
New Records of the Psychrophilic Tetracladium Species Isolated from Freshwater Environments in Korea

1
Fungal Resources Research Division, Freshwater Bioresources Research Department, Nakdonggang National Institute of Biological Resources, 137 Donam 2-gil, Sangju-si 37242, Korea
2
Wildlife Disease Research Team, National Institute of Wildlife Disease Control and Prevention(NIWDC), Ministry of Environment, 1 Songam-gil, Gwangsan-gu, Gwangju 62407, Korea
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(10), 789; https://doi.org/10.3390/d14100789
Submission received: 29 August 2022
/
Revised: 18 September 2022
/
Accepted: 20 September 2022
/
Published: 23 September 2022
Abstract
:We investigated the diversity of aquatic fungi from freshwater environments in Korea. From the survey of aquatic fungi, three Tetracladium (Leotiomycetes, Helotiales) strains, NNIBRFG814, NNIBRFG1997, and NNIBRFG3891, were isolated from decaying leaves of Fraxinus rhynchophylla located in Taebaek-si, Gangwon-do; freshwater foam in the freshwater of the Gam stream in Gimcheon-si; and from filtered freshwater of the Yangsan stream in Mungyeong-si, Gyeongsangbuk-do, respectively. The isolated fungal strains were identified using ITS sequence analysis and morphological characters. A sequence analysis indicated that the fungal isolates were closely related to Tetracladium breve MK371730 (100%, NNIBRFG814), Tetracladium setigerum KU519120 (98.96%, NNIBRFG1997), and T. marchalianum MK353126 (98.14%, NNIBRFG3891). In terms of growth conditions, it was confirmed that NNIBRFG814 grew best at 20 °C, did not grow above 30 °C, and grew at 5 °C as a psychrophilic fungus. NNIBRFG814 was included in the T. breve clade but its morphology was not similar to that of T. breve. Thus, NNIBRFG814 is a new species, and T. fraxineum sp. nov. NNIBRFG1997 and NNIBRFG3891 are the first records in Korea of T. setigerum and T. marchalianum, respectively.
1. Introduction
Aquatic hyphomycetes are common fungi that play key roles in the decomposition of plant matter in freshwater [1]. Ingold was the first to describe them, documenting a collection of new fungal species with aquatic life cycles [2]. The classification and identification of aquatic hyphomycetes are based on their morphology and development, and, in particular, on conidia (asexual spores) [3]. This class of fungi predominantly produces branched multiradiate or sigmoid conidia, which can easily attach to leaves or other smooth surfaces [4]. Although many aquatic hyphomycetes share morphological characters, they are not a monophyletic group, but belong to a polyphyletic taxon [5]. In a previous study in Korea, six aquatic hyphomycetes were isolated from plant litter [6,7].
Tetracladium De Wild. (Leotiomycetes, Helotiales) was the first aquatic hyphomycete genus to be described [2]. To date, eleven Tetracladium species have been reported from different continents [8,9], including T. apiense R.C. Sinclair & Eicker, T. breve A. Roldán, T. ellipsoideum M.M. Wang & Xing Z. Liu, T. furcatum Descals, T. globosum M.M. Wang & Xing Z. Liu, T. marchalianum De Wild., T. maxilliforme (Rostr.) Ingold, T. nainitalense (Sati & Arya), T. palmatum A. Roldán, T. psychrophilum M.M. Wang & Xing Z. Liu, and T. setigerum (Grove) Ingold [2,9,10,11,12,13]. The type species of the Tetracladium genus, T. marchalianum, was first reported in nature by de Wildeman [2]. Since Ingold described T. setigerum as an aquatic hyphomycete in 1942, the majority of Tetracladium species have been found in freshwater environments, although some have been found in terrestrial environments as well [14]. Most Tetracladium species produce tetracladiate conidia with several knobs or finger-like processes that are a unique character distinguishing this genus from other aquatic hyphomycetes [15].
In this study, we isolated three Tetracladium species from freshwater environments in Korea. Through a molecular phylogenetic analysis and morphological characterization, the three fungal strains were identified as T. fraxineum, T. setigerum, and T. marchalianum. Here, we present mycological descriptions of the three Tetracladium species.
