Lophophorata is a clade composed of three Phyla: Phoronida, Brachiopoda, and Bryozoa. They are grouped like this due to one uniting structure, the lophophore (Jang & Hwang, 2009; Temereva & Kuzmina, 2017).
Morphology
Let’s begin with the pièce de resistance: the lophophore. It is a feeding structure, similar to a bunch of ciliated tentacles, that surround the mouth; therefore, lophophorates are suspension feeders. The cilia create strong water circulation, allowing for gas exchange, the exportation of gametes, and food particle delivery. The lophophore is thought to be the result of convergent evolution (Halanych, 1996). The gut of a lophophorate is U-shaped, so the direction of water flow prevents the mixing of food and waste products.
Phylum Phoronida
Phoronids are also known as horseshoe worms and build chitinous tubes to protect and support their soft bodies (Abele, Gilmour, & Gilchrist, 1983). Phoronids can retract and extend their lophophore, and cilia manipulate food into their mouth. Phoronids actively assess the flow of the water current and can reorient themselves as water flow changes to maximise their food-capturing ability. Their diet includes zooplankton, detritus, and invertebrate larvae.
Phoronis australis.
Phylum Brachiopoda
Please don’t be a pleb and confused them with Bivalvia. Brachiopoda has upper and lower valves, as opposed to left and right valves of bivalves. Anyway, they don’t even share the same ancestry.
The lophophore is connected to the lower brachial valve and is supported by cartilage and, sometimes, a brachidium (calcareous support attached to the brachial valve). Most brachiopods attach themselves to hard substrates by a stalk called the pedicle, a connective tissue that is part of the upper pedicle valve.
Inarticulate brachiopods (Lingula anatine).
Class Articulata
This class is defined by its “tooth-and-socket” hinge arrangement and a simple muscle strategy that opens and closes the hinges.
Class Inarticulata
This class has untoothed hinges and a more complex muscular strategy for aligning the valves.
Phylum Bryozoa
Bryozoans resemble phoronids, except bryozoans are microscopic, typically about 0.5mm in length. All genera, except one, form colonies that resemble moss. Each individual is called a zooid and has a hard casing called a cystid and a polypide that holds the organs in place. Within the colony, individual zooids may share resources through internal connections, and some zooids may specialise in a function. Vibracula zooids have a long bristle thought to function as defence or vibrate to keep the colony from becoming covered with sediment.
A freshwater bryozoan species.
References
Abele, L. G., Gilmour, T., & Gilchrist, S. (1983). Size and shape in the phylum Phoronida. Journal of Zoology, 200(3), 317–323.
Halanych, K. M. (1996). Convergence in the feeding apparatuses of lophophorates and pterobranch hemichordates revealed by 18S rDNA: an interpretation. The Biological Bulletin, 190(1), 1–5.
Jang, K. H., & Hwang, U. W. (2009). Complete mitochondrial genome of Bugula neritina (Bryozoa, Gymnolaemata, Cheilostomata): phylogenetic position of Bryozoa and phylogeny of lophophorates within the Lophotrochozoa. Bmc Genomics, 10(1), 1–18.
Temereva, E. N., & Kuzmina, T. V. (2017). The first data on the innervation of the lophophore in the rhynchonelliform brachiopod Hemithiris psittacea: what is the ground pattern of the lophophore in lophophorates?. BMC evolutionary biology, 17(1), 1–19.
There’s a mollusc, see? And he walks, well he doesn’t walk up, he swims up. Well, actually, the mollusc isn’t moving, he’s in one place and then the sea cucumber… I mixed up. There was a mollusc and a sea cucumber. None of them were walking. Normally, they don’t talk, but in a joke, everyone talks. So, the sea mollusc says to the cucumber, “with fronds like these, who needs anemones!”
Mollusca forms the second largest invertebrate phylum and has a diverse evolutionary history and wide range of feeding and life-history strategies that have led to its success on land, freshwater, and the ocean (Rosenberg, 2014). Molluscs can be herbivores, carnivores, scavengers, and filter feeders, and they are responsible for the consumption of large amounts of organic matter. They themselves serve as food for a range of predators, and thus, are essential links in the food chain.
Morphology
All species of Mollusca have bilateral symmetry, lack of segmentation, a structured nervous system, and a mantle. When discussing mollusc morphology, a generalised overall structure is usually referred to, though many species are an exception to this structure.
Their head contains the mouth and feeding structures and a ganglion of nervous systems. The visceral mass is the metabolic region containing the stomach, heart, intestines, gonads, etc. Covering the visceral mass is the mantle which secretes the shell, and a fold on the mantle known as the mantle cavity contains excretory and respiratory organs. The shell thickens with age as it is secreted by the mantle and consists of three layers:
Outer layer – aka periostracum is composed of a durable organic material and may develop as a thin smooth coating, into hairs or into flexible spine-like outgrowths.
Middle layer – aka prismatic is made of columnar calcite.
Inner layer – is often nacreous (think iridescent and pearl-like) and laid down in thin sheets by the epithelial parts of the mantle.
Torsion
Torsion is evident in gastropods (snails) during larval development. Two torsion events of 90° result in 180° rotation of the mantle cavity and the organs it contains to an anterior position above the head (Page, 2006). A possible disadvantage of torsion is that the anus excreting above the head could cause fouling of the mouth and sensory organs. However, the success of the class Gastropoda suggests this may not be an issue. Possible advantages of torsion are that it allows the animal to retract its vulnerable head into the shell efficiently and in marine species, the anterior positioning may prevent sediment from entering the mantle cavity.
Feeding
The radula is unique to molluscs and is found in every class except bivalves (Steneck & Watling, 1982). It is a chitinous ribbon studded with small, hard teeth, used for scraping or cutting food before it enters the oesophagus. The radula protrudes from the floor of the buccal cavity, where the odontophore underlies the radula membrane and controls its protrusion and return. As the radula retracts into the buccal cavity, the teeth rasp food particles from the substrate and food is deposited into the pharynx. As teeth wear, new teeth are continuously being secreted, shaped, and added to the cuticle ribbon inside the radula sac.
Food particles pass from the pharynx to the oesophagus and then to the stomach, where digestion occurs intracellularly and extracellularly within folds of the stomach called diverticula. The stomach may have several functions: sorting, grinding, and digesting food particles.
Systems
The circulatory system is usually open, and blood flows through the haemocoel cavity. The respiratory pigment is called hemocyanin, which is pale blue when oxygenated and clear when deoxygenated. This blue is from the copper contained in the oxygen-binding molecules, as opposed to the red blood of mammal’s iron found in haemoglobin.
The nervous system is typically a mass of nerve cell bodies that associate with the sensory organs. The sensory organs often include eyes, statocysts (the sensory organ that orients animal to gravity, located in the foot), osphradia (sensory epithelium which act as chemoreceptors), and tentacles.
