
Potential of the Endophytic Fungus Phialocephala fortinii Rac56 Found in Rhodiola Plants to Produce Salidroside and p-Tyrosol

Abstract
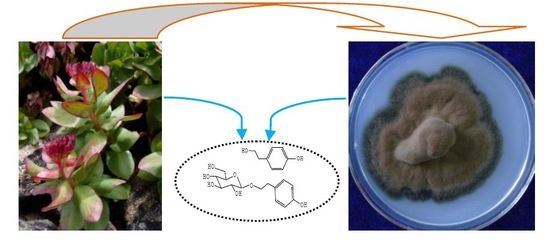
:
1. Introduction
2. Results
2.1. Isolation and Screening of SAL- and TYR-Producing Fungi
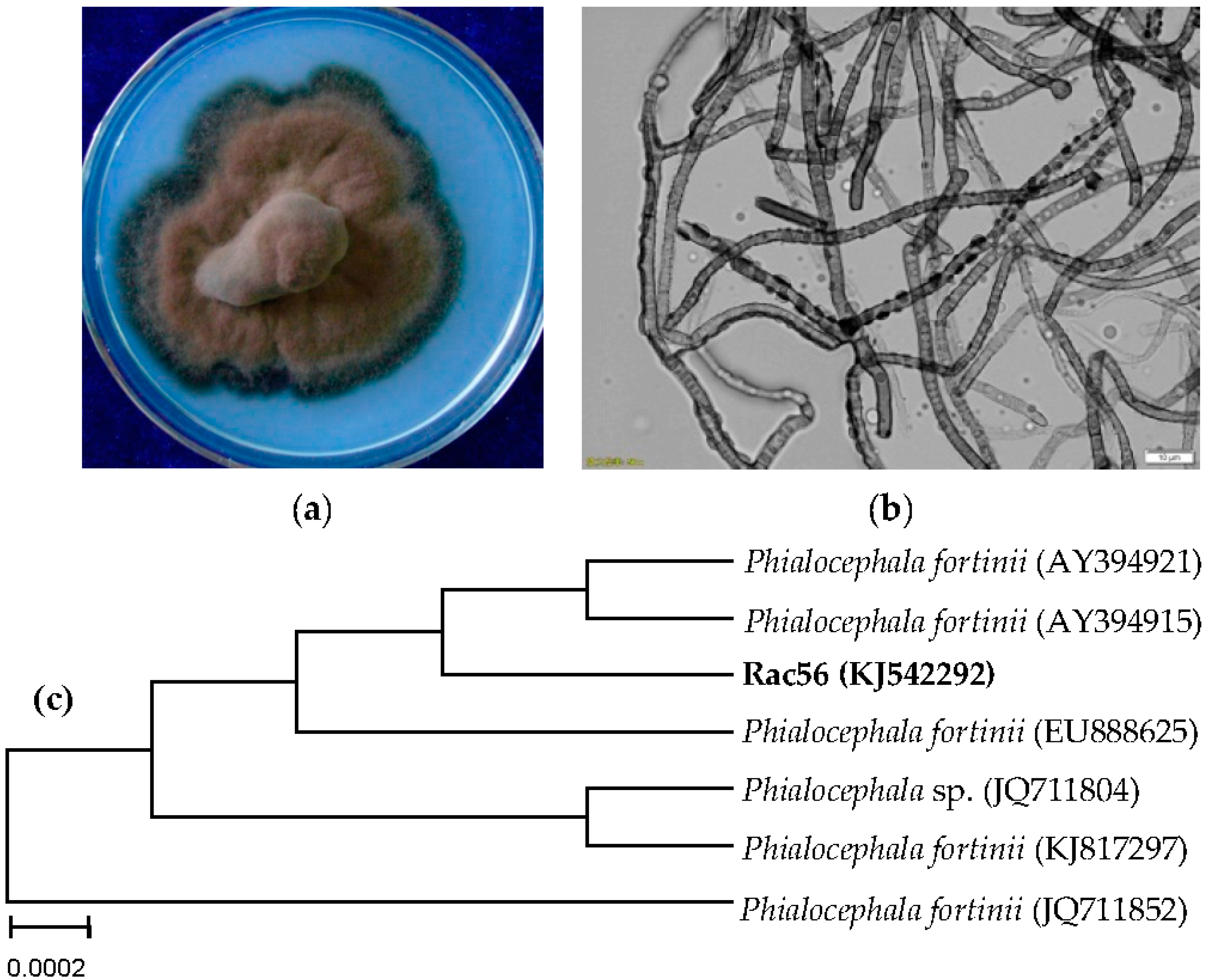
2.2. Identification of Rac56 as the Ideal SAL- and TYR-Producing Fungus
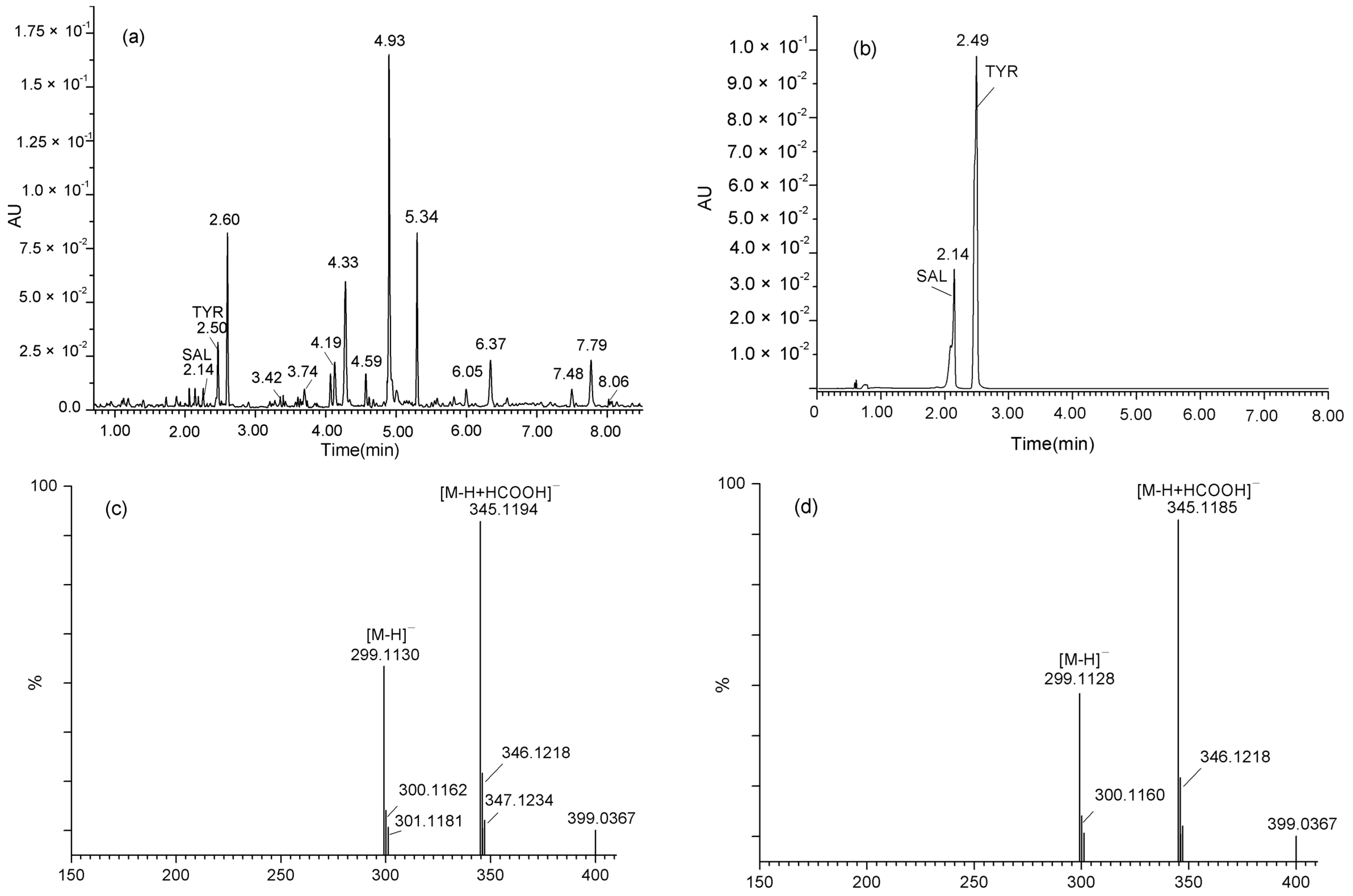
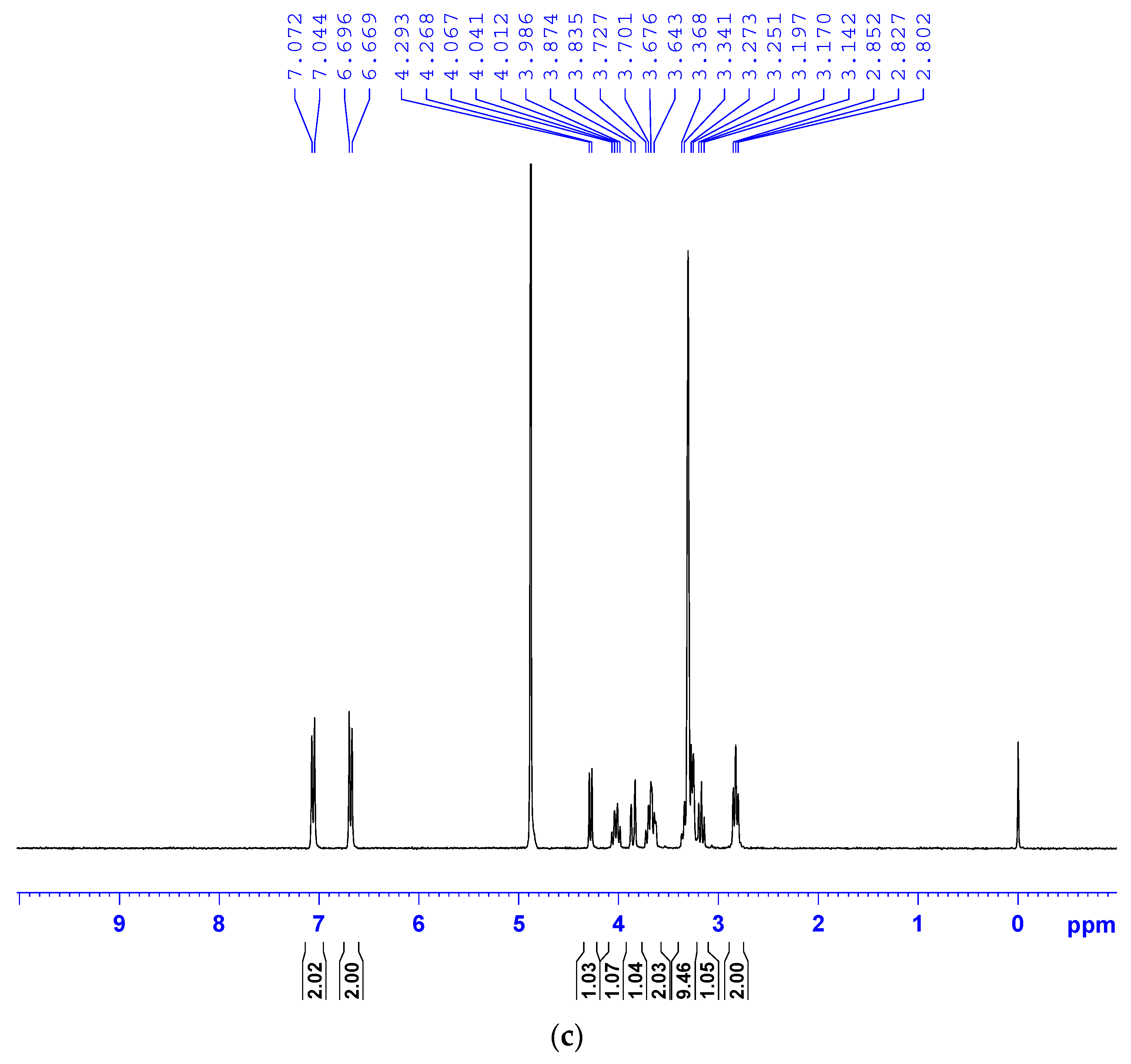
2.3. Validation of SAL and TYR Produced from Rac56 by LC-MS and 1H-NMR
2.4. Fermentation and Optimal Growth Conditioning for SAL and TYR Production from Rac56
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Isolation of Endophytic Fungi
