corrêa et al. 2016; Chang and Wasser 2017; Rathore et al. 2017; Wu et al. 2019). The key importance of mushroom cultivation lies not only in its nutritional and medicinal properties, but also in its economic value (Niego et al. 2023). Mushroom cultivation has always been a central issue (Hyde et al. 2019; Mapook et al. 2022). However, with the rapid expansion of cultivation, diseases have become a bottleneck restricting mushroom production, especially those caused by fungi.
By 2023, there were 133 fungal pathogens reported on cultivated mushrooms, which lead to economic losses to different degrees. Fungal pathogens have high species diversity in 58 genera, 40 families, 20 orders, 12 classes, and six phyla (Fig. 19, Table 7). These pathogens are more diverse within Sordariomycetes, mostly exclusive pathogens reported in Hypocreaceae (Hypocreales), followed by some Eurotiomycetes in Aspergillaceae. At least 23 species of cultivated mushrooms are affected by fungal pathogens to varying degrees. The most affected cultivated mushrooms are Agaricus bisporus, Lentinula edodes, Morchella spp., and Pleurotus spp. (Table 7).

Species diversity of fungal pathogens in phylum (a), class (b), order (c) and family (d) on cultivated mushrooms
Fungal pathogens of cultivated mushrooms*
Note: * The classification follows Wijayawardene et al. (2022).
Ascomycota Caval.-Sm.
Dothideomycetes sensu O.E. Erikss & Winka.
Cladosporiales Abdollahz. & Crous.
Cladosporiaceae Chalm. & R.G. Archibald.
Cladosporium Link.
Notes: Cladosporium is a monophyletic genus in Cladosporiaceae whose species are cosmopolitan in distribution and commonly encountered in indoor and outdoor environments, including extreme ecological niches (Bensch et al. 2012). Some Cladosporium species are saprobic, and some have been reported as endophytes, hyperparasites on other fungi and plants, as well as animal pathogens, including humans (Bensch et al. 2012; Iturrieta-González et al. 2021; Salvatore et al. 2021). As the largest and most heterogeneous genus of hyphomycetes, Cladosporium currently encompasses more than 500 names in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 2 June 2023). Sun et al. (2019a) listed 13 Cladosporium species that can parasitize or colonize other fungi, e.g., Balladyna magnifica, Lophodermium pinastri, and Teratosphaeria proteae-arboreae. Only Cladosporium allicinum was reported to infect the mycelium of Pleurotus eryngii, which led to a decline in production yield and caused deterioration of commercial value (Kim et al. 2013).
Cladosporium species causing disease of cultivated mushrooms
Cladosporium allicinum (Fr.) Bensch, U. Braun & Crous (Kim et al. 2013).
Eurotiomycetes Tehler ex O.E. Eriksson & K. Winka.
Eurotiales G.W. Martin ex Benny & Kimbr.
Aspergillaceae Link.
Aspergillus P. Micheli ex Haller.
Notes: Aspergillus was introduced in 1768, and typified by A. glaucus in 1931 (Haller 1768). Aspergillus comprises diverse species that occur worldwide in various habitats and are common food spoilage taxa (Krijgsheld et al. 2013; Samson et al. 2014). Some Aspergillus species are opportunistic pathogens of animals and even humans, while others are used in biotechnology to produce various metabolites or as agents in food fermentations (Krijgsheld et al. 2013; Samson et al. 2014; Niego et al. 2023). Currently, Aspergillus encompasses about 580 taxa in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 2 June 2023). Some are fungicolous and parasitize wild fruiting bodies of Earliella scabrosa and Sclerotinia sclerotiorum, e.g., Aspergillus aculeatus, A. polyporicola, and A. terreus (Melo et al. 2006; Hu et al. 2013; Hubka et al. 2015). Aspergillus taxa can be causative agents of “aspergillus mold” in cultivated mushrooms, they often infect and inhabit the fruiting bodies, compost, substrate, and even air in a greenhouse of several cultivated mushrooms, e.g., Agaricus bisporus, Cordyceps militaris, Cyclocybe aegerita, Lentinula edodes, Macrocybe gigantea, and Pleurotus ostreatus (Table 7).
Aspergillus species causing disease of cultivated mushrooms
Aspergillus flavus Link (Fletcher and Gaze 2008; Wang et al. 2017; Liu et al. 2021a).
Aspergillus fumigatus Fresen. (Wickremasinghe et al. 1998, Fletcher and Gaze 2008).
Aspergillus niger Tiegh. (Fletcher and Gaze 2008; Wang et al. 2017; Liu et al. 2021a).
Aspergillus pulverulentus (McAlpine) Thom (Kwon et al. 2015).
Aspergillus tubingensis Mosseray (Choi et al. 2010).
Penicillium Link.
Notes: Penicillium was established by Link (1809a, b) with the type species P. expansum. Penicillium species occur in various habitats, such as soil, vegetation, air, indoor environments, food products, and other fungi (Visagie et al. 2014). Some are considered to play vital roles in natural ecosystems, agriculture, and medicine, while others are reported as the postharvest pathogens on a wide range of fruits, vegetables, and mushrooms (Visagie et al. 2014; Park et al. 2019). According to Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 02 June 2023), the widespread Penicillium genus currently encompasses about 540 taxa. Among them, several members are fungicolous, e.g., P. angulare colonizes old Polyporaceae, P. jiangxiense parasitizes Cordyceps jiangxiensis, and P. pancosmium on Armillaria mellea (Kang and Liang 2003; Peterson et al. 2004; Houbraken et al. 2011). Penicillium-mold and smoky mold caused by Penicillium species are common diseases of fruiting bodies, composts, and substrates of cultivated Agaricus bisporus, Cordyceps militaris, Cyclocybe aegerita, Lentinula edodes, Macrocybe gigantea, Phallus rubrovolvatus, and Pleurotus eryngii (Table 7). These pathogens effect and inhibit the growth of the mycelium and fruiting bodies, resulting in economic losses.
Penicillium species causing disease of cultivated mushrooms
Penicillium chermesinum Biourge (Fletcher and Gaze 2008).
Penicillium citreonigrum Dierckx (Fletcher and Gaze 2008).
Penicillium corylophilum Dierckx (Jo et al. 1999).
Penicillium dierckxii Biourge (Fletcher and Gaze 2008).
Penicillium expansum Link (Liu et al. 2021a).
Penicillium hermansii Houbraken, Seifert & Samson (Houbraken et al. 2019).
Penicillium implicatum Biourge (Fletcher and Gaze 2008).
Penicillium oxalicum Currie & Thom (Fletcher and Gaze 2008).
Penicillium solitum Westling (Choi et al. 2010).
Penicillium spp. (Hassan 2013; Sharma et al. 2016; Wang et al. 2017).
Thermoascaceae Apinis.
Paecilomyces Bainier.
Notes: Paecilomyces was described by Bainier (1907a) based on the only species, P. variotii. Paecilomyces species are pathogenic and saprobic, and distributed in a wide range of habitats, including soil, decomposing plant material or food, compost, insects, nematodes, and the rhizosphere of various plants (Senthilkumar et al. 2020). In addition, some endophytic Paecilomyces can be used as potential bio-stimulants, which play an important role in the development of numerous plants and crops (Moreno-Gavíra et al. 2020). Currently, Paecilomyces has more than 70 taxa assigned in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 2 June 2023). Sun et al. (2019a) listed nine members of Paecilomyces as fungicolous, which can parasitize or grow on Armillaria mellea, Cordyceps spp., and some Polyporaceae (Liu et al. 1996; Kang and Liang 2003; Peterson et al. 2004; Houbraken et al. 2011). Paecilomyces variotii is reported as a pathogen that causes the fruiting body of Phallus rubrovolvatus to rot (Sang et al. 2016).
Paecilomyces species causing disease of cultivated mushrooms
Paecilomyces variotii Bainier [as 'varioti'] (Sang et al. 2006).
Onygenales Cif. ex Benny & Kimbr.
Onygenaceae Berk.
Chrysosporium Corda.
Notes: Chrysosporium was established by Corda (1833) with C. corii as a type species. Members of this genus are mostly saprobic and keratinolytic (Bowman et al. 1996), and are known to be occur in various habitats, e.g., air, sea water, sludge, and waste water (Zhang et al. 2016). Some of these species are responsible for occasional opportunistic infections in humans (Kamath et al. 2015). Currently, more than 50 taxa are listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 6 June 2023). Notably, Chrysosporium merdarium and C. synchronum are agents of confetti mold on cultivated Agaricus bisporus (Table 7).
Chrysosporium species causing disease of cultivated mushrooms
Chrysosporium merdarium (Ehrenb.) J.W. Carmich. (Fletcher and Gaze 2008).
Chrysosporium synchronum Oorschot (Van Oorschot 1980).
Sporendonema Desm.
Notes: As a rare and old genus, Sporendonema was described by Desmazières (1827) to accommodate an orange cheese fungus S. casei as the type species. Currently, Sporendonema comprises only two species (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 6 June 2023), i.e., S. casei and S. purpurascens, which can sometimes contaminate mushroom growing facilities (Wood 1957). Sporendonema purpurascens is regarded as the agent of lipstick mold of cultivated Agaricus bisporus, and it can appear and live on compost and substrate in the greenhouse (Table 7).
Sporendonema species causing disease of cultivated mushrooms
Sporendonema purpurascens (Bonord.) E.W. Mason & S. Hughes (Van Greuning and Eicker 1991; Largeteau and Savoie 2010).
Uncinocarpus Sigler & G.F. Orr.
Notes: Sigler and Carmichael (1976) described Uncinocarpus to accommodate the type species U. reesii, a soil species having reddish brown, oblate, smooth ascospores. To date, two species are recorded in Species Fungorum, distributed in North America, Europe, Asia, and the Middle East (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 06 June 2023). Uncinocarpus species are of saprotrophic micro-fungi that grows in/on dung, soil and on keratinous materials (e.g., bone, hair, feathers and skin) (Sigler et al. 2011). Uncinocarpus uncinatus is the causative agent of confetti disease in cultivated Agaricus bisporus, and it can appear on the substrate in greenhouse (Table 7).
Uncinocarpus species causing disease of cultivated mushrooms
Uncinocarpus uncinatus (Eidam) Currah (Eicker and Greuning 1991).
Leotiomycetes O.E. Erikss. & Winka.
Helotiales Nannf. ex Korf & Lizoň.
Helotiaceae Rehm.
Scytalidium Pesante.
Notes: Scytalidium was established by Pesante (1957) with the type species of S. lignicola. It has a widespread distribution, often infecting and growing on various plants, crops, other fungi and even human feet and toes (Geng et al. 2016). Scytalidium has 26 species listed in the Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 2 June 2023). Among them, four species infect cultivated mushrooms. Scytalidium auriculariicola, the causative agent of slippery scar, is one of the most destructive pathogens encountered in cultivating Auricularia polytricha. The typical symptom of a slippery scar is that once the mushroom bag is infected, the brown lesions quickly spread to the whole mycelium, which delays basidiocarp development and causes severe economic losses (Peng et al. 2014). Three Scytalidium species, including S. cuboideum, S. ganodermophthorum, and S. sphaerosporum, were reported as pathogens that cause yellow rot disease in cultivated Ganoderma lucidum (Table 7).
Scytalidium species causing disease of cultivated mushrooms
Scytalidium auriculariicola W.H. Peng, Z.R. Yang bis & Qun Sun (Sun and Bian 2012; Peng et al. 2014).
Scytalidium cuboideum (Sacc. & Ellis) Sigler & Kang (Oh et al. 1998).
Scytalidium ganodermophthorum Kang, Sigler, Y.W. Lee & S.H. Yun (Kang et al. 2010; Liu et al. 2019).
Scytalidium sphaerosporum Sigler & Kang (Lee et al. 1996).
Myxotrichaceae Currah.
Oidiodendron Robak.
Notes: Oidiodendron was circumscribed by Robak (1932) for three species isolated from wood pulp, i.e., O. fuscum, O. nigrum, and O. rhodogenum (Robak 1932). Species of Oidiodendron have been isolated worldwide, especially in temperate regions. They are known as saprobes, and have a wide range of hosts, such as decaying plant materials, animal and fungal substrates/composts, peat, soil, lichens, marine sediments and holothurians, and even human skin and decomposing human hair (Rice and Currah 2005a, b). Currently, the genus comprises 26 species (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 2 June 2023). Oidiodendron fimicola and an undetermined Oidiodendron species have been reported on cultivated mushroom composts (Table 7).
Oidiodendron species causing disease of cultivated mushrooms
Oidiodendron fimicola A.V. Rice & Currah (Rice and Currah 2005b).
Oidiodendron sp. (an undetermined species) (Fletcher and Gaze 2008).
Sclerotiniaceae Whetzel ex Whetzel.
Botrytis P. Micheli ex Pers.
Notes: Botrytis species are efficient pathogens, causing devastating diseases and significant crop losses in a wide variety of plant species (Elad et al. 2004). Botrytis is reported to infect over 1,400 plant species, many of which are important agricultural crops (Elad et al. 2004; Valero-Jiménez et al. 2019). Currently, there are about 76 species recorded in Species Fungorum (https://www.speciesfungorum.org/Names/names.asp?pg=1, accessed date: 2 June 2023). Botrytis cinerea is the most destructive necrotrophic species, with a wide range of hosts (Williamson et al. 2007). Eicker and Greuning (1991) reported B. cinerea infecting the substrate of Agaricus bisporus, causing brown mold (Table 7).
Botrytis species causing disease of cultivated mushrooms
Botrytis cinerea Pers. (Eicker and Greuning 1991).
Phacidiales C.E. Bessey.
Helicogoniaceae Baral.
Eleutheromyces Fuckel.
Notes: Eleutheromyces has two species, i.e., E. pseudosubulatus and E. subulatus (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 12 June 2023). Both are fungicolous, the former species is isolated from Lactarius scrobiculatus, and later is reported on fruiting bodies of cultivated Lentinula edodes causing black spot disease (Tsuneda et al. 1997; Crous et al. 2015).
Eleutheromyces species causing disease of cultivated mushrooms
Eleutheromyces subulatus (Tode) Fuckel (Tsuneda et al. 1997).
Orbiliomycetes O.E. Erikss. & Baral.
Orbiliales Baral, O.E. Erikss., G. Marson & E. Weber.
Orbiliaceae Nannf.
Orbilia Fr.
Notes: Orbilia was established by Fries (1836), and later the typification O. xanthostigma was designated by Bachman in 1909. Members of Orbilia are widely distributed as saprobic, parasitic or superficial on decayed wood or bark of plants, dung, and other fungi (Qin et al. 2011; Qiao et al. 2012; Baral et al. 2018; Zhang et al. 2020a, b). Orbilia is important in establishing the sexual–asexual morphs connections, as it has produced several asexual genera, including Arthrobotrys, Dactylella, Dicranidion, Dwayaangam, Drechslerella, Helicoon, Monacrosporium, Pseudotripoconidium, and Trinacrium (Qin et al. 2011; Qiao et al. 2012; Zhang et al. 2020a, b). Orbilia species are globally distributed, comprising more than 500 species (https://www.speciesfungorum.org/Names/names.asp?pg=1, accessed date: 2 June 2023). Orbilia inflatula is reported as fungicolous that colonizes the fruiting bodies of Hypoxylon fragiforme (Helfer 1991), and O. oligospora can be a pathogen, which forms arthrobotrys brown mold on compost of cultivated Agaricus bisporus (Table 7).
Orbilia species causing disease of cultivated mushrooms
Orbilia oligospora (Fresen.) Baral & E. Weber (Eicker and Greuning 1991; van Greuning and Ficker 1992).
Pezizomycetes O.E. Erikss. & Winka.
Pezizales J. Schröt.
Ascodesmidaceae J. Schröt.
Cephaliophora Thaxt.
Notes: Cephaliophora was described by Thaxter (1903) to accommodate two coprophilous species, C. tropica (type species) and C. irregularis. Three species are placed in Cephaliophora (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 6 June 2023). They are endophytic, pathogenic, and saprobic, and have a range of host/habit, including soil, dung, plant, diverse substrates, moth pupae, rotifer, and even humans (Tanabe et al. 1999). It is reported that C. tropica can cause mold on the compost and substrate of cultivated Agaricus bisporus (Table 7).
Cephaliophora species causing disease of cultivated mushrooms
Cephaliophora tropica Thaxt. (Fletcher and Gaze 2008).
Pezizaceae Dumort.
Chromelosporium Corda.
Notes: Chromelosporium is common in soil and plant substrate. Currently, eight species are placed in Chromelosporium (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 6 June 2023). Previous research revealed that C. fulvum is the agent of peat/brown mold on the compost and substrate of cultivated Agaricus bisporus (Table 7).
Chromelosporium species causing disease of cultivated mushrooms
Chromelosporium fulvum (Link) McGinty, Hennebert & Korf (Coetzee and Eicker 1990; Eicker and Greuning 1991; Hassan 2013).
Pyronemataceae Corda.
Diehliomyces Gilkey.
Notes: Diehliomyces was described by Gilkey (1955) with the type species D. microsporus (Gilkey 1954). Diehliomyces microspores is originally discovered in soil, and later it’s frequently reported as a fungal disease agent in mushroom farms worldwide (Parmar and Sodavadiya 2019). The false truffle resulting from infection by D. microsporus is one of the main problems affecting the yield of cultivated Agaricus mushrooms (Table 7).
Diehliomyces species causing disease of cultivated mushrooms
Diehliomyces microsporus (Diehl & E.B. Lamb.) Gilkey (Fletcher and Gaze 2008; do Nascimento and da Eira 2013; Hassan 2013).
Pezizales genera incertae sedis.
Oedocephalum Preuss.
Notes: Preuss (1851) erected Oedocephalum with the type species O. elegans (Preuss 1851). Oedocephalum species are widely distributed and grow on a wide range of hosts, including soils, dungs, decaying wood and vegetables, paper production, mushrooms and cheese (Sharma et al. 2006). Currently, 21 taxa of Oedocephalum are listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 6 June 2023). Oedocephalum glomerulosum is reported as the agent of oedocephalum-mold on the compost and substrate of cultivated Agaricus bisporus (Table 7).
Oedocephalum species causing disease of cultivated mushrooms
Oedocephalum glomerulosum (Bull.) Sacc. (Botha et al. 1990; Fletcher and Gaze 2008).
Sordariomycetes O.E. Erikss. & Winka.
Coronophorales Nannf. Genera incertae sedis.
Papulaspora Preuss.
Notes: Papulaspora, as unusual taxa, was erected by Preuss (1851), and was typified by P. sepedonioides. Papulaspora species are presumed to be saprobes, mycoparasites, and opportunistic pathogens, which can be distributed in a wide variety of substrates, including plant debris, pulp products, wood, dung, soil, other fungi and animal tissues (Davey et al. 2008). Currently, there are 32 names listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 2 June 2023). Papulaspora byssina is the causative agent of brown plaster mold on Agaricus bisporus, Calocybe indica, and Pleurotus spp., which can cause complete crop failure in severe cases (Anonymous 2012; Binit et al. 2013; Murmu et al. 2014).
Papulaspora species causing disease of cultivated mushrooms
Papulaspora byssina Hotson (Dar et al. 1995; Largeteau and Savoie 2010; Binit et al. 2013; Hassan 2013; Murmu et al. 2014).
Diaporthales Nannf.
Diaporthaceae Höhn. ex Wehm.
Diaporthe Fuckel.
Asexual morph: Phomopsis Sacc. & Roum.
Notes: Fuckel (1867) established Diaporthe to accommodate D. alnea (the type species) in 1867. With the implementation of the “One Fungus = One Name” nomenclatural principle, the name Diaporthe takes precedence over its asexual genus Phomopsis (Rossman 2014; Norphanphoun et al. 2022). Diaporthe species have a broad geographic distribution and are reported as plant-pathogens, endophytes, saprobes, and even causing animal health problems (Udayanga et al. 2011; Norphanphoun et al. 2022). Some Diaporthe cause stem cankers, diebacks, rots, leaf spots, blights, seed decay, and wilts on a wide range of plant hosts, including sunflower, grapevine, and soybean (Udayanga et al. 2011). Currently, more than 620 taxa are listed in Diaporthe (https://www.speciesfungorum.org/Names/names.asp?pg=1, accessed date: 23 July 2023), and 13 species complexes are introduced by Norphanphoun et al. (2022). An unidentified species of Diaporthe is reported to infect the mycelium of cultivated Pleurotus eryngii, and caused severe damage to the commercial production (Table 7).