Classifications
Neogastropoda
Neogastropoda includes sea snails and is primarily carnivorous. They have a proboscis that extends out and can drill through shells of bivalves or is used to suck up nutrients from its prey (much like a butterfly feeding on nectar). Some species have a siphon to draw water into the mantle cavity to oxygenate the gill.
Mud whelks (Nassarius jacksoniania) eating a dead fish.
Bivalvia
Bivalvia includes your classic shellfish, e.g., clams, mussels, oysters, scallops, and cockles. They lack a radula, an odontophore, and a head. The name bivalve means “two shells”, which is exactly what they have, and these shells are connected by a hinge and are left and right, as opposed to top and bottom as with Brachiopoda (don’t worry, we’ll get there). The lack of head is made up for with their foot. It is usually well-developed and excellent for digging and ploughing through sediment. Some bivalves, e.g., mussels, have a byssus thread used to attach to hard substrates, and I guarantee you will notice it the next time you eat a mussel.
Little black mussel (Xenostrobus pulex).
Small giant clam (Tridacna maxima).
Nudibranchia
Nudibranchia contains the crazy, colourful nudibranchs: a group of around 3,000 species of soft-bodied molluscs. But how can they be molluscs if they don’t have a shell? They shed their shell in the larval stage (Thompson, 1959). Along with a naked body, they also lack a mantle cavity, meaning that the nudibranchs probably like the term mollusc to be used loosely! To add to their peculiar nature, they are all carnivorous mostly feeding with a radula, and some store nematocysts from their Cniadrian prey and use them as a defence mechanism (Frick, 2003). They are hermaphroditic, but cannot fertilise themselves, and mate after a courting dance takes place.
Phyllidia babai.
Mexichromis macropus.
Dendronotus albus.
Cephalopoda
And if you thought nudibranchs didn’t fit into the Mollusca mould, then don’t even bother reading about cephalopods. With their name meaning “head-feet”, Cephalopoda contains over 800 living species of octopus, squid, cuttlefish, and nautilus. They have bilateral symmetry and, as their name suggests, a prominent head atop a set of arms or tentacles that have evolved from the molluscan foot. And before you ask, “arms” are the suction cup ones, and “tentacles” only have suction cups at the end – some species have one or the other or both.
When you think of squid, you don’t immediately associate them with their bivalve or gastropod relatives mainly because, well, the latter two have a shell and a squid does not, right? Well, the answer is tricky, and, like with every marine invertebrate, I will forgive you for thinking something is something that it is not. Nautiluses have an external shell that is visible to the naked eye, still the case of cuttlefish, octopuses, and squid is slightly more complex. Some cephalopods have a vestigial shell, some have organic, internal, calcium carbonate structures, and some may have just evolved to lose their shell entirely (Baratte, Andouche, & Bonnaud, 2007; Furuhashi, Schwarzinger, Miksik, Smrz, & Beran, 2009; Warnke & Keupp, 2005).
Cephalopods are often regarded as extremely intelligent, with complex nervous systems and the ability to use tools and problem solve (Budelmann, 1995; Finn, Tregenza, & Norman, 2009; Richter, Hochner, & Kuba, 2016; Schnell, Amodio, Boeckle, & Clayton, 2021).
The genus Hapalochlaena contains four extremely venomous octopus species, more commonly known as the blue-ringed octopuses. They are tiny, reaching maximum sizes of 20cm, but deadly, with one animal containing enough tetrodotoxin to kill 26 adult humans with a painless bite that can paralyse within minutes.
The greater blue-ringed octopus (Hapalochlaena lunulata.).
The Palau nautilus (Nautilus belauensis).
The common cuttlefish (Sepia officinalis) – note its “W” shaped pupil thought to be useful in improving horizontal vision (Mäthger, Hanlon, Håkansson, & Nilsson, 2013).
The Hawaiian bobtail squid (Euprymna scolopes) reaches a max of 3cm.
References
Baratte, S., Andouche, A., & Bonnaud, L. (2007). Engrailed in cephalopods: a key gene related to the emergence of morphological novelties. Development genes and evolution, 217(5), 353–362.
Budelmann, B. U. (1995). The cephalopod nervous system: what evolution has made of the molluscan design. In The nervous systems of invertebrates: An evolutionary and comparative approach (pp. 115–138). Birkhäuser Basel.
Finn, J. K., Tregenza, T., & Norman, M. D. (2009). Defensive tool use in a coconut-carrying octopus. Current biology, 19(23), R1069–R1070.
Frick, K. (2003). Predator suites and flabellinid nudibranch nematocyst complements in the Gulf of Maine. DIVING FOR SCIENCE… 2003, 37.
Furuhashi, T., Schwarzinger, C., Miksik, I., Smrz, M., & Beran, A. (2009). Molluscan shell evolution with review of shell calcification hypothesis. Comparative biochemistry and physiology Part B: Biochemistry and molecular biology, 154(3), 351–371.
Mäthger, L. M., Hanlon, R. T., Håkansson, J., & Nilsson, D. E. (2013). The W-shaped pupil in cuttlefish (Sepia officinalis): functions for improving horizontal vision. Vision research, 83, 19-24.Page, L. R. (2006). Modern insights on gastropod development: reevaluation of the evolution of a novel body plan. Integrative and Comparative Biology, 46(2), 134–143.
Richter, J. N., Hochner, B., & Kuba, M. J. (2016). Pull or push? Octopuses solve a puzzle problem. PloS one, 11(3), e0152048.
Rosenberg, G. (2014). A new critical estimate of named species-level diversity of the recent Mollusca. American Malacological Bulletin, 32(2), 308–322.
Schnell, A. K., Amodio, P., Boeckle, M., & Clayton, N. S. (2021). How intelligent is a cephalopod? Lessons from comparative cognition. Biological Reviews, 96(1), 162–178.
Steneck, R. S., & Watling, L. (1982). Feeding capabilities and limitation of herbivorous molluscs: a functional group approach. Marine Biology, 68(3), 299–319.
Thompson, T. E. (1959). Feeding in nudibranch larvae. Journal of the Marine Biological Association of the United Kingdom, 38(2), 239–248.
Warnke, K., & Keupp, H. (2005). Spirula—a window to the embryonic development of ammonoids? Morphological and molecular indications for a palaeontological hypothesis. Facies, 51(1), 60–65.
There are less than 200 known species of ctenophores, all of which are found exclusively in marine habitats. Ctenophores, more commonly known as the comb jellies, resemble cnidarian medusa (I will forgive you for confusing them with jellyfish), but ctenophores have a few specific features that make them unique.
Morphology
Opposed to jellyfish, who have radial symmetry, ctenophores have bilateral symmetry (Pang & Martindale, 2008). They don’t use jet propulsion like our Scyphozoan friends, but rather are the largest animals to swim with the help of cilia, with adults range from a few millimetres to 1.5 metres (Tamm, 2015). The cilia are packed in the thousands into ctenes, or comb plates, which are organised into comb rows around the body (Tamm, 2014).
