
Rhizomaticola guizhouensis gen. et sp. nov. and Five Rosellinia Like Species Isolated from Decaying Wood

1
State Key Laboratory of Functions and Applications of Medicinal Plants, Guizhou Medical University, Guiyang 550004, China
2
Engineering and Research Center for Southwest Bio-Pharmaceutical Resources of National Education Ministry of China, Guizhou University, Guiyang 550025, China
3
Immune Cells and Antibody Engineering Research Center of Guizhou Province, Guizhou Medical University, Guiyang 550004, China
4
Key Laboratory of Biology and Medical Engineering, Guizhou Medical University, Guiyang 550004, China
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(9), 703; https://doi.org/10.3390/d14090703
Submission received: 6 July 2022
/
Revised: 17 August 2022
/
Accepted: 22 August 2022
/
Published: 25 August 2022
(This article belongs to the Section Microbial Diversity and Culture Collections)
Abstract
:During the investigation of xylarialean taxa in China and Thailand, six rosellinia like taxa were collected. Rhizomaticola gen. nov. with type species of Rh. guizhouensis is established based on its morphology and multi-gene molecular data. Rhizomaticola owns no carbonaceous stromata and has black ascospores without a germ slit which are distinguished from those of Rosellinia, Dematophora, Stilbohypoxylon and Xylaria. Five rosellinia like species are introduced based on their morphology, inducing three new species (Dematophora populi, Rosellinia thailandica, Ro. vitis), one new record for China (Ro. cainii) and one known species (D. necatrix). Their descriptions and illustrations are detailed.
1. Introduction
Xylariaceae is one of the largest families of Xylariomycetidae, distributed worldwide, with high biodiversity [1,2,3,4,5]. Most species of the family were reported in tropical and subtropical regions [4,5,6,7]. Many xylariaceous species are saprophytes, including wood, dung, litter, termite nests and other substrates [8,9,10]. There were also a large number of endophytes and pathogenic fungi of plants were reported in Xylariaceae [8,11,12]. Many novel compounds with biological activity were found in the species of Xylariaceae [13,14]. A total of 32 genera were included in Xylariaceae by Wijayawardene et al. [15]. Xylaria Hill ex Schrank, Rosellinia De Not., Nemania Gray and Stilbohypoxylon Henn. are the most common genera in this family.
Rosellinia is characterized by carbonized, hard, uniperithecioid stromata usually growing from a subiculum, asci with a J+ apical apparatus bluing in Melzer’s reagent, ascospores with or without a germ slit [16]. According to morphological characteristics, 142 species were accepted by Petrini [16]. Only 48 species of Rosellinia have been reported from China [16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31]. Based on chemotaxonomic markers and a multi-locus phylogeny, the genus Dematophora R. Hartig was resurrected to include D. arcuata (Petch) C. Lamb., Wittstein and M. Stadler, D. buxi (Fabre) C. Lamb., Wittstein and M. Stadler, D. bunodes (Berk. and Broome) C. Lamb., Wittstein and M. Stadler, D. necatrix R. Hartig and D. pepo (Pat.) C. Lamb., Wittstein and M. Stadler and other allied species [32]. Dematophora differs from Rosellinia by its dematophora like asexual morph and forming a well-defined clade in phylogeny. With regard to the morphology of sexual morph, Wittstein et al. [32] did not point out the differences between Rosellinia and Dematophora.
In the process of investigating xylarialean taxa in China and Thailand, six rosellinia like species were collected from the forests, which were regard as an undescribed xylariaceous genus, Rhizomaticola to accommodate Rh. guizhouensis, as well as three species of Rosellinia and two species of Dematophora. Their descriptions and illustrations are provided.
2. Materials and Methods
2.1. Collection and Isolation
Samples with black dots were collected from forests in China and Thailand in the rainy season. Samples were put into paper bags with some silica gel desiccant. Macroscopic characteristics of stromata were observed and photographed under an Olympus SZ61 stereomicroscope. Materials were mounted in water and Melzer’s reagent for anatomical examination [31]. Macroscopic photographs of asci and ascospores were taken under a Nikon digital camera (700 D) fitted to a light microscope (Nikon Ni, Nikon Corporation, Tokyo, Japan). At least 30 ascospores and 30 asci were measured using the Tarosoft ® image framework (v. 0.9.0.7). Macroscopic photographs were made with the necessary changes and arranged for a plate. Single-spore isolation was used to obtain pure cultures [33]. Herbarium materials were deposited in the Herbarium of Guizhou Agricultural College (GACP); the Herbarium of the Engineering and Research Center for Southwest Bio-Pharmaceutical Resources of National Education Ministry of China, Guizhou University (GZUH); the Herbarium of Mae Fah Luang University (MFLU); and the living cultures are deposited in Guizhou University Culture Collection (GZUHC) or Mae Fah Luang University Culture Collection (MFLUCC).
2.2. DNA Extraction, Polymerase Chain Reaction (PCR) Amplification and Sequencing
Culture was grown on potato dextrose agar (PDA) and the hyphae were scraped off with a scalpel to extract DNA. Some stromata were cut and the contents were picked for DNA extraction directly. Total DNA was extracted by BIOMIGA Fungus Genomic DNA Extraction Kit (BW-GD2416-02, Biomiga, USA), following the manufacturer’s instructions. Regions of internal transcribed spacers (ITS), segments of large-subunit ribosomal RNA (LSU), partial β-tubulin gene (tub2), the second largest subunit of the RNA polymerase II (rpb2) were amplified with primer pairs ITS1/ITS4, LROR/LR5, Bt2a/Bt2b, RPB2-5F/RPB2-7Cr, respectively [34,35,36,37]. The components of a 25 μL volume PCR mixture were used as follows: double distilled water 9.5 μL, PCR master mix 12.5 μL, 1 μL of each primer, 1 μL template DNA [38]. PCR reaction systems were as follows: Long et al. [38] and Pi et al. [39]. Qualified PCR products checked with 1.5% agarose gel electrophoresis stained with GoldenView were sent to Sangon Co., China, for sequencing.