Diaporthe species causing disease of cultivated mushrooms
Diaporthe sp. (an unidentified species) (Kim et al. 2013).
Glomerellales Chadef. ex Réblová, W. Gams & Seifert.
Plectosphaerellaceae W. Gams, Summerb. & Zare.
Acrostalagmus Corda.
Notes: Acrostalagmus was established by Corda (1838), and its type species A. cinnabarinus was later designated by Clements and Shear in 1931. Acrostalagmus is characterized with verticillate conidiophores, and hyaline, egg-shaped conidia formed singly (Nguyen et al. 2019). Acrostalagmus is distributed in soil, freshwater, plant branches, mushrooms, and vermicompost (Nguyen et al. 2019). Currently, 13 taxa are listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp., accessed date: 03 June 2023). Zhang and Tang (2015) reported that A. luteoalbus infects the cultivated mushroom Flammulina velutipes, and causes the substrate bag to be contaminated with the red rust mold. As a result, this pathogen leads to a 40% yield reduction in each production site in Hebei Province, China. Its symptom is that a large number of rust-colored sporulation produced by A. luteoalbus covers the substrate surface, which will inhibit the growth and development of the fruiting body of needle mushroom (Table 7).
Acrostalagmus species causing disease of cultivated mushrooms
Acrostalagmus luteoalbus (Link) Zare, W. Gams & Schroers (Tang et al. 2012; Zhang and Tang 2015).
Hypocreales Lindau.
Albomorchellophilaceae F.M. Yu, K.D. Hyde & Q. Zhao.
Albomorchellophila F.M. Yu, K.D. Hyde & Q. Zhao.
Notes: Albomorchellophila is proposed in this study. It contains only one species that appears on the fruiting bodies of cultivated morels.
Albomorchellophila species causing disease of cultivated mushrooms
Albomorchellophila morchellae F.M. Yu, K.D. Hyde & Q. Zhao (this study).
Bionectriaceae Samuels & Rossman.
Clonostachys Corda.
Notes: Clonostachys Corda, as the asexual morph of Bionectria, was circumscribed by Corda with C. rosea as its type species (Corda 1839; Schroers et al. 1999; Schroers 2001; Perera et al. 2023). According to the International Code of Nomenclature for algae, fungi and plants (McNeill et al., 2012), Rossman et al. (2013) recommend that the name of Clonostachys take precedence over the sexual morph-typified genus Bionectria. Clonostachys species usually have a wide range of lifestyles (endo- and epiphytes, saprotrophs, and destructive mycoparasites), appearing in the bark of dead trees, rotting leaves, nematodes, insects, and rarely in other fungi (Schroers 2001; Chen et al. 2016; Torcato et al. 2020; Zeng and Zhuang 2022; Perera et al. 2023). At present, about 60 scientific names have been placed under Clonostachys (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 2 June 2023). Four asexual species, i.e., C. byssicola, C. pseudosetosa, C. rosea, and C. subquaternata, are associated with other fungi (Sun et al. 2019a, b). Clonostachys rosea is a ubiquitous mycoparasite whose hyphae penetrate and destroy those of many host fungi, including some Mucorales, Ceratocystis fimbriata, Morchella sextelata, Ophiocordyceps sinensis and Trichothecium roseum (Gams et al. 2004; Sun et al. 2019b; Fu et al. 2023).
Clonostachys species causing disease of cultivated mushrooms
Clonostachys rosea (Link) Schroers, Samuels, Seifert & W. Gams (Chen and Huang 2004; Liu et al. 2021a; Fu et al. 2022; this study).
Clonostachys solani (Harting) Schroers & W. Gams (this study).
Sexual morph: Bionectria solani (Reinke & Berthold) Schroers.
Roumegueriella Speg.
Notes: Roumegueriella consists of three species, i.e., R. muricospora (type species), R. pulchella, and R. rufula (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 2 June 2023). Roumegueriella species dwell in soil, and also occur on paper products, rotten leaves and twigs in Asia and Europe. Fletcher and Gaze (2008) reported that R. rufula can form mold disease on compost and substrate of Agaricus bisporus.
Roumegueriella species causing disease of cultivated mushrooms
Roumegueriella rufula (Berk. & Broome) Malloch & Cain (Fletcher and Gaze 2008).
Calcarisporiaceae Jing Z. Sun, X.Z. Liu & K.D. Hyde.
Calcarisporium Preuss.
Notes: Calcarisporium was established by Preuss (1851) and is typified by C. arbuscula. It was assigned to Sordariomycetes genera incertae sedis by De Hoog (1974) based on its morphological features and by Maharachchikumbura et al. (2015, 2016) based on phylogenetic analyses. Sun et al. (2017) provided well-resolved taxonomic placement for Calcarisporium taxa and erected a new family Calcarisporiaceae in Hypocreales to accommodate the sole genus Calcarisporium. Eight species are listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 02 June 2023). Calcarisporium species are fungicolous, and they can occur or parasitize Cordyceps militaris, Hirsutella citriformis, rust fungi, and xylarialean taxa (Sun et al. 2019a). Sun et al. (2016) and Liu et al. (2021a) reported a fungal parasite Calcarisporium cordycipiticola, which infected the fruiting bodies of Cordyceps militaris, causing significant quality and yield losses.
Calcarisporium species causing disease of cultivated mushrooms
Calcarisporium cordycipiticola Jing Z. Sun, C.H. Dong, Xing Z. Liu & K.D. Hyde (Liu et al. 2021a).
Clavicipitaceae (Lindau) Earle ex Rogerson.
Ustilaginoidea Bref.
Notes: Ustilaginoidea currently comprises 15 taxa in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 02 June 2023). Ustilaginoidea virens is a devastating fungal pathogen worldwide and causes rice false smut, pseudo-smut, or green smut (Sun et al. 2020). According to Liu et al. (2021a), U. virens can occur on the fruiting bodies of artificially cultivated Cordyceps militaris. This pathogen occurred sporadically in the early stage of C. militaris cultivation. White velvet hyphae of pathogen appear and occupy the whole substrate, competing with C. militaris hyphae for nutrition (Liu et al. 2021a).
Ustilaginoidea species causing disease of cultivated mushrooms
Ustilaginoidea virens (Cooke) Takah. (Liu et al. 2021a).
Cordycipitaceae Kreisel ex G.H. Sung, J.M. Sung, Hywel-Jones & Spatafora.
Beauveria Vuill.
Notes: Beauveria, as an entomopathogenic genus, is of great economic and ecological value (Sinha et al. 2016; Chen et al. 2019). Some Beauveria species grow naturally in soil worldwide, and act as pathogens on various insect species. Currently, 46 Beauveria species are listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 02 June 2023). Eicker and Greuning (1991) reported Beauveria brongniartii appearing on the substrate of Agaricus bisporus, and forming “flour mold” on its surface (Table 7).
Beauveria species causing disease of cultivated mushrooms
Beauveria brongniartii (Sacc.) Petch (Eicker and Greuning 1991).
Lecanicillium W. Gams & Zare.
Notes: Lecanicillium has been found parasitizing various hosts including arthropods, nematodes, plants, and even fungi (Zare and Gams 2001; Sukarno et al. 2009). It is considered to be entomopathogenic, fungicolous, and nematophagous (Goettel et al. 2008). Currently, there are 19 names in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 02 June 2023). Among them, nine fungicolous Lecanicillium species can parasite macro-fungi, e.g., Agaricus spp., Coltricia perennis, Crinipellis perniciosa, and even micro-fungi, e.g., Phakopsora pachyrhizi, Sphaerotheca fuliginea, and rust fungi (Sun et al. 2019a). Five Lecanicillium taxa are found on cultivated mushrooms or on mushroom composts as competitors or saprobes, i.e., L. antillanum, L. aphanocladii, L. dimorphum, L. fungicola, and L. psalliotae. They form “white mold” or “dry bubble” on host’s fruiting bodies and mycelia. Affected mushrooms involve Agaricus bisporus, Coprinus comatus, Hericium erinaceus, Lentinus edodes, Morchella sextelata, and Pleurotus ostreatus (Table 7).
Lecanicillium species causing disease of cultivated mushrooms
Lecanicillium antillanum (R.F. Castañeda & G.R.W. Arnold) Zare & W. Gams (Kwon et al. 2015).
Lecanicillium aphanocladii Zare & W. Gams (Chen et al. 1984; Lv et al. 2022).
Lecanicillium dimorphum (J.D. Chen) Zare & W. Gams (Chen et al. 1985).
Lecanicillium fungicola (Preuss) Zare & W. Gams (Marlowe 1982; Gea et al. 2003; Zaree et al. 2005; Dong 2013; Hassan 2013).
Lecanicillium psalliotae (Treschew) Zare & W. Gams (Fletcher and Gaze 2008; Kwon et al. 2015).
Simplicillium W. Gams & Zare.
Notes: Zare and Gams (2001) introduced Simplicillium to accommodate four taxa, i.e., S. lanosoniveum (type species), S. lamellicola, S. obclavatum and S. wallacei. Simplicillium species have a wide spectrum of hosts and substrates, e.g., insects, plants, rust fungi, nematodes, animal tissues, mushrooms, algae (Chroococcus spp.), soil, freshwater, marine and even terrene environments (Zare and Gams 2001; Dong et al. 2014; Sun et al. 2019a). Therefore, they are considered to be animal- and plant-parasitic, symbiotic, entomopathogenic, fungicolous and nematophagous fungi. Currently, Simplicillium comprises 21 taxa in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 2 June 2023). Two species are reported as fungicolous, i.e., S. lamellicola and S. lanosoniveum. Simplicillium lamellicola can form gill mildew and brown spots on the fruiting body and compost of Agaricus bisporus (Eicker and Greuning 1991; Zare and Gams 2001).
Simplicillium species causing disease of cultivated mushrooms
Simplicillium lamellicola (F.E.V. Sm.) Zare & W. Gams (Eicker and Greuning 1991; Zare and Gams 2001).
Hypocreaceae De Not.
Hypomyces (Fr.) Tul. & C. Tul.
Asexual morph: Cladobotryum Nees, Mycogone Link, Sepedonium Link, Stephanoma Wallr.
Notes: Hypomyces was originally introduced as a subgenus of Hypocrea (Fries 1825), and then Tulasne and Tulasne (1860) raised it to a genus and designated H. lactifluorum as its type. Almost all Hypomyces species are parasites on other fungi, e.g., Agaricales, Boletales, Helotiales, Pezizales and Polyporales (Rossman et al. 1999; Tamm and Põldmaa 2013; Sun et al. 2019a). Karimi et al. (2022) reported a lignicolous species H. iranica that expanded the host range of Hypomyces from fungi to plants. Currently, there are 118 taxa listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 03 June 2023). Hypomyces species are reported as the causing-agent of cobweb/yellow mold/wet bubble/cotton leak disease of cultivated mushrooms, e.g., Agaricus bisporus, Auricularia spp., Coprinus comatus, Ganoderma spp., Flammulina velutipes, Hypsizigus marmoreus, Lentinula edodes, Morchella spp., Pleurotus spp., and Volvariella volvacea (Table 7).
Hypomyces/Cladobotryum/Sepedonium species causing disease of cultivated mushrooms
Cladobotryum asterophorum de Hoog (Mckay et al. 1999; Back et al. 2012a, b, 2013; Carrasco et al. 2017).
Cladobotryum cubitense R.F. Castañeda & G.R.W. Arnold (Wang et al. 2018).
? Cladobotryum hypsigum (Lee et al. 2015, Back et al. 2015).
Cladobotryum multiseptatum de Hoog (De Hoog 1978; Põldmaa 2011).
Cladobotryum protrusum K. Põldmaa (Wang et al. 2015a, b; Lan et al. 2020).
Hypomyces aurantius (Pers.) Fuckel (Kim et al. 1998, 1999; Mckay et al. 1999; Back et al. 2012a, 2012b; Lee et al. 2015; Zhang et al. 2015; Lan et al. 2019; this study).
Hypomyces chrysospermus Tul. & C. Tul. (Largeteau and Savoie 2010).
Hypomyces mycophilus Rogerson & Samuels (Tubaki 1955; Eicker et al. 1990).
Hypomyces odoratus G.R.W. Arnold (Mckay et al. 1999; Back et al. 2010, 2012a; Gea et al. 2011, 2012, 2017, 2019; Kim et al.2012d, 2014; Lee et al. 2015; Carrasco et al. 2016; Lan et al. 2016; Sharma et al. 2016; Zuo et al. 2016; Chakwiya et al. 2019; Xu et al.2019; Wu and Li 2020; Liu et al. 2023; this study).
Hypomyces perniciosus Magnus (Chang 1978; Fletcher et al. 1995; Glamočlija et al. 2008; Hassan 2013; Naik and Pani 2019).
Hypomyces pseudocorticiicola Tokiwa & Okuda (Okuda et al. 2016).
Hypomyces rosellus (Alb. & Schwein.) Tul. & C. Tul. (Mckay et al. 1999; Zaree et al. 2005; Hassan and Abdulkareem 2013; Potočnik et al. 2015; Sharma et al. 2016; Gea et al. 2018; Naik and Pani 2019; Xu et al. 2020; this study).
Hypomyces semicircularis (G.R.W. Arnold, R. Kirschner & Chee J. Chen) R. Kirschner (Kirschner et al. 2007).
Asexual morph: Cladobotryum semicirculare G.R.W. Arnold, R. Kirschner & Chee J. Chen.
Sepedonium niveum Massee & E.S. Salmon (Botha et al. 1990).
Trichoderma Pers.
Notes: Trichoderma, a species-rich genus in Hypocreaceae, was described by Persoon (1794). It is well adapted to various ecological niches, and ubiquitous in soils, plants, natural or artificial substrate, and other fungi (Zhuang 2020). Species in Trichoderma are considered as widely distributed endophytic, parasitic, and saprobic taxa (Zhuang 2020). Currently, more than 430 taxa are documented in Species Fungorum (https://www.speciesfungorum.org/Names/names.asp?pg=1, accessed date: 3 June 2023). In cultivated mushrooms, Trichoderma is the causative agent of “green mold” disease, which produces visible green sporangium on the fruiting bodies, mycelia and composts (Miles and Chang 2004; Zeng et al. 2022). “Green mold” disease frequently occurs on the wood of shelves, trays, mushrooms bags and fruiting bodies, initially white in color, but turning green within 2–4 days as the spores are produced (Fletcher and Gaze 2008). More than 13 cultivated mushrooms are infected by Trichoderma, which led to a decline in the quality, including Agaricus bisporus, Cyclocybe aegerita, Flammulina velutipes, Ganoderma lingzhi, Hymenopellis raphanipes, Lentinula edodes, and Pleurotus spp. (Table 7).
Trichoderma species causing disease of cultivated mushrooms
Trichoderma aggressivum Samuels & W. Gams (Seaby 1996; Samuels et al. 2002; Hatvani et al. 2007; Baars et al. 2011; Kosanović et al. 2013).
Trichoderma asperellum Samuels, Lieckf. & Nirenberg (Hatvani et al. 2007).
Trichoderma atrobrunneum F.B. Rocha, P. Chaverri & Jaklitsch (Talavera-Ortiz et al. 2020).
Trichoderma atroviride P. Karst. (Hatvani et al. 2007; Choi et al. 2010; Kim et al. 2012a; Wang et al. 2016a).
Trichoderma aureoviride Rifai (Pukahuta et al. 2000; Fletcher and Gaze 2008).
? Trichoderma cf. virens (Choi et al. 2003).
Trichoderma citrinoviride Bissett (Kim et al. 2012a, 2012c).
Trichoderma deliquescens (Sopp) Jaklitsch (Fletcher and Gaze 2008; Kim et al. 2010).
Trichoderma ghanense Yoshim. Doi, Y. Abe & Sugiy. (Hatvani et al. 2007).
Trichoderma hamatum (Bonord.) Bainier (Zargarzadeh et al. 2011).
Trichoderma harzianum Rifai (Wickremasinghe et al. 1999; Pukahuta et al. 2000; Choi et al. 2003, 2010; Zaree et al. 2005; Hatvani et al. 2007, 2012; Xiong 2011; Zargarzadeh et al. 2011; Kim et al. 2012a; Hassan 2013; Kosanović et al. 2013; Wang et al. 2016a; Liu et al. 2021a).
Trichoderma hymenopellicola X.Y. Zeng, X.X. Yuan & F.H. Tian (Zeng et al. 2022).
Trichoderma inhamatum Veerkamp & W. Gams (Zargarzadeh et al. 2011).
Trichoderma koningii Oudem. (Pukahuta et al. 2000; Xiong 2011; Kosanović et al. 2013).
Trichoderma koningiopsis Samuels, Carm. Suárez & H.C. Evans (Kim et al. 2013).
Trichoderma longibrachiatum Rifai (Choi et al. 2003, 2010; Hatvani et al. 2007; Xiong 2011; Kim et al. 2012a; Sharma et al. 2016; Wang et al. 2016a; Zhang et al. 2019).
Trichoderma oblongisporum Bissett (Cao et al. 2014; Wang et al. 2016a).
Trichoderma piluliferum J. Webster & Rifai (Pukahuta et al. 2000).
Trichoderma pleuroti S.H. Yu & M.S. Park [as 'pleurotum'] (Park et al. 2006; Komoń-Zelazowska et al. 2007; Gea 2009; Hatvani et al. 2012; Blaszczyk et al. 2013; Talavera-Ortiz et al. 2020).
Trichoderma pleuroticola S.H. Yu & M.S. Park (Park et al. 2006; Komoń-Zelazowska et al. 2007; Choi et al. 2010; Hatvani et al. 2012; Blaszczyk et al. 2013; Wang et al. 2016a).
Trichoderma pseudogelatinosum (M. Komatsu & Yoshim. Doi) Chang S. Kim [as 'pseudogelatinosa'] (Kim et al. 2012a, 2012b).
Trichoderma pseudokoningii Rifai (Choi et al. 1998; Pukahuta et al. 2000; Fletcher and Gaze 2008).
Trichoderma pseudostramineum (Yoshim. Doi) Chang S. Kim [as 'pseudostraminea'] (Kim et al. 2012a, 2012c).
Trichoderma virens (J.H. Mill., Giddens & A.A. Foster) Arx (Choi et al. 2003; Zargarzadeh et al. 2011; Kosanović et al. 2013).
Trichoderma viride Pers. (Sang et al. 2006; Xiong 2011; Hassan 2013; Wang et al. 2016a).
Trichoderma sp. (Wang et al. 2017).
Trichoderma sp. DAOM 175924 (Kredics et al. 2006; Hatvani et al. 2007).
Myrotheciomycetaceae Crous.
Trichothecium Link.
Notes: This genus was reported by Link (1809a, b). Members of Trichothecium occur on decaying vegetation and in the soil with cosmopolitan distribution (Summerbell et al. 2011). Currently, 26 species are listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 19 June 2023). Some Trichothecium taxa are reported as fungicolous, and its type species T. roseum could parasitize Sclerotinia sclerotiorum (Huang and Kokko 1993). Three species of Trichothecium are regarded as the agents of “white mold” and “pink flour mold” on the fruiting bodies of cultivated Agaricus bisporus, Coprinus comatus, Cordyceps militaris, and Morchella sp. (Table 7).
Trichothecium species causing disease of cultivated mushrooms
Trichothecium crotocinigenum (Schol-Schwarz) Summerb., Seifert & Schroers (Zaree et al. 2005; Liu et al. 2021a).
Trichothecium ovalisporum (Seifert & S.A. Rehner) Seifert & S.A. Rehner (Liu et al. 2021a).
Trichothecium roseum (Pers.) Link (Botha et al. 1990; Zaree et al. 2005; Dong and Bian 2013; this study).
Nectriaceae Tul. & C. Tul.
Aphanocladium W. Gams.
Notes: Aphanocladium was erected by Gams (1971) with the typification A. album. The generic concept of Aphanocladium was revised as having solitary conidia borne on phialide-like conidiogenous cells (Gams 1973; O’Donnell et al. 2011). Currently, four taxa are placed in Aphanocladium (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 03 June 2023), which can be found in other fungi, bark, and fallen leaves. Aphanocladium album can grow on Agaricus bisporus and produce brown spots on the fruiting bodies (Table 7).