Ctenophores are known to be bioluminescent, where the photoproteins, located in canals under the comb rows, are activated (Haddock & Case, 1999; Pang & Martindale, 2008). This is not to be confused with the rainbow effect of the comb rows produced as their cilia beat and scatter light (Welch, Vigneron, Lousse, & Parker, 2006).
A brief annotation of the anatomy of a ctenophore.
Feeding
Ctenophores have sticky cells in the epidermis of their tentacles called colloblasts which capture food. The tentacles expand when ready to capture food and the branches on the tentacles separate. The stickiness of the colloblasts allows the organism to “fire” a fibre with adhesive granules to capture food (e.g., copepods).
Colloblasts are linked to a nervous system that triggers the retraction of the tentacles towards the mouth when prey is captured. Prey is then wiped inside the mouth, swallowed, and liquified into a slurry by enzymes and muscular contractions in the pharynx. Cilia beat and distribute the slurry through the canal system where digestion occurs intra and extracellularly. Some waste is released through the anal pores but usually they regurgitate waste from the mouth.
Colloblasts are unique to ctenophores and are found in the epidermis of the tentacles and, similar to nematocysts of Cnidaria, the colloblasts are discharged from the tentacles and capture prey (Franc, 1978). However, colloblasts are not venomous but rather they are adhesive and stick to their prey. The rare ctenophore, Haeckelia rubra, has rid itself of colloblasts completely and instead collects nematocysts from their cnidarian prey (Mills & Miller, 1984).
Nervous system and navigation
Ctenophores have a structure, know as the statocyst, that aids in their navigation and orientation through gravitational sensitivity (Pang & Martindale, 2008; Tamm, 2015). If a ctenophore is pulled off balance, its statocyst will direct beating of specific comb rows in order to right itself. For movement over long distances, ctenophores mainly rely on ocean currents (Pang & Martindale, 2008). Instead of a nervous system, ctenophores have a complex nerve net that works closely with the statocyst and ctenes (Pang & Martindale, 2008; Tamm, 2014; Tamm, 2015).
Reproduction
Floating somewhat aimlessly around in the ocean as a relatively tiny and hard-to-see individual means the likelihood of you meeting someone you can have babies with is quite low. To combat this, almost all ctenophores are self-fertile hermaphrodites (Pang & Martindale, 2008).
Classifications
Class Tentaculata
Tentaculata have, you guessed it, tentacles. Commonly, they have long, feathery tentacles which are equipped with colloblasts.
Mertensia ovum, a species of ctenophore from Class Tentaculata (note the two, prominent tentacles trailing behind.
Class Nuda
If Class Tentaculata have tentacles, then Class Nuda must have no tentacles. Organisms from Nuda are known as beroids (from the monophyletic order Beroida), and they feed by using their large mouths to engulf prey. Alternatively, some species spread their lips over prey whilst a sword-like structure chops the prey up (Tamm & Tamm, 1991). Beroids actively hunt their prey, which is usually soft-bodied organisms such as ctenophores – yes, they eat their own kind.
A species of beroid ctenophore from the Class Nuda (note its lack of tentacles).
References
Franc, J. M. (1978). Organization and function of ctenophore colloblasts: an ultrastructural study. The Biological Bulletin, 155(3), 527–541.
Haddock, S. H., & Case, J. F. (1999). Bioluminescence spectra of shallow and deep-sea gelatinous zooplankton: ctenophores, medusae and siphonophores. Marine Biology, 133(3), 571–582.
Mills, C. E., & Miller, R. L. (1984). Ingestion of a medusa (Aegina citrea) by the nematocyst-containing ctenophore Haeckelia rubra (formerly Euchlora rubra): phylogenetic implications. Marine Biology, 78(2), 215–221.
Pang, K., & Martindale, M. Q. (2008). Ctenophores. Current Biology, 18(24), R1119–R1120.
Tamm, S. L. (2014). Cilia and the life of ctenophores. Invertebrate Biology, 133(1), 1–46.
Tamm, S. L. (2015). Functional consequences of the asymmetric architecture of the ctenophore statocyst. The Biological Bulletin, 229(2), 173–184.
Tamm, S. L., & Tamm, S. (1991). Reversible epithelial adhesion closes the mouth of Beroe, a carnivorous marine jelly. The Biological Bulletin, 181(3), 463–473.
Welch, V., Vigneron, J. P., Lousse, V., & Parker, A. (2006). Optical properties of the iridescent organ of the comb-jellyfish Beroë cucumis (Ctenophora). Physical Review E, 73(4), 041916.
Phylum Cnidaria contains all your favourite ocean stingers from jellyfish to corals to anenomes, and we can’t forget our favourite Hydrozoan, the Portuguese man o’ war.
Description
Morphology
The body wall is composed of three layers:
Epidermis tissue – outer layer.
Mesoglea – “jelly in the middle” composed of mucopolysaccharides & collagen; is not true tissue but provides support, buoyancy, and locomotion.
Gastrodermis tissue – inner layer which lines the gastrovascular cavity.
A distinguishing feature of cnidaria is their simple gastrovascular cavity, present in only one other primitive phylum. It is a two-way system, where food enters through an opening that serves as a mouth and an anus and is extracellularly digested within the gastrovascular cavity, then waste exits back through the same hole (Shostak, 2001).
Medusa & polyp form
Cnidaria can exist in two forms: medusa or polyp (Piraino, Boero, Aeschbach, & Schmid, 1996; Seipel & Schmid, 2004). Some cnidarians only exhibit a polyp or medusa form, or have one predominantly over the other. Some may pass through both forms throughout their life histories.
Polyp form:
Sessile
Cylindrical tubes point upwards
The oral end is on top
The aboral end usually attached to the substrate
Tentacles point upward
E.g., sea anemone
Some polyps form colonies (e.g., coral)
Medusa form:
Swims
Is the inverse of the polyp
The oral end is below a bell-shaped body
Th aboral end is to top of the umbrella-like structure (velum)
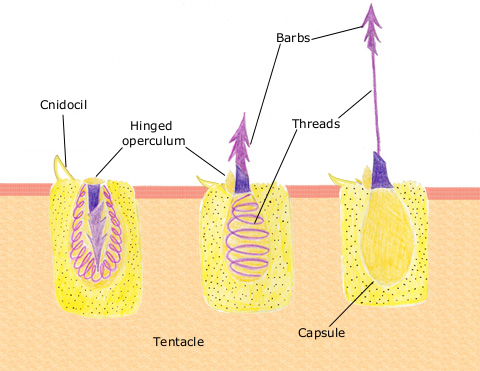
The nematocyte is a specialised, ectodermal stinging cell involved in defence and prey capture. It contains the nematocyst (Beckmann & Özbek, 2012; David et al., 2008; Östman, 2000).