2. Materials and Methods
2.1. Isolation of Fungal Strains
We collected decaying leaf deposits and freshwater foam from streams. The decaying leaves were washed with distilled water (DW) and incubated in a pretreatment liquid medium (0.05% 3-morpholinopropane-1-sulfonic acid (weight/volume (w/v), 0.05% KNO3 (w/v), 0.025% KH2PO4 (w/v), and 0.025% K2HPO4 (w/v)) at 20 °C overnight. The incubated supernatant, 100 µL, was spread on water agar (WA) (20 g/L) after 5 to 10 days. Hyphal tips and germinated conidia were detached under a microscope and transferred onto 24-well plates containing V8 agar (V8A) (8% V8 juice (v/v) and 1.5% agar (w/v) adjusted to pH 6.0 using 10 N NaOH). Then, the colonies were transferred onto potato dextrose agar (PDA) (Difco; BD, Franklin Lakes, NJ, USA) for pure culture. All strains used in this study were grown on PDA at 25 °C.
2.2. Induction of Sporulation and Investigation of Culture Conditions
For sporulation, the three Tetracladium species were incubated in sporulation medium (0.01% CaCl3∙2H2O [w/v], 0.001% MgSO4∙7H2O [w/v], 0.001% KNO3 [w/v], 0.055% K2HPO4 [w/v], 0.05% MOPs buffer [w/v]) [16], at 20 °C, for 10 days. The strains were observed using a model Eclipse Ni-U microscope (Nikon, Tokyo, Japan).
To study the optimal growth conditions, the strains were cultivated on PDA, MEA (malt extract 2%; agar 2%/L), oatmeal agar (OA) ( Difco; BD, Franklin Lakes, NJ, USA), and yeast extract peptone dextrose agar (YPDA) (Duchefa Biochemie, Haarlem, The Netherlands), at 5, 15, 20, 25, 30, and 35 °C.
2.3. DNA Extraction and Phylogenetic Analysis
Fungal genomic DNA was isolated by means of a NucleoSpin Plant II DNA extraction kit (Macherey-Nagel, Duren, Germany). To identify the fungi, an internal transcribed spacer (ITS) region was amplified using primers ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) [17]. DNA Homology searches were carried out by BLAST algorithms from the National Center for Biotechnology Information (NCBI). For the phylogenetic analysis, the MEGAX software was used [18]. A phylogenetic tree was constructed using a maximum parsimony (MP) analysis, the maximum likelihood (ML) method, and the neighbor-joining (NJ) method with 1000 bootstrap replications. The MP tree was obtained using the tree-bisection-regrafting (TBR) algorithm [19] with search level 1, in which the initial trees were obtained by the random addition of sequences (10 replicates). This analysis involved 36 nucleotide sequences. The final dataset contained 430 positions in total. The evolutionary distances were calculated using the Kimura 2-parameter method [20] and were in units of the number of base substitutions per site using the ML and NJ methods. The rate of variation among sites was modeled with a gamma distribution (shape parameter = 1). This analysis involved 36 nucleotide sequences. All positions with less than 95% site coverage were eliminated, i.e., fewer than 5% alignment gaps, missing data, or ambiguous bases were allowed at any position (partial deletion option). There was a total of 394 positions in the final dataset. Reference sequences of other fungi were obtained from GenBank at NCBI (Table 1).