Aphanocladium species causing disease of cultivated mushrooms
Aphanocladium album (Preuss) W. Gams (Nair et al. 1980; Eicker and Greuning 1991).
Cosmospora Rabenh.
Notes: Cosmospora was erected by Rabenh (1862) with the typification C. coccinea, and reduced to a subgenus of Nectria by Saccardo (1883). Over a century later, the concept of Cosmospora was redefined by Rossman et al. (1999), which became commonly accepted (Perera et al. 2023). However, phylogenetic analyses revealed that not all species accepted in the genus were monophyletic (Zhang and Zhuang 2006; Luo and Zhuang 2008). Combining molecular data with morphological evidences, Gräfenhan et al. (2011) restricted Cosmospora in a much narrower sense to include only the species growing on polypores and xylariaceous fungi, and having acremonium-like or verticillium-like asexual states. Currently, 50 species are accepted in the genus (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 11 August 2023). Kwon et al. (2015) isolated a strain C. butyri from indoor air of greenhouse of shiitake cultivation. However, the pathogenicity of C. butyri to Lentinula edodes has yet to be confirmed (Table 7).
Cosmospora species causing disease of cultivated mushrooms
Cosmospora butyri (J.F.H. Beyma) Gräfenhan, Seifert & Schroers (Kwon et al. 2015).
Cylindrodendrum Bonord.
Notes: Cylindrodendrum was erected by Bonorden (1851) with C. album as type species. It is regarded as a semi-aquatic saprobe, because its taxa can grow on decaying plant materials in aquatic (including marine and fresh-water) and terrestrial environments (Buffin and Hennebert 1984; Lombard et al. 2014). At present, Cylindrodendrum contains four species, i.e., C. album, C. alicantinum, C. articulatum, and C. orthosporum (= C. hubeiense) (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 03 August 2023). This study finds that C. alicantinum is able to infect the pileus of cultivated morels, and causes white mold-like symptoms.
Cylindrodendrum species causing disease of cultivated mushrooms
Cylindrodendrum alicantinum Agustí-Bris., Armengol & A. Cabral (this study).
Fusarium Link.
Notes: Fusarium is a large cosmopolitan genus and is generally regarded as the most important plant-, mushroom-, and even human-pathogens (Crous et al. 2021). Currently, about 500 records in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 04 June 2023). Several studies reported that Fusarium species cause white mildew/stipe rot/black scar/yellow mold on fruiting bodies and composts of cultivated mushrooms. The infected mushrooms included Agaricus bisporus, Auricularia spp., Cordyceps militaris, Morchella importuna, and Phallus rubrovolvatus (Table 7).
Fusarium species causing disease of cultivated mushrooms
Fusarium chlamydosporum Wollenw. & Reinking (Kong et al. 2011).
Fusarium equiseti (Corda) Sacc. (Guo et al. 2016; Zhang et al. 2017).
Fusarium fujikuroi Nirenberg (Zaree et al. 2005; Li et al. 2022).
Fusarium oxysporum Schltdl. (Fletcher and Gaze 2008; Kong et al. 2011).
Fusarium solani (Mart.) Sacc. (He and Zhang 1995; Sang et al. 2006; Fletcher and Gaze 2008).
Fusarium sporotrichioides Sherb. (Zhang et al. 2017).
Fusarium sp. (Liu et al. 2021a).
Luteonectria Sand.-Den., L. Lombard, Schroers & Rossman.
Notes: Based on analysis of combined ITS, LSU, RPB1, RPB 2 and TEF1 multiple sequence, Crous et al. (2021) erected a new genus Luteonectria to accommodate two combinations Luteonectria albida (type species) and Luteonectria nematophila. Its sexual morph is characterized by off-white to pale luteous perithecia, colour-invariant in KOH or lactic acid, well-developed stroma with clavate asci and fusiform, 3-septate, finely striate ascospores, and its fusarioid asexual morph is characterized by mono-phialides producing robust multi-septate conidia from aerial conidiophores, lacking micro- and meso-conidia, and chlamydospores (Crous et al. 2021). Luteonectria has two species recorded in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 15 August 2023), which occur on bark of woody stem and dwell in soil with roots of Hedera helix (Rossman 1983; Guu et al. 2007; Nirenberg and Hagedorn 2008; Schroers et al. 2011; Crous et al. 2021). Liu et al. (2021b) reported a stipe rot disease caused by Luteonectria nematophila (previously known as Fusarium nematophilum) on cultivated Morchella sextelata in China. The main symptom is the formation of dark-black lesions on the stipe of fruiting bodies, which cause 50%–70% loss in severe cases.
Luteonectria species causing disease of cultivated mushrooms
Luteonectria nematophila (Nirenberg & Hagedorn) Sand.-Den. & L. Lombard (Liu et al. 2021b).
Ophiocordycipitaceae G.H. Sung, J.M. Sung, Hywel-Jones & Spatafora.
Purpureocillium Luangsa-ard, Hywel-Jones, Houbraken & Samson.
Notes: Based on analysis of combined ITS, SSU, and TEF1-α regions, Luangsa-ard et al. (2011) erected a new genus Purpureocillium to accommodate Paecilomyces lilacinus, with a new combination Purpureocillium lilacinum as its type. To date, six Purpureocillium taxa are listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 04 June 2023). Taxa are isolated from a wide range of habitats, including soil, forest, grassland, desert, estuarine sediment and sewage sludge, nematode, plant, insect, other fungi, and even eye of Homo sapiens (Luangsa-ard et al. 2011). Purpureocillium lilacinum can form “purple mold” disease on the compost of Cordyceps militaris, compete with C. militaris mycelia for nutrition, and then cover the host’s mycelia (Liu et al. 2021a). Masaphy (2022) reported P. lilacinum can infect the indoor-cultivated Morchella rufobrunnea. The symptom is that the mycelia of P. lilacinum covers the surface of the growth substrate, as well as the initials and young ascocarps. The infected fruiting body of morels stops growing, turns dark brown and finally disintegrated (Table 7).
Purpureocillium species causing disease of cultivated mushrooms
Purpureocillium lilacinum (Thom) Luangsa-ard, Houbraken, Hywel-Jones & Samson (Liu et al. 2021a; Masaphy 2022).
Pseudodiploosporeaceae Jing Z. Sun, X.Z. Liu & H.W. Liu.
Pseudodiploospora Jing Z. Sun, X.Z. Liu & H.W. Liu.
Notes: Based on the multi-locus phylogenetic analyses and divergence time estimation of combined SSU, ITS, LSU, TEF, and RPB 2 regions in Hypocreales, Sun et al. (2022) proposed a new genus Pseudodiploospora to accommodate four combinations, i.e., P. cubensis, P. longispora, P. fungicola (type species), and P. zinnia. Pseudodiploospora species can be found on other fungi as fungicolous taxa, seeds and dead/fallen leaves of plants (such as Zinnia elegans, Colocasia esculenta var. antiquorum and Leguminosae spp.) as plant-pathogens, on skin and feet as animal-pathogens, and porcupine dung as saprobic fungi (Sun et al. 2022). Pseudodiploospora longispora is the causative agent of “white mold” disease on cultivated Morchella, which results in up to 80% of morel yield losses each year in China (Table 7).
Pseudodiploospora species causing disease of cultivated mushrooms
Pseudodiploospora longispora (Matsush.) Jing Z. Sun, X.Z. Liu & H.W. Liu (He et al. 2017, 2018; Wang et al. 2020; Shi et al. 2022; this study).
Microascales Luttr. ex Benny & Kimbr.
Microascaceae Luttr. ex Malloch.
Cephalotrichum Link.
Notes: Cephalotrichum was described by Link (1809a, b), and is characterized by the formation of dry-spored, indeterminate synnemata and enteroblastic percurrent conidiogenesis. Currently, 38 of Cephalotrichum taxa is listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 4 June 2023). Some Cephalotrichum species occur on decaying plant material, straw, dung, wood and soil (Domsch et al. 2007; Woudenberg et al. 2017), and are infrequently reported from the indoor environment. Three Cephalotrichum species are reported as contaminants that infect the compost of Agaricus bisporus and form black whisker mold (Table 7).
Cephalotrichum species causing disease of cultivated mushrooms
Cephalotrichum microsporum (Sacc.) P.M. Kirk (Eicker and Greuning 1991).
Cephalotrichum purpureofuscum (S. Hughes) S. Hughes (Fletcher and Gaze 2008).
Cephalotrichum stemonitis (Pers.) Nees (Fletcher and Gaze 2008).
Scopulariopsis Bainier.
Notes: Scopulariopsis was erected by Bainier for three asexual taxa S. brevicaulis (the type species), S. rubellus and S. rufulus (Bainier 1907b). The sexual morph of Scopulariopsis has been shown to be Microascus (Curzi 1930, 1931; Abbott et al. 1998; Issakainen et al. 2003). Recently, Microascus and Scopulariopsis were divided into two distinct lineages based on phylogenetic analyses combined four loci ITS + LSU + TEF1-α + β-tubulin (Sandoval-Denis et al. 2016). Some Scopulariopsis species are saprobes commonly isolated from soil, air, and plant debris, as well as paper production, and some are opportunistic pathogens, such as the agent of superficial tissue infections (Sandoval-Denis et al. 2013, 2016). There are 70 members placed in Scopulariopsis (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 4 June 2023). Several studies have reported that S. coprophila is the agent of “white plaster mold” disease, which can grow and infect the fruiting bodies, composts and substrates of cultivated Agaricus bisporus and Flammulina velutipes (Table 7).
Scopulariopsis species causing disease of cultivated mushrooms
Scopulariopsis coprophila (Cooke & Massee) W. Gams (Bhardwaj et al. 1989; Largeteau and Savoie 2010; Hassan 2013; Zhao et al. 2013).
Sordariales Chad. ex D. Hawksw. & O.E. Erikss.
Chaetomiaceae G. Winter.
Chaetomidium (Zopf) Sacc.
Notes: Chaetomidium is characterized by non-ostiolate and globose ascomata, some members having a cephalothecoid peridium, evenly hairy, clavate or sometimes ovoidal (4)–8-spored asci, and limoniform to broadly fusiform, triangular, often bilaterally flattened ascospores (Doveri 2018). Currently, 19 Chaetomidium species are accepted in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 5 June 2023). Chaetomidium thermophilum is the agent of “heat mold” disease, and it can contaminate compost of cultivated Agaricus bisporus (Table 7).
Chaetomidium species causing disease of cultivated mushrooms
Chaetomidium thermophilum (Fergus & Sinden) Lodha (Fergus and Sinden 1969; Fletcher and Gaze 2008).
Chaetomium Kunze.
Notes: Chaetomium was established by Kunze with C. globosum as its generic type (Kunze and Schmidt 1817). It is commonly characterized by ostiolate ascomata, membranaceous perithecial wall covered by well-developed hairs or setae, clavate or fusiform or sometimes cylindrica asci, scarce paraphyses (disappear before ascocarps mature), and 1-celled, smooth and pigmented ascospores (brown or gray-brown) with germ pores (Wang et al. 2016c). Species of Chaetomium are cosmopolitan, and usually as saprobes that are exist in soil, air and the plant debris (Wang et al. 2016c; Abdel-Azeem 2019). Currently, more than 200 species have been described in Chaetomium (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 5 June 2023). Three species are reported as contaminants in mushroom bed, straw and compost of cultivated Agaricus bisporus and Pleurotus ostreatus (Table 7).
Chaetomium species causing disease of cultivated mushrooms
Chaetomium globosum Kunze (Tautorus and Townsley 1983; Botha et al. 1990; Eicker and Greuning 1991; Hassan 2013).
Chaetomium piluliferum J. Daniels (Eicker and Greuning 1991; Fletcher and Gaze 2008).
Chaetomium thermophilum La Touche [as 'thermophile'] (Wickremasinghe et al. 1998).
Myceliophthora Costantin.
Notes: Myceliophthora was introduced by Costantin (1892) to accommodate a mushroom pathogen M. lutea. It is morphologically characterized by the blastic conidia borne terminally or laterally on hyphae, short protrusions or side branches on ampulliform swellings with pyriform to globose conidia (Zhuang et al. 2014; Raze et al. 2019). Myceliophthora species are widespread and associated with plant litter and substrata, especially in high temperature environments (Zhuang et al. 2014; Raze et al. 2019). Currently, Myceliophthora comprises five species, i.e., M. fergusii, M. lutea, M. officinarum, M. sepedonium and M. thermophila (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 5 June 2023). Myceliophthora lutea frequently occurs on compost of cultivated Agaricus bisporus, and causes mat disease and yellow mold (Table 7).
Myceliophthora species causing disease of cultivated mushrooms
Myceliophthora lutea Costantin (Eicker and Greuning 1991; Kamal et al. 2000).
Sordariaceae G. Winter.
Neurospora Shear & B.O. Dodge.
Notes: Neurospora was established by Shear & B.O. Dodge (1927), and was typified by N. sitophila in 1954. Currently, 57 taxa of Neurospora are listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 5 June 2023). Neurospora species are widely distributed, and can be obtained from soil, plant, substrate, and bakeries (Barratt 1974). Neurospora crassa is reported as causing “fire mold” disease on compost of cultivated Agaricus bisporus (Table 7).
Neurospora species causing disease of cultivated mushrooms
Neurospora crassa Shear & B.O. Dodge (Eicker and Greuning 1991; Fletcher and Gaze 2008).
Xylariales Nannf.
Hypoxylaceae DC.
Daldinia Ces. & De Not.
Notes: Daldinia was described by Cesati and De Notaris (1863), and its type species is Daldinia concentrica. Currently, 56 taxa of Daldinia are listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 1 August 2023). Most Daldinia species are saprobes, and several members occur as endophytes (Pažoutová et al. 2013; Samarakoon et al. 2023). Gajanayake et al. (2021) reported that Daldinia eschscholtzii causes fungal contamination in oyster mushroom growing substrate. Daldinia eschscholtzii forms black mycelium in mushroom bags, and rapidly and widely colonizes the substrate and fruit (Gajanayake et al. 2021).
Daldinia species causing disease of cultivated mushrooms
Daldinia eschscholtzii (Ehrenb.) Rehm [as ‘eschscholzii’] (Gajanayake et al. 2021).
Xylariaceae Tul. & C. Tul.
Xylaria Hill ex Schrank.
Notes: Xylaria is the largest genus of the family Xylariaceae. Some xylarialean species are considered as saprotrophic wood-destroyers, while others are ubiquitous as endophytes in plants (Samarakoon et al. 2022). Currently, about 300 accepted species of Xylaria are listed in Species Fungorum (https://www.speciesfungorum.org/Names/names.asp?pg=1, accessed date: 5 June 2023). Xylaria coprinicola is reported as an antagonist for the cultivation of the edible mushroom Coprinus comatus (Table 7).
Xylaria species causing disease of cultivated mushrooms
Xylaria coprinicola Y.M. Ju, H.M. Hsieh & X.S. He (Ju et al. 2011).
Ascomycota genera incertae sedis.
Hormiactis Preuss.
Notes: Hormiactis was established by Preuss (1851). Hormiactis species are distributed worldwide, and usually occur on the dead/rotten trunks, branches and leaves of plants, and even vegetables. Currently, four taxa of Hormiactis are recorded in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 5 June 2023). Hormiactis alba is the agent of cap spot disease in commercial Agaricus bisporus mushroom farm (Cunnington et al. 2008).
Hormiactis species causing disease of cultivated mushrooms
Hormiactis alba Preuss (Fermor 1978; Cunnington et al. 2008).
Basidiomycota R.T. Moore.
Agaricomycetes Doweld.
Agaricales Underw.
Agaricaceae Chevall.
Coprinus Pers.
Notes: Coprinus species are common wild mushrooms worldwide, and they have the liquefied fruiting bodies when mature. Currently, about 150 taxa are listed in Species Fungorum (https://www.speciesfungorum.org/Names/names.asp?pg=1, accessed date: 5 June 2023). Some undetermined Coprinus species are described as ink cap disease on the compost of cultivated Agaricus bisporus (Table 7).
Coprinus species causing disease of cultivated mushrooms
Coprinus spp. (Undetermined species) (Fletcher and Gaze 2008; Hassan 2013).
Schizophyllaceae Quél.
Schizophyllum Fr.
Notes: Schizophyllum members are environmental, wood-rotting taxa, sometimes as plant and animal pathogens, while some are edible (Ohm et al. 2010; Siqueira et al. 2016). Currently, 11 taxa of Schizophyllum are listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 5 June 2023). White mycelium of S. commune can infect and cover the fruiting bodies of Cordyceps militaris, and inhibit the growth of artificially cultivated C. militaris (Table 7).
Schizophyllum species causing disease of cultivated mushrooms
Schizophyllum commune Fr. [as ‘Schizophyllus communis’] (Liu et al. 2021a).
Boletales E.-J. Gilbert.
Coniophoraceae Ulbr.
Coniophora DC.
Notes: Coniophora was introduced by De Candolle (1815) for C. puteana as its type species. Many Coniophora species grow on wood, causing brown rot and serious damage (Kirk et al. 2008). Currently, 26 taxa Coniophora species are recorded in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 5 June 2023). Coniophora puteana var. puteana caused coniophora rot on the compost of cultivated Agaricus bisporus (Table 7).
Coniophora species causing disease of cultivated mushrooms
Coniophora puteana var. puteana (Schumach.) P. Karst. (Fletcher and Gaze 2008).
Cystobasidiomycetes families incertae sedis.
Symmetrosporaceae Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout.
Symmetrospora Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout.
Notes: With the implementation of the “One Fungus = One Name” nomenclatural principle, Wang et al. (2015a, b) revised the yeast taxa in Pucciniomycotina, and the novel family Symmetrosporaceae with sole genus Symmetrospora were established to accommodate the aurantiac taxa in the gracilis/marina clade. Currently, 13 taxa of Symmetrospora are listed in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 5 June 2023), which occur on wilting/rotten leaves and fruits of plants. Symmetrospora symmetrica is the causal agent of red spot disease on cultivated Pleurotus eryngii (Xu et al. 2014). Orange-red spots with a diameter of 2–4 cm are produced on the surface of stipes near pilei of P. eryngii, which directly affects the development of fruiting bodies and causes 10–15% commercial losses (Table 7).
Symmetrospora species causing disease of cultivated mushrooms
Symmetrospora symmetrica (F.Y. Bai & Q.M. Wang) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout (Xu et al. 2014).
Entomophthoromycota Humber.
Entomophthoromycetes Humber.
Entomophthorales G. Winter.
Ancylistaceae J. Schröt.
Conidiobolus Bref.
Notes: Conidiobolus was established to accommodate C. utriculosus (the type species) and C. minor. The genus is characterized by simple conidiophores, globose to pyriform conidia and resting spores formed in the axis of the hypha (Humber 1997). Conidiobolus species are found in soil and decaying plant debris, and are widely distributed, mostly in tropical areas. Members of Conidiobolus are regarded as saprobes, insect pathogens, and even facultative human pathogens (Nie et al. 2002). Currently, Conidiobolus encompasses 34 names in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 9 June 2023). Two Conidiobolus taxa are reported as mycoparasites of Agaricales in India (Srinivasan and Thirumalachar 1965). Fletcher and Gaze (2008) described a third species, C. coronatus, occurring on the compost and fruiting bodies of cultivated Agaricus bisporus (Table 7).
Conidiobolus species causing disease of cultivated mushrooms
Conidiobolus coronatus (Costantin) A. Batko (Fletcher and Gaze 2008).
Mortierellomycota Tedersoo, Sanchez-Ramirez, Kõljalg, Bahram, M. Döring, Schigel, T.W. May, M. Ryberg & Abarenkov.
Mortierellomycetes Doweld.
Mortierellales Caval.-Sm.
Mortierellaceae A. Fisch.
Mortierella Coem.