The nematocyst is an organelle inside the nematocyte consisting of an ejectable thread that causes a sting and injects toxins into predators or prey. The nematocyst capsule is firm and made from a type of collagen, and it holds a coiled thread that can be barbed, smooth, or hold toxins. A chemical or physical stimulant stimulates the ejection of this thread, and once stimulated, will uncoil and extrude out, penetrating or wrapping around the prey. Once paralysed, captured, or killed (who knew sea anemones are so gruesome??), the tentacles will move the prey to the oral cavity to be digested by the gastrovascular cavity.
Showing the discharge of a barbed nematocyst from a nematocyte.
There are three types of nematocysts:
Penetrant – barbed thread with open tip; when discharged, it pierces the skin/exoskeleton and injects venom to paralyse or kill.
Glutenant – smooth or bristled thread with an open tip that is sticky and has toxins.
Volvent – smooth, lasso-like thread with closed tip entangles prey.
Reproduction
Cnidaria are capable of both sexual and asexual reproduction (Shostak, 2001). Sexual reproduction involves gametes, usually produced in separate individuals, and are fertilised in the gut, ovary, or water after being released by the mouth, tentacles, or breaks in the epidermal layer. Female gametes may produce a substance that attracts male gametes. Asexual reproduction usually happens in warmer months, where a bud develops via evagination from the adult body wall and contains an extension of the gastrovascular cavity. Once fully developed, it detaches from the parent.
Coral
Coral have a unique symbiotic relationship with zooxanthellae which gives the coral a range of different colours (Shostak, 2001). Coral produce carbon dioxide (CO2) and ammonium (NH4+) as a by-product of cellular respiration, and zooxanthellae use the CO2 and NH4+ to conduct photosynthesis which, in turn, supplies the coral with sugars, lipids, and oxygen.
Coral bleaching is a phenomenon where the coral consumes or expels their symbiotic inhabitants to ensure short-term survival when exposed to stressful conditions such as rising water temperature, leading to a white “bleached” appearance (Hoegh-Guldberg, 1999; Lesser, 2011; Nir, Gruber, Shemesh, Glasser, & Tchernov, 2014). The coral continues to live after bleaching, but under a prolonged, stressful environment, they will die from starvation.
Classifications
Class Anthozoa
This group includes sea anemones, stony corals, and soft corals.
Chrysaora melanaster, commonly known as the northern sea nettle or brown jellyfish, is a species of jellyfish native to the northern Pacific Ocean and adjacent parts of the Arctic Ocean.
A species of jellyfish from the genus Cephea, or the cauliflower jellyfish.
Class Cubozoan
Box jellyfish are distinguishable by their cube-shaped medusae. Some species have potent venom that can be extremely painful and potentially fatal to humans.
Chironex fleckeri, or the sea wasp, is thought to be the most lethal jellyfish in the world and is responsible for sixty-four deaths in Australia from 1884-2021 (Fenner & Williamson, 1996). C. fleckeri is said to contain enough venom to kill sixty adult humans, and stings are typically excruciatingly painful and, if left untreated, can kill within two to five minutes.
Chironex fleckeri.
Malo kingi, or the common kingslayer, is a species of Irukandji jellyfish named after one of its victims, Robert King (Gershwin, 2007). The Irukandji jellyfish are any of several box jellies that cause Irukandji syndrome after stinging their victims; Irukandji syndrome is characterised by severe pain, vomiting, and rapid rise in blood pressure. M. kingi are very small and inconspicuous in the water, making it hard for victims to see them.
Malo kingi.
Class Hydrozoa
To end on a fun note, let’s finish with Hydrozoa. They are a group of very small, predatory individuals that can live in solitude or within a colony. The colonial species can be large and sometimes the specialised individuals cannot survive outside of their colony. So, the next time you hear someone calling a Portuguese man o’ war (Physalia physalis) a jellyfish, feel free to roll your eyes and let them know they are actually a Hydrozoan.
Portuguese man o’ war (Physalia physalis) washed up on a beach.
References
Beckmann, A., & Özbek, S. (2012). The nematocyst: a molecular map of the cnidarian stinging organelle. International Journal of Developmental Biology, 56(6–7–8), 577–582.
David, C. N., Özbek, S., Adamczyk, P., Meier, S., Pauly, B., Chapman, J., … & Holstein, T. W. (2008). Evolution of complex structures: minicollagens shape the cnidarian nematocyst. Trends in genetics, 24(9), 431–438.
Hoegh-Guldberg, O. (1999). Climate change, coral bleaching and the future of the world’s coral reefs. Marine and freshwater research, 50(8), 839–866.
Lesser, M. P. (2011). Coral bleaching: causes and mechanisms. In Coral reefs: an ecosystem in transition (pp. 405–419). Springer, Dordrecht.
Nir, O., Gruber, D. F., Shemesh, E., Glasser, E., & Tchernov, D. (2014). Seasonal mesophotic coral bleaching of Stylophora pistillata in the Northern Red Sea. PLoS One, 9(1), e84968.
Östman, C. (2000). A guideline to nematocyst nomenclature and classification, and some notes on the systematic value of nematocysts. Scientia Marina, 64(S1), 31–46.
Piraino, S., Boero, F., Aeschbach, B., & Schmid, V. (1996). Reversing the life cycle: medusae transforming into polyps and cell transdifferentiation in Turritopsis nutricula (Cnidaria, Hydrozoa). The Biological Bulletin, 190(3), 302–312.
Seipel, K., & Schmid, V. (2004). Mesodermal anatomies in cnidarian polyps and medusae. International Journal of Developmental Biology, 50(7), 589–599.
Shostak, S. (2001). Cnidaria (Coelenterates). e LS.
Phylum Porifera, better known as the sponges, are an interesting group of animals. Yes, they are animals, albeit they are the most primitive of all multicellular creatures as they were the first to separate from the common ancestor of animals, but animals nonetheless (Feuda et al., 2017; Giribet, 2016). Porifera are the first group of marine invertebrates I will be chatting about over the next few weeks, so move over megafauna cause the little guys are in town!
Description
Morphology
Porifera is estimated to contain around 15,000 species of sponge, many of which have not even been described yet (Degnan et al., 2015). Sponges don’t exactly have any true tissues or organs; what they do have is a mesohyl (a gelatinous matrix that resembles a type of connective tissue) sandwiched between two thin layers of cells (more on the different types of cells soon). One of the craziest things about sponges is that they are an aggregation of cells in space and time; at that moment, they are part of that sponge, but they could be part of another sponge at a different time. Lavrov and Kosevich (2016) took this to the next level when they mechanically separated sponge cells and saw them reaggregate again before their very eyes!