3. Results
3.1. Phylogeny
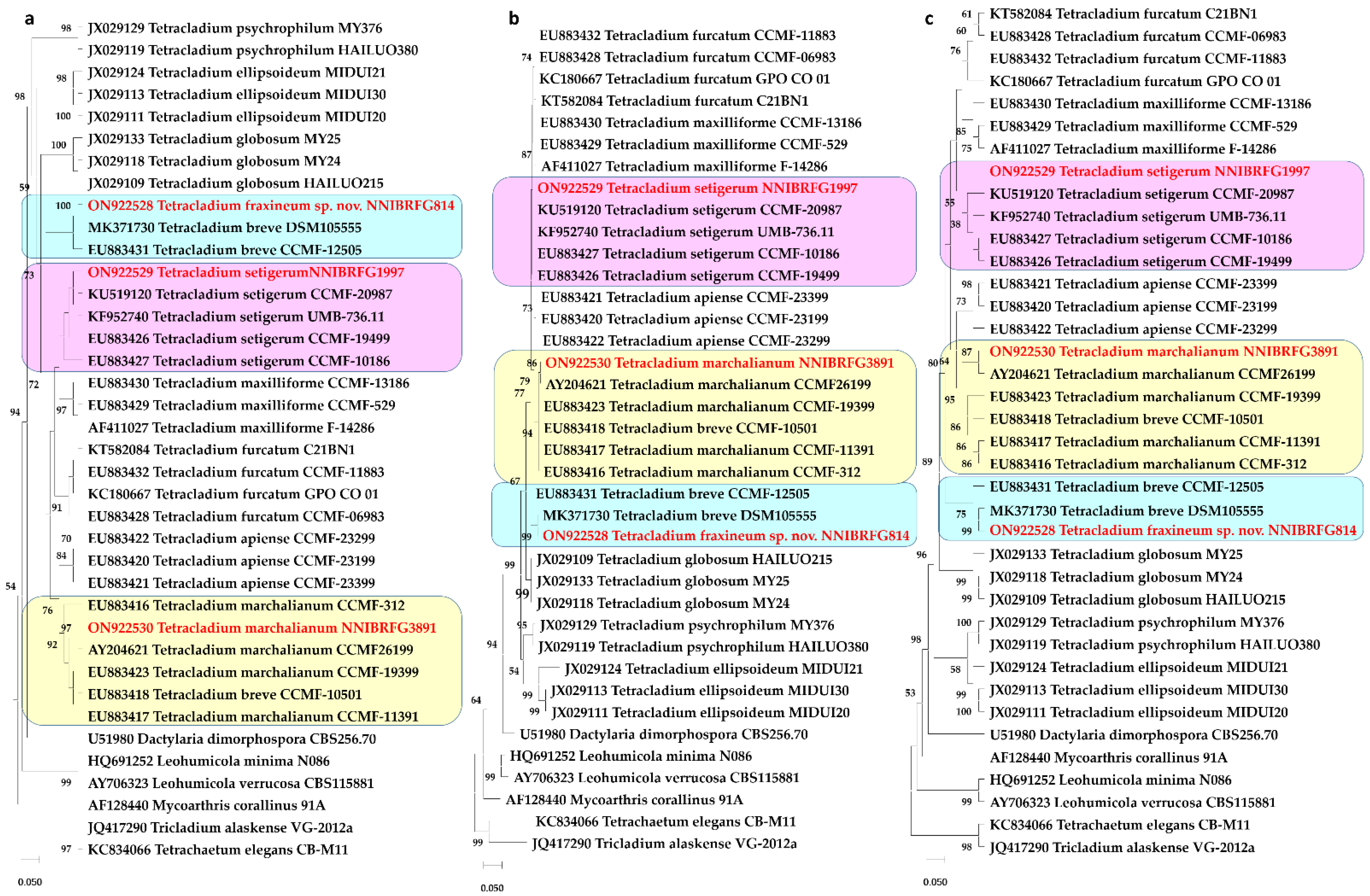
A BLASTn (Nucleotide BLAST) search of the ITS region of NNIBRFG814, NNIBRFG1997, and NNIBRFG3891 revealed similarities of 100% (513/513), 98.96% (476/481), and 98.14% (475/484) with T. breve (MK371730), T. setigerum (KU519120), T. marchalianum (MK353126), respectively. A phylogenetic analysis was performed to identify three fungal strains, and this analysis involved 36 nucleotide sequences. There was a total of 394 positions in the final dataset. NNIBRFG1997 and NNIBRFG3891 were placed in clades with T. setigerum and T. marchalianum, respectively. NNIBRFG814 was placed in a clade with T. breve on the MP and NJ trees, but not in any clade on the ML tree. The estimated value of the shape parameter for the discrete gamma distribution was 0.4286. Substitution patterns and rates were estimated under the Kimura [20] model (+G). The nucleotide frequencies were A = 25.00%, T/U = 25.00%, C = 25.00%, and G = 25.00%. The maximum log-likelihood for this computation was −2584.030.
Figure 1.
Phylogenetic analysis using: (a) the maximum parsimony method; (b) maximum likelihood method; (c) neighbor-joining method, based on the ITS region of three Tetracladium species. This analysis involved 36 nucleotide sequences. Tetrachaetum elegans and Tricladium alaskense were used as outgroups. At branches, bootstrap values of more than 50% (1000 replications) were shown. The unrecorded strains are shown in bold and red.