4.2. Fermentation and Secondary Metabolite Extraction Fromendophytic Fungal spp.
4.3. Screening of SAL and TYR-Producing Endophytic Fungi
4.4. Molecular Weight Determination by UPLC/Q-TOF-MS
4.5. Structure Determination of SAL and TYR in Fungal Extracts by 1H-NMR
4.6. Identification of SAL and TYR-Producing Fungus
4.7. Optimization of Fermentation Conditions for SAL and TYR Production from Rac56
4.8. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| SAL | salidroside |
| YAL | p-tyrosol |
| DSE | dark septate endophytes |
| CD | Czapek-Dox medium |
| PD | potato dextrose medium |
| SD | sabouraud medium |
| LBP | lactose-beef extract-peptone medium |
| HPLC | high performance liquid chromography |
| UPLC/Q-TOF-MS | ultra-performance liquid chromatography/quadrupole time–off light mass spectrometry |
| 1H-NMR | 1H nuclear magnetic resonance |
References
- Panossian, A.; Hamm, R.; Wikman, G.; Efferth, T. Mechanism of action of Rhodiola, salidroside, tyrosol and triandrin in isolated neuroglial cells: An interactive pathway analysis of the downstream effects using RNA microarray data. Phytomedicine 2014, 21, 1325–1348. [Google Scholar] [CrossRef] [PubMed]
- Hung, S.K.; Perry, R.; Ernst, E. The effectiveness and efficacy of Rhodiola rosea L.: A systematic review of randomized clinical trials. Phytomedicine 2011, 18, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Chiang, H.M.; Chen, H.C.; Wu, C.S.; Wu, P.Y.; Wen, K.C. Rhodiola plants: Chemistry and biological activity. J. Food Drug Anal. 2015, 23, 359–369. [Google Scholar] [CrossRef]
- Peschel, W.; Prieto, J.M.; Karkour, C.; Williamson, E.M. Effect of provenance, plant part and processing on extract profiles from cultivated European Rhodiola rosea L. for medicinal use. Phytochemistry 2013, 86, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.S.; Ma, L.Q.; Zhang, J.X.; Shi, G.L.; Hu, Y.H.; Wang, Y.N. Characterization of glycosyltransferases responsible for salidroside biosynthesis in Rhodiola sachalinensis. Phytochemistry 2011, 72, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Petrini, O.; Sieber, T.N.; Toti, L.; Viret, O. Ecology, metabolite production, and substrate utilization in endophytic fungi. Nat. Toxins 1992, 1, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.; Daisy, B. Bioprospecting for microbial endophytes and their natural products. Microbiol. Mol. Biol. R. 2003, 67, 491. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.X.; Zou, W.X. Endophytes: A rich source of functional metabolites. Nat. Prod. Rep. 2001, 18, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.H.; Su, Z.Z.; Wang, C.; Kubicek, C.P.; Feng, X.X.; Mao, L.J.; Wang, J.Y.; Chen, C.; Lin, F.C.; Zhang, C.L. The rice endophyte Harpophora oryzae genome reveals evolution from a pathogen to a mutualistic endophyte. Sci. Rep. UK 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.J.; Henson, J.; Van Volkenburgh, E.; Hoy, M.; Wright, L.; Beckwith, F.; Kim, Y.O.; Redman, R.S. Stress tolerance in plants via habitat-adapted symbiosis. Isme J. 2008, 2, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.S.; Zeng, X.; Zhang, D.W.; Guo, S.X. Diverse fungi associated with partial irregular heartwood of Dalbergia odorifera. Sci. Rep. UK 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by taxomyces-andreanae, an endophytic fungus of pacific yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, R.; Ferranti, P.; Caira, S.; Misso, G.; Castellano, M.; di Lorenzo, G.; Caraglia, M. Myrtucommulone production by a strain of Neofusicoccum australe endophytic in myrtle (Myrtus communis). World J. Microbiol. Biotechnol. 