Notes: Mortierella was circumscribed with a type species M. polycephala (Coemans 1863). Mortierella species are saprotrophic and widespread; some are detected as root endophytes and saprobes in many natural and agricultural habitats, including soil, dung, forest, pasture, cropland, and even plant tissue (Zhang et al. 2020a, b; Ozimek and Hanaka 2021). In some cases, some members of this genus are regarded as the plant growth-promoting taxa (Zhang et al. 2020a, b; Ozimek & Hanaka 2021). Mortierella currently encompasses 80 names in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 6 June 2023). Mortierella polycephala is isolated from Polyporus perennis and Daedalea spp., and eight other species are found to be fungicolous on some basidiomycetes, Chromelosporium spp., and Mucor species (Sun et al. 2019a). Two taxa of Mortierella, M. bainieri and M. reticulata, are reported as causative agents of shaggy stipe in cultivated Agaricus bisporus (Table 7).
Mortierella species causing disease of cultivated mushrooms
Mortierella bainieri Costantin (Fletcher 1973).
Mortierella reticulata Tiegh. & G. Le Monn. (Fletcher and Gaze 2008).
Mucoromycotina Benny.
Mucoromycetes Doweld.
Mucorales Fr.
Lichtheimiaceae Kerst. Hoffm., Walther & K. Voigt.
Rhizomucor Lucet & Costantin.
Notes: Rhizomucor, a filamentous genus, is commonly found in soil, decaying organic matter, and various other environments (Kirk 2008). Some Rhizomucor taxa are human pathogens with high mortality rates, and they can affect the lung, sinuses, brain, and skin (Gomes et al. 2011). Rhizomucor currently encompasses six names in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 9 June 2023). Rhizomucor pusillus is found on the straw and compost of cultivated Pleurotus ostreatus, and causes “rhizomucor mold” disease (Table 7).
Rhizomucor species causing disease of cultivated mushrooms
Rhizomucor pusillus (Lindt) Schipper (Wickremasinghe et al. 1998).
Mucoraceae Dumort.
Mucor Fresen.
Notes: Mucor was described by Fresenius (1850), and it is characterized by simple or branched sporangiophores, non-apophysate and globose sporangia, deliquescent, and incrusted sporangial walls, and zygospores on opposed or tong-like suspensors. Mucor, is the most species-rich genus within Mucoraceae, commonly found in soil, digestive systems, plant surfaces, and decomposed vegetables (Karimi and Zamani 2013). Most Mucor species are saprobes, but also endophytes, parasites of plants and human pathogens (Hurdeal et al. 2021). Mucor currently encompasses more than 100 names in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 6 June 2023). The earliest record showed that M. fusiger infects some species of Agaricales (Willdenow et al. 1833), and five other Mucor taxa are reported to be parasitic on Agaricus spp., Amanita muscaria, Boletus spp., and Rhizopus nigricans (Sun et al. 2019a). Mucor circinelloides and M. racemosus cause mucor disease and infect the mycelia, fruiting bodies and compost of cultivated Pleurotus eryngii and Cyclocybe aegerita (Table 7).
Mucor species causing disease of cultivated mushrooms
Mucor circinelloides Tiegh. (Kim et al. 2013).
Mucor racemosus Fresen. (Choi et al. 2010).
Rhizopodaceae K. Voigt & P.M. Kirk.
Rhizopus Ehrenb.
Notes: Rhizopus is a saprobic genus on fermented foods, plants and specialized parasites on animals. Some species are opportunistic human pathogens that often cause fatal disease called mucormycosis (Zheng et al. 2007). Rhizopus currently encompasses 12 names in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 6 June 2023). Rhizopus nigricans is fungicolous on many other fungi (Rudakov 1978). Sang et al. (2016) reported Rhizopus stolonifera infects the fruiting bodies of cultivated Dictyophora rubrovolvata, causing rot disease (Table 7).
Rhizopus species causing disease of cultivated mushrooms
Rhizopus stolonifer (Ehrenb.) Vuill. (Sang et al. 2006).
Oomycota Arx.
Peronosporomycetes M.W. Dick.
Peronosporales A.N. Beketov.
Pythiaceae Schroet.
Pythium Pringsh.
Notes: Pythium is a worldwide genus which may be destructive saprobes, plant or animal parasites, and mycoparasites. Pythium species are characterized by filamentous sporangia, smooth-walled spherical oogonia, and stalked antheridia. Some species occur in terrestrial habitats and others are aquatic (Gaastra et al. 2010; Koji Kageyama 2014). Pythium currently encompasses 148 names in Species Fungorum (https://www.speciesfungorum.org/Names/Names.asp, accessed date: 06 June 2023). Pythium hydnosporum and P. oligandrum are pathogens of black compost disease, infecting the compost of cultivated Agaricus bisporus (Table 7).
Pythium species causing disease of cultivated mushrooms
Pythium hydnosporum (Mont.) J. Schröt. (Fletcher and Gaze 2008).
Pythium oligandrum Drechsler (Fletcher et al. 1990; Fletcher and Gaze 2008).
Notes: An “?” before the species name indicates that the species is in doubt.
Discussion
With the booming mushroom industries, fungal diseases become problematic. Failure to control early disease outbreaks can be very costly as vast numbers of propagules spread throughout the farm (Fletcher and Gaze 2008; He et al. 2017, 2018; Sun et al. 2022; Fu et al. 2023). Frequent disease outbreaks reduce high yields and incur additional costs for disease control (Fletcher and Gaze 2008). A common and effective way to control pathogens is to use fungal fungicides, although this can cause other problems, such as affecting fruiting body production and increasing pathogens’ resistance to the fungicides (Gea et al. 2021). Many researches have been conducted on various aspects of fungal pathogens on cultivated mushrooms, including species identification, the application of fungal fungicides, and pathologic processes (Sossah et al. 2019; Sun et al. 2019b; Zeng et al. 2022; Fu et al. 2023; Liu et al. 2023; Lu et al. 2023). Nevertheless, cultivated mushrooms are still often disturbed by various fungal diseases. Key problems lie in the gap between disease management practices and pathogens identification. In this study, 133 fungal pathogens on cultivated mushrooms are summarised, most of which were based on morphological identification and culture-based sequencing techniques. However, the number of pathogenic fungi is far more than these according to high-throughput sequencing. Through the ITS amplicon sequencing on the morels ascocarp lesions and culture soils, more potential unreported pathogens were detected (Shi et al. 2022; Yu et al. 2022). Given the essential importance of fungal pathogens for mushroom cultivation, it is necessary to investigate fungal diversity using the most diverse methods, and to address their pathogenicity in the different hosts and emphasize the nutritional relationship between pathogens and cultivated mushrooms.
Species diversity of fungal pathogens on cultivated Morchella
Fungal diseases on cultivated morel have multifarious effects. They lead to fruiting body deformity, large-scale production reduction, and cause devastating damage to the morel industry. Since 2015, due to the uncontrollable spread of fungal diseases, a large number of morel farms have suffered serious losses every year. By 2023, 13 fungal causative agents have been reported to infect the fruiting bodies of cultivated morels, and they mainly are Hypocreales (Summarized by this study). Different pathogens affect different parts of morels, some caps and others are found only on stipes.
Most causal agents of “white mold” disease on cultivated morels were identified as Pseudodiploospora longispora, formerly known as Diploospora longispora. This species was first isolated from a dead leaf of Colocasia esculenta var. antiquorum in Japan (Matsushima 1975), then reported on human skin and feet in Canada (https://www.uamh.ca/index.html). However, with the large-scale cultivation of Morchella, it has become prevalent in China. Previous investigations showed that “white mold” disease resulted in up to 80% of morel yield losses each year, which was attributed to the rapid spread of a large number of conidia around the cultivation areas (Wang et al. 2020; Shi et al. 2022; Sun et al. 2022). In our study, Pseudodiploospora longispora (Pseudodiploosporeaceae) is frequently found on the fruiting bodies of Morchella spp. and Peziza varia in morel’s farm, and it is speculated that this species has host preference on discomycetes. Albomorchellophila morchellae (Albomorchellophilaceae) is another agent of “white mold” disease on cultivated Morchella. At present, it has only been found on morels. Hypomyces / Cladobotryum (Hypocreaceae) taxa are important causal agents of wet bubble/cobweb disease of cultivated mushrooms (Table 7). Four species infect cultivated morels and cause different degrees of damage, i.e., Cladobotryum protrusum, Hypomyces aurantius, H. odoratus and H. rosellus (Lan et al. 2020; Liu et al. 2023; this study). They are generalists as they have a wide range of hosts, from Ascomycota to Basidiomycota (Põldmaa 2011; Zhuang 2020). In Bionectriaceae, Clonostachys solani can be found in the bark of woody plants and from soils (Schroers 2001), and on the fruiting bodies of cultivated morels (Shi et al. 2023), suggesting that it is a facultative species. Clonostachys rosea, a generalist, exists in many habitats, and appears most frequently in soil. Sun et al. (2019b) reported its variant Clo. rosea f. catenulata found on the sclerotium of Ophiocordyceps sinensis. This study adds a new host record of Clo. rosea. Cylindrodendrum (Nectriaceae) has a variety of hosts, which can cause root rot (Agustí-Brisach et al. 2016). In our study, we found Cylindrodendrum alicantinum on the caps of cultivated morels, expanding its host range from plants to fungi. Trichothecium roseum (Myrotheciomycetaceae) is a common facultative pathogen, and causes pink flour mold and stipe canker in Agaricus bisporus, Coprinus comatus, and Morchella sextelata (Botha et al. 1990; Dong and Bian 2013; Zaree et al. 2005).
It is difficult to distinguish the symptoms of different fungal diseases in the cultivation of Morchella. Thus leads to diseases being easily confused and ignored. In a broad sense, “white mold” diseases can be caused by a variety of pathogenic fungi, which can form white mycelial plaques on the fruiting bodies. However, Pseudodiploospora longispora is considered the pathogen of “white mold” disease in a narrow sense because it was the main and most serious fungal pathogen of cultivated Morchella.
Clarify the causative pathogen of “white mold” disease on Morchella
Fungal diseases of cultivated Morchella are easily confused and must not be underestimated. The main disease commonly known as “white mold” is the most serious, and has been reported by many authors (He et al. 2017, 2018; Chen et al. 2019, 2021; Sun et al. 2019a, 2022; Mapook et al. 2022; Shi et al. 2022). However, the wrong identification of agents responsible for “white mold” disease and the mixed names have led to the fungal diseases on Morchella, which have not been clarified so far. He et al. (2017) provided three ITS items (KY490042, KY490043, and KY490044) generated from “white mold” disease on cultivated morels in China, and identified them as Paecilomyces penicillatus based on the ITS phylogeny. Later, He et al. (2018) reported the pileus rot disease on Morchella sextelata in Chongqing and Wuhan, China, and placed these pathogens in Diploospora longispora. They also provided the ITS (KX427537 and KX223838) and LSU (KY765315 and KY765316) information. However, these five ITS genes are identical, indicating that they are produced from the same species. In our phylogeny of Fig. 3, three strains labelled as BH, mp-9 and BHY of Paecilomyces penicillatus (He et al. 2017), and two samples 60319 and 60320 of “Diploospora longispora” in He et al. (2018) nest in Pseudodiploospora longispora lineage. The above evidences clearly show that the cause of “white mold” disease mentioned in these two publications is Ps. longispora (Sun et al. 2022). In addition, “Paecilomyces penicillatus” in Chen et al. (2019, 2021), and “Diploospora longispora” in Sun et al. (2019a), Mapook et al. (2022) and Shi et al. (2022) should also be Ps. longispora according the morphological and molecular traits.
Speculation: ecological process of Pseudodiploospora infecting morel fruiting bodies
Strains of Pseudodiploospora longispora isolated from cultivated morels share the same host habits and ecological niches in China. They all form white, loose fluffy mycelia on morels (mostly on the caps), and may cover the whole fruiting body at the right temperature and humidity. Previous studies have shown that Ps. longispora can be found on skin, foot, humus, decayed mushrooms, seeds and leaves of plants (Matsushima 1975, 1981; Castañeda 1987; Sun et al. 2022). In China, the re-discovery of this species is related to the large-scale cultivation of Morchella throughout the country. It is speculated that this pathogen dwells in the soil and prefers the host of discomycetous taxa. The process of Ps. longispora infecting morels’ fruiting body can be generally divided into three stages (He et al. 2017; Liu et al. 2017; He et al. 2018; Sun et al. 2022): the colonization stage (stage I), the expansion and spore-producing period (stage II), and the withering stage (stage III). Stage I: this stage may occur in the hyphal growth or primordial differentiation of morel. The ambient temperature at this stage is usually < 18 °C. In this process, pathogenic propagules in soil are hypothesized to colonize the primordia or young morels, but no white colonies can be seen by naked eyes. Therefore, it is extremely difficult manage. Stage II: Many white fluffy colonies with abundant conidia powder erupt and grow on the fruiting bodies of morel. The temperature is usually between 18 and 23 °C. During this period, this species undergoes nutrition and reproduction. In severe cases, such as high temperature and high humidity, the pathogen can break out in a short time in the culture shed, and may cover the whole youths to prevent them from growing. Stage III: the temperature at this stage is around 25 °C. With the continuous expansion of the disease spots, the infected parts of morel rot and wither, and even lead to deformity and decay of the whole fruit body.
In the fungus-fungus interaction, the successful infection caused by pathogen usually goes through host recognition, attachment, infection and degradation, which depends on the gene content, expression or regulation (Quandt et al. 2016). Chen et al. (2021) studied a set of genes differentially regulated in the infection mechanism of Pseudodiploospora longispora (it is called Paecilomyces penicillatus in the publication), and the response mechanism of Morchella importuna, which filled the gaps in the fungal pathogen-morel interaction. They found genes had variable expression trends at different stages of infection, indicating P. longispora had diverse regulation patterns to infect M. importuna. Infection mechanism involves diphthamide biosynthesis, aldehyde reductase, and NAD (P) H-hydrate epimerase, and the response mechanism includes cyclin-dependent protein kinase inhibitor, and enriched fatty acid biosynthesis and metabolism pathways (Chen et al. 2021).
Future research of fungal diseases on cultivated mushrooms
Accurately identifying the pathogens or competitors in the mushroom industry is the crucial first step, so that effective control measures can be developed later to better control disease. Second, understanding the mechanism of pathogens is as important as clarifying the pathogen–mushroom interactions. Recently, genomes of Cladobotryum dendroides, Cladobotryum mycophilum, Cladobotryum protrusum, and Hypomyces perniciosus showed a large number of the predicted expanded gene families, including secreted peptidases, carbohydrate-active enzymes, and cytochrome P450 enzymes, which are essential for mycoparasitism (Li et al. 2019; Sossah et al. 2019; Xu et al. 2020; Liu et al. 2023). In addition, it is also important to determine whether fungal pathogens are ubiquitous or host specific (Sun et al. 2019a). For the sustainable development of mushroom cultivation industry, researchers should give priority to the research of fungal pathogens on mushrooms (wild and cultivated mushrooms), including species identification, symptom description, Koch's postulates, disease control, infection mechanism, and the pathogen-host interactions.
Conclusion
This study focuses on fungal pathogens affecting cultivated mushrooms, with an emphasis on Morchella. The results show that the pathogenetic fungi related to cultivated mushrooms have high species diversity. A total of 133 pathogenic species have been reported on cultivated mushrooms, involving six phyla of fungi Kingdom. Among them, more than half (70/133, 52.6%) are concentrated in Hypocreales (Sordariomycetes). Up to 23 kinds of mushrooms are infected by these pathogens, accounting for one third of commercially cultivated mushrooms in the world. The cultivated mushrooms most reported to be affected by fungal pathogens are Agaricus bisporus, Lentinus edodes and Morchella species. Despite all this, research on fungal pathogen of cultivated mushrooms is still in the primary stage. The potential diseases caused by fungi still need further investigation, as well as the diseases on wild mushroom should also be given attention to. In addition, the follow-up research should focus on the biological characteristics of typical fungal pathogens, such as origin, transmission, pathogenesis, and the pathogen-mushroom interactions. For cultivated morels, it is necessary to study pathogenic microorganisms (including fungi, bacteria and viruses), and carry out targeted research to control the spread or growth of harmful diseases.
Data availability
All sequences have been deposited at GenBank database (Tables S1 and S2).
References
Abbott SP, Sigler L, Currah RS (1998) Microascus brevicaulis sp. nov., the teleomorph of Scopulariopsis brevicaulis, supports placement of Scopulariopsis with the Microascaceae. Mycologia 90:297–302
Abdel-Azeem AM (2019) Recent developments on genus Chaetomium. Springer, Cham
Agustí-Brisach C, Cabral A, González-Domínguez E, Pérez-Sierra A, León M, Abad-Campos P, García-Jiménez J, Oliveira H, Armengol J (2016) Characterization of Cylindrodendrum, Dactylonectria and Ilyonectria isolates associated with loquat decline in Spain, with description of Cylindrodendrum alicantinum sp. nov. Eur J Plant Pathol 145:103–118. https://doi.org/10.1007/s10658-015-0820-7
Anonymous (1955) Shen Nong materia medica, 102-200 AD (E. Reprinted. People's Hygiene Press, Beijing, Han)
Błaszczyk L, Siwulski M, Sobieralski K, Frużyńska-Jóźwiak D (2013) Diversity of Trichoderma spp. causing Pleurotus green mould diseases in Central Europe. Folia Microbiol 58:325–333. https://doi.org/10.1007/s12223-012-0214-6
Baars J, Rutjens J, Mumm R (2011) Can volatiles emitted by compostduring spawn run be used to detect green mould infection early? In: Savoie J-M, Foulongne-Oriol M, Largeteau M, Barroso, G. (eds) Proceedings of the 7th International Conference on MushroomBiology and Mushroom Products (ICMBMP7), Vol 1. INRA, Villenave d'Ornon Cedex 474–483.
Back CG, Kim YH, Jo WS, Chung HJ (2010) Cobweb disease on Agaricus bisporus caused by Cladobotryum mycophilum in Korea. J Gen Plant Pathol 76:232–235. https://doi.org/10.1007/s10327-010-0236-3
Back CG, Lee CY, Seo GS, Jung HY (2012a) Characterization of species of Cladobotryum which cause cobweb disease in edible mushrooms grown in Korea. Mycobiology 40:189–194. https://doi.org/10.5941/MYCO.2012.40.3.189
Back CG, Lee CY, Seo GS, Jung HY, Ohga S (2012b) First Occurrence of cobweb disease on Hypsizigus marmoreus caused by Cladobotryum varium in Korea. J Fac Agric Kyushu Univ 57:373–377. https://doi.org/10.5109/25193
Back CG, Lee SY, Lee CY, Seo GS, Jung HY (2013) Topic F: Infection and Pathogenesis: New Cladobotryum Species causing Cobweb Disease in Beech Mushroom. 균학회소식 25:96–96.