Figure 1.
Phylogenetic analysis using: (a) the maximum parsimony method; (b) maximum likelihood method; (c) neighbor-joining method, based on the ITS region of three Tetracladium species. This analysis involved 36 nucleotide sequences. Tetrachaetum elegans and Tricladium alaskense were used as outgroups. At branches, bootstrap values of more than 50% (1000 replications) were shown. The unrecorded strains are shown in bold and red.

3.2. Taxonomy
3.2.1. Tetracladium fraxineum sp. nov. J. Goh & H.Y. Mun (2022)
Etymology: Referring to the host genus Fraxinus
Mycobank No.: MB827330
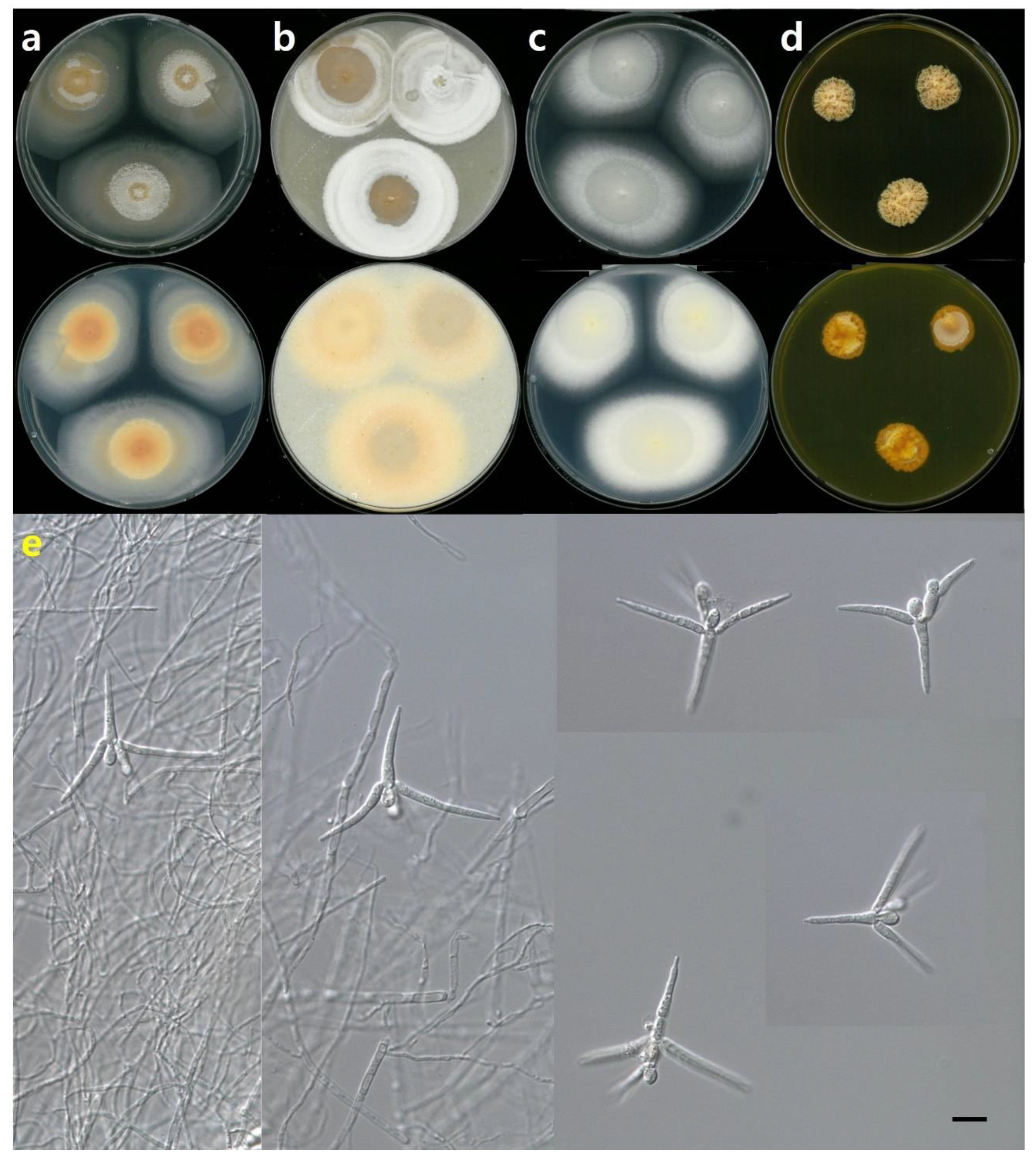
Description: Colonies on PDA, with a diameter of 58–60 mm after 14 days at 20 °C, covered by aerial mycelia, flat floccose white to pale pink, floccose white concentric ring, margin regular, reverse white, and apricot near the center. Colonies on OA, with a diameter of 40–46 mm after 14 days at 20 °C, covered by cottony aerial mycelia, flat floccose pale pink concentric ring, white margin regular, and reverse apricot. Colonies on MEA, with a diameter of 56–58 mm after 14 days at 20 °C, covered by white floccose aerial mycelia, a white concentric ring, margin filiform, reverse white, and a light yellow near the center. Colonies on YPDA, with a diameter of 16–18 mm after 14 days at 20 °C, flat floccose aerial mycelia, umbonate, margin undulate, dense, front, and reverse apricot. Sporulation when submerged. Conidia were three digitiform 20–32 μm and short 1–3 septate, distal cell globose to ellipsoid 6–8 μm long × 4–5 μm width, primary branches at one level, immediately below the globose cell, typically 1-septate, acicular secondary branches were inserted below the second globose cell, and straight and slightly curved branches (Figure 1, Figure 2 and Figure 3).
Habitat: Decaying leaves of Fraxinus rhynchophylla from freshwater
Specimen examined: Taebaek-si, Gangwon-do, Republic of Korea, 3 February 2016, NNIBRFG814, Nakdonggang National Institute of Biological Resources (Sangju-si, Gyeongsangbuk-do, Republic of Korea)