2.3. Sequence Alignment and Phylogenetic Analyses
All sequences for phylogenetic tree construction were chosen following published literature [40] and top hits of ITS blasted in the GenBank database (Table 1). ITS, LSU, tub2, rpb2 sequence data including all introns and exons were aligned separately using the MAFFT v.7.110 online programme (http://mafft.cbrc.jp/alignment/server/ (accessed on 15 August 2022) [41]) with the default settings. Multiple sequence alignments were generated and adjusted using BioEdit v.7.0.5.3 [42]. The MrModeltest 2.2 was used to perform the model of evolution [43]. The final ML search was conducted using the GTRGAMMA + I model. The phylogenetic analyses were carried out for maximum likelihood in CIPRES web portal [44] using RAxML 7.4.2 Black Box [45].
Bayesian analyses were performed in CIPRES web portal by using MrBayes on XSEDE [46]. The model of evolution was calculated by using MrModeltest v. 2.2 [43]. Markov Chain Monte Carlo sampling (MCMC) was used to determine posterior probabilities (PP) [47] in MrBayes on XSEDE. Six simultaneous Markov chains were run for 1,000,000 generations and trees were sampled every 1000th generation. The first 25% of trees were discarded during the burn-in phase of each analysis [48]. Phylogenetic trees were visualized and arranged using FigTree v1.4.0. and were edited with Adobe Photoshop CS6 [38]. The alignments were uploaded in TreeBASE (www.treebase.org/treebase-web/home.html (accessed on 15 August 2022) under ID 24609 for ITS-LSU-rpb2-tub2 alignment.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of taxa used for phylogenetic reconstruction.
| Species | Strains | GenBank Accession Number | References | |||
|---|---|---|---|---|---|---|
| ITS | LSU | rpb2 | tub2 | |||
| Amphirosellinia fushanensis | HAST 91111209HT | NR_153514 | N/A | GQ848339 | GQ495950 | [3] |
| A. nigrospora | HAST 91092308HT | NR_153513 | N/A | GQ848340 | GQ495951 | [3] |
| Annulohypoxylon annulatum | CBS 140775ET | NR_153579 | KY610418 | KY624263 | N/A | [49] |
| A. truncatum | CBS 140778ET | NR_153580 | KY610419 | KY624277 | N/A | [49] |
| Anthostomella thailandica | MFLUCC 15-0017HT | NR_153556 | KX533448 | KX599538 | KX600496 | [50] |
| Astrocystis bambusae | HAST 89021904 | GU322449 | N/A | GQ844836 | GQ495942 | [3] |
| A. mirabilis | HAST 94070803 | GU322448 | N/A | GQ844835 | GQ495941 | [3] |
| Barrmaelia rappazii | Cr2 = CBS 142771HT | MF488989 | MF488989 | MF488998 | MF489017 | [51] |
| B. rhamnicola | BR = CBS 142772ET | MF488990 | MF488990 | MF488999 | MF489018 | [51] |
| Biscogniauxia arima | 122 WSPIT | EF026150 | N/A | GQ304736 | AY951672 | [3] |
| Brunneiperidium gracilentum | MFLUCC:14-0011HT | KP297400 | KP340549 | KP340529 | KP406611 | [52] |
| Cainia anthoxanthis | MFLUCC 15-0539HT | NR_138407 | NG_070382 | N/A | N/A | [53] |
| C. globosa | MFLUCC 13-0663HT | NR_171724 | KX822123 | N/A | N/A | [50] |
| Camillea tinctor | YMJ 363 | JX507806 | N/A | JX507790 | JX507795 | [54] |
| Clypeosphaeria mamillana | WU 33598ET | NR_153909 | NG_067338 | MF489001 | MH704637 | [55] |
| Collodiscula japonica | CJ = CBS:124266 | JF440974 | MH874889 | KY624273 | KY624316 | [56] |
| C. leigongshanensis | GZ70 = GZUH0107HT | KP054281 | KP054282 | KR002588 | KR002587 | [29] |
| Coniocessia cruciformis | IRAN 1475CHT | NR_145220 | GU553347 | N/A | N/A | [57] |
| C. nodulisporioides | CBS 125778 | MH863756 | MH875224 | N/A | N/A | [57] |
| Coniolariella gamsii | IRAN 2506C | KY052004 | KY052005 | N/A | N/A | (submitted directly) |
| C. hispanica | CBS 124506T | MH863381 | MH874902 | N/A | N/A | [57] |
| Creosphaeria sassafras | CBS 127876 | MH864737 | MH876173 | N/A | N/A | [57] |
| Dematophora pepo | CBS:123592 | MN984620 | N/A | N/A | MN987246 | [32] |
| Diabolocovidia claustri | CPC37593HT | NR_170827 | NG_074445 | N/A | N/A | [58] |
| Diatrype lijiangensis | MFLU 19-0717HT | NR_165229 | MK810546 | N/A | MK852583 | [59] |
| Diatrypella heveae | MFLU:17-1216HT | MF959501 | NG_069531 | N/A | MG334557 | [60] |
| D. vulgaris | CBS 128327T | NR_159873 | NG_069986 | N/A | N/A | [57] |
| Durotheca comedens | YMJ 90071615 | EF026128 | N/A | JX507793 | EF025613 | [38] |
| D. guizhouensis | GMBC0065HT | MH645423 | MH645421 | MH645422 | MH645420 | [38] |
| Emarcea eucalyptigena | CBS 139908HT | MK762711 | NG_066346 | MK791286 | N/A | [61] |
| Engleromyces sinensis | BJTC 200803 | MZ622705 | MZ622702 | N/A | N/A | (submitted directly) |
| Entalbostroma erumpens | ICMP:21152HT | NR_154013 | N/A | KX258204 | KX258205 | [62] |
| Entoleuca mammata | JDR 100 | GU300072 | N/A | GQ844782 | GQ470230 | [3] |
| Eutypa lata | CBS 208.87NT | MH862066 | MH873755 | KF453595 | DQ006969 | [57] |
| Eutypella citricola | CBS 128332 | MH864883 | MH876331 | N/A | N/A | [57] |
| Furfurella nigrescens | CBS:143622HT | MK527844 | MK527844 | MK523275 | MK523333 | [51] |
| F. stromatica | CBS 144409HT | MK527846 | MK527846 | MK523277 | MK523334 | [51] |