Bainier G (1907a) Mycothèque de l’École de Pharmacie. XI. Paecilomyces, genre nouveau de Mucédinées. Bull Soc Mycol Fr 23:26–27
Bainier G (1907b) Mycothèque de lÉcole de Pharmacie, XII-XVI. Bull Soc Mycol Fr 23:90–110
Baral HO, Weber E, Gams W, Hagedorn G, Liu B, Liu X, Marson G, Marvanová L, Stadler M, Weiß M (2018) Generic names in the Orbiliaceae (Orbiliomycetes) and recommendations on which names should be protected or suppressed. Mycol Prog 17:5–31
Barratt RW (1974) Neurospora crassa. In: King RC (ed) Bacteria, Bacteriophages, and Fungi. Springer, Boston. https://doi.org/10.1007/978-1-4899-1710-2_27
Bensch K, Braun U, Groenewald JZ, Crous PW (2012) The genus Cladosporium. Stud Mycol 72:1–401. https://doi.org/10.3114/sim0003
Bhardwaj SC, Seth PK, Guleria DS (1989) A note on the occurrence of white plaster disease of cultivated mushrooms (Agaricus bisporus) caused by Scopulariopsis fimicola—a new fungus from India. Curr Sci Bangalore 58:320–321
Binit K, Lal AA, Sobita S, Shivam S (2013) Papulaspora byssina on substrate of milky mushroom (Calocybe indica). Ann Plant Prot Sci 21:210–211
Botha WJ, Eicker A, Meillon SD (1990) A comparative physiological study of four weed moulds of Agaricus bisporus mushroom compost. Phytophylactica 22:219–223
Bouckaert R, Heled J, Kühnert D, Vaughan T, Wu CH, Xie D, Suchard MA, Rambaut A, Drummond AJ (2014) BEAST 2: a software platform for Bayesian evolutionary analysis. PLoS Comput Biol 10:e1003537. https://doi.org/10.1371/journal.pcbi.1003537
Bouckaert R, Vaughan TG, Barido-Sottani J, Duchêne S, Fourment M, Gavryushkina A, Heled J, Jones G, Kühnert D, De Maio N (2019) BEAST 2.5: an advanced software platform for Bayesian evolutionary analysis. PLoS Comput Biol 15:650. https://doi.org/10.1371/journal.pcbi.1006650
Bowman BH, White TJ, Taylor JW (1996) Human pathogeneic fungi and their close nonpathogenic relatives. Mol Phylogenet Evol 6:89–96. https://doi.org/10.1006/mpev.1996.0061
Cao XT, Bian YB, Xu ZY (2014) First report of Trichoderma oblongisporum causing green mold disease on Lentinula edodes (shiitake) in China. Plant Dis 98:1440. https://doi.org/10.1094/PDIS-05-14-0537-PDN
Carrasco J, Navarro MJ, Gea FJ (2017) Cobweb, a serious pathology in mushroom crops: a review. Spanish J Agric Res 15:e10R01. https://doi.org/10.5424/sjar/2017152-10143
Carrasco J, Navarro MJ, Santos M, Diánez F, Gea FJ (2016) Incidence, identification and pathogenicity of Cladobotryum mycophilum, causal agent of cobweb disease on Agaricus bisporus mushroom crops in Spain. Ann Appl Biol 168:214–224. https://doi.org/10.1111/aab.12257
Castañeda-Ruiz RF, Iturriaga T, Heredia Abarca G, Minter DW, Gene J, Stadler M, Saikawa M, Silvera-Simon C (2008) Notes on Heteroconium and a new species from Venezuela. Mycotaxon 105:175–184
Castañeda R (1987). Fungi cubenses II: instituto de investigaciones fundamentales en agricultura tropical “Alejandro Humboldt”. Cuba (Academia de Ciencas de Cuba): la Habana
Cesati V, De Notaris G (1863) Schema di classificazione degle sferiacei italici aschigeri piúo meno appartenenti al genere Sphaeria nell’antico significato attribuitoglide Persono. Comm Soc Crittog 1:177–420
Chakwiya A, Van der Linde EJ, Chidamba L, Korsten L (2019) Diversity of Cladobotryum mycophilum isolates associated with cobweb disease of Agaricus bisporus in the South African mushroom industry. Eur J Plant Pathol 154:767–776. https://doi.org/10.1007/s10658-019-01700-7
Chang ST (1978) Volvariella volvacea. The biology and cultivation of edible mushrooms. 573–600.
Chang ST, Wasser SP (2017) The cultivation and environmental impact of mushrooms. In: Oxford Research Encyclopedia of Environmental Science. CRC Press, Boca Raton
Chen C, Fu R, Wang J, Li X, Chen X, Li Q, Lu D (2021) Genome sequence and transcriptome profiles of pathogenic fungus Paecilomyces penicillatus reveal its interactions with edible fungus Morchella importuna. Comput Struct Biotechnol J 19:2607–2617. https://doi.org/10.1016/j.csbj.2021.04.065
Chen JD, Liu JG, Chen SS, Cai FJ, Zhang ZC (1984) Studies on Aphanocladium aranearum (Petch) gams var sinense var. nov.—a fungus parasitic on Agaricus bisporus (Lange) Sing. Acta Mycol Sin 3:96
Chen JD, Liu JG, Chen SS, Cai FJ, Zhang ZC (1985) A new species of Aphanocladium on Agaricus bisporus. Acta Mycologica Sinica 4:227–233
Chen JT, Huang JW (2004) Identification of Gliocladium roseum, the causal agent of brown spot of the king oyster mushroom Pleurotus eryngii. Plant Pathol Bull 13:17–26 ((In Chinese))
Chen WH, Han YF, Liang JD, Zou X, Liang ZQ, Jin DC (2016) A new araneogenous fungus of the genus Clonostachys. Mycosystema 35:1061–1069 ((In Chinese))
Chen ZH, Chen K, Dai YD, Zheng Y, Wang YB, Yang XN, Yu H, Yang YM, Xu L (2019) Beauveria species diversity in the Gaoligong Mountains of China. Mycol Prog 18:933–943. https://doi.org/10.1007/s11557-019-01497-z
Chethana KWT, Manawasinghe IS, Hurdeal VG, Bhunjun CS, Appadoo MA, Gentekaki E, Raspé O, Promputtha I, Hyde KD (2021) What are fungal species and how to delineate them? Fungal Diversity 109:1–25. https://doi.org/10.1007/s13225-021-00483-9
Choi IY, Choi JN, Sharma PK, Lee WH (2010) Isolation and identification of mushroom pathogens from Agrocybe aegerita. Mycobiology 38:310–315. https://doi.org/10.4489/MYCO.2010.38.4.310
Choi IY, Hong SB, Yadav MC (2003) Molecular and morphological characterization of green mold, Trichoderma spp. isolated from oyster mushrooms. Mycobiology 31:74–80. https://doi.org/10.4489/MYCO.2003.31.2.074
Choi IY, Lee WH, Choi JS (1998) Forest green mold disease caused by Trichoderma pseudokoningii in winter mushroom, Flammulina velutipes. Korean J Mycol 26:531–537
Coemans E (1863) Quelques Hyphomycètes nouveaux. 1. notice: Mortierella polycephala et Martinella pectinata. Bulletin De L’académie Royale Des Sciences De Belgique Classe Des Sciences 15:536–540
Coetzee JC, Eicker A (1990) Chromelosporium fulvum in the mushroom industry: a review. Suid-Afrikaanse Tydskrif Vir Natuurwetenskap En Tegnologie 9:112–115
Corda ACJ (1833) Deutschlands Flora, Abt. III. Die Pilze Deutschlands 3-13:65–96.
Corda ACJ (1838) Icones fungorum hucusque cognitorum 2:1–43.
Corda ACJ (1839) Pracht-Flora Europaeischer Schimmelbildungen 1–55.
Costantin J (1892) Sur quelques maladies du blanc de Champignon. Comptes Rendus Hebdomadaires Des Séances De L’academie Des Sciences 114:849–851
Crous PW, Lombard L, Sandoval-Denis M, Seifert KA, Schroers HJ, Chaverri P, Gené J, Guarro J, Hirooka Y, Bensch K, Kema GHJ, Lamprecht SC, Cai L, Rossman AY, Stadler M, Summerbell RC, Taylor JW, Ploch S, Visagie CM, Yilmaz N, Frisvad JC, Abdel-Azeem AM, Abdollahzadeh J, Abdolrasouli A, Akulov A, Alberts JF, Araújo JPM, Ariyawansa HA, Bakhshi M, Bendiksby M, Ben Hadj Amor A, Bezerra JDP, Boekhout T, Câmara MPS, Carbia M, Cardinali G, Castañeda-Ruiz RF, Celis A, Chaturvedi V, Collemare J, Croll D, Damm U, Decock CA, de Vries RP, Ezekiel CN, Fan XL, Fernández NB, Gaya E, González CD, Gramaje D, Groenewald JZ, Grube M, Guevara-Suarez M, Gupta VK, Guarnaccia V, Haddaji A, Hagen F, Haelewaters D, Hansen K, Hashimoto A, Hernández-Restrepo M, Houbraken J, Hubka V, Hyde KD, Iturriaga T, Jeewon R, Johnston PR, Jurjević Ž, Karalti İ, Korsten L, Kuramae EE, Kušan I, Labuda R, Lawrence DP, Lee HB, Lechat C, Li HY, Litovka YA, Maharachchikumbura SSN, Marin-Felix Y, Matio Kemkuignou B, Matočec N, McTaggart AR, Mlčoch P, Mugnai L, Nakashima C, Nilsson RH, Noumeur SR, Pavlov IN, Peralta MP, Phillips AJL, Pitt JI, Polizzi G, Quaedvlieg W, Rajeshkumar KC, Restrepo S, Rhaiem A, Robert J, Robert V, Rodrigues AM, Salgado-Salazar C, Samson RA, Santos ACS, Shivas RG, Souza-Motta CM, Sun GY, Swart WJ, Szoke S, Tan YP, Taylor JE, Taylor PWJ, Tiago PV, Váczy KZ, van de Wiele N, van der Merwe NA, Verkley GJM, Vieira WAS, Vizzini A, Weir BS, Wijayawardene NN, Xia JW, Yáñez-Morales MJ, Yurkov A, Zamora JC, Zare R, Zhang CL, Thines M (2021) Fusarium: more than a node or a foot-shaped basal cell. Stud Mycol 98:100116. https://doi.org/10.1016/j.simyco.2021.100116
Crous PW, Schumacher RK, Wingfield MJ, Lombard L, Giraldo A, Christensen M, Gardiennet A, Nakashima C, Pereira OL, Smith AJ, Groenewald J (2015) Fungal systematics and evolution: FUSE 1. Sydowia Horn 67:81–118. https://doi.org/10.12905/0380.sydowia67-2015-0081
Cruickshank RD, Ko K (2003) Geology of an amber locality in the Hukawng Valley, northern Myanmar. J Asian Earth Sci 21:441–455. https://doi.org/10.1016/S1367-9120(02)00044-5
Cunnington JH, Ratnayake K, Salib S, Irvine G, Priest MJ, Shivas RG (2008) First record of Hormiactis cap spot of mushrooms in Australia. Aust Plant Dis Notes 3:19–20. https://doi.org/10.1071/DN08008
Curzi M (1930) Una nuova specie di Microascus. Bolletino Della Stazione Patol Veg Roma 10:302–309
Curzi M (1931) Rapporti fra i generi Microascus Zukal e Scopulariopsis Bainier. Bolletino Della Stazione Patol Veg Roma 11:55–60
D’Alessandro CP, Jones LR, Humber RA, López Lastra CC, Sosa-Gomez DR (2014) Characterization and phylogeny of Isaria spp. strains (Ascomycota: Hypocreales) using ITS1-5.8S-ITS2 and elongation factor 1-alpha sequences. J Basic Microbiol 54:S21-31. https://doi.org/10.1002/jobm.201300499
Dar GM, Beigh GM, Mir NA (2012) Influence of some mushroom cultivation factors on the development of brown plaster mould (Papulaspora byssina) on mushroom beds in Kashmir. Integrated disease management and plant health. 284–289.
De Hoog GS (1974) The genera Blastobotrys, Sporothrix, Calcarisporium and Calcarisporiella gen. nov. Stud Mycol 7:1–83. https://doi.org/10.1016/s0007-1536(75)80099-4
De Hoog GS (1978) Notes on some fungicolus hyphomycetes and their relatives. Persoonia Mol Phylogeny Evol Fungi 10:33–81
Desmazières JBHJ (1827) Observations sur le Sporendonema casei, nouveau genre des Mucédinées. Ann Sci Nat Bot 11:246–249
do Nascimento JS, da Eira AF (2003) Occurrence of the false truffle (Diehliomyces microsporus Gilkey) and damage on the himematsutake medicinal mushroom (Agaricus brasiliensis S. Wasser (et al.). Int J Med Mushrooms 5:8. https://doi.org/10.1615/IntJMedMushr.v5.i1.100
Dong QL, Dong RZ, Xing XY, Li YK (2018) A new antibiotic produced by the cyanobacterium-symbiotic fungus Simplicillium lanosoniveum. Nat Prod Res 32:1348–1352. https://doi.org/10.1080/14786419.2017.1343320
Dong WH, Bian YB (2013) Stipe canker caused by Trichothecium roseum on the edible shaggy mane Coprinus comatus in China. Plant Dis 97:1507–1507. https://doi.org/10.1094/PDIS-03-13-0344-PDN
Doveri F (2018) On a new species of Chaetomidium, C. vicugnae, with a cephalothecoid peridium and its relationships with Chaetomiaceae (Sordariales). Ascomycete.org 10:86–96. https://doi.org/10.25664/art-0231
Du XH, Yang ZL (2021) Mating Systems in True Morels (Morchella). Microbiol Mol Biol Rev 85:e00220-e320. https://doi.org/10.1128/MMBR.00220-20
Eicker A, Peng JT, Chen ZC (1990) A Pseudohansfordia-disease of sawdust-cultivated Auricularia mesenterica in Taiwan. Bot Bull Acad Sin 31:205–210
Eicker A, van Greuning M (1991) Fungi in the cultivation of Agaricus bisporus—an undated list of species. a Pudoc Wageningen 199:88.
Elad Y, Williamson B, Tudzynski P, Delen N (2004) Botrytis: biology, pathology and control. Springer, New York
Fazenda ML, Seviour R, McNeil B, Harvey LM (2008) Submerged culture fermentation of “higher fungi”: the macrofungi. Adv Appl Microbiol 63:33–103. https://doi.org/10.1016/S0065-2164(07)00002-0
Fergus CL, Sinden JW (1969) A new thermophilic fungus from mushroom compost: Thielavia thermophila spec. nov. Can J Bot 47:1635–1637. https://doi.org/10.1139/b69-235
Fermor TR (1978) Hormiactis alba, an uncommon fungal pathogen of the cultivated mushroom Agaricus bisporus. Glasshouse Crops Research Institute Annual Report 191–193
Fletcher JT (1973) Shaggy stipe, a new disease of cultivated mushroom caused Mortierella bainieri. Plant Pathol 22:25–27. https://doi.org/10.1111/j.1365-3059.1973.tb01763.x
Fletcher JT, Gaze RH (2008) Mushroom pest and disease control, a colour handbook. Academic Press, San Diego
Fletcher JT, Jaffe B, Muthumeenakshi S, Brown AE, Wright DM (1995) Variations in isolates of Mycogone perniciosa and in disease symptoms in Agaricus bisporus. Plant Pathol 44:130–140. https://doi.org/10.1111/j.1365-3059.1995.tb02725.x
Fletcher JT, Smewin BJ, O’Brien A (1990) Pythium oligandmm associated with a cropping disorder of Agaricus bisporus. Plant Pathol 39:603–605. https://doi.org/10.1111/j.1365-3059.1990.tb02540.x
Fresenius G (1850) Beiträge zur Mykologie 1:1–38
Fu Y, Xu X, Wu H, Li L, Wang J, Sun Y, Gu L, Yu Q (2022) First Report of Clonostachys rosea Causing rot of Morchella sextelata in Anhui Province, China. China. Plant Dis 1:10. https://doi.org/10.1094/PDIS-08-22-1794-PDN
Fuckel L (1867) Fungi Rhenani Exsiccati. Supplementi. Fasc. 5:no. 1901-no. 2000
Gaastra W, Lipman LJA, De Cock AWAM, Exel TK, Pegge RBG, Scheurwater J, Vilela R, Mendoza L (2010) Pythium insidiosum: an overview. Vet Microbiol 146:1–16. https://doi.org/10.1016/j.vetmic.2010.07.019
Gajanayake AJ, Samarakoon MC, Bundhun D, Jayawardena RS, Luangharn T, Hyde KD (2021) A challenge for mushroom growers: Daldinia eschscholtzii associated with oyster mushroom growing substrate. Chiang Mai J Sci 48:774–786
Gams W (1971) Cephalosporium-artige Schimmelpilze. 1–262
Gams W (1973) Phialides with solitary conidia? Remarks on conidium ontogeny in some hyphomycetes. Persoonia 7:161–169
Gams W, Diederich P, Põldmaa K (2004) Fungicolous fungi. In: Mueller GM, Bills GF, Foster MS (eds) Biodiversity of fungi. Inventory and monitoring methods. Elsevier Academic Press, Burlington, pp 343–392
Gams W, O’Donnell K, Schroers HJ, Christensen M (2011) Generic classification of some more hyphomycetes with solitary conidia borne on phialides. Can J Bot. https://doi.org/10.1139/cjb-76-9-1570
Gardes M, Bruns TD (1993) ITS primers with enhanced specifity for Basidiomycetes: application to identification of mycorrhizae and rusts. Mol Ecol 2:113–118. https://doi.org/10.1111/j.1365-294x.1993.tb00005.x
Gea FJ (2009) First report of Trichoderma pleurotum on oyster mushroom crops in Spain. J Plant Pathol 91:504
Gea FJ, Carrasco J, Suz LM, Navarro MJ (2017) Characterization and pathogenicity of Cladobotryum mycophilum in Spanish Pleurotus eryngii mushroom crops and its sensitivity to fungicides. Eur J Plant Pathol 147:129–139. https://doi.org/10.1007/s10658-016-0986-7
Gea FJ, Navarro MJ, Carrasco J, González AJ, Suz LM (2012) First report of cobweb on white button mushroom (Agaricus bisporus) in Spain caused by Cladobotryum mycophilum. Plant Dis 96:1067. https://doi.org/10.1094/PDIS-02-12-0120-PDN
Gea FJ, Navarro MJ, Suz LM (2018) First report of cobweb disease caused by Cladobotryum dendroides on shiitake mushroom (Lentinula edodes) in Spain. Plant Dis 102:1030. https://doi.org/10.1094/PDIS-09-17-1481-PDN
Gea FJ, Navarro MJ, Suz LM (2019) Cobweb disease on oyster culinary-medicinal mushroom (Pleurotus ostreatus) caused by the mycoparasite Cladobotryum mycophilum. J Plant Pathol 101:349–354. https://doi.org/10.1007/s42161-0180-0174-z
Gea FJ, Tello JC, Navarro MJ (2003) Occurrence of Verticillium fungicola var. fungicola on Agaricus bitorquis mushroom crops in Spain. J Phytopathol 151:98–100. https://doi.org/10.1046/j.1439-0434.2003.00687.x
Geng YH, Wu YM, Zhang TY (2016) Three new species of Scytalidium from soil in Tibet of China. Mycosystema 35:1310–1316. https://doi.org/10.13346/j.mycosystema.160102
Gilkey HM (1954) Taxonomic notes on Tuberales. Mycologia 46:783–793
Glamočlija J, Soković M, Ljaljević-Grbić M, Vukojević J, Milenković I, Van Griensven L (2008) Morphological characteristics and mycelial compatibilityof different Mycogone perniciosa isolates. J Microsc 232:489–492. https://doi.org/10.1111/j.1365-2818.2008.02145.x
Goettel MS, Koike M, Kim JJ, Aiuchi D, Shinya R, Brodeur J (2008) Potential of Lecanicillium spp. for management of insects, nematodes and plant diseases. J Invertebr Pathol 98:256–261
Gomes MZ, Lewis RE, Kontoyiannis DP (2011) Mucormycosis caused by unusual mucormycetes, non-Rhizopus, -Mucor, and -Lichtheimia species. Clin Microbiol Rev 24:411–445. https://doi.org/10.1128/CMR.00056-10
Gräfenhan T, Schroers HJ, Nirenberg HI, Seifert KA (2011) An overview of the taxonomy, phylogeny and typification of nectriaceous fungi in Cosmospora, Acremonium, Fusarium, Stilbella and Volutella. Stud Mycol 68:79–113
Grove WB (1916) New or noteworthy fungi. Part v. Lond J Bot 54:220
Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O (2010) New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol 59:307–321. https://doi.org/10.1093/sysbio/syq010
Guo MP, Chen K, Wang GZ, Bian YB (2016) First report of stipe rot disease on Morchella importuna caused by Fusarium incarnatum-F. equiseti species complex in China. Plant Dis 100:2530. https://doi.org/10.1094/PDIS-05-16-0633-PDN
Guu JR, Ju YM, Hsieh HJ (2007) Nectriaceous fungi collected from forests in Taiwan. Bot Stud 48:187–203
Hall T (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 41:95–98
Haller A von (1768) Historia stirpium indigenarum Helvetiae inchoate 3:1–204.