Respiration, digestion, and excretion
Sponges have no nervous system, no digestive system, no excretory system, and no circulatory system, so how are they even… alive??? Sponges are sessile and are therefore attached to a substrate via their pinacocytes (contractile cells that line the outer wall). By orienting themselves perpendicular to the water flow to create low pressure at the excurrent opening (osculum), they then open and close incurrent pores (ostia) to regulate water flow; up to 20,000 times the volume of the sponge can be filtered through the sponge in 24 hours and up to 90% of the bacteria in the water may be filtered out. The structure of the sponge maximises the efficiency of water flowing into the ostia through the central cavity, where respiration and digestion occur (Hutchings, Kingsford, & Hoegh-Guldberg, 2019). As water flows through the body, cells absorb oxygen by diffusion and dump waste products into the outgoing current. This water flow also delivers food particles to the sponge. If the food is larger (>50μm), it cannot enter the ostia, so pinacocytes (remember these from earlier?) grab ‘em and digest ‘em. Usually, though, food particles are <0.5μm, so they can easily pass through the ostia to where the choanocytes are waiting. Choanocytes have a flagellum that beats, creating a unidirectional flow of water, drawing in food particles. Choanocytes have a collar of microvilli which filters nutrients from the water; the choanocytes then store the nutrients in vacuoles of adjacent cells, usually amoebocytes which distribute nutrients around the sponge. Some sponges, like Clarohizdae, are carnivorous and will passively capture small invertebrates via their sticky surface, where cells will migrate to and envelop the prey (Hestetun, Tompkins-Macdonald, & Rapp, 2017).
Morphology of a sponge, showing the osculua and ostia.
Spicules
A peculiar characteristic of Porifera is their spicules, structures made from either calcium carbonate (CaCO3) or silica that vary in size and shape from rods to three-dimensional stars (Renard et al., 2013). They are held in place by collagen fibres (one of the places where “marine collagen” comes from) and produced in the mesohyl by sclerocyte cells. Spicules are thought to be a deterrent from predators or to provide skeletal structure or support.
A six-pointed star spicule of a sponge.Three-pointed star and rod spicules of a sponge.
Classifications
Calcarea
These are calcareous sponges with CaCO3 spicules.
A calcareous sponge (Leucetta primigenia).
Hexactinellida
These are the glass sponges that mainly inhabit deep water. They have siliceous spicules that form stable lattices. You may remember these sponges from David Attenborough’s Blue Planet II, a romantic story of 2 shrimp larvae, male and female, that get swept into a Hexactinellid sponge and grow too large to be able to escape; a twist of fate that leaves them stuck together forever… awww.
White hexactinellida glass sponge known as a venus flower basket (Euplectella aspergillum).
Demospongiae
Some Demospongiae don’t have spicules, but if they do, they are siliceous spicules that are held together by collagen.
A purple encrusting sponge from the Strongylacidon genus.
References
Degnan, B. M., Adamska, M., Richards, G. S., Larroux, C., Leininger, S., Bergum, B., … & Degnan, S. M. (2015). Porifera. In Evolutionary developmental biology of invertebrates 1 (pp. 65–106). Springer, Vienna.
Feuda, R., Dohrmann, M., Pett, W., Philippe, H., Rota-Stabelli, O., Lartillot, N., … & Pisani, D. (2017). Improved modeling of compositional heterogeneity supports sponges as sister to all other animals. Current Biology, 27(24), 3864-3870.
Giribet, G. (2016). Genomics and the animal tree of life: conflicts and future prospects. Zoologica Scripta, 45, 14–21.
Hestetun, J. T., Tompkins-Macdonald, G., & Rapp, H. T. (2017). A review of carnivorous sponges (Porifera: Cladorhizidae) from the Boreal North Atlantic and Arctic. Zoological Journal of the Linnean Society, 181(1), 1–69.
Hutchings, P., Kingsford, M., & Hoegh-Guldberg, O. (Eds.). (2019). The Great Barrier Reef: biology, environment and management. Csiro publishing.
Lavrov, A. I., & Kosevich, I. A. (2016). Sponge cell reaggregation: Cellular structure and morphogenetic potencies of multicellular aggregates. Journal of Experimental Zoology Part A: Ecological Genetics and Physiology, 325(2), 158–177.
Renard, E., Gazave, E., Fierro‐Constain, L., Schenkelaars, Q., Ereskovsky, A., Vacelet, J., & Borchiellini, C. (2013). Porifera (sponges): recent knowledge and new perspectives. eLS.
The shortfin mako shark (Isurus oxyrinchus) is a pelagic, cartilaginous fish with a wide distribution range that covers most oceans and undergoes migrations that can be as large as 5,300km in just under 1.5 years (Barreto, de Farias, Andrade, Santana, & Lessa, 2016; Kohler, Turner, Hoey, Natanson, & Briggs, 2002). There is high demand for mako shark meat, and it is a prize game species in recreational fishing worldwide (Barreto et al., 2016).
As of 2019, I. oxyrinchus is classified as “Endangered” on the International Union for the Conservation of Nature’s (IUCN) Red List (Rigby et al., n.d.). Rigby et al. (n.d.) concluded that shortfin mako’s population trend is decreasing; there is an estimated decline everywhere except in the South Pacific and an overall estimated average reduction of 46.6% over 72-75 years. There is an estimated decline in biomass and abundance of 99.9% since the early 1800s, the main reason being overfishing (Ferretti et al., 2008).
Morphology
Shortfin mako circulatory systems utilise a heat-exchanging technique that raises their internal temperature above that of the surrounding environment (Carey, Teal, & Kanwisher, 1981; Kohler et al., 2002). They have streamlined bodies, and aerobic muscles centred closer to their rear, which aids in thunniform swimming and increases power (Donley, Sepulveda, Konstantinidis, Gemballa, & Shadwick, 2004; Wegner, Sepulveda, Olson, Hyndman, & Graham, 2010). Emery and Szczepanski (1986) concluded that the gill area of I. oxyrinchus is 2-3 times larger than other pelagic shark species, which could aid in the mako’s speed, agility, and ability to swim long distances. Shortfin mako habitat extends globally in tropical and temperate oceans and they can be found inshore in coastal areas or at least 500m down in oceanic zones (Kohler et al., 2002).