| Graphostroma platystomum | CPC:37153 | MT223799 | MT223894 | MT223680 | MT223734 | [63] |
| Halorosellinia oceanica | BCC < THA > :60405 | MK606079 | MK629003 | N/A | N/A | (submitted directly) |
| H. xylocarpi | MFLU 18-0545HT | NR_166290 | NG_068301 | N/A | MN077076 | [40] |
| Hansfordia pulvinata | CBS 194.56 | KU683763 | MH869122 | KU684307 | N/A | [12] |
| Hansfordia pulvinata | CBS:144422 | MK442587 | MK442527 | N/A | N/A | [64] |
| Helicogermslita clypeata | MFLU 18-0852HT | NR_175685 | NG_081506 | MW658647 | MW775614 | [65] |
| Hypocopra rostrata | NRRL 66178 | KM067909 | KM067909 | N/A | N/A | (submitted directly) |
| Hypocreodendron sanguineum | 169 (JDR) | GU322433 | N/A | GQ844819 | GQ487710 | [3] |
| Hypoxylon rickii | MUCL 53309ET | NR_137115 | KY610416 | KY624281 | KC977288 | [49] |
| Idriella lunata | CBS:204.56T | MH857584 | MH869129 | N/A | N/A | [57] |
| Induratia thailandica | MFLU 18-0784HT | MK762707 | MK762714 | MK791283 | N/A | [61] |
| Jackrogersella multiformis | CBS 119016ET | NR_154784 | KY610473 | KY624290 | KX271262 | [49] |
| Kretzschmaria clavus | JDR 114 | EF026126 | N/A | GQ844789 | EF025611 | [3] |
| Kretzschmaria lucidula | JDR 112 | EF026125 | N/A | GQ844790 | EF025610 | [3] |
| Kretzschmariella culmorum | JDR 88 | KX430043 | N/A | KX430045 | KX430046 | (submitted directly) |
| Lopadostoma turgidum | CBS 133207ET | NR_132036 | KC774618 | KC774563 | MF489024 | [66] |
| Lunatiannulus irregularis | MFLUCC:14-0014HT | KP297398 | KP340540 | KP340526 | KP406609 | [52] |
| Microdochium phragmitis | CBS:285.71ET | MH860125 | KP858949 | KP859122 | MH704636 | [67] |
| Nemania bipapillata | HAST 90080610 | GU292818 | N/A | GQ844771 | GQ470221 | [3] |
| N. serpens | HAST 235 | GU292820 | N/A | GQ844773 | GQ470223 | [3] |
| N. sphaeriostoma | JDR 261 | GU292821 | N/A | GQ844774 | GQ470224 | [3] |
| Neoxylaria arengae | MFLUCC 15-0292HT | NR_171264 | N/A | MT502418 | N/A | [68] |
| Nigropunctata nigrocircularis | MFLU 19-2130HT | NR_175683 | NG_081504 | N/A | MW775612 | [65] |
| N. bambusicola | MFLU 19-2145HT | NR_175684 | NG_081505 | MW658646 | N/A | [65] |
| Podosordaria mexicana | 176 WSP | GU324762 | N/A | GQ853039 | GQ844840 | [3] |
| Podosordaria muli | 167 WSPHT | GU324761 | N/A | GQ853038 | GQ844839 | [3] |
| Poronia pileiformis | WSP 88113001ET | NR_158882 | N/A | GQ853037 | GQ502720 | [3] |
| Requienella fraxini | CBS 140475HT | NR_138415 | MH878686 | N/A | N/A | [57] |
| R. seminuda | CBS 140502ET | NR_154630 | MH878683 | MK523300 | N/A | [57] |
| Rhizomaticola guizhouensis | FJS12 = GZUH0101HT | ON815473 | 0N815474 | ON897692 | ON924997 | This study |
| Rh. guizhouensis | 2022FJS24 = GZUH0335 | OP177724 | OP177725 | OP184058 | OP184057 | This study |
| Rosellinia aquila | MUCL 51703 | KY610392 | KY610460 | KY624285 | KX271253 | [49] |
| Ro. corticium | STMA 13324 | MN984621 | MN984627 | MN987237 | MN987241 | [32] |
| Sarcoxylon compunctum | CBS:359.61 | KT281903 | KY610462 | KY624230 | KX271255 | [49] |
| Stilbohypoxylon elaeicola | 94082615 (HAST) | GU322440 | N/A | GQ844827 | GQ495933 | [68] |
| Stilbohypoxylon elaeidis | MFLUCC 15-0295aHT | MT496745 | NG_074460 | MT502416 | MT502420 | [68] |
| Stilbohypoxylon quisquiliarum | JDR 172 | EF026119 | N/A | GQ853020 | EF025605 | [3] |
| Vamsapriya bambusicola | MFLUCC11-0477 HT | KM462835 | NG_067527 | KM462834 | KM462833 | [58] |
| V. indica | MFLUCC 12-0544 | KM462839 | KM462840 | KM462841 | KM462838 | [58] |
| Wawelia regia | CBS:110.10 | MH854595 | MH866123 | N/A | N/A | [57] |
| Xylaria bambusicola | WSP 205HT | EF026123 | N/A | GQ844801 | AY951762 | [3] |
| X. feejeensis | HAST 92092013 | GU322454 | N/A | GQ848336 | GQ495947 | [3] |
| X. hypoxylon | HAST 95082001 | GU300095 | N/A | GQ844811 | GQ487703 | [3] |
| Zygosporium pseudomasonii | CBS 146059HT | MN562147 | MN567654 | MN556815 | N/A | (submitted directly) |
Notes: Type specimens are labeled with HT (holotype), ET (epitype), IT (isotype), NT (neotype), T (type). N/A: sequence not available. New sequences are marked as bold. HAST: Herbarium, research Center for Biodiversity, Academia Sinica, Taipei; CBS: Westerdijk Fungal Biodiversity Institute (CBS-KNAW Fungal Biodiversity Centre), Utrecht; MFLUCC: Mae Fah Luang University Culture Collection, Thailand; WSP: Washington State University, U.S.A.; YMJ: Herbarium of Yu-Min Ju; Wu: Herbarium of the Institute of Botany, University of Vienna, Austria; GZUH: Herbarium of Guizhou University; IRAN: Iranian Research Institute of Plant Protection, Tehran, Iran; ICMP: International Collection of Microorganisms from Plants; JDR: Herbarium of Jack D. Rogers; CPC: Culture collection of Pedro Crous, housed at CBS; BCC: Universitat de Barcelona; MFLU: Mae Fah Luang University Herbarium, Chiang Rai, Thailand; NRRI: Natural Resources Research Institute, University of Minnesota Duluth, Duluth, Minnesota; MUCL: University Catholique de Louvain.