2014, 30, 1047–1052. [Google Scholar] [CrossRef] [PubMed]
- El-Hawary, S.S.; Mohammed, R.; AbouZid, S.F.; Bakeer, W.; Ebel, R.; Sayed, A.M.; Rateb, M.E. Solamargine production by a fungal endophyte of Solanum nigrum. J. Appl. Microbiol. 2016, 120, 900–911. [Google Scholar] [CrossRef] [PubMed]
- Addy, H.D.; Hambleton, S.; Currah, R.S. Distribution and molecular characterization of the root endophyte Phialocephala fortinii along an environmental gradient in the boreal forest of Alberta. Mycol. Res. 2000, 104, 1213–1221. [Google Scholar] [CrossRef]
- Brenn, N.; Menkis, A.; Grunig, C.R.; Sieber, T.N.; Holdenrieder, O. Community structure of Phialocephala fortinii s. lat. in European tree nurseries, and assessment of the potential of the seedlings as dissemination vehicles. Mycol. Res. 2008, 112, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Grunig, C.R.; Brunner, P.C.; Duo, A.; Sieber, T.N. Suitability of methods for species recognition in the Phialocephala fortinii-Acephala applanata species complex using DNA analysis. Fungal Genet. Biol. 2007, 44, 773–788. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.J.K.; Wilcox, H.E. New species of ectendomycorrhizal and pseudomycorrhizal fungi—Phialophora-Finlandia, Chloridium-Paucisporum, and Phialocephala-Fortinii. Mycologia 1985, 77, 951–958. [Google Scholar] [CrossRef]
- Wilson, B.J.; Addy, H.D.; Tsuneda, A.; Hambleton, S.; Currah, R.S. Phialocephala sphaeroides sp nov. a new species among the dark septate endophytes from a boreal wetland in Canada. Can. J. Bot. 2004, 82, 607–617. [Google Scholar] [CrossRef]
- Ioset, K.N.; Nyberg, N.T.; Van Diermen, D.; Malnoe, P.; Hostettmann, K.; Shikov, A.N.; Jaroszewski, J.W. Metabolic profiling of Rhodiola rosea rhizomes by H-1 NMR spectroscopy. Phytochem. Anal. 2011, 22, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.H.; Debbab, A.; Proksch, P. Fungal endophytes: Unique plant inhabitants with great promises. Appl. Microbiol. Biotechnol. 2011, 90, 1829–1845. [Google Scholar] [CrossRef] [PubMed]
- Shiono, Y.; Shimanuki, K.; Hiramatsu, F.; Koseki, T.; Tetsuya, M.; Fujisawa, N.; Kimura, K. Pyrrospirones A and B, apoptosis inducers in HL-60 cells, from an endophytic fungus, Neonectria ramulariae Wollenw KS-246. Bioorg. Med. Chem. Lett. 2008, 18, 6050–6053. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.Y. A new species of Lachnum on leaves of Livistona and a key to the Chinese species of the genus. Mycotaxon 2003, 86, 375–382. [Google Scholar]
- Jumpponen, A.; Trappe, J.M. Dark septate endophytes: A review of facultative biotrophic root-colonizing fungi. New Phytol. 1998, 140, 295–310. [Google Scholar] [CrossRef]