Life history
Sexual dimorphism is prevalent in I. oxyrinchus,with females often occurring larger than males, with males reaching a maximum size of 2.6m and females reaching a maximum size of 3.4m (Barreto et al., 2016; Cema & Lincandeo, 2009; Chan, 2001; Doño, Montealegre-Quijano, Domingo, & Kinas, 2014; Hsu, 2001; Natanson et al., 2006; Semba, Nakana, & Aoki, 2009). Shortfin mako sharks are oophagous and ovoviviparous (Kohler et al., 2002), and a study by Mollet et al. (2000) predicts a gestation period of 15-18 months, although a study by Duffy and Francis (2001) puts makos in New Zealand waters at a 21-month gestation period. They have a 3-year reproductive cycle (Mollet & Cailliet, 2002). Bishop, Francis, Duffy, and Montgomery (2006) concluded that New Zealand shortfin mako births are concentrated in spring and gave a theoretical birth date of 1 October, with the average length of the shark at birth to be 61cm. Francis and Duffy’s (2005) study on sexual maturity of New Zealand shortfin makos concluded that maturity occurs between 7-9 years for males and 19-21 years for females (Bishop et al., 2006). Bishop et al. (2006) found evidence of New Zealand shortfin makos living to 29 years, although this number is probably higher because there is a lower chance of catching older sharks which make up a small percentage of the overall population. The same study found the sharks grow quickly within their first year after birth; this growth rate rapidly reduces in the subsequent years to steadier growth. Late maturity, moderately long longevity, the estimated low natural mortality rate, and low annual fecundity causes low productivity in the species (Bishop et al., 2006; Mollet et al., 2000).
Diet
Stillwell and Kohler (1982) analysed the stomach contents of shortfin mako sharks and found evidence of bony fish, swordfish, and cephalopods. As shark body length increased, so did the average volume of food, indicating that as makos grow larger, they may switch to larger prey items (Kohler et al., 2002).
Mortality
Sharks have evolved for 400 million years (Donley et al., 2004), leading species such as I. oxyrinchus to be apex predators at the top of their food chain; they have no natural predators which results in low natural mortality. There is evidence of parasitic activity in shortfin makos, although it could not be determined if there was a negative effect on the shark (Borucinska & Hege, 1999). One example of anthropogenically influenced mortality is seen in 1966 when a longfin mako (Isurus paucus) died from a fishing hook retained in its flesh (Adams, Borucinska, Maillett, Whitburn, & Sander, 2015).
Ecology
Sharks are keystone species and have a strong effect on multiple ecosystems due to their predatory role and wide dispersal range (Feretti, Worm, Britten, Heithaus, & Lotze, 2010). I. oxyrinchus is considered to be a large shark with “strong, top-down forces” (Feretti et al., 2010, p1055). Therefore, their removal from an ecosystem is highly likely to drastically alter communities, induce trophic cascades, release mesopredators such as smaller sharks and rays, and consequently cause a decline in commercial fish stocks (Fig. 1).
Figure 1 “Documented ecosystem effects of fishing large sharks. Depicted are trophic (solid arrows) and behavioral (dotted arrows) interactions between humans, large and mesopredator elasmobranchs and their prey species. Block arrows represent overall population trends of the various functional groups. Regions in which particular interactions have been documented (see text) are indicated by letters (A, Australia; C, Caribbean; E, Europe; G, Gulf of Mexico; M, Mediterranean Sea; N, North American East Coast; P, Central Pacific; S, South Africa; W, North American West Coast).” (Feretti et al. 2010)
Threats
There is a high commercial demand for shortfin mako shark meat, and the species is exploited globally (Barreto et al., 2016). Peru is one of the biggest shark-fishing nations, and I. oxyrinchus is one of the top 2 most caught species with a rarely enforced catch size limit (Gonzalez-Pestana, Kouri, & Velez-Zuazo, 2016; Fischer, Erikstein, D’Offay, Guggisberg, & Barone, 2012). Brazilian fleets in the western and central South Atlantic exploit immature shortfin makos, specifically females (Barreto, 2016); it was rare for them to catch individuals greater than 2m. On a global scale, it is both a target and bycatch species in commercial and small-scale fisheries, including longline, gillnet, purse seine, trammel net, and trawls (Camhi, Pikitch, & Babcock, 2008; Rigby et al., 2019). There is likely underreporting of catch, and commercial post-release mortality from longlines alone is reported at 30-33% (Campana, Joyce, Fowler, & Showell, 2016; Rigby et al., 2019). Shortfin mako fins made up 1.2% of shark fin imported to Hong Kong in 2014 (Fields et al., 2017), and their skin, jaws, and liver oil are also used (Compagno, 2001). In New Zealand’s EEZ, I. oxyrinchus is a common bycatch species on tuna longlines and less commonly on pelagic longlines, trawls, and set nets (Bagley et al., 2000; Francis, 1998; Francis, Griggs, Baird, Murray, & Dean, 2000; Martinsohn & Muller, 1992). Since 1993, commercial catch averaged 60 tonnes annually, but observer reporting estimated 100-200 tonnes per year from the longline tuna fishery alone (Francis, 1998; Francis et al. 2000); the discrepancy between these numbers could be related to lack of accurate recording. From 1988-2015, the New Zealand tuna longline fishery total catch was comprised of 11.1% target species (southern bluefin tuna, bigeye tuna, and swordfish) and 88.9% bycatch (including albacore tuna, lancetfish, porbeagle shark, deepwater dogfish, dealfish, mako shark, moonfish, escolar, sunfish, and butterfly tuna) according to a report by Fisheries New Zealand [FNZ] (2018). Since October 2014, shark finning is illegal in New Zealand, yet over half of all makos caught by charter vessels in the 2014-15 tuna longline fishery year were kept for their flesh (FNZ, 2018). The low productivity of mako sharks causes them to be vulnerable to overfishing and makes it hard for the species to replace lost individuals. CITES (2019) reported that shortfin mako is in danger of population collapse due to the overfishing of most of the juveniles since the 1980s; this means that sharks dying of old age now will not be replaced with mature individuals leaving a 10-20-year gap.
The mining of precious metals increases the presence of toxic contaminants in the environment, especially in coastal environments, specifically mercury (Maz-Courrau et al., 2011). Sharks bioaccumulate mercury through their tissues and organs from the environment and through the food they eat; mercury travels up the trophic web and magnifies at each level, eventually accumulating in shark muscular tissue (Maz-Courrau et al., 2011). Maz-Courrau et al. (2011) found that 33% of shortfin mako sharks they studied off the Baja California coast had levels of mercury higher than was estimated to be fit for human consumption by Mexican Law and Watling et al. (1981) found mercury concentrations in mako sharks off the coast of Australia to be almost twice that. As Maz-Corrau et al. (2011) only sampled juveniles, it is likely that as sharks grow and eat more, more mercury is accumulated into their tissue over time. When people eat this contaminated tissue, it bioaccumulates up the food chain again and causes adverse health effects in humans.
Current/Proposed Management Actions
As of 2019, I. oxyrinchus officially met criteria to be listed on the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) Appendix II, which “includes species not necessarily threatened with extinction, but in which trade must be controlled in order to avoid utilisation incompatible with their survival” (CITES, 2019, p. 2).