3. Results
3.1. Phylogenetic Analyses
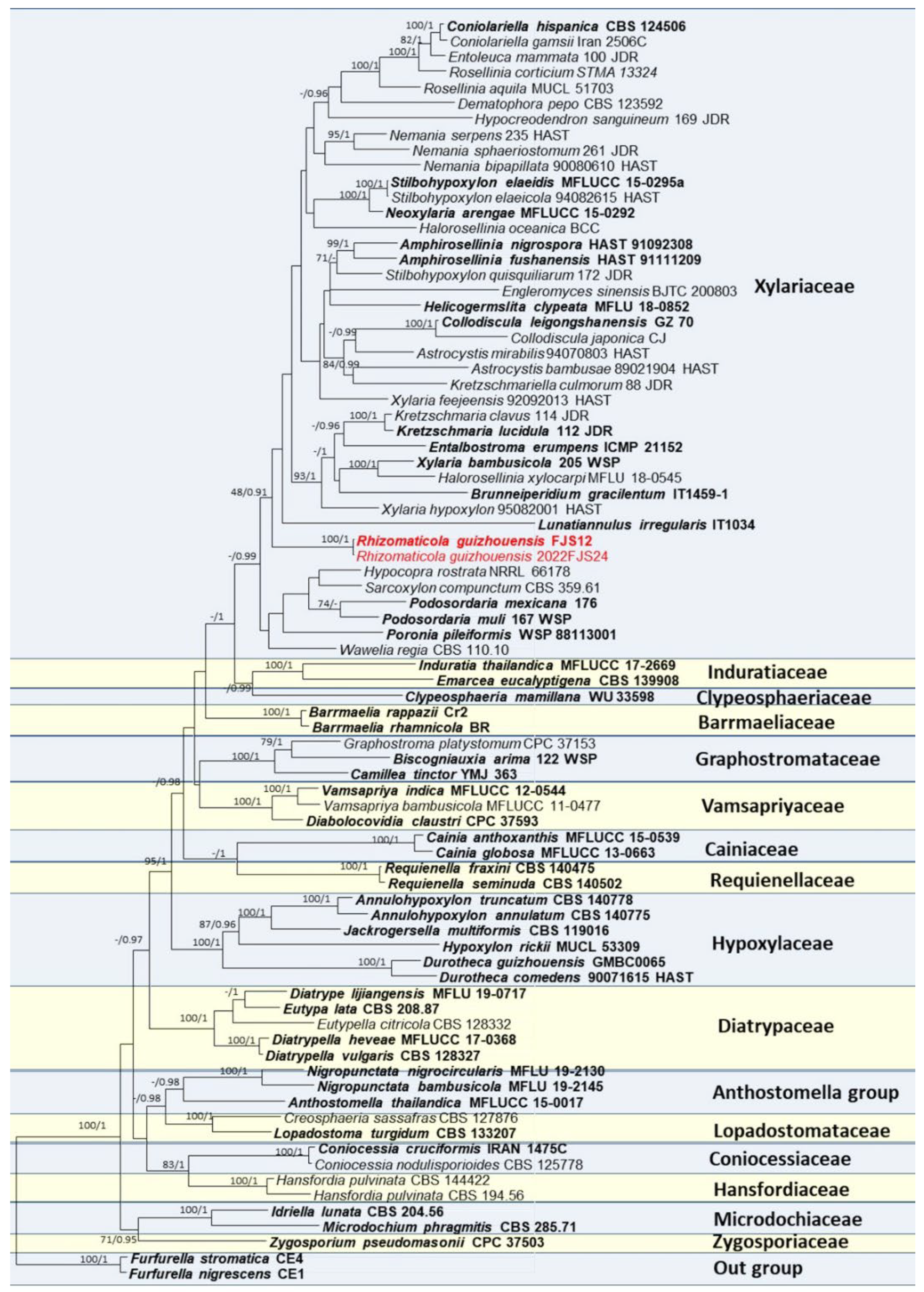
Multiple sequence alignment for constructing the phylogenetic tree (Figure 1) included 81 taxa, 2988 positions including gaps (ITS: 1–538, LSU: 539–1372, rpb2: 1373–2435, tub2: 2436–2988). All characters have equal weight. Of these characters, 1424 characters are constant, 315 variable characters are parsimony-uninformative. Number of parsimony-informative characters is 1249. Gaps were treated as “missing”. Rhizomaticola guizhouensis showed a distinct clade on the base of Xylariaceae (Figure 1), but bootstrap support values were not high (48, 0.91).
3.2. Taxonomy
Rhizomaticola Q.R. Li and J.C. Kang gen. nov.
MycoBank no.: 844445
Etymology: In reference to rhizome where the fungus is inhabited.
Holotype: GZUH0101
Type species: Rhizomaticola guizhouensis Q.R. Li and J.C. Kang, sp. nov.
Saprobic on dead rhizoma of Phragmites australis (Cav.) Trin. ex Steud., Sexual morph: No subiculum observed. Stromata scattered or gregarious, solitary, superficial, orbicular in outline, black, no carbonaceous, containing a single perithecium; Surface convex, black alternating with white, crack. The tissue between surface and perithecia white. Ostioles papillate on the central, black. Peridium black. Paraphyses hyaline, unbranched, septate, longer than asci. Asci 8-spored, unitunicate, long-cylindrical, long-stipitate, apically rounded with a J+, barrel-shaped apical apparatus. Ascospores overlapping uniseriate, dark brown to black, unicellular, long ellipsoidal to fusiform, prominent at ends, smooth-walled, lacking germ slits, appendages and clear sheaths. Asexual morph: Undetermined.
Notes—Morphologically, this genus is similar to Dematophora, Rosellinia, Stilbohypoxylon and Xylaria, all of which have large stromata visible to the naked eye and unitunicate asci with a J+ apical ring bluing in Melzer’s reagent, ascospores with germ slits [3,16,69,70]. However, Rhizomaticola has no subiculum, non-carbonaceous stromata cracking on its surface, white external stromata and ascospores lacking germ slits which are different from those close genera. Rhizomaticola differs from Collodiscula and Astrocystis by its non-carbonaceous stromata. Moreover, Collodiscula has ascospores with one too many septa, most species of Astrocystis have the ascospores with germ slits [19,29]. Molecular phylogenetic studies based on ITS, LSU, tub2 and rpb2 sequences in this study showed Rhizomaticola formed a distinct branch in Xylariaceae. Although the support values (48/0.91) are not high, its morphological characteristics are consistent with those of Xylariaceae. We would like to propose to temporarily place it in the Xylariaceae.