Hassan AA (2013) First report of diseases and pests of an edible mushroom Agaricus bisporus in Iraq1.The fungal infections. Tikrit J Agric 13:60–71
Hatvani L, Antal Z, Manczinger L, Szekeres A, Druzhinina IS, Kubicek CP, Nagy A, Nagy EV, Kredics L (2007) Green mold diseases of Agaricus and Pleurotus spp. are caused by related but phylogenetically different Trichoderma species. Phytopathology 97:532–537. https://doi.org/10.1094/PHYTO-97-4-0532
Hatvani L, Sabolić P, Koscubé S, Kredics L, Czifra D, Vágvölgyi C, Kaliterna J, Ivić D, Dermić E, Kosalec I (2012) The first report on mushroom green mould disease in Croatia. Arch Ind Hyg Toxicol 63:481–487. https://doi.org/10.2478/10004-1254-63-2012-2220
Hawksworth DL (2012) Global species numbers of fungi: are tropical studies and molecular approaches contributing to a more robust estimate? Biodivers Conserv 21:l2425-2433. https://doi.org/10.1007/s10531-012-0335-x
He P, Li C, Cai Y, Zhang Y, Bian Y, Liu W (2018) First report of pileus rot disease on cultivated Morchella importuna caused by Diploöspora longispora in China. J Gen Plant Pathol 84:65–69. https://doi.org/10.1007/s10327-017-0754-3
He XL, Peng WH, Miao RY, Tang J, Chen Y, Liu LX, Wang D, Gan BC (2017) White mold on cultivated morels caused by Paecilomyces penicillatus. FEMS Microbiol Lett. https://doi.org/10.1093/femsle/fnx037
He XS, Zhang L (1995) Study of Fusarium Pathogen on Auricularia polytricha Cultivated with Plastic Bag in Sichuan Province. Acta Edulis Fungi 2:44–47
Helfer W (1991) Pilze auf Pilzfruchtkӧrpern. Untersuchungen Zurӧkologie. Syst Chem Libri Bot 1:1–157
Ho LH, Zulkifli NA, Tan TC (2020) Edible Mushroom: Nutritional Properties, Potential Nutraceutical Values, and Its Utilisation in Food Product Development. In An Introduction to Mushroom. IntechOpen. https://doi.org/10.5772/intechopen.91827
Hoang DT, Chernomor O, Von Haeseler A, Minh BQ, Vinh LS (2018) UFBoot2: improving the ultrafast bootstrap approximation. Mol Biol Evol 35:518–522. https://doi.org/10.1093/molbev/msx281
Hongsanan S, Sánchez-Ramírez S, Crous PW, Ariyawansa HA, Zhao RL, Hyde KD (2016) The evolution of fungal epiphytes. Mycosphere 7:1690–1712. https://doi.org/10.5943/mycosphere/7/11/6
Houbraken J, Seifert KA, Samson RA (2019) Penicillium hermansii, a new species causing smoky mould in white button mushroom production. Mycol Prog 18:229–236. https://doi.org/10.1007/s11557-018-1407-4
Houbraken J, Frisvad JC, Samson RA (2011) Taxonomy of Penicillium section Citrina. Stud Mycol 70:53–138
Hu XJ, Webster G, Xie LH, Yu CB, Li YS, Liao X (2013) A new mycoparasite, Aspergillus sp. ASP-4, parasitizes the sclerotia of Sclerotinia sclerotiorum. Crop Prot 54:15–22
Huang HC, Kokko EG (1993) Trichothecium roseum, a mycoparasite of Sclerotinia sclerotiorum. Can J Bot 71:1631–1638
Hubka V, Nováková A, Kolařík M, Jurjević Ž, Peterson SW (2015) Revision of Aspergillus section Flavipedes: seven new species and proposal of section Jani sect. nov. Mycologia 107:169–208. https://doi.org/10.3852/14-059
Humber RA (1997) Fungi: identification. In: Lacey LA (ed) Manual of techniques in insect pathology. Academic Press, London
Hurdeal VG, Gentekaki E, Hyde KD, Nguyen TTT, Lee HB (2021) Novel Mucor species (Mucoromycetes, Mucoraceae) from northern Thailand. MycoKeys 84:57–78. https://doi.org/10.3897/mycokeys.84.71530
Hyde KD, Maharachchikumbura SSN, Hongsanan S, Samarakoon MC, Lücking R, Pem D, Harishchandra D, Jeewon R, Zhao RL, Xu JC, Liu JK, Al-Sadi AM, Bahkali AH, Elgorban AM (2017) The ranking of fungi: a tribute to David L. Hawksworth on his 70th birthday. Fungal Diversity 84:1–23. https://doi.org/10.1007/s13225-017-0383-3
Hyde KD, Norphanphoun C, Maharachchikumbura SSN, Bhat DJ, Jones EBG, Bundhun D, Chen YJ, Bao DF, Boonmee S, Calabon MS, Chaiwan N, Chethana KWT, Dai DQ, Dayarathne MC, Devadatha B, Dissanayake AJ, Dissanayake LS, Doilom M, Dong W, Fan XL, Goonasekara AJ, Dissanayake LS, Doilom M, Dong W, Fan XL, Goonasekara ID, Hongsanan S, Huang SK, Jayawardena RS, Jeewon R, Karunarathna A, Konta S, Kumar V, Lin CG, Liu JK, Liu NG, Luangsaard J, Lumyong S, Luo ZL, Marasinghe DS, McKenzie EHC, Niego AGT, Niranjan M, Perera RH, Phukhamsakda C, Rathnayaka AR, Samarakoon MC, Samarakoon SMBC, Sarma VV, Senanayake IC, Shang QJ, Stadler M, Tibpromma S, Wanasinghe DN, Wei DP, Wijayawardene NN, Xiao YP, Yang J, Zeng XY, Zhang SN, Xiang MM (2020) Refined families of Sordariomycetes. Mycosphere 11:305–1059. https://doi.org/10.5943/mycosphere/11/1/7
Hyde KD, Xu J, Rapior S, Jeewon R, Lumyong S, Niego AGT, Abeywickrama PD, Aluthmuhandiram JVS, Brahamanage RS, Brooks S, Hyde KD, Xu J, Rapior S, Jeewon R, Lumyong S, Niego AGT, Abeywickrama PD, Aluthmuhandiram JVS, Brahamanage RS, Brooks S, Chaiyasen A, Chethana KWT, Chomnunti P, Chepkirui C, Chuankid B, de Silva NI, Doilom M, Faulds C, Gentekaki E, Gopalan V, Kakumyan P, Harishchandra D, Hemachandran H, Hongsanan S, Karunarathna A, Karunarathna SC, Khan S, Kumla J, Jayawardena RS, Liu JK, Liu N, Luangharn T, Macabeo APG, Marasinghe DS, Meeks D, Mortimer PE, Mueller P, Nadir S, Nataraja KN, Nontachaiyapoom S, O’Brien M, Penkhrue W, Phukhamsakda C, Ramanan US, Rathnayaka AR, Sadaba RB, Sandargo B, Samarakoon BC, Tennakoon DS, Siva R, Sriprom W, Suryanarayanan TS, Sujarit K, Suwannarach N, Suwunwong T, Thongbai B, Thongklang N, Wei DP, Wijesinghe SN, Winiski J, Yan J, Yasanthika E, Stadler M (2019) The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Diversity 97:1–136. https://doi.org/10.1007/s13225-019-00430-9
Ignazio C, Linda MK (1999) A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 91:553–556. https://doi.org/10.1080/00275514.1999.12061051
Issakainen J, Jalava J, Hyvönen J, Sahlberg N, Pirnes T, Campbell CK (2003) Relationships of Scopulariopsis based on LSU rDNA sequences. Med Mycol 41:31–42
Iturrieta-González I, García D, Gené J (2021) Novel species of Cladosporium from environmental sources in Spain. MycoKeys 77:1–25. https://doi.org/10.3897/mycokeys.77.60862
Jayasiri SC, Hyde KD, Ariyawansa HA, Bhat J, Buyck B, Cai L, Dai YC, Abd-Elsalam KA, Ertz D, Hidayat I (2015) The Faces of Fungi database: fungal names linked with morphology, phylogeny and human impacts. Fungal Diversity 74:3–18. https://doi.org/10.1007/s13225-015-0351-8
Jo WS, Rew YH, Kim SH, Yun JT, Choi BS (1999) Occurrence of bluish green mold of Pleurotus eryngii by Penicillium corylophilum. Korean J Mycol 27:412–414
Ju YM, Hsieh HM, He XS (2010) Xylaria coprinicola, a new species that antagonizes cultivation of Coprinus comatus in China. Mycologia 103:424–430. https://doi.org/10.3852/10-215
Kageyama K (2014) Molecular taxonomy and its application to ecological studies of Pythium species. J Gen Plant Pathol 80:314–326. https://doi.org/10.1007/s10327-014-0526-2
Kamal S, Verma RN, Sharma HSS (2000) Role of lytic enzymes in yellow mould disease of Agaricus bisporus. Indian Phytopathol 53:273–278
Kamath PM, Prasad V, Shenoy VS, Mukundan A, Shenoy S (2015) Chrysosporium: an uncommon fungus in chronic rhinosinusitis. J Clin Diagn Res 9:MD01-2. https://doi.org/10.7860/JCDR/2015/11676.5688
Kang HJ, Sigler L, Lee J, Gibas CFC, Yun SH, Lee YW (2010) Xylogone ganodermophthora sp. nov., an ascomycetous pathogen causing yellow rot on cultivated mushroom Ganoderma lucidum in Korea. Mycologia 102:1167–1184. https://doi.org/10.3852/09-304
Karimi K, Zamani A (2013) Mucor indicus: biology and industrial application perspectives: a review. Biotechnol Adv 31:466–481. https://doi.org/10.1016/j.biotechadv.2013.01.009
Katoh K, Standley DM (2013) MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol 30:772–780. https://doi.org/10.1093/molbev/mst010
Kepler RM, Luangsa-Ard JJ, Hywel-Jones NL, Quandt CA, Sung GH, Rehner SA, Aime MC, Henkel TW, Sanjuan T, Zare R, Chen M, Li Z, Rossman AY, Spatafora JW, Shrestha B (2017) A phylogenetically-based nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 8:335–353. https://doi.org/10.5598/imafungus.2017.08.02.08
Kim CS, Park MS, Kim SC, Maekawa N, Yu SH (2012a) Identification of Trichoderma, a competitor of shiitake mushroom (Lentinula edodes), and competition between Lentinula edodes and Trichoderma species in Korea. Plant Pathol J 28:137–148. https://doi.org/10.5423/PPJ.OA.02.2011.0030
Kim CS, Yu SH, Nakagiri A, Shirouzu T, Sotome K, Kim SC, Maekawa N (2012b) Re-evaluation of Hypocrea pseudogelatinosa and H. pseudostraminea isolated from shiitake mushroom (Lentinula edodes) cultivation in Korea and Japan. Plant Pathol J 28:341–356. https://doi.org/10.5423/PPJ.OA.05.2012.0068
Kim HK, Seok SJ, Kim GP, Moon BJ, Terashita T (1999) Occurrence of disease caused by Cladobotryum varium on Flammulina velutipes in Korea. Korean J Mycol 27:415–419
Kim JY, Kwon HW, Tang L, Kim H (2012c) Identification and characterization of Trichoderma citrinoviride isolated from mushroom fly-infested oak log beds used for shiitake cultivation. Plant Pathol J 28:219–219. https://doi.org/10.5423/PPJ.DR.03.2011.006
Kim JY, Yun YH, Hyun MW, Kim MH, Kim SH (2010) Identification and characterization of Gliocladium viride isolated from mushroom fly infested oak log beds used for shiitake cultivation. Mycobiology 38:7–12. https://doi.org/10.4489/MYCO.2010.38.1.007
Kim MK, Lee YH, Cho KM, Lee JY (2012d) First report of cobweb disease caused by Cladobotryum mycophilum on the edible mushroom Pleurotus eryngii in Korea. Plant Dis 96:1374–1374. https://doi.org/10.1094/PDIS-01-12-0015-PDN
Kim MK, Seuk SW, Lee YH, Kim HR, Cho KM (2014) Fungicide sensitivity and characterization of cobweb disease on a Pleurotus eryngii mushroom crop caused by Cladobotryum mycophilum. Plant Pathol J 30:82–89. https://doi.org/10.5423/PPJ.OA.09.2013.0098
Kim SW, Kim S, Lee HJ, Park JW, Ro HS (2013) Isolation of fungal pathogens to an edible mushroom, Pleurotus eryngii, and development of specific ITS primers. Mycobiology 41:252–255. https://doi.org/10.5941/MYCO.2013.41.4.252
Kim T, Lee H, Song G, Shin W (1998) King oyster mushroom (Pleurotus eryngii) white mold disease caused by Cladobotryum varium. KSM News Lett 11:46
Kirk PM, Cannon PF, Minter DW, Stalpers JA (2008) Dictionary of the fungi, 10th edn. CABI, Wallingford, UK
Kirschner R, Arnold GR, Chen C (2007) Cladobotryum semicirculare sp. nov. (Hyphomycetes) from commercially grown Ganoderma tsugae in Taiwan and other Basidiomycota in Cuba. Sydowia-Horn 59:114–124
Komoń-Zelazowska M, Bissett J, Zafari D, Hatvani L, Manczinger L, Woo S, Lorito M, Kredics L, Kubicek CP, Druzhinina IS (2007) Genetically closely related but phenotypically divergent Trichoderma species cause green mold disease in oyster mushroom farms worldwide. Appl Environ Microbiol 73:7415–7426. https://doi.org/10.1128/AEM.01059-07
Kong HZ, Liang ZQ (2003) A new Penicillium species isolated from Jiangxi, China. Mycosystema 22:4–5
Kong XH, Liu JN, Zhang PQ, Dai XD, Han XD, Ma QF, Zhang JC (2011) The pathogens of white mildew disease of cultivated Auricularia auricula-judae in Northeast China. Mycosystema 30:551–555
Kosanović D, Potočnik I, Duduk B, Vukojević J, Stajić M, Rekanović E, Milijašević-Marčić S (2013) Trichoderma species on Agaricus bisporus farms in Serbia and their biocontrol. Ann Appl Biol 163:218–230. https://doi.org/10.1111/aab.1204
Kredics L, Hatvani L, Antal Z, Manczinger L, Druzgining IS, Kubicek CP, Szekeres A, Nagy A, Vágvölgyi C, Nagy E (2006) Green mould disease of oyster mushroom in Hungary and Transylvania. Acta Microbiol Immunolog Hung 53:306–307
Krijgsheld P, Bleichrodt R, van Veluw GJ, Wang F, Müller WH, Dijksterhuis J, Wösten HA (2013) Development in Aspergillus. Stud Mycol 74:1–29. https://doi.org/10.3114/sim0006
Kunze G, Schmidt JC (1817) Mykologische Hefte. 1:1–109
Kwon H, Yun Y, Kim J, Kim S, Ko H (2015) New records of fungi isolated from indoor air of greenhouse used for shiitake cultivation in Korea. Korean J Mycol 43:58–63. https://doi.org/10.4489/KJM.2015.43.1.58
Lan YF, Cong QQ, Cui X (2019) Cladobotryum hypzigus, newly reported pathogen causing Cobweb disease on Beech Mushroom and comparison of Cladobotryum spp. in Korea. Edible Fungi China 38:76–79
Lan YF, Cong QQ, Wang QW, Tang LN, Li XM, Yu QW, Cui X, An XR, Yu CX, Kong FH, Li XD (2020) First report of Cladobotryum protrusum causing cobweb disease on cultivated Morchella importuna. Plant Dis 104:977. https://doi.org/10.1094/PDIS-08-19-1611-PDN
Lan YF, Wang QW, Yu CX, Cong QQ, Tang LN, An XR, Kong FH, Li XD (2016) First report of Cladobotryum mycophilum causing cobweb disease on Agaricus bisporus in China. Plant Dis 100:2334–2334. https://doi.org/10.1094/PDIS-05-16-0754-PDN
Largeteau ML, Savoie JM (2010) Microbially induced diseases of Agaricus bisporus: biochemical mechanisms and impact on commercial mushroom production. Appl Microbiol Biotechnol 86:63–73. https://doi.org/10.1007/s00253-010-2445-2
Lee JK, Choi GJ, Cho KY, Oh SJ, Park JS (1996) Xylogone sphaerospora, a new fungal pathogen of cultivated Ganoderma lucidum. Korean J Mycol 24:246–254
Lee SY, Back CG, Park JH, Kang IK, Jung HY (2015) D-8: Cladobotryum hypzigus, newly reported pathogen causing Cobweb disease on Beech Mushroom and Comparison of Cladobotryum spp. in Korea. 균학회소식 27:117.
Li D, Sossah FL, Sun L, Fu Y, Li Y (2019) Genome analysis of Hypomyces perniciosus, the causal agent of wet bubble disease of button mushroom (Agaricus bisporus). Genes 10:417. https://doi.org/10.3390/genes10060417
Link HF (1809) Observationes in ordines plantarum naturales. Dissertatio I. Mag Ges Naturf Freunde Berlin 3:3–42
Link HF (1809) Observationes in ordines plantarum naturales. Dissertatio I. Magazin der Gesellschaft Naturforschenden Freunde Berlin 3:3–42.