Pressure from overfishing appears to be the most significant contributor to shortfin mako shark decline globally. Due to the widespread distribution of I. oxyrinchus and migratory nature, conservation effort and management is needed at both a local and international scale (Adams, Flores, Flores, Aarestrup, & Svendsen, 2016; Corrigan et al., 2018). Because of their “Endangered” status on the with a decreasing population trend, it is essential to increase their population trend and bring them to the next highest, more stable level on the Red List, “Vulnerable”, and take measures to ensure they do not go the other direction and become classified as “Critically Endangered”. The most effective way this can be achieved is through strict management of fisheries and the implementation of Marine Protected Areas (MPA) in habitats that are suitable for shortfin makos by means of international treaties (Birkmanis, Partridge, Simmons, Heupel, & Sequeira, 2020; Dulvy et al., 2008; Rigby et al., 2019).
Community Education & Awareness
Films, documentaries, and TV series are a great way of moving an audience and opening their eyes to what is going on in the world around them in an entertaining way. Saving Jaws by Ocean Ramsey, Shark Week by The Discovery Channel, and BBC’s Blue Planet, narrated by David Attenborough, are a few.
For the booklovers, don’t worry, we got you. Ocean Ramsey’s What You Should Know About Sharks: Shark Language, social behavior, human inter-actions, and life saving information educates readers on shark behaviours, how to swim with them safely, and debunks common shark myths. Sylvia A. Earle’s book The World is Blue: How Our Fate and the Ocean’s are One shows readers how every one of us literally breathes the ocean, and her other book, Sea Change: A Message of the Oceans, has been compared to Rachel Carson’s Silent Spring. For those who do not enjoy reading, Christian Vizl’s Silent Kingdom is a photography-based book that shows the raw beauty, power, and elegance of marine creatures, such as sharks, in stunning black and white photos.
NGO’s also play an essential role in community education and awareness. In New Zealand, groups such as Auckland Whale & Dolphin Safari and Whale Watch Kaikōura take their customers to visit sea life up close. SEA LIFE Kelly Tarlton’s Aquarium educates both adults and children alike and cares for sick and dying turtles.
Monitoring & Research
There is enough research on I. oxyrinchus populations to know something has to change, but what lacks is monitoring the factors contributing to their dismal fate. As of 2019, CITES began to monitor the commercial catch and trade of shortfin mako but agreed that it might not be enough (CITES, 2019). In 2013, MPI released a National Plan of Action for the Conservation and Management of Sharks (NPOA). In the plan, they stated one of their objectives was to “systematically review management categories and protection status to ensure they are appropriate to the status of individual shark species” (MPI, 2013, p. 19) among others, yet there is a lack of reporting available for public access to back this up. They also had intentions of issuing a revised NPOA in 2018, which seems not to exist altogether.
Bycatch records need to be scrutinised and fisheries must be forced to show accurate record-keeping. Data collected from accurate bycatch records of I. oxyrinchus can then be accumulated around the globe to highlight the fisheries that are exploiting shortfin makos.
References
Adams, D. H., Borucinska, J. D., Maillett, K., Whitburn, K., & T. E. Sanders. (2015). Mortality due to a retained circle hook in a longfin mako shark Isurus paucus (Guitart-Manday). Journal of Fish Diseases, 38(7), 621–628. https://doi.org/10.1111/jfd.12277
Adams, G. D., Flores, D., Flores, O. G., Aarestrup, K., & Svendsen, J. C. (2016). Spatial ecology of blue shark and shortfin mako in Southern Peru: local abundance, habitat preferences and implications for conservation. Endangered Species Research, 31, 19–32. doi: 10.3354/esr00744
Barreto, R. R., de Farias, W. K. T., Andrade, H., Santana, F. M., & Lessa, R. (2016). Age, growth and spatial distribution of the life stages of the shortfin mako, Isurus oxyrinchus (Rafinesque 1810) caught in the western and central Atlantic. PLOS One, 11(4). doi:10.1371/journal.pone.0153062
Birkmanis, C. A., Partridge, J. C., Simmons, L. W., Heupel, M. R., & Sequeira, A. M. M. (2020). Shark conservation hindered by lack of habitat protection. Global Ecology and Conservation, 21. https://doi.org/10.1016/j.gecco.2019.e00862
Bishop, S. D. H., Francis, M. P., Duffy, C, & Montgomery, J. C. (2006). Age, growth, maturity, longevity and natural mortality of the shortfin mako shark (Isurus oxyrinchus) in New Zealand waters. Marine and Freshwater Research, 57, 143–154. doi: 10.1071/MF05077
Borucinska, J. D., & Hege, K. (1999). Disseminated granulomas associated with nematode larvae in a shortfin mako shark. Journal of Wildlife Diseases, (35)1, 98–100. https://doi.org/10.7589/0090-3558-35.1.98
Camhi, M. D., Pikitch, E. K., & Babcock, E. A. (2008). Sharks of the open ocean: biology, fisheries, and conservation. Oxford, United Kingdom: Blackwell Publishing Ltd.
Campana, S.E., Joyce, W., Fowler, M. & Showell, M. (2016). Discards, hooking, and post-release mortality of porbeagle (Lamna nasus), shortfin mako (Isurus oxyrinchus), and blue shark (Prionace glauca) in the Canadian pelagic longline fishery. ICES Journal of Marine Science, 73(2), 520–528. https://doi.org/10.1093/icesjms/fsv234
Carey, F. G., Teal, J. M., & Kanwisher, J. W. (1981). The visceral temperatures of mackerel sharks (Lamindae). Physiological and Biochemical Zoology, 54, 334–344. https://doi.org/10.1086/physzool.54.3.30159948
Cema, F., & Licandeo, R. (2009). Age and growth of the shortfin mako (Isurus oxyrinchus) in the south-eastern Pacific off Chile. Marine and Freshwater Research, 60, 394–403
Chan, R. W. K. (2001). Biological studies on sharks caught off the coast of New South Wales (Doctoral thesis, University of New South Wales, Sydney, Australia)
Compagno, L. J. V. (2001). Sharks of the world: an annotated and illustrated catalogue of shark species known to date. Volume 2: bullhead, mackerel and carpet Sharks (Heterodontiformes, Lamniformes and Orectolobiformes). Rome, Italy: FAO.