Rhizomaticola guizhouensis Q.R. Li and J.C. Kang, sp. nov. Figure 2.
Mycobank No.: 844446
Etymology: In reference to its collection location, Guizhou province, China.
Holotype—CHINA, Guizhou Province, Tongren city, The Fanjing Mountain Nature Reserve, on dead rhizome of P. australis, March, 2015, Q.R. Li and Lili Liu, FJS12 (GZUH0101, holotype, DNA was extracted directly from specimen; GACP QR0159). CHINA, Guizhou Province, Tongren city, The Fanjing Mountain Nature Reserve, on dead rhizome of P. australis (Cav.) Trin. ex Steud., July, 2022, Q.R. Li, 2022FJS24 (GZUH0335).
Description—Saprobic on dead rhizoma of P. australis. Sexual morph: No subiculum observed. Stromata 600–1000 µm diam., 500–800 µm high, scattered or gregarious, solitary, superficial, orbicular in outline, black, non carbonaceous, containing a single perithecium; Surface convex, black alternating with white, cracked. The tissue between surface and perithecia white. Ostioles papillate on the central, black. Peridium 55–65 µm thick, black. Paraphyses 2.5–4 μm wide, hyaline, unbranched, septate, longer than asci. Asci 221.5–320.5 × 12–18.5 µm (av. = 271.5 × 15.5 µm, n = 30), 8-spored, unitunicate, long-cylindrical, long-stipitate, the spore bearing part up to 155 µm long, apically rounded with a J+, barrel-shaped apical ring, 5–6.5 μm high, 4–5 μm broad. Ascospores 29.5–34.5 × 9–11 µm (av. = 32.5 × 9.5 µm, n = 30), overlapping uniseriate, dark brown to black, unicellular, long ellipsoidal to fusiform, prominent at both ends, smooth-walled, lacking germ slits, without appendages and sheaths. Asexual morph: Undetermined.
Culture characteristics—no culture was obtained; DNA was extracted directly from asci and ascospores in stromata.
Notes—Rhizomaticola guizhouensis is designated as the type species of Rhizomaticola. Rh. guizhouensis was found from Guizhou, China. Rhizomaticola guizhouensis differs from the uniperithecial species of Xylaria by it non-carbonaceous stromata, the ascospores lacking germ slits and observation of no multiple perithecia on a stroma [3,4,5]. Unfortunately, we could not obtain the pure culture of this species after many attempts. Ascospores did not germinate on PDA, OA (oatmeal agar) and MEA (malt extract agar) media.
Dematophora necatrix R. Hartig, Untersuch. Forstbot. Inst. München 3: 126 (1883). Figure 3.
Synonymy:
Rosellinia necatrix Berl. ex Prill., Bull. Soc. mycol. Fr. 20: 34 (1904)
Hypoxylon necatrix (Berl. ex Prill.) P.M.D. Martin, Jl S. Afr. Bot. 34: 187 (1968)
Hypoxylon necatrix (Berl. ex Prill.) P.M.D. Martin, Jl S. Afr. Bot. 42(1): 73 (1976)
Rosellinia radiciperda sensu auct. NZ; fide NZfungi (2008)
Pleurographium necator (R. Hartig) Goid., Ann. Bot., Roma 21(1): 48 (1935)
Rhizomorpha necatrix R. Hartig, Untersuch. Forstbot. Inst. München 3: 125 (1883)
Mycobank no.: 216282
Descrption—see Petrini (2013).
Distribution—China, France, Italy, Portugal, Spain
Specimen examined—CHINA, Guizhou Province, Guiyang city, Huaxi District, on unidentified plant stem, June 2014, Qirui Li, GZ28 (GZUH0139, GACP QR0198).
Notes—Dematophora necatrix is distributed worldwide. Morphologically, D. necatrix resembles D. bothrina (Berk. and Broome) C. Lamb., Wittstein and M. Stadler, D. compacta (Takemoto) C. Lamb., Wittstein and M. Stadler, D. paraguayensis (Starbäck) C. Lamb., Wittstein and M. Stadler, D. grantii (L.E. Petrini) C. Lamb., Wittstein and M. Stadler, D. siggersii (L.E. Petrini) C. Lamb., Wittstein and M. Stadler and D. acutispora (Theiss.) C. Lamb., Wittstein and M. Stadler [16,71]. Petrini [16] pointed out their differences. Morphological characteristics of new collection (GZUH0139) are consistent with those of D. necatrix.
Dematophora populi Q.R. Li and J.C. Kang sp. nov. Figure 4.
Mycobank no.: 844442
Etymology: The name refers the name of host, Populus sp.
Holotype—CHINA: Guizhou Province, Guiyang city, Baiyun park, on dead branch of Populus sp., May 2014, Qirui Li, GZ7 (GZUH0116, holotype; GACP QR0214); CHINA: Guizhou Province, Guiyang city, Guiyang Forest Park, on dead branch of an unknown plant, June 2014, Qirui Li, GYSLGY09 (GZUH0117; GACP QR0215).
Description—Saprobic on dead branch of Populus sp., Sexual morph: Subiculum evanescent, black, felty. Stromata 1.5–2 mm diam., 1.0–2.0 mm high, carbonaceous, globose to subglose, with fine papillate. Ectostroma up to 150 μm thick, black. Entostroma disappearing at matutity. Perithecia 1000–1500 μm diam., 800–1425 μm high, black. Asci 240–315 × 11–13 μm (av. = 287 × 12 μm, n = 15), 8-spores, unitunicate, long-cylindrical, apically rounded, with an urn-shaped apical ring, 10.5–12.5 μm high (av. = 11.6 μm, n = 20), upper width 3.5–4.5 μm (av. = 4.2 μm, n = 20), lower width 6–7 μm (av. = 6.7 μm, n = 20). Ascospores 34–44 × 6–8 μm (av. = 38.5 × 7.1 μm, n = 30), extremely narrowly ellipsoidal to almost fusiform, brown to dark brown, smooth-walled, with short cell appendages on the ends; germ slit short, central, straight, far less than half of spore length, lacking sheaths. Asexual morph: Undetermined.