Liu K, Zheng Q, Jiang J, Wu Z (2019) Isolation and identification of Xylogone ganodermophthora, a new record to China. J Fungal Res 17:74–78
Liu Q, Wang F, Xu FX, Xu YY, Dong CH (2021a) Pathogenic fungi of artificially cultivated Cordyceps militaris. Mycosystema 40:2962–2980
Liu TH, Zhou J, Wang D, He XL, Tang J, Chen Y, Wang J, Peng WH (2021b) A new stipe rot disease of the cultivated Morchella sextelata. Mycosystema 40:2229–2243
Liu W, Zhang Y, He PX (2017) Morel biology and cultivation. Jilin Science and Technology Press, Changchun
Liu YJ, Whelen S, Hall BD (2000) Phylogenetic relationships among ascomycetes: evidence from an RNA Polymerse II subunit. Mol Biol Evol 16:1799–1808. https://doi.org/10.1093/oxfordjournals.molbev.a026092
Liu ZH, Cong YL, Sossah FL, Lu YZ, Kang JC, Li Y (2023) Characterization and Genome Analysis of Cladobotryum mycophilum, the Causal Agent of Cobweb Disease of Morchella sextelata in China. J Fungi 9:411. https://doi.org/10.3390/jof9040411
Liu ZY, Liang ZQ, Liu AY (1996) A new species Paecilomyces isolated from Cordyceps odonatae. Mycosystema 9:83–87
Lu ML, Wen TC, Guo M, Li QH, Peng XC, Zhang Y, Lu ZH, Wang J, Xu YJ, Zhang C (2023) Regulation of Intracellular Reactive Oxygen Species Levels after the Development of Phallus rubrovolvatus Rot Disease Due to Trichoderma koningii Mycoparasitism. J Fungi 9:525. https://doi.org/10.3390/jof9050525
Luangsa-Ard J, Houbraken J, van Doorn T, Hong SB, Borman AM, Hywel-Jones NL, Samson RA (2011) Purpureocillium, a new genus for the medically important Paecilomyces lilacinus. FEMS Microbiol Lett 321:141–149. https://doi.org/10.1111/j.1574-6968.2011.02322.x
Luangsa-Ard JJ, Hywel-Jones NL, Manoch L, Samson RA (2005) On the relationships of Paecilomyces sect. Isarioidea species. Mycol Res 109:581–589. https://doi.org/10.1017/S0953756205002741
Luangsa-Ard JJ, Hywel-Jones NL, Samson RA (2004) The polyphyletic nature of Paecilomyces sensu lato based on 18S-generated rDNA phylogeny. Mycologia 96:773–780. https://doi.org/10.2307/3762111
Luo J, Zhuang WY (2008) Two new species of Cosmospora (Nectriaceae, Hypocreales) from China. Fungal Divers 31:83–93
Lv B, Yu S, Chen Y, Yu H, Mo Q (2022) First report of Lecanicillium aphanocladii causing rot of Morchella sextelata in China. Plant Dis 106:3202. https://doi.org/10.1094/PDIS-12-21-2656-PDN
Maharachchikumbura SSN, Chen YP, Ariyawansa HA, Hyde KD, Haelewaters D, Perera RH, Samarakoon MC, Wanasinghe DN, Bustamante DE, Liu JK, Lawrence DP, Cheewangkoon R, Stadler M (2021) Integrative approaches for species delimitation in Ascomycota. Fungal Diversity 109:155–179. https://doi.org/10.1007/s13225-021-00486-6
Maharachchikumbura SSN, Hyde KD, Jones EBG, McKenzie EHC, Bhat JD, Dayarathne MC, Huang SK, Xiao YP, D’souza MJ, Hongsanan S, Jayawardena RS, Daranagama DA, Konta K, Goonasekara ID, Zhuang WY, Jeewon R, Phillips AJL, Abdel-Wahab MA, Al-Sadi AM, Dissanayake AJ, Kang JC, Li QR, Liu JK, Liu XZ, Liu ZY, Luangsaard JJ, Pang KL, Phookamsak R, Promputtha I, Suetrong S, Stadler M, Wen TC, Wijayawardene NN (2016) Families of Sordariomycetes. Fungal Divers 79:1–317. https://doi.org/10.1007/s13225-016-0369-6
Maharachchikumbura SSN, Hyde KD, Jones EBG, McKenzie EHC, Huang SK, Abdel-Wahab MA, Daranagama DA, Dayarathne M, D’souza MJ, Goonasekara ID, Hongsanan S, Jayawardena RS, Kirk PM, Konta S, Liu J-K, Liu ZY, Norphanphoun C, Pang KL, Perera R, Senanayake IC, Shang QJ, Shenoy BD, Xiao YP, Bahkali A, Kang JC, Somrothipol S, Suetrong S, Wen TC, Xu JC (2015) Towards a natural classification and backbone tree for Sordariomycetes. Fungal Divers 72:1–103. https://doi.org/10.1007/s13225-015-0331-z
Mapook A, Hyde KD, Hassan K, Hassan K, Kemkuignou BM, Čmoková A, Surup F, Kuhnert E, Paomephan P, Cheng T, de Hoog S, Song YG, Jayawardena RS, Al-Hatmi AMS, Mahmoudi T, Ponts N, Studt-Reinhold L, Richard-Forget F, Chethana KWT, Harishchandra DL, Mortimer PE, Li HL, Lumyong S, Aiduang W, Kumla J, Suwannarach N, Bhunjun CS, Yu FM, Zhao Q, Schaefer D, Marc Stadler M (2022) Ten decadal advances in fungal biology leading towards human well-being. Fungal Diversity 116:547–614. https://doi.org/10.1007/s13225-022-00510-3
Marlowe A, Romaine CP (1982) Dry bubble of oyster mushroom caused by Verticillium fungicola. Plant Dis 66:859–860
Masaphy S (2022) First report on Purpureocillium lilacinum infection of indoor-cultivated morel primordia. Agriculture 12:695. https://doi.org/10.3390/agriculture12050695
Matsushima T (1976) Icones Microfungorum a Matsushima Lectorum. Mycologia 68:955. https://doi.org/10.2307/3758819
Matsushima T (1981) Matsushima Mycological Memoirs No. 2. Matsushima Mycol Memoirs 2:1–68
McKay GJ, Egan D, Morris E, Scott C, Brown AE (1999) Genetic and morphological characterization of Cladobotryum species causing cobweb disease of mushrooms. Appl Environ Microbiol 65:606–610. https://doi.org/10.1128/AEM.65.2.606-610.1999
Melo IS, Faull JL, Nascimento RS (2006) Antagonism of Aspergillus terreus to Sclerotinia sclerotiorum. Braz J Microbiol 37:417–499
Miles PG, Chang ST (2004) Mushrooms: cultivation, nutritional value, medicinal effect, and environmental impact. CRC Press, Boca Raton
Moreira DC, Oliveira MME, Borba CM (2018) Human pathogenic paecilomyces from food. Microorganisms 6:64
Moreno-Gavíra A, Huertas V, Diánez F, Sánchez-Montesinos B, Santos M (2020) Paecilomyces and its importance in the biological control of agricultural pests and diseases. Plants 9:1746
Murmu R, Lal AA, Kumar B, Singh S, Simon S (2014) Incidence of Papulaspora byssina (brown plaster mould) on casing mixture and compost of white button mushroom (Agaricus bisporus). Bioscan 9:449–451
Naik J, Pani BK (2019) Unwanted microbial invasion and first report of two new disease encountered during paddy straw mushroom cultivation in Odisha. Int J Curr Agric Sci 9:430–433
Nair NG, Letham DB, Walker J (1980) Incidence of Aphanocladium album on the cultivated mushroom, Agaricus bisporus, in New South Wales 20–22.
Nguyen TTT, Hee Lee S, Jeong Jeon S, Burm Lee H (2019) First Records of Rare Ascomycete Fungi, Acrostalagmus luteoalbus, Bartalinia robillardoides, and Collariella carteri from Freshwater Samples in Korea. Mycobiology 47:1–11. https://doi.org/10.1080/12298093.2018.1550894
Nie Y, Cai Y, Gao Y, Yu DS, Wang ZM, Liu XY, Huang B (2020) Three new species of Conidiobolus sensu stricto from plant debris in eastern China. MycoKeys 73:133–149. https://doi.org/10.3897/mycokeys.73.56905
Niego AGT, Lambert C, Mortimer P, Thongklang N, Rapior S, Grosse M, Schrey H, Charria-Girón E, Walker A, Hyde KD, Stadler M (2023) The contribution of fungi to the global economy. Fungal Diversity. https://doi.org/10.1007/s13225-023-00520-9
Nirenberg HI, Hagedorn G (2008) Fusarium nematophilum spec. nov—.ein neuer Nematoden-assoziierter Pilz. Nachrichtenblatt Des Deutschen Pflanzenschutzdienstes 60:213–216
Norphanphoun C, Gentekaki E, Hongsana S, Jayawardena R, Senanayake IC, Manawasinghe IS, Abeywickrama PD, Bhunjun CS, Hyde KD (2022) Diaporthe: Formalizing the species-group concept. Mycosphere 13:752–819. https://doi.org/10.5943/mycosphere/13/1/9
O’Donnell K, Cigelnik E, Benny GL (1998) Phylogenetic relationships among the Harpellales and Kickxellales. Mycologia 90:624–639. https://doi.org/10.1080/00275514.1998.12026952
O’Donnell K, Cigelnik E, Weber NS, Trappe JM (1997) Phylogenetic relationships among ascomycetous truffles and the true and false morels inferred from 18S and 28S ribosomal DNA sequence analysis. Mycologia 89:48–65. https://doi.org/10.1080/00275514.1997.12026754
Oh SJ, Chun CS, Lee JK, Kim HK (1998) Occurrence and identification of the fungus causing yellow rot on Ganoderma lucidum. Korean J Mycol 26:31–38
Ohm RA, De Jong JF, Lugones LG, Aerts A, Kothe E, Stajich JE, De Vries RP, Record E, Levasseur A, Baker SE (2010) Genome sequence of the model mushroom Schizophyllum commune. Nat Biotechnol 28:957–963. https://doi.org/10.1038/nbt.1643
Okuda Y, Nagasawa E, Tokiwa T, Hase K, Murakami S (2016) Cottony leak on cultivated Auricularia nigricans caused by Hypomyces pseudocorticiicola. Rep Tottori Mycol Inst 46:23–29
Ozimek E, Hanaka A (2021) Mortierella species as the plant growth-promoting fungi present in the agricultural soils. Agriculture 11:7. https://doi.org/10.3390/agriculture11010007
Pažoutová S, Follert S, Bitzer J, Keck M, Surup F, Šrůtka P, Holuša J, Stadler M (2013) A new endophytic insect-associated Daldinia species, recognised from a comparison of secondary metabolite profiles and molecular phylogeny. Fungal Diversity 60:107–123. https://doi.org/10.1007/s13225-013-0238-5
Perera RH, Hyde KD, Jones EBG, Maharachchikumbura SSN, Bundhun D, Camporesi E, Akulov A, Liu JK, Liu ZY (2023) Profile of Bionectriaceae, Calcarisporiaceae, Hypocreaceae, Nectriaceae, Tilachlidiaceae, Ijuhyaceae fam. nov., Stromatonectriaceae fam. nov. and Xanthonectriaceae fam. nov. Fungal Divers 118:5–271. https://doi.org/10.1007/s13225-022-00512-1
Põldmaa K (2000) Generic delimitation of the fungicolous Hypocreaceae. Stud Mycol 45:83–94
Põldmaa K (2011) Tropical species of Cladobotryum and Hypomyces producing red pigments. Stud Mycol 68:1–34. https://doi.org/10.3114/sim.2011.68.01
Park MS, Bae KS, Yu SH (2006) Two new species of Trichoderma associated with green mold of oyster mushroom cultivation in Korea. Mycobiology 34:111–113
Park MS, Oh SY, Fong JJ, Houbraken J, Lim YW (2019) The diversity and ecological roles of Penicillium in intertidal zones. Sci Rep 9:1–11. https://doi.org/10.1038/s41598-019-49966-5
Parmar HV, Sodavadiya HB (2019) Diseases of mushroom and their management. Agric Food 21530:151
Peng W, He X, Wang Y, Zhang Y, Ye X, Jia D, Guo Y, Gan B, Zheng C, Yang Z, Sun Q (2014) A new species of Scytalidium causing slippery scar on cultivated Auricularia polytricha in China. FEMS Microbiol Lett 359:72–80. https://doi.org/10.1111/1574-6968.12564
Persoon CH (1794) Neuer Versuch einer systematischen Eintheilung der Schwämme. Neues Magazin für die Botanik in ihrem ganzen Umfange 1:63–80.
Pesante A (1957) Osservazioni su una carie del platana. Ann Sperimentazione Agaria 11(1):249–266
Peterson SW, Bayer EM, Wicklow DT (2004) Penicillium thiersii, Penicillium angulare and Penicillium decutarense, new species isolated from wood-decay fungi in North America and their phylogenetic placement from multilocus DNA sequences. Mycologia 96:1280–1293
Posada D (2008) jModelTest: phylogenetic model averaging. Mol Biol Evol 25:1253–1256. https://doi.org/10.1093/molbev/msn083
Potocnik I, Rekanovic E, Milijasevic S, Todorovic B, Stepanović M (2015) Morphological and pathogenic characteristics of the fungus Cladobotryum dendroides, the causal agent of cobweb disease of the cultivated mushroom Agaricus bisporus in Serbia. Pesticidi i Fitomedicina 23:175–181
Preuss CGT (1851) Uebersicht untersuchter Pilze, besonders aus der Umgegend von Hoyerswerda. Linnaea 24:99–153
Pukahuta C, Limtong S, Suwanarit P, Nutalaya S (2000) Species diversity of Trichoderma contaminating shiitake production houses in Thailand. Agric Nat Resour 34:478–485
Qiao M, Zhang Y, Li SF, Baral HO, Weber E, Su HY, Xu JP, Zhang KQ, Yu ZF (2012) Orbilia blumenaviensis and its Arthrobotrys anamorph. Mycol Progress 11:255–262. https://doi.org/10.1007/s11557-011-0744-3
Qin L, Qiao M, Yang Y, Yang GZ, Lu KP, Zhang K-Q, Xu JP, Yu ZF (2011) Dactylella pseudobrevistipitata, a new species from China. Ann Microbiol 61:591–595. https://doi.org/10.1007/s13213-010-0177-2
Quandt CA, Di Y, Elser J, Jaiswal P, Spatafora JW (2016) Differential expression of genes involved in host recognition, attachment, and degradation in the mycoparasite Tolypocladium ophioglossoides. G3 6:731–741. https://doi.org/10.1534/g3.116.027045
Rambaut A, Suchard MA, Xie D, Drummond AJ (2013) Tracer version 1.6. University of Edinburgh. [Online]. http://tree.bio.ed.ac.uk/software/tracer
Raza M, Zhang ZF, Hyde KD, Diao YZ, Cai L (2019) Culturable plant pathogenic fungi associated with sugarcane in southern China. Fungal Diversity 99:1–104. https://doi.org/10.1007/s13225-019-00434-5
Rice AV, Currah RS (2005a) Oidiodendron: a survey of the named species and related anamorphs of Myxotrichum. Stud Mycol 53:83–120. https://doi.org/10.3114/sim.53.1.83
Rice AV, Currah RS (2005b) Profiles from Biolog FF plates and morphological characteristics support the recognition of Oidiodendron fimicola sp. nov. Stud Mycol 53:75–82. https://doi.org/10.3114/sim.53.1.75
Robak H (1932) Investigations regarding fungi on Norwegian ground wood pulp and fungal infection at wood pulp mills. Nytt Mag Nat 71:185–330
Rogerson CT, Samuels GJ (1989) Boleticolous species of Hypomyces. Mycologia 81:413–432
Rogerson CT, Samuels GJ (1993) Polyporicolous species of Hypomyces. Mycologia 85:213–272
Rogerson CT, Samuels GJ (1994) Agaricicolous species of Hypomyces. Mycologia 86:839–866
Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Höhna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP (2012) MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol 61:39–42. https://doi.org/10.1093/sysbio/sys029
Ropars J, Cruaud C, Lacoste S, Dupont J (2012) A taxonomic and ecological overview of cheese fungi. Int J Food Microbiol 155:199–210
Rossman AY, Samuels GJ, Rogerson CT, Lowen R (1999) Genera of Bionectriaceae, Hypocreaceae and Nectriaceae (Hypocreales, Ascomycetes). Stud Mycol 42:1–248
Rossman AY (1983) The phragmosporous species of Nectria and related genera. Mycol Pap 150:1–164
Rossman AY (2014) Lessons learned from moving to one scientific name for fungi. IMA Fungus 5:81–89. https://doi.org/10.5598/imafungus.2014.05.01.10
Salvatore MM, Andolfi A, Nicoletti R (2021) The genus Cladosporium: a rich source of diverse and bioactive natural compounds. Molecules 26:3959. https://doi.org/10.3390/molecules26133959
Samarakoon MC, Hyde KD, Maharachchikumbura SS, Stadler M, Gareth Jones E, Promputtha I, Suwannarach N, Camporesi E, Bulgakov TS, Liu JK (2022) Taxonomy, phylogeny, molecular dating and ancestral state reconstruction of Xylariomycetidae (Sordariomycetes). Fungal Diversity 112:1–88. https://doi.org/10.1007/s13225-021-00495-5
Samarakoon MC, Hyde KD, Promputtha I, Hongsanan S, Ariyawansa H, Maharachchikumbura S, Daranagama D, Stadler M, Mapook A (2016) Evolution of Xylariomycetidae (Ascomycota: Sordariomycetes). Mycosphere 7:1746–1761. https://doi.org/10.5943/mycosphere/7/11/9
Samson RA (1974) Paecilomyces and some allied hyphomycetes. Stud Mycol 6:1–119
Samson RA, Visagie CM, Houbraken J, Hong SB, Hubka V, Klaassen CH, Perrone G, Seifert KA, Susca A, Tanney JB, Varga J, Kocsubé S, Szigeti G, Yaguchi T, Frisvad JC (2014) Phylogeny, identification and nomenclature of the genus Aspergillus. Stud Mycol 78:141–173. https://doi.org/10.1016/j.simyco.2014.07.004
Samuels GJ, Dodd SL, Gams W, Castlebury LA, Petrini O (2002) Trichoderma species associated with the green mold epidemic of commercially grown Agaricus bisporus. Mycologia 94:146–170. https://doi.org/10.1080/15572536.2003.11833257
Sandoval-Denis M, Sutton DA, Fothergill AW, Cano-Lira J, Gené J, Decock CA, de Hoog GS, Guarro J (2013) Scopulariopsis, a poorly known opportunistic fungus: spectrum of species in clinical samples and in vitro responses to antifungal drugs. J Clin Microbiol 51:3937–3943. https://doi.org/10.1128/JCM.01927-13
Sandoval-Denis M, Gené J, Sutton DA, Cano-Lira JF, De Hoog GS, Decock CA, Wiederhold NP, Guarro J (2016) Redefining Microascus, Scopulariopsis and allied genera. Persoonia 36:1–36. https://doi.org/10.3767/003158516X688027
Sang WJ, Song BA, Lian QX, Li XX, Hu DY, Huang RM (2006) Pathogen identification for the rot of Dictyophra rubrovolvata and selection of fungicdes for its conitrol. J Southwest Agric Univ (Nat Sci) 28:302–304
Schoch CL, Robbertse B, Robert V, Vu D, Cardinali G, Irinyi L, Meyer W, Nilsson RH, Hughes K, Miller AN, Kirk PM, Abarenkov K, Aime MC, Ariyawansa HA, Bidartondo M, Boekhout T, Buyck B, Cai Q, Chen J, Crespo A, Crous PW, Damm U, De Beer ZW, Dentinger BTM, Divakar PK, Dueñas M, Feau N, Fliegerova K, García MA, Ge ZW, Griffith GW, Groenewald JZ, Groenewald M, Grube M, Gryzenhout M, Gueidan C, Guo L, Hambleton S, Hamelin R, Hansen K, Hofstetter V, Hong SB, Houbraken J, Hyde KD, Inderbitzin P, Johnston PR, Karunarathna SC, Kõljalg U, Kovács GM, Kraichak E, Krizsan K, Kurtzman CP, Larsson KH, Leavitt S, Letcher PM, Liimatainen K, Liu JK, Lodge DJ, Jennifer Luangsa-ard J, Lumbsch HT, Maharachchikumbura SSN, Manamgoda D, Martín MP, Minnis AM, Moncalvo JM, Mulè G, Nakasone KK, Niskanen T, Olariaga I, Papp T, Petkovits T, Pino-Bodas R, Powell MJ, Raja HA, Redecker D, Sarmiento-Ramirez JM, Seifert KA, Shrestha B, Stenroos S, Stielow B, Suh SO, Tanaka K, Tedersoo L, Telleria MT, Udayanga D, Untereiner WA, Diéguez Uribeondo J, Subbarao KV, Vágvölgyi C, Visagie C, Voigt K, Walker DM, Weir BS, Weiß M, Wijayawardene NN, Wingfield MJ, Xu JP, Yang ZL, Zhang N, Zhuang WY, Federhen S (2014) Finding needles in haystacks: linking scientific names, reference specimens and molecular data for Fungi. Database. https://doi.org/10.1093/database/bau061
Schroers HJ, Gräfenhan T, Nirenberg HI, Seifert KA (2011) A revision of Cyanonectria and Geejayessis gen. nov., and related species with fusarium-like anamorphs. Stud Mycol 68:115–138. https://doi.org/10.3114/sim.2011.68.05
Schroers H, Samuels GJ, Seifert KA, Gams W (1999) Classification of the mycoparasite Gliocladium roseum in Clonostachys as C. rosea, its relationship to Bionectria ochroleuca, and notes on other Gliocladium-like fungi. Mycologia 91:365–385
Schroers HJ (2001) A monograph of Bionectria (Ascomycota, Hypocreales, Bionectriaceae) and its Clonostachys anamorphs. Stud Mycol 46:1–214
Seaby DA (1996) Investigation of the epidemiology of green mould of mushroom (Agaricus bisporus) compost caused by Trichoderma harzianum. Plant Pathol 45:913–923
Seifert K (2008) Compendium of soil fungi-by Domsch KH, Gams W, Anderson TH. Eur J Soil Sci 59:1007–1007. https://doi.org/10.1111/1.1365-2389.2008.01052.x
Senanayake IC, Rathnayaka AR, Sandamali DS, Calabon MS, Gentekaki E, Lee HB, Hurdeal VG, Pem D, Dissanayake LS, Wijesinghe SN, Bundhun D, Nguyen TT, Goonasekara ID, Abeywickrama PD, Bhunjun CS, Jayawardena RS, Wanasinghe DN, Jeewon R, Bhat DJ, Xiang MM (2020) Morphological approaches in studying fungi: collection, examination, isolation, sporulation and preservation. Mycosphere 11:2678–2754. https://doi.org/10.5943/mycosphere/11/1/20
Senthilkumar M, Anandham R, Krishnamoorthy R (2020) Paecilomyces. In: Amaresan N, SenthilKumar M, Annapurna K, Kumar K, Sankaranarayanan A (eds) Beneficial microbes in agro-ecology. Academic Press, Cambridge, pp 793–808
Sharma VP, Kumar S, Mahajan S (2006) Yield loss and management of Oedocephalum cultivation of Calocybe indica. Ann Plant Prot Sci 14:306–309
Sharma VP, Sharma S, Kumar S, Gupta M, Kamal S (2016) Cob web and dry bubble diseases in Lentinula edodes cultivation-a new report. International Society for Mushroom Science 130–134.