Corrigan, S., Lowther, A. D., Beheregaray, L. B., Bruce, B. D., Cliff, G., Duffy, C. A., Foulis, A., Francis, M. P., Goldsworthy, S. D., Hyde, J. R., Jabado, R. W., Kacev, D., Marshall, L., Mucientes, G. R., Naylor, G. J. P., Pepperell, J. G., Queiroz, N., White, W. T., Wintner, S. P., & Rogers, P. J. (2018). Population connectivity of the highly migratory shortfin mako (Isurus oxyrinchus Rafinesque 1810) and implications for management in the Southern Hemisphere. Frontiers in Ecology and Evolution, 6. https://doi.org/10.3389/fevo.2018.00187
Donley, J. M., Sepulveda, C. A., Konstantinidis, P., Gemballa, S., & Shadwick, R. E. (2004). Convergent evolution in mechanical design of lamnid sharks and tunas. Nature, 429(6987), 61–65. doi:10.1038/nature02435
Doño F., Montealegre-Quijano, S., Domingo, A., & Kinas, P. G. (2014). Bayesian age and growth analysis of the short-fin mako shark Isurus oxyrinchus (Rafinesque, 1810), in the Western South Atlantic Ocean using a flexible model. Environmental Biology of Fishes, 98, 517–533. Retrieved from https://link.springer.com/article/10.1007/s10641-014-0284-1
Duffy, C., & Francis, M. P. (2001). Evidence of summer parturition in shortfin mako (Isurus oxyrinchus) sharks from New Zealand waters. New Zealand Journal of Marine and Freshwater Research, 35(2), 319–324. https://doi.org/10.1080/00288330.2001.9517001
Dulvy, N. K., Baum, J. K., Clarke, S., Compagno, L. J. V., Cortés, E., Domingo, A … Valenti, S. (2008). You can swim but you can’t hide: the gobal status and conservation of oceanic pelagic sharks and rays. Aquatic Conservation: Marine and Freshwater Ecosystems, 18(5), 459–482. https://doi.org/10.1002/aqc.975
Emery, S. H., & Szczepanski, A. (1986). Gill dimensions in pelagic elasmobranch fishes. Biol Bulletin, 171, 441–449. https://doi.org/10.2307/1541685
Ferretti, F., Myers, R.A., Serena, F. & Lotze, H.K. (2008). Loss of large predatory sharks from the Mediterranean Sea. Conservation Biology, 22, 952–964. https://doi.org/10.1111/j.1523-1739.2008.00938.x
Feretti, F., Worm, B., Britten, G. L., Heithaus, M. R., & Lotze, H. K. (2010). Patterns and ecosystem consequences of shark declines in the ocean. Ecology Letters, 13(8), 1055–1071. doi:10.1111/j.1461-0248.2010.01489.x
Fields, A. T., Fischer, G. A., Shea, S. K. H., Zhang, H., Abercrombie, D. L., Feldheim, K. A., Babcock, E. A. & Chapman, D. D. (2017). Species composition of the international shark fin trade assessed through retail-market survey in Hong Kong. Conservation Biology, 32(2), 376–389. https://doi.org/10.1111/cobi.13043
Francis, M. P. (1998). New Zealand shark fisheries: development, size and management. Marine and Freshwater Research, 49(7), 579-591. https://doi.org/10.1071/MF97076
Gonzalez-Pestana, A., Kouri, C. J., & Velez-Zuazo, X. (2016). Shark fisheries in the South-East Pacific: a 61-year analysis form Peru. F100 Research, 3(164). doi:310.12688/f1000research.4412.2
Hsu, H. H. (2003). Age, growth, and reproduction of shortfin mako, Isurus oxyrinchus in the North-Eestern Pacific (Masters thesis, National Taiwan Ocean University, Keelung, Taiwan)
Martinsohn, J. T. & Muller, M. (1992). Sharks of Otago region. (Unpublished thesis, University of Otago, Dunedin, New Zealand)
Maz-Courrau, A., López-Vera, C., Galván-Magaña, F., Escobar-Sánchez, O., Rosíles-Martínez, R., & Sanujuán-Muñoz, A. (2011). Bioaccumulation and biomagnification of total mercury in four exploited shark species in the Baja California Peninsula, Mexico. Bulletin of Environmental Contamination and Toxicology, 88(2), 129–134. doi:10.1007/s00128-011-0499-1
Mollet, H.F., & Cailliet, G. M. (2002). Comparative population demography of elasmobranchs using life history tables, Leslie matrices and stage-based matrix models. Marine and Freshwater Research, 53(8), 503–516.
Mollet, H. F., Cliff, G., Pratt, H. L. Jr., & Stevens, J. D. (2000). Reproductive biology of the female shortfin mako, Isurus oxyrinchus Ratlnesque, 1810, with comments on the embryonic development of lamnoids. Fishery Bulletin, 98, 299–318. Retrieved from http://www.vliz.be/en/imis?refid=50
Natanson, L. J., Kohler, N. E., Ardizzone, D., Cailliet, G. M., Wintner, S. P., & Mollet, H. F. (2006). Validated age and growth estimates for the shortfin mako, Isurus oxyrinchus, in the North Atlantic Ocean. Environmental Biology of Fishes, 77, 367–383. Retrieved from https://link.springer.com/chapter/10.1007/978-1-4020-5570-6_16
Rigby, C.L., Barreto, R., Carlson, J., Fernando, D., Fordham, S., Francis, M.P., Jabado, R.W., Liu, K.M., Marshall, A., Pacoureau, N., Romanov, E., Sherley, R.B. & Winker, H. (2019). Isurus oxyrinchus, shortfin mako. The IUCN Red List of Threatened Species. http://dx.doi.org/10.2305/IUCN.UK.2019-1.RLTS.T39341A2903170.en
Semba, Y., Nakano, H., & Aoki, I. (2009). Age and growth analysis of the shortfin mako, Isurus oxyrinchus, in the western and central North Pacific Ocean. Environmental Biology of Fishes, 84, 377–391. Retrieved from https://link.springer.com/article/10.1007/s10641-009-9447-x
Stillwell, C. E., & Kohler, N. E. (1982). Food, feeding habits, and estimates of daily ration of the shortfin mako (Isurus oxyrinchus) in the northwest Atlantic. Canadian Journal of Fisheries and Aquatic Sciences, 39(3), 407–414. https://doi.org/10.1139/f82-058
Watling, R. J., McClurg, T. P., Stanton, R. C. (1981). Relation between mercury concentration and size in the mako shark. Bulletin of Environmental Contamination and Toxicology, 26, 352–358. https://doi.org/10.1007/BF01622101
Wegner, N. C., Sepulveda, C. A., Olson K. R., Hyndman, K. A., & Graham, J. B. (2010). Functional morphology of the shortfin mako, Isurus oxyrinchus, a lamnid shark. Journal of Morphology, 271, 937–948. doi:10.1002/jmor.10845
My name is Emily Jupp. I’m a studying Marine Biologist and Geospatial Scientist, and I’ve got some things I’d like to say. So, grab yourself a hot bevvy and buckle up because it’s all underwater from here.
There’s a plethora of information out there, and some of that information is about the ocean. The only real way to figure out the truth about something is to do your own research, which can be really hard when there’s so much research material to choose from. That’s where I come in. I hope to provide you with as much reliable, honest, and up-to-date oceanic information as possible, including references that will help you with your own study.
You don’t need to be a scientist to read about science; all you need is an inquisitive mind and a childlike curiosity about the world that surrounds you.