Notes—In term of ascospores dimension, D. populi belongs to Rosellinia necatrix group, and which is close to D. bothrina (≡ Ro. arcuata Petch), D. necatrix, D. paraguayensis Starbäck in this group [16]. However, possessing cell appendages on both ends of ascospores, D. populi are clearly different from them. Dematophora populi morphologically shows similarities to Ro. desmazieri (Berk. and Br.) Sacc. but differs by its longer ascospores (34–44 × 6–8 μm vs. 25–30 × 6.6–8.1 μm) and shorter germ slit of ascospores [16]. Although its anamorph was not observed, we put it in Dematophora here, since the Dematophora populi belongs to Rosellinia necatrix group, and the species of this group were transferred to Dematophora [32].
Rosellinia cainii L.E. Petrini, Index Fungorum 25: 1 (2013). Figure 5.
Mycobank no.: 550201
Description—Saprobic on dead branches of an unknown plant. Sexual morph: Subiculum woolly, wiry, felted brown to black, persistent or not. Stromata 600–800 μm diam., 500–900 μm high, solitary or densely, smooth, superficial, spherical with a papillate ostiole, containing single perithecia in one stroma. Ostioles finely papillate. Ectostroma up to 30 μm thick, black. Entostroma black, easily separated from ectostroma at matutity. Paraphyses 3–5 μm, hyaline, unbranched, septate. Asci 170–206 × 10.5–16 μm (av. = 187.8 × 13.5 μm, n = 30), 8-spores, unitunicate, cylindrical, apically rounded, with a blue, urn-shaped, J+, apical rings stained in Melzer’s reagent, 10.5–12.5 μm high (av. = 11 μm, n = 30), upper width 3.5–4.5 μm (av. = 4 μm, n = 30), lower width 5–7.5 μm (av. = 6.5 μm, n = 30). Ascospores 22.5–30.5 × 7.5–9.5 μm (av. = 25.7 × 8.4 μm, n = 30), overlapping uniseriate, ellipsoidal to asymmetrically ellipsoidal with broadly rounded ends, dark brown to black, smooth-walled, with a straight germ slit nearly spore length, lacking sheaths and appendages. Asexual morph: undetermined.
Distribution—Canada, China
Specimen examined—CHINA, Yunnan Province, Pu’er city, Wuliangshan National Nature Reserve, on unknown plant, June 2014, Qirui Li, WLS1 (GZUH0119, GACP QR0217).
Notes—Rosellinia cainii was introduced by Petrini [16] as a new species, since its broadly rounded ascospores without slimy sheaths and caps. No available description for asci in the original literature. The first collection was found on Corylus rostrata hort. ex Dippel from Canada. This is the first report for Ro. cainii collected in China.
Rosellinia thailandica Q.R. Li and J.C. Kang sp. nov. Figure 6.
Mycobank no.: 844443
Etymology: in reference to the collection country, Thailand.
Holotype—THAILAND, Chiang Mai, Campus of Mae Fah Luang University, on deadwood, November 2012, Q.R. Li, T24 (MFLU12-2136 holotype, ex-type culture MFLUCC 13-0166; GZUH0058); THAILAND, Chiang Mai, on unknown plant dead branches, December 2012, Qirui Li, T35 (MFLU 12-2146, GZUH0065)
Description—Saprobic on dead branches of unknown angiosperm plant. Sexual morph: Subiculum woolly, brown to black, persistent, gathering at the bottom of stromata. Stromata 950–1200 μm wide, 600–800 μm high, carbonaceous, subglobose to globose, solitary or gregarious, embedded up to the middle the subiculum, smooth, with metallic luster. Ostioles finely papillate, well-developed. Ectostroma 120 μm, black. Entostroma black. Asci 170–235 × 26–39 μm (av. = 203 × 33 μm, n = 30) 8-spored, unitunicate, cylindrical to clavate, short pedicellate, apically rounded, with a J+, urn–shaped apical ring, bluing in Melzer’s reagent, 19–24 μm (av. = 22 μm, n = 30) high, 9–11 μm (av. = 10 μm, n = 30) wide. Ascospores 72.5–144.5 × 10.0–15.0 μm (av. = 109.1 × 12.7 μm, n = 30), overlapping, fusiform, with round ends, asymmetrically equilateral, dark brown at maturity, unicellular, smooth, with a germ slit in the center of the ascospores, nearly half of spore-length, possessing thin, slimy sheaths covering ends of ascospores, lacking appendages. Asexual morph: Undetermined.
Notes—In term of stromata and ascospores dimension, Rosellinia thailandica belongs to Ro. emergens group [16]. Species with similar ascorspore dimension are Ro. macrosperma Speg., Ro. markhamiae Sivan., Ro. megalosperma Syd. and P. S.yd. [16,72]. However, Ro. thailandica possess the higher apical rings than those of them. Moreover, Ro. megalosperma and Ro. markhamiae have spore-length germ slit. Sheaths were not observed on the ascospores of Ro. megalosperma [16,72]. Rosellinia macrosperma owns narrower ascospores (10 ± 1.7 μm vs. 10–14 μm) without germ slits [16].
Rosellinia vitis Q.R. Li and J.C. Kang, sp. nov. Figure 7.
Mycobank no.: 844444
Etymology: The name refers the host of vine.
Holotype—CHINA, Yunnan Province, Pu’er city, Xishuangbanna Nature Reserve, on the dead vines of unknown plants, June 2014, Qirui Li, XSBN25 (GZUH0123 holotype, GACP QR0222)
Description—Saprobic on the dead vines of unknown plants. Sexual morph: Subiculum woolly, brown to black, persistent, gathering at the bottom of stromata. Stromata 550–1100 μm wide, 420–790 μm high, carbonaceous, subglobose to globose, solitary or gregarious, embedded up to the bottom the subiculum, smooth. Ostioles finely papillate, well-developed. Ectostroma 80 μm, black. Entostroma black. Asci 165–270 × 27–35 μm (av. = 210.5 × 31.4 μm, n = 30), 8-spored, unitunicate, short cylindrical to clavate, short pedicellate, apically rounded, with a J+, urn-shaped apical rings, bluing in Melzer’s reagent, 21.5–26.5 μm (av. = 24 μm, n = 30) high, 7–15 μm (av. = 12 μm, n = 30) wide. Ascospores 92–116.5 × 12.5–18.5 μm (av. = 109.9 × 13.7 μm, n = 30), overlapping, fusiform, with round ends, asymmetrically equilateral, dark brown at maturity, unicellular, smooth, lacking germ slits, sheathes and appendages. Asexual morph: Undetermined.