Note: Based on the MP and NJ trees, this strain was in a clade with T. breve. Sequence similarity to T. breve was very high, 100% (MK371730) (Figure 1), but morphology was similar to T. marchlianum. Conidia were found which were very similar to those of T. marchalianum but consistently much smaller and shorter (Figure 2). Therefore, NNIBRFG814 was named T. fraxineum, as a new species.
NNIBRFG814 grew very slowly, approximately 2.5 mm/day, which was the best growth on OA at 20 °C. NNIBRFG814 grew at 5 °C as a psychrophilic fungus, but did not grow at 30 °C (Figure 3).
3.2.2. Tetracladium setigerum (Grove) Ingold
Basionym: Tetracladium setigerum (Grove) Ingold, Transactions of the British Mycological Society 25 (4): 369 (1942)
Mycobank No.: MB291361
Description: Colonies on PDA were 14–18 mm in diameter after 14 days at 20 °C, covered by flat floccose aerial mycelia, yellow, irregular sulcate, margin undulate, umbonate elevation, and reverse yellow. Colonies on OA, 25–34 mm after 14 days at 20 °C, covered by floccose aerial mycelia, white to yellow, with a white concentric ring, margin regular and reverse apricot. Colonies on MEA, 33–36 mm after 14 days at 20 °C, covered by aerial mycelia, white to hyaline, white concentric ring, and reverse white to hyaline, white near the center. Colonies on YPDA, after 14 days at 20 °C, covered by hyaline aerial mycelia. The strain was not formed into a colony but grew into hyaline mycelia. Sporulation when submerged. Conidiophores were semi-macronematous and typically simple. Conidia were two or three digitiforms, 30–80 μm long, and three narrow obclavate elements, 12–20 μm long and 4–8 μm wide, with the upper digitiform having primary branches arising at two levels, curved adaxially, 3 or 5 septate, long and pointed branches (Figure 1 and Figure 4).
Habitat: Freshwater foam in a stream
Specimen examined: Gam cheon, Gimcheon-si, Gyeonsangbuk-do, Republic of Korea, 23 March 2016, NNIBRFG1997, Nakdonggang National Institute of Biological Resources (Sangju-si, Gyeongsangbuk-do, Republic of Korea)
Note: Based on the phylogenetic tree, this strain was in the T. setigerum clade. Similarity was very low, i.e., 98.96% (KU519120), but the morphology was similar to T. setigerum. As a result, NNIBRFG1997 was considered to be T. setigerum.