- Jumpponen, A.; Trappe, J.M. Performance of Pinus contorta inoculated with two strains of root endophytic fungus, Phialocephala fortinii: Effects of synthesis system and glucose concentration. Can. J. Bot. 1998, 76, 1205–1213. [Google Scholar] [CrossRef]
- Narisawa, K.; Usuki, F.; Hashiba, T. Control of verticillium yellows in Chinese cabbage by the dark septate endophytic fungus LtVB3. Phytopathology 2004, 94, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Bartholdy, B.A.; Berreck, M.; Haselwandter, K. Hydroxamate siderophore synthesis by Phialocephala fortinii, a typical dark septate fungal root endophyte. Biometals 2001, 14, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Addy, H.D.; Piercey, M.M.; Currah, R.S. Microfungal endophytes in roots. Can. J. Bot. 2005, 83, 1–13. [Google Scholar] [CrossRef]
- Zhang, J.X.; Ma, L.Q.; Yu, H.S.; Zhang, H.; Wang, H.T.; Qin, Y.F.; Shi, G.L.; Wang, Y.N. A tyrosine decarboxylase catalyzes the initial reaction of the salidroside biosynthesis pathway in Rhodiola sachalinensis. Plant Cell Rep. 2011, 30, 1443–1453. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.Q.; Liu, B.Y.; Gao, D.Y.; Pang, X.B.; Lu, S.Y.; Yu, H.S.; Wang, H.; Yan, F.; Li, Z.Q.; Li, Y.F.; Ye, H.C. Molecular cloning and overexpression of a novel UDP-glucosyltransferase elevating salidroside levels in Rhodiola sachalinensis. Plant Cell Rep. 2007, 26, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Gyorgy, Z.; Tolonen, A.; Neubauer, P.; Hohtola, A. Enhanced biotransformation capacity of Rhodiola rosea callus cultures for glycosid production. Plant Cell Tiss Org. 2005, 83, 129–135. [Google Scholar] [CrossRef]
- Commission, C.P. Pharmacopoeia of the People’s Republic of China; Chinese Medical Science and Technology Press: Beijing, China, 2010; p. 144. [Google Scholar]
- Sample Availability: Samples of the compoundsare not available from the authors.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Rac12 (mg/mL) | Rct30 (mg/mL) | Rac56 (mg/mL) | Rac63 (mg/mL) | ||||
|---|---|---|---|---|---|---|---|---|
| Fil. Ext. 3 | Myc. Ext. 4 | Fil. Ext. | Myc. Ext. | Fil. Ext. | Myc. Ext. | Fil. Ext. | Myc. Ext. | |
| SAL 1 | - | 0.131 ± 0.009 | - | 0.343 ± 0.465 | 1.729 ± 0.063 | 0.028 ± 0.032 | 0.402 ± 0.248 | - |
| TYR 2 | - | 0.113 ± 0.010 | - | - | 1.990 ± 0.050 | - | 0.083 ± 0.117 | - |
| 1 SAL | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 | 19.298 | 19.086 | 19.471 | 19.1383 | 18.4791 | 2 V | SS | df | S2 | F | p |
| T2 | 24.2256 | 19.8705 | 19.1425 | 18.6516 | 21.8906 | 3 TE | 3.6368 | 3 | 1.2123 | 40.3207 | 0.0001 |
| T3 | 15.607 | 18.4464 | 17.0162 | 18.4303 | 17.2551 | 4 pH | 0.106 | 3 | 0.0353 | 1.1751 | 0.3346 |
| T4 | 16.8242 | 18.5519 | 20.3251 | 19.7346 | 18.33 | 5 MV | 0.4944 | 3 | 0.1648 | 5.4815 | 0.0037 |
| x1 | 1.6082 | 1.5905 | 1.6226 | 1.5949 | 1.5399 | 6 RS | 0.0837 | 3 | 0.0279 | 0.9278 | 0.4386 |
| x2 | 2.0188 | 1.6559 | 1.5952 | 1.5543 | 1.8242 | 7 FT | 1.01 | 3 | 0.3367 | 11.1977 | 0.0001 |
| x3 | 1.3006 | 1.5372 | 1.418 | 1.5359 | 1.4379 | error | 0.9621 | 32 | 0.0301 | ||
| x4 | 1.402 | 1.546 | 1.6938 | 1.6446 | 1.5275 | F0.05 | 2.90 | ||||
| R | 0.7182 | 0.1187 | 0.2757 | 0.1087 | 0.3863 | F0.01 | 4.46 | ||||