Notes—In term of stromata and ascospores dimension, Rosellinia vitis belongs to Ro. emergens group [16]. Rosellinia vitis is most close to Ro. capetribulensis, Ro. markhamiae, and Ro. macrosperma [16,72,73]. Rosellinia vitis, however, differs from Ro. capetribulensis and Ro. markhamiae by lacking germ slit on ascospores. Entostroma of Ro. macrosperma is white and its ascospores (103.3 ± 8.5 × 10 ± 1.7 μm) are narrower than those of Ro. vitis.
4. Discussion
Xylariaceae is a worldwide distributed group which includes common characteristics such as ascomata visible to the naked eye, unitunicate asci with or without a J+, apical apparatus, brown to black, rarely hyaline, 1–2-celled ascospores mostly with a germ slit, geniculosporium-like or nodulisporium-like asexual morph [40]. Here, we introduce a new genus, Rhizomaticola to accommodate the type species of Rh. guizhouensis isolated from China. Rhizomaticola have black ascospores without a germ slit and no hard carbonaceous stromata which can be distinguished from its similar genera.
Rosellinia and Dematophora are widely distributed in tropical and subtropical regions and mainly saprophytes on plant branches [27,32,74]. In this paper, we introduced five species of Rosellinia and Dematophora which were collected from China and Thailand and identified them based on their morphology. We have attempted to isolate the pure cultures of these specimens, but only a part of the isolations has been obtained. We found that the larger the ascospores, the less likely it is to germinate in Rosellinia and Dematophora. Moreover, the culture is likely to die after being stored for a while at 4 ℃.
Many taxonomic features are used for the identification of species within Rosellinia and Dematophora. The commonly used morphological characteristics mainly include: the size and shape of the stromata; the size and shape of the apical ring of ascus; the size and shape of the ascospores; the length of germ slits; the type of appendages; and the presence and type of sheaths covering the ascospores [16,75,76,77]. Anamorph is used for species identification as well, although only a few species of asexual stage have been observed [32,49,74]. DNA sequences have also been carried out for the identification of species within those genera [30,73]. However, there are only a few DNA sequences of Rosellinia available on Genbank. Secondary metabolites were attempted to be taken as a taxonomic feature to identify species of Rosellinia and Dematophora [32]. Dematophora was resurrected as inferred from polythetic taxonomy, while the results of utility of secondary metabolites as genus-specific chemotaxonomic markers were inconclusive [32].
Author Contributions
Investigation and collected specimens, Q.L., L.L.; checked specimens and DNA extract, S.L.; Microphotography, Q.L.; Validation, J.K.; Visualization, Y.L.; writing, original draft preparation Q.L., L.L.; writing, review and editing, S.L., Y.L., X.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (31960005, 32000009 and 32170019); the Open Fund Program of Engineering Research Center of Southwest Bio-Pharmaceutical Resources, Ministry of Education, Guizhou University No. GZUKEY20160; the Fund of the Science and Technology Foundation of Guizhou Province ([2020]1Y059); the Fund of Special Project of Academic New Seedling Cultivation and Innovation Exploration in Guizhou Medical University [2018]5779-64; Guizhou Province Ordinary Colleges and Universities Youth Science and Technology Talent Growth Project [2021]154.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study were included in this published article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Peláez, F.; González, V.; Platas, G.; Sánchez-Ballesteros, J.; Rubio, V. Molecular phylogenetic studies within the Xylariaceae based on ribosomal DNA sequences. Fungal Divers. 2008, 31, 111–134. [Google Scholar]
- Tang, A.; Jeewon, R.; Hyde, K.D. A re-evaluation of the evolutionary relationships within the Xylariaceae based on ribosomal and protein-coding gene sequences. Fungal Divers. 2009, 34, 127–155. [Google Scholar]
- Hsieh, H.-M.; Lin, C.-R.; Fang, M.-J.; Rogers, J.D.; Fournier, J.; Lechat, C.; Ju, Y.M. Phylogenetic status of Xylaria subgenus Pseudoxylaria among taxa of the subfamily Xylarioideae (Xylariaceae) and phylogeny of the taxa involved in the subfamily. Mol. Phylogenet. Evol. 2010, 54, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Fournier, J.; Lechat, C.; Courtecuisse, R. The genus Xylaria sensu lato (Xylariaceae) in Guadeloupe and Martinique (French West Indies) II. Taxa with robust upright stromata. Ascomycete.org 2019, 11, 77–115. [Google Scholar]
- Fournier, J.; Lechat, C.; Courtecuisse, R. The genus Xylaria sensu lato (Xylariaceae) in Guadeloupe and Martinique (French West Indies) III. Taxa with slender upright stromata. Ascomycete.org 2020, 12, 81–164. [Google Scholar]
- Carmona, A.; Fournier, J.; Williams, C.; Piepenbring, M. New records of Xylariaceae from Panama. N. Am. Fungi 2009, 4, 1–11. [Google Scholar] [CrossRef]
- Stadler, M.; Læssøe, T.; Fournier, J.; Decock, C.; Schmieschek, B.; Tichy, H.V.; Peršoh, D. A polyphasic taxonomy of Daldinia (Xylariaceae). Stud. Mycol. 2014, 77, 1–143. [Google Scholar] [CrossRef]