3.2.3. Tetracladium marchalianum De Wild
Basionym: Tetracladium marchalianum De Wild., Annales de la Societé Belge de Microscopie 17: 39 (1893)
Mycobank No.: MB243554
Description: Colonies on PDA, 45–47 mm in diameter after 14 days at 20 °C, covered by floccose aerial mycelia, white to pale apricot, pale apricot concentric ring, irregular sulcate, white margin filiform, and reverse apricot. After 14 days at 20 °C, colonies on OA, 37–43 mm in diameter, were covered by floccose aerial mycelia, pale yellow mixed white, pale yellow concentric ring, regular margin, and reverse mustard. Colonies on MEA were 28–35 mm in diameter after 14 days at 20 °C, covered by flat floccose near the center, pale apricot to white, floccose margin filiform, and reverse pale apricot. Colonies on YPDA were 30–32 mm in diameter after 14 days at 20 °C, floccose aerial mycelia, light yellow to pale yellow, irregular sulcate, and reverse mustard. Sporulation when submerged. Conidia had an axis of 1–3 septate and a length of 15–25 μm, three or four digitiforms, 1–3 septate, and a length of 25–40 μm. The distal cell was bicellular and constricted at the septum. Globose to ellipsoid was 9–10 μm long and 4–6 μm wide, primary branches at one level were immediately below the globose cell, acicular secondary branch was inserted below the second globose cell, and branches were a little curved (Figure 1 and Figure 5).
Habitat: Filtered freshwater in stream
Specimen examined: Yangsan cheon, Mungyeon-si, Gyeonsangbuk-do, Republic of Korea, 24 February 2017, NNIBRFG3891, Nakdonggang National Institute of Biological Resources (Sangju-si, Gyeongsangbuk-do, Republic of Korea)
Note: Based on the phylogenetic tree, this strain was in the T. marchalianum clade. Similarity was very low, i.e., 98.14% (MK353126), but morphology was similar to T. marchlianum. As a result, NNIBRFG3891 was considered to be T. marchalianum.
4. Discussion
The morphology of T. breve was three long and thin digitiform elements, two narrow obclavate, filiform elements on a stalk, similar to T. setigerum [15]. In contrast, NNIBRFG814 had three short digitiform, globose distal cells and two level branches similar to those of T. marchalianum. T. setigerum was similar to T. breve, but it had three narrow obclavate elements [15]. The morphology of NNIBRFG1997 included three narrow obclavate elements, similar to T. setigerum. Three species, T. fraxineum NNIBRFG814, T. stigerum NNIBRFG1997, and T. marchalianum NNIBRFG 3891, could not produce conidia on solid media, but could produce conidia and conidiophores in liquid media at 20 °C. Almost Ingoldian fungi could not produce conidia in solid media, but they could form conidia when they were completely submerged in water [2].
We collected Tetracladium species from decaying leaves, freshwater foam, and filtered freshwater in late winter. Wang et al. studied cold-adapted fungi and reported three Tetracladium species, which grew well at temperatures below 20 °C. They collected Tetracladium species from soil on a glacier [13]. Our strains were collected from freshwater, in cold weather in Feburary and March, at temperatures under 0 °C. Our three strains grew well below 20 °C as psychrophilic fungi. Czeczuga and Orlowska investigated the aquatic hyphomycetes diversity in melting snow water, rain, and ice. According to them, as the water temperature decreased, the increasing surface tension of the upper layer seemed to pull various aquatic fungi. They found most species in their study from melting snow water [21]. For effective isolation of aquatic hyphomycetes including Tetracladium, we would consider sampling in freshwaters during cold weather.