| 1 TYR | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 | 13.7101 | 15.9705 | 15.8391 | 15.244 | 13.4073 | 2 V | SS | df | S2 | F | p |
| T2 | 19.664 | 16.223 | 15.9399 | 14.6122 | 13.4654 | 3 TE | 2.1205 | 3 | 0.7068 | 21.3276 | 0.0001 |
| T3 | 15.1039 | 14.2837 | 14.0013 | 16.6605 | 17.3406 | 4 pH | 0.1871 | 3 | 0.0624 | 1.8822 | 0.1525 |
| T4 | 13.319 | 15.3198 | 16.0167 | 15.2803 | 17.5837 | 5 MV | 0.2343 | 3 | 0.0781 | 2.3563 | 0.0903 |
| x1 | 1.1425 | 1.3309 | 1.3199 | 1.2703 | 1.1173 | 6 RS | 0.1865 | 3 | 0.0622 | 1.8762 | 0.1535 |
| x2 | 1.6387 | 1.3519 | 1.3283 | 1.2177 | 1.1221 | 7 FT | 1.3532 | 3 | 0.4511 | 13.6102 | 0.0001 |
| x3 | 1.2587 | 1.1903 | 1.1668 | 1.3884 | 1.4451 | error | 1.0605 | 32 | 0.0331 | ||
| x4 | 1.1099 | 1.2767 | 1.3347 | 1.2734 | 1.4653 | F0.05 | 2.90 | ||||
| R | 0.5288 | 0.1616 | 0.1679 | 0.1707 | 0.3480 | F0.01 | 4.46 | ||||
| Codes | Factors | Content (mg/mL) | |||||
|---|---|---|---|---|---|---|---|
| TE 1 | pH 2 | MV 3 | RS 4 | FT 5 | SAL 6 | TYR 7 | |
| 1 | 20 | 5 | 25 | 100 | 3 | 1.6264 ± 0.1482 | 1.0312 ± 0.0071 |
| 2 | 20 | 6 | 50 | 150 | 7 | 1.9082 ± 0.0720 | 1.0128 ± 0.0069 |
| 3 | 20 | 7 | 100 | 200 | 11 | 1.2076 ± 0.1359 | 1.1833 ± 0.0524 |
| 4 | 20 | 8 | 150 | 250 | 15 | 1.6904 ± 0.0822 | 1.3428 ± 0.2233 |
| 5 | 25 | 5 | 50 | 200 | 15 | 1.9383 ± 0.0475 | 2.0018 ± 0.0009 |
| 6 | 25 | 6 | 25 | 250 | 11 | 2.0502 ± 0.0477 | 1.8792 ± 0.0860 |
| 7 | 25 | 7 | 150 | 100 | 7 | 2.3392 ± 0.1093 | 1.4064 ± 0.1196 |
| 8 | 25 | 8 | 100 | 150 | 3 | 1.7475 ± 0.0789 | 1.2673 ± 0.0111 |
| 9 | 30 | 5 | 100 | 250 | 7 | 1.4483 ± 0.1551 | 1.0020 ± 0.0014 |
| 10 | 30 | 6 | 150 | 200 | 3 | 1.3964 ± 0.0129 | 1.3012 ± 0.1997 |
| 11 | 30 | 7 | 25 | 150 | 15 | 1.2126 ± 0.1286 | 1.3021 ± 0.2278 |
| 12 | 30 | 8 | 50 | 100 | 11 | 1.1450 ± 0.1473 | 1.4293 ± 0.2049 |
| 13 | 35 | 5 | 150 | 150 | 11 | 1.3489 ± 0.2708 | 1.2885 ± 0.1504 |
| 14 | 35 | 6 | 100 | 100 | 15 | 1.2687 ± 0.1773 | 1.2145 ± 0.1107 |
| 15 | 35 | 7 | 50 | 250 | 3 | 1.3894 ± 0.0478 | 0.8694 ± 0.0961 |
| 16 | 35 | 8 | 25 | 200 | 7 | 1.6011 ± 0.0927 | 1.0672 ± 0.0888 |
| Factor | Levels | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| Temperature (°C) | 20 | 25 | 30 | 35 |
| Initial pH | 5 | 6 | 7 | 8 |
| Medium volume (mL) | 25 | 50 | 100 | 150 |
| Rotation speed (rpm) | 100 | 150 | 200 | 250 |
| Fermentation time (day) | 3 | 7 | 11 | 15 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, J.; Guo, T.; Chao, J.; Wang, M.; Wang, J. Potential of the Endophytic Fungus Phialocephala fortinii Rac56 Found in Rhodiola Plants to Produce Salidroside and p-Tyrosol. Molecules 2016, 21, 502. https://doi.org/10.3390/molecules21040502
Cui J, Guo T, Chao J, Wang M, Wang J. Potential of the Endophytic Fungus Phialocephala fortinii Rac56 Found in Rhodiola Plants to Produce Salidroside and p-Tyrosol. Molecules. 2016; 21(4):502. https://doi.org/10.3390/molecules21040502
Chicago/Turabian StyleCui, Jinlong, Tingting Guo, Jianbin Chao, Mengliang Wang, and Junhong Wang. 2016. "Potential of the Endophytic Fungus Phialocephala fortinii Rac56 Found in Rhodiola Plants to Produce Salidroside and p-Tyrosol" Molecules 21, no. 4: 502. https://doi.org/10.3390/molecules21040502