- Edwards, R.L.; Jonglaekha, N.; Kshirsagar, A.; Maitland, D.J.; Mekkamol, S.; Nugent, L.K.; Phosri, C.; Rodtong, S.; Ruchikachorn, N.; Sangvichien, E.; et al. The Xylariaceae as phytopathogens. Recent Res. Dev. Plant Sci. 2003, 1, 1–19. [Google Scholar]
- Visser, A.A.; Ros, V.I.D.; de Beer, Z.W.; Debets, A.J.M.; Hartog, E.; Kuyper, T.W.; Læssøe, T.; Slippers, B.; Aanen, D.K. Levels of specificity of Xylaria species associated with fungus-growing termites: A phylogenetic approach. Mol. Ecol. 2009, 18, 553–567. [Google Scholar] [CrossRef]
- Pažoutová, S.; Šrůtka, P.; Holuša, J.; Chudíčková, M.; Kolařík, M. Diversity of xylariaceous symbionts in Xiphydria woodwasps: Role of vector and a host tree. Fungal Ecol. 2010, 3, 392–401. [Google Scholar] [CrossRef]
- Okane, I.; Toyama, K.; Nakagiri, A.; Suzuki, K.I.; Srikitikulchai, P.; Sivichai, S.; Hywel-Jones, N.; Potacharoen, W.; Læssøe, T. Study of endophytic Xylariaceae in Thailand: Diversity and taxonomy inferred from rDNA sequence analyses with saprobes forming fruit bodies in the field. Mycoscience 2008, 49, 359–372. [Google Scholar] [CrossRef]
- U’Ren, J.M.; Miadlikowska, J.; Zimmerman, N.B.; Lutzoni, F.; Stajich, J.E.; Arnold, A.E. Contributions of North American endophytes to the phylogeny, ecology, and taxonomy of Xylariaceae (Sordariomycetes, Ascomycota). Mol. Phylogenet. Evol. 2016, 98, 210–232. [Google Scholar] [CrossRef]
- Stadler, M. Importance of secondary metabolites in the Xylariaceae as parameters for assessment of their taxonomy, phylogeny, and functional biodiversity. Curr. Res. Environ. Appl. Mycol. 2011, 1, 75–133. [Google Scholar] [CrossRef]
- Surup, F.; Wiebach, V.; Kuhnert, E.; Stadler, M. Truncaquinones A and B, asterriquinones from Annulohypoxylon truncatum. Tetrahedron Lett. 2016, 47, 2183–2185. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungi-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Petrini, L.E. Rosellinia—A World Monograph; Gebrüder Borntraeger Verlagsbuchhandlung: Stuttgart, Germany, 2013; pp. 1–410. [Google Scholar]
- Teng, S.C. Fungi of China; Science Press: Beijing, China, 1963; pp. 1–808. [Google Scholar]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979; pp. 1–1527. [Google Scholar]
- Ju, Y.M.; Rogers, J.D. Astrocystis reconsidered. Mycologia 1990, 82, 342–349. [Google Scholar] [CrossRef]
- Ju, Y.M.; Rogers, J.D. The Xylariaceae of Taiwan (excluding Anthostomella). Mycotaxon 1999, 73, 343–440. [Google Scholar]
- Yuan, Z.Q.; Zhao, Z.Y. Studies on the genera Amphisphaerella, Coniochaeta and Rosellinia of XingJiang, China. Acta Mycol. Sin. 1993, 12, 180–186. [Google Scholar]
- Lu, B.S.; Hyde, K.D.; Ho, W.H.; Tsui, K.M.; Taylor, J.E.; Wong, K.M.; Zhou, D.Y. Checklist of Hong Kong Fungi; Fungal Diversity Press: Hongkong, China, 2000; pp. 1–207. [Google Scholar]
- Liu, C.F.; Lu, T.; Gao, J.M.; Wang, M.Q.; Lu, B.S. Two new Chinese records of Rosellinia. Mycosystema 2010, 29, 459–462. [Google Scholar]
- Petrini, L.E. Nomenclatural novelties [Rosellinia]. Index Fungorum. 2013, 25, 1–6. [Google Scholar]
- Li, W.; Guo, L. Rosellinia brunneola sp. nov. and R. beccariana new to China. Mycotaxon 2015, 130, 233–236. [Google Scholar] [CrossRef]
- Li, W.; Guo, L. Rosellinia hainanensis sp. nov. and three Rosellinia species new to China. Mycotaxon 2016, 131, 541–545. [Google Scholar] [CrossRef]
- Li, W.; Guo, L. Rosellinia jiangxiensis and R. yunnanensis spp. nov. and a new Rosellinia record from China. Mycotaxon 2018, 133, 31–35. [Google Scholar] [CrossRef]
- Li, W.; Guo, L. Rosellinia angusta and R. menglana spp. nov. and two new Rosellinia records from China. Mycotaxon 2018, 133, 591–595. [Google Scholar] [CrossRef]
- Li, Q.R.; Kang, J.C.; Hyde, K.D. Two new species of the genus Collodiscula (Xylariaceae) from China. Mycol. Prog. 2015, 205, 187–196. [Google Scholar] [CrossRef]
- Su, H.; Li, Q.R.; Kang, J.C.; Wen, T.C.; Hyde, K.D. Rosellinia convexa, sp. nov. (Xylariales, Pezizomycotina) from China. Mycoscience 2016, 57, 164–170. [Google Scholar] [CrossRef]
- Xie, X.; Liu, L.; Zhang, X.; Long, Q.D.; Sheng, X.C.; Boonmee, S.; Kang, J.C.; Li, Q.R. Contributions to species of Xylariales in China—2. Rosellinia pervariabilis and R. tetrastigmae spp. nov. and a new record of R. caudata. Mycotaxon 2019, 134, 183–196. [Google Scholar] [CrossRef]
- Wittstein, K.; Cordsmeier, A.; Lambert1, C.; Wendt, L.; Sir, E.B.; Weber, J.; Wurzler, N.; Petrini, L.E.; Stadler, M. Identification of Rosellinia species as producers of cyclodepsipeptide PF1022 A and resurrection of the genus Dematophora as inferred from polythetic taxonomy. Stud. Mycol. 2020, 96, 1–16. [Google Scholar] [CrossRef]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alisa, A.S.; Xu, J.C.; Liu, X.Z.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes–application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Long, Q.D.; Liu, L.L.; Zhang, X.; Wen, T.C.; Kang, J.C.; Hyde, K.D.; Shen, X.C.; Li, Q.R. Contributions to species of Xylariales in China-1. Durotheca species. Mycol. Prog. 2019, 18, 495. [Google Scholar] [CrossRef]
- Pi, Y.H.; Zhang, X.; Liu, L.L.; Long, Q.D.; Shen, X.C.; Kang, Y.Q.; Hyde, K.D.; Boonmee, S.; Kang, J.C.; Li, Q.R. Contributions to species of Xylariales in China–4. Hypoxylon