Author Contributions
H.Y.M., conceptualization, identification, illustrations editing, writing the orginal draft, editing, and correspondence; J.G., conceptualization, collection, identification, editing, assistance in reviewing, funding; Y.J.J., methodology, illustrations editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the Nakdonggang National Institute of Biological Resources (NNIBR), funded by the Ministry of Environment (MOE) of the Republic of Korea (NNIBR202201107).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The types were deposited in the Fungal research team, Nakdonggang National Institute of Biological Resources, Korea.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Krauss, G.J.; Solé, M.; Krauss, G.; Schlosser, D.; Wesenberg, D.; Bärlocher, F. Fungi in freshwaters: Ecology, physiology and biochemical potential. FEMS Microbiol. Rev. 2011, 35, 620–651. [Google Scholar] [CrossRef] [PubMed]
- Ingold, C.T. Aquatic Hyphomycetes of decaying alder leaves. Trans. Br. Mycol. Soc. 1942, 25, 339–417, IN1–IN6. [Google Scholar] [CrossRef]
- Seena, S.; Pascoal, C.; Marvanová, L.; Cássio, F. DNA barcoding of fungi: A case study using ITS sequences for identifying aquatic hyphomycete species. Fungal Divers. 2010, 44, 77–87. [Google Scholar] [CrossRef]
- Bärlocher, F.; Seena, S.; Wilson, K.P.; Dudley Williams, D.D. Raised water temperature lowers diversity of hyporheic aquatic Hyphomycetes. Freshw. Biol. 2008, 53, 368–379. [Google Scholar] [CrossRef]
- Belliveau, M.J.-R.; Bärlocher, F. Molecular evidence confirms multiple origins of aquatic Hyphomycetes. Mycol. Res. 2005, 109, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- Mun, H.Y.; Goh, J.; Oh, Y.; Chung, N. New records of three aquatic fungi isolated from freshwater in Samcheok and Yeongju, Korea. Korean J. Mycol. 2016, 44, 247–251. [Google Scholar]
- Mun, H.Y.; Oh, Y.; Goh, J.; Chung, N. First report of three Filosporella species isolated from freshwater ecosystem in Korea. Korean J. Mycol. 2019, 47, 165–172. [Google Scholar]
- Ranzoni, F.V. Aquatic Hyphomycetes from Hawaii. Mycologia 1979, 71, 786–795. [Google Scholar] [CrossRef]
- Sinclair, R.C.; Eicker, A. Tetracladium apiense, a new aquatic species from South Africa. Trans. Br. Mycol. Soc. 1981, 76, 515–517. [Google Scholar] [CrossRef]
- Descals, E.; Webster, J. Four new staurosporous Hyphomycetes from mountain streams. Trans. Br. Mycol. Soc. 1983, 80, 67–75. [Google Scholar] [CrossRef]
- Roldán, A.; Descals, E.; Honrubia, M. Pure culture studies on Tetracladium. Mycol. Res. 1989, 93, 452–465. [Google Scholar] [CrossRef]
- Sati, S.C.; Arya, P.; Belwal, M. Tetracladium nainitalense sp. nov., a root endophyte from Kumaun Himalaya, India. Mycologia 2009, 101, 692–695. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jiang, X.; Wu, W.; Hao, Y.; Su, Y.; Cai, L.; Xiang, M.; Liu, X. Psychrophilic fungi from the world’s roof. Pers. Mol. Phylogeny Evol. Fungi 2015, 34, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Bråthen, I. The aquatic, stauroconidial Hyphomycetes of Norway with notes on the Nordic species. Nord. J. Bot. 1984, 4, 375–392, IN12–IN13. [Google Scholar] [CrossRef]
- Letourneau, A.; Seena, S.; Marvanová, L.; Bärlocher, F. Potential use of barcoding to identify aquatic Hyphomycetes. Fungal Divers. 2010, 40, 51–64. [Google Scholar] [CrossRef]
- Chauvet, E.; Suberkropp, K. Temperature and sporulation of aquatic Hyphomycetes. Appl. Environ. Microbiol. 1998, 64, 1522–1525. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Czeczuga, B.; Orlowska, M. Hyphomycetes in rain water, melting, snow and ice. Acta Mycol. 1999, 34, 181–200. [Google Scholar] [CrossRef]
Figure 2.
Colony shape and conidia of Tetracladium fraxineum NNIBRFG814. Colony shape of front and back on: (a) PDA; (b) OA; (c) MEA; (d) YPDA; (e) conidia, on SM media, 400× (size bar = 10 µm).
Figure 2.
Colony shape and conidia of Tetracladium fraxineum NNIBRFG814. Colony shape of front and back on: (a) PDA; (b) OA; (c) MEA; (d) YPDA; (e) conidia, on SM media, 400× (size bar = 10 µm).

Figure 3.
The growth rate of NNIBRFG814 depending on four media and six temperature.

Figure 4.
Colony shape and conidia of Tetracladium setigerum NNIBRFG1997 Colony shape of front and back on: (a) PDA; (b) OA; (c) MEA; (d) YPDA; (e) conidia, on SM media, 400× (size bar = 10 µm). Red arrows point at conidiophores.