Shi XF, Liu D, He XH, Liu W, Yu F (2022) Epidemic Identification of Fungal Diseases in Morchella Cultivation across China. Journal of Fungi 8:1107. https://doi.org/10.3390/jof8101107
Shigehiro K, Christian MZ, Osamu N, Kazutaka K (2013) aLeaves facilitates on-demand exploration of metazoan gene family trees on MAFFT sequence alignment server with enhanced interactivity. Nucleic Acids Res 41:W22–W28. https://doi.org/10.1093/nar/gkt389
Sigler L, Carmichael JW (1976) Taxonomy of Malbranchea and some other Hyphomycetes with arthroconidia. Mycotaxon 4:349–488
Sigler L, Flis A, Carmichael J (2011) The genus Uncinocarpus (Onygenaceae) and its synonym Brunneospora: new concepts, combinations and connections to anamorphs in Chrysosporium, and further evidence of relationship with Coccidioides immitis. Can J Bot 76:1624–1636. https://doi.org/10.1139/b98-110
Sinha KK, Choudhary AK, Kumari P (2016) Entomopathogenic fungi. In: Omkar BT (ed) Ecofriendly pest management for food security. Academic Press, Cambridge, pp 475–505
Siqueira JP, Sutton D, Gené J, García D, Guevara-Suarez M, Decock C, Wiederhold N, Guarro J (2016) Schizophyllum radiatum, an Emerging Fungus from Human Respiratory Tract. J Clin Microbiol 54:2491–2497. https://doi.org/10.1128/JCM.01170-16
Sossah FL, Liu Z, Yang C, Okorley BA, Sun L, Fu Y, Li Y (2019) Genome sequencing of Cladobotryum protrusum provides insights into the evolution and pathogenic mechanisms of the cobweb disease pathogen on cultivated mushroom. Genes 10:124. https://doi.org/10.3390/genes10020124
Srinivasan MC, Thirumalachar M (1965) Studies on species of Conidiobolus from India. Sydowia 19:86–91
Stephen AR, Ellen B (2005) A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 97:84–98. https://doi.org/10.3852/mycologia.97.1.84
Sukarno N, Kurihara Y, Park JY, Inaba S, Ando K, Harayama S, Ilyas M, Mangunwardoyo W, Sjamsuridzal W, Yuniarti E, Saraswati R (2009) Lecanicillium and Verticillium species from Indonesia and Japan including three new species. Mycoscience 50:369–379
Summerbell RC, Gueidan C, Schroers HJ, de Hoog GS, Starink M, Rosete YA, Guarro J, Scott JA (2011) Acremonium phylogenetic overview and revision of Gliomastix, Sarocladium, and Trichothecium. Stud Mycol 68:139–162. https://doi.org/10.3114/sim.2011.68.06
Sun J, Bian Y (2012) Slippery scar: a new mushroom disease in Auricularia polytricha. Mycobiology 40:129–133
Sun JZ, Liu XZ, Hyde KD, Zhao Q, Maharachchikumbura SS, Camporesi E, Bhat J, Nilthong S, Lumyong S (2017) Calcarisporium xylariicola sp. nov. and introduction of Calcarisporiaceae fam. nov. in Hypocreales. Mycol Prog 16:433–445. https://doi.org/10.1007/s11557-017-1290-4
Sun JZ, Liu XZ, McKenzie EH, Jeewon R, Liu JK, Zhang XL, Zhao Q, Hyde KD (2019a) Fungicolous fungi: terminology, diversity, distribution, evolution, and species checklist. Fungal Diversity 95:337–430. https://doi.org/10.1007/s13225-019-00422-9
Sun JZ, Liu XZ, Jeewon R, Li YL, Lin CG, Tian Q, Zhao Q, Xiao XP, Hyde KD, Nilthong S (2019b) Fifteen fungicolous Ascomycetes on edible and medicinal mushrooms in China and Thailand. Asian J Mycol 2:129–169. https://doi.org/10.5943/ajom/2/1/7
Sun JZ, Yu S, Lu YZ, Liu HW, Liu XZ (2022) Proposal of a new family Pseudodiploösporeaceae fam. nov. (Hypocreales) based on phylogeny of Diploöspora longispora and Paecilomyces penicillatus. Mycology 14:60–73. https://doi.org/10.1080/21501203.2022.2143919
Sun W, Fan J, Fang A, Li Y, Tariqjaveed M, Li D, Hu D, Wang WM (2020) Ustilaginoidea virens: insights into an emerging rice pathogen. Annu Rev Phytopathol 58:363–385. https://doi.org/10.1146/annurev-phyto-010820-012908
Sung GH, Poinar GO Jr, Spatafora JW (2008) The oldest fossil evidence of animal parasitism by fungi supports a Cretaceous diversification of fungal-arthropod symbioses. Mol Phylogenet Evol 49:495–502. https://doi.org/10.1016/j.ympev.2008.08.028
Talavera-Ortiz A, Chaverri P, Díaz-Godínez G, de Lourdes Acosta-Urdapilleta M, Villegas E, Téllez-Téllez M (2019) Mycelial inhibition of Trichoderma spp. (Hypocreaceae) isolated from the cultivation of Pleurotus ostreatus (Pleurotaceae) with an extract of Pycnoporus sp. (Polyporaceae). Acta Bot Mex. https://doi.org/10.21829/abm127.2020.1537
Tamm H, Põldmaa K (2013) Diversity, host associations, and phylogeography of temperate aurofusarin-producing Hypomyces/Cladobotryum including causal agents of cobweb disease of cultivated mushrooms. Fungal Biol 117:348–367. https://doi.org/10.1016/j.funbio.2013.03.005
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30:2725–2729. https://doi.org/10.1093/molbev/mst197
Tanabe Y, Nagahama T, Saikawa M, Sugiyama J (1999) Phylogenetic relationship of Cephaliophora to nematophagous hyphomycetes including taxonomic and nomenclatural emendations of the genus Lecophagus. Mycologia 91:830–835
Tanney JB, Nguyen HD, Pinzari F, Seifert KA (2015) A century later: rediscovery, culturing and phylogenetic analysis of Diploöspora rosea, a rare onygenalean hyphomycete. Antonie Van Leeuwenhoek 108:1023–1035. https://doi.org/10.1007/s10482-015-0555-7
Tautorus TE, Townsley PM (1983) Biological control of olive green mold in Agaricus bisporus cultivation. Appl Environ Microbiol 45:511–515
Thaxter R (1903) New or peculiar North American Hyphomycetes III. Bot Gazette Crawfordsville 35:153–159
Torcato C, Gonçalves MFM, Rodríguez-Gálvez E, Alves A (2020) Clonostachys viticola sp. nov., a novel species isolated from Vitis vinifera. Int J Syst Evol Microbiol 70:4321–4328. https://doi.org/10.1099/ijsem.0.004286
Tsuneda A, Murakami S, Gill WM, Maekawa N (1997) Black spot disease of Lentinula edodes caused by the Hyphozyma synanamorph of Eleutheromyces subulatus. Mycologia 89:867–875
Udayanga D, Liu X, McKenzie EH, Chukeatirote E, Bahkali AH, Hyde KD (2011) The genus Phomopsis: biology, applications, species concepts and names of common phytopathogens. Fungal Diversity 50:189–225
Valero-Jiménez CA, Veloso J, Staats M, van Kan JAL (2019) Comparative genomics of plant pathogenic Botrytis species with distinct host specificity. BMC Genomics 20:203. https://doi.org/10.1186/s12864-019-5580-x
Van Greuning M, Eicker A (1991) The identity of the lipstick mould of cultivated mushrooms, Agaricus bisporus. Bot Bull Acad Sin 32:175–181
Van Greuning M, Ficker A (1992) The morphology and physiology of Arthrobotrys oligospora, a weed mould on mushroom beds of Agaricus bisporus. Mushroom Res 1:33–38
Van Oorschot CA (1980) A revision of Chrysosporium and allied genera. Stud Mycol 20:1–89
Vilgalys R, Hester M (1990) Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cyptococcus species. J Bacteriol 172:4238–4246. https://doi.org/10.1128/jb.172.8.4238-4246.1990
Visagie CM, Houbraken J, Frisvad JC, Hong SB, Klaassen CH, Perrone G, Seifert KA, Varga J, Yaguchi T, Samson RA (2014) Identification and nomenclature of the genus Penicillium. Stud Mycol 78:343–371. https://doi.org/10.1016/j.simyco.2014.09.001
Vu D, Groenewald M, De Vries M, Gehrmann T, Stielow B, Eberhardt U, Al-Hatmi A, Groenewald JZ, Cardinali G, Houbraken J, Boekhout T, Crous PW, Robert V, Verkley GJM (2019) Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud Mycol 92:135–154. https://doi.org/10.1016/j.simyco.2018.05.001
Walker JD (2019) GSA Geologic Time Scale v. 5.0. Elsevier, Amsterdam
Wang GZ, Guo MP, Bian YB (2015a) First report of Cladobotryum protrusum causing cobweb disease on the edible mushroom Coprinus comatus. Plant Dis 99:287–287. https://doi.org/10.1094/PDIS-07-14-0757-PDN
Wang QM, Yurkov AM, Göker M, Lumbsch HT, Leavitt SD, Groenewald M, Theelen B, Liu XZ, Boekhout T, Bai FY (2015b) Phylogenetic classification of yeasts and related taxa within Pucciniomycotina. Stud Mycol 81:149–189
Wang GZ, Cao XT, Ma XL, Guo MP, Liu CH, Yan LI, Bian YB (2016a) Diversity and effect of Trichoderma spp. associated with green mold disease on Lentinula edodes in China. Microbiologyopen 5:709–718. https://doi.org/10.1002/mbo3.364
Wang XH, Huhtinen S, Hansen K (2016b) Multilocus phylogenetic and coalescent-based methods reveal dilemma in generic limits, cryptic species, and a prevalent intercontinental disjunct distribution in Geopyxis (Pyronemataceae sl, Pezizomycetes). Mycologia 108:1189–1215. https://doi.org/10.3852/16-100
Wang XW, Houbraken J, Groenewald JZ, Meijer M, Andersen B, Nielsen KF, Crous PW, Samson RA (2016c) Diversity and taxonomy of Chaetomium and chaetomium-like fungi from indoor environments. Stud Mycol 84:145–224. https://doi.org/10.1016/10.1016/j.simyco.2016.11.005
Wang Q, Long YY, Li DP, Chen TS (2017) Investigation of fungal diseases and identification of main pathogens on Tricholoma giganteum. Edible Fungi 5:71–74 ((In Chinese))
Wang GZ, Ma CJ, Zhou SS, Põldmaa K, Tamm H, Luo Y, Guo MP, Ma XL, Bian YB, Zhou Y (2018) First Report of cobweb disease of Auricularia polytricha (Mont.) Sacc. caused by Cladobotryum cubitense in Xuzhou, China. Plant Dis. https://doi.org/10.1094/PDIS-11-17-1827-PDN
Wang Y, Yu FQ, Zhang C, Liu C, Yang M, Li S (2020) Edible Ectomycorrhizal Fungi and Their Cultivation in China. In Mushrooms, Humans and Nature in a Changing World (pp. 31–60). Springer, Cham.
White TJ, Bruns T, Lee JWT, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc 18:315–322. https://doi.org/10.1016/B978-0-12-372180-8.50042-1
Wickremasinghe R, Abeywickrama KP, Abeythunga DTU (1999) Isolation and identification of fungi from mushroom composts and evaluation their biological activity. https://doi.org/10.4038/jnsfsr.v27i1.2974
Willdenow CL, von Linne´ C, Link HF, Dietrich A, Schwa¨grichen CF, von Linne´ C, Link HF (1833) Caroli a Linne´ Species plantarum exhibentes plantas rite cognitas ad genera relatas cum differentiis specificis, nominibus trivialibus, synonymis selectis, locis natalibus, secundum systema sexuale digestas olim curante Carolo Ludovico Willdenow. Impensis GC Nauck, Berolini.
Williamson B, Tudzynski B, Tudzynski P, Van Kan JA (2007) Botrytis cinerea: the cause of grey mould disease. Mol Plant Pathol 8:561–580
Wood FC (1957) Diseases of cultivated mushrooms. Nat Lond 179:328–328
Woudenberg JHC, Sandoval-Denis M, Houbraken J, Seifert KA, Samson RA (2017) Cephalotrichum and related synnematous fungi with notes on species from the built environment. Stud Mycol 88:137–159. https://doi.org/10.1016/j.simyco.2017.09.001
Wu F, Zhou LW, Yang ZL, Bau T, Li TH, Dai YC (2019) Resource diversity of Chinese macrofungi: edible, medicinal and poisonous species. Fungal Diversity 98:1–76. https://doi.org/10.1007/s13225-019-00432-7
Wu XY, Li Y (2020) First Report of Cobweb Disease Caused by Cladobotryum mycophilum on Cultivated Shiitake Mushroom (Lentinula edodes) in China. Plant Dis 104:573. https://doi.org/10.1094/PDIS-07-19-1434-PDN
Xiong XL (2011) A research of pathogenic Trichoderma of Flammulina velutipes in Sichuan and the analysis of resistantce for Flammulina velutipes strains. Sichuan Agricultral University, Master degree dissertation.
Xu F, Wang SX, Liu Y, Ma YW, Zhang DP, Zhao S (2014) First report of Sporobolomyces symmetricus induced red spot disease of Pleurotus eryngii in China. Plant Dis 98:693–693. https://doi.org/10.1094/PDIS-09-13-0928-PDN
Xu R, Liu X, Pen B, Liu P, Li Z, Dai Y, Xiao S (2020) Genomic features of Cladobotryum dendroides, which causes Cobweb disease in edible mushrooms, and identification of genes related to pathogenicity and mycoparasitism. Pathogens 9:232. https://doi.org/10.3390/pathogens9030232
Xu R, Liu ZH, Fu YP, Li Y (2019) Identification and biological characteristics of Cladobotryum mycophilum causing cobweb disease on Ganoderma lingzhi. Mycosystema 38:669–678. https://doi.org/10.13346/j.mycosystema.180328
Zare R, Gams W (2001) The genera Lecanicillium and Simplicillium gen. nov. Nov Hedwigia 73:1–50
Zaree R, Ghanbari Z, Sadeghi AA, Kalanaki S (2005) Fungi isolated from the cultivated mushroom, Agaricus bisporus, in Tehran province with special reference to Verticillium fungicola in Iran.
Zargarzadeh Z, Mohammadi Goltapeh E, Danesh RY, DaneshMehrparvar M (2011) Identification of aggressive species of Trichoderma from button mushroom farms (Agaricus bisporus) using morphological and molecular methods. Rostanih 12:83–89. https://doi.org/10.22092/BOTANY.2011.101434
Zeng XY, Yuan XX, Peng KQ, Pan YT, Tan TJ, Wu N, Tian F-H (2022) Taxonomy and control of Trichoderma hymenopellicola sp. nov. responsible for the first green mold disease on Hymenopellis raphanipes. Front Microbiol 13:991987. https://doi.org/10.3389/fmicb.2022.991987
Zeng ZQ, Zhuang WY (2022) Three New Species of Clonostachys (Hypocreales, Ascomycota) from China. J Fungi 8:1027. https://doi.org/10.3390/jof8101027
Zhang GZ, Tang CY (2015) First report of Acrostalagmus luteo-albus causing red rust of needle mushroom (Flammulina velutipes) in China. Plant Dis 99:158–158. https://doi.org/10.1094/PDIS-07-14-0728-PDN
Zhang JC, Kong XH, Zhang PQ, Liu JN, Ma YP, Dai XD, Han ZH, Ma QF, Wang XY, Yu LP (2017) Identification of a new fungal pathogen causing white villous disease on the fruiting body of the culinary-medicinal mushroom Auricularia auricula-judae (Agaricomycetes) in China. International Journal of Medicinal Mushrooms 19:155–161. https://doi.org/10.1615/IntJMedMushrooms.v19.i2.70
Zhang K, Bonito G, Hsu CM, Hameed K, Vilgalys R, Liao HL (2020a) Mortierella elongata increases plant biomass among non-Leguminous crop species. Agronomy 10:754. https://doi.org/10.3390/agronomy10050754
Zhang T, Lu MZ, Zhang CL, Xu JZ (2019) First Report of Trichoderma longibrachiatum causing green mold disease on Ganoderma lingzhi. Plant Dis 103:156. https://doi.org/10.1094/PDIS-05-18-0818-PDN
Zhang XM, Zhuang WY (2006) Phylogeny of some genera in the Nectriaceae (Hypocreales, Ascomycetes) inferred from 28S nrDNA partial sequences. Mycosystema 25:15–22
Zhang Y, Qiao M, Baral HO, Xu J, Zhang KQ, Yu ZF (2020b) Morphological and molecular characterization of Orbilia pseudopolybrocha and O. tonghaiensis, two new species of Orbiliaceae from China. Int J Syst Evol Microbiol 70:2664–2676. https://doi.org/10.1099/ijsem.0.004088
Zhang Y, Wu WP, Hu DM, Su YY, Cai L (2014) A new thermophilic species of Myceliophthora from China. Mycol Prog 13:165–170. https://doi.org/10.1007/s11557-013-0904-8
Zhang YW, Chen W, Zeng G, Zou X, Wen T, Han Y, Qiu SY, Liang ZQ (2016) Two new Chrysosporium (onygenaceae, onygenales) from China. Phytotaxa 270:210–216. https://doi.org/10.11646/phytotaxa.270.3.5
Zhao XJ, Shi YX, Xie XW, Li BJ (2013) Identification of a new pathogen of Flammulina velutipes. Edible Fungi 5:57–58 ((In Chinese))
Zheng RY, Chen GQ, Huang H, Liu XY (2007) A monograph of Rhizopus. Sydowia 59:273–372
Zhu ZX, Zhuang WY (2013) Resources of nonlichenized fungicolous Ascomycota from China. Mycosystema 32:79–88
Zhuang WY (2020) Flora Fungorum Sinicorum Vol. LX Hypocreaceae. Science Press, Beijing
Zuo B, Lu BH, Liu XL, Wang Y, Ma GL, Gao J (2016) First report of Cladobotryum mycophilum causing cobweb on Ganoderma lucidum cultivated in Jilin province, China. Plant Dis 100:1239–1239. https://doi.org/10.1094/PDIS-12-15-1431-PDN
Acknowledgements
This study is supported by the Second Tibetan Plateau Scientific Expedition and Research (STEP) Program (Grant No. 2019QZKK0503), Major science and technology projects and key R&D plans/programs, Yunnan Province (202202AE090001), Natural Science Foundation of Guizhou Province (Grant No. Qian Ke Zhong Yin Di [2021]4031, Qian Ke He Zhi Cheng [2021] Generally 200), the open research project of “Cross-Cooperative Team” of the Germplasm Bank of Wild Species, Kunming Institute of Botany, Chinese Academy of Sciences (Grant No. 292019312511043), the Biodiversity Survey and Assessment Project of the Ministry of Ecology and Environment, PR China (2019HJ2096001006), the National Research Council of Thailand (NRCT) grant “Total fungal diversity in a given forest area with implications towards species numbers, chemical diversity and biotechnology” (Grant No. N42A650547), Natural Science Foundation of China (No. 32072645). Guizhou Provincial Science and Technology Projects ZK [2023] general 087. Feng-Ming Yu thanks Dr. Xiang-Hua Wang (Kunming Institute of Botany, Chinese Academy of Sciences) for providing valuable guidance for this paper.
Author information
Authors and Affiliations
Contributions
F-MY is responsible for culture isolations, identifications and descriptions, data analyses, drafting and editing the manuscript; F-MY, C-J-YL and QZ are responsible for sampling; KDH and QZ for funding acquisition; RSJ, TL, Xiang-Yu Zeng, C-J-YL, S-XB, HB, D-QZ, S-MT, KDH and QZ conceiving this study, and improving the manuscript; RSJ, TL, X-YZ, KDH and QZ contributed to the data analyses.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Handling Editor: Hiran Ariyawansa.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Yu, FM., Jayawardena, R.S., Luangharn, T. et al. Species diversity of fungal pathogens on cultivated mushrooms: a case study on morels (Morchella, Pezizales). Fungal Diversity 125, 157–220 (2024). https://doi.org/10.1007/s13225-023-00531-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13225-023-00531-6