Figure 4.
Colony shape and conidia of Tetracladium setigerum NNIBRFG1997 Colony shape of front and back on: (a) PDA; (b) OA; (c) MEA; (d) YPDA; (e) conidia, on SM media, 400× (size bar = 10 µm). Red arrows point at conidiophores.

Figure 5.
Colony shape and conidia of Tetracladium marchalianum NNIBRFG3891: Colony shape of front and back on: (a) PDA; (b) OA; (c) MEA; (d) YPDA; (e) conidia, on SM media, 400× (size bar = 10 µm).
Figure 5.
Colony shape and conidia of Tetracladium marchalianum NNIBRFG3891: Colony shape of front and back on: (a) PDA; (b) OA; (c) MEA; (d) YPDA; (e) conidia, on SM media, 400× (size bar = 10 µm).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Reference sequences of strains from GenBank at the NCBI.
| Species | Strain | Accession Number |
|---|---|---|
| ITS | ||
| Dactylaria dimorphospora | CBS 256.70 | U51980 |
| Leohumicola minima | N086 | HQ691252 |
| Leohumicola verrucosa | CBS 115881 | AY706323 |
| Mycoarthris corallinus | 91A | AF128440 |
| Tetrachaetum elegans | CB-M11 | KC834066 |
| Tetracladium apiense | CCM F-23199 | EU883420 |
| Tetracladium apiense | CCM F-23299 | EU883422 |
| Tetracladium apiense | CCM F-23399 | EU883421 |
| Tetracladium breve | CCM F-12505 | EU883431 |
| Tetracladium ellipsoideum | MIDUI20 | JX029111 |
| Tetracladium ellipsoideum | MIDUI21 | JX029124 |
| Tetracladium ellipsoideum | MIDUI30 | JX029113 |
| Tetracladium fraxineum sp. nov. | NNIBRFG814 | ON922528 |
| Tetracladium furcatum | C21BN1 | KT582084 |
| Tetracladium furcatum | CCM F-06983 | EU883428 |
| Tetracladium furcatum | CCM F-11883 | EU883432 |
| Tetracladium furcatum | GPO CO 01 | KC180667 |
| Tetracladium globosum | HAILUO215 | JX029109 |
| Tetracladium globosum | MY24 | JX029118 |
| Tetracladium globosum | MY25 | JX029133 |
| Tetracladium marchalianum | CCM F-11391 | EU883417 |
| Tetracladium marchalianum | CCM F-19399 | EU883423 |
| Tetracladium marchalianum | CCM F-26199 | AY204621 |
| Tetracladium marchalianum | CCM F-312 | EU883416 |
| Tetracladium marchalianum | NNIBRFG3891 | ON922530 |
| Tetracladium maxilliforme | CCM F-13186 | EU883430 |
| Tetracladium maxilliforme | CCM F-529 | EU883429 |
| Tetracladium maxilliforme | F-14286 | AF411027 |
| Tetracladium psychrophilum | HAILUO380 | JX029119 |
| Tetracladium psychrophilum | MY376 | JX029129 |
| Tetracladium setigerum | CCM F-10186 | EU883427 |
| Tetracladium setigerum | CCM F-19499 | EU883426 |
| Tetracladium setigerum | CCM F-20987 | KU519120 |
| Tetracladium setigerum | NNIBRFG1997 | ON922529 |
| Tetracladium setigerum | UMB 736.11 | KF952740 |
| Tricladium alaskense | VG-2012a | JQ417290 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Goh, J.; Jeon, Y.J.; Mun, H.Y. New Records of the Psychrophilic Tetracladium Species Isolated from Freshwater Environments in Korea. Diversity 2022, 14, 789. https://doi.org/10.3390/d14100789
AMA Style
Goh J, Jeon YJ, Mun HY. New Records of the Psychrophilic Tetracladium Species Isolated from Freshwater Environments in Korea. Diversity. 2022; 14(10):789. https://doi.org/10.3390/d14100789
Chicago/Turabian StyleGoh, Jaeduk, Yu Jeong Jeon, and Hye Yeon Mun. 2022. "New Records of the Psychrophilic Tetracladium Species Isolated from Freshwater Environments in Korea" Diversity 14, no. 10: 789. https://doi.org/10.3390/d14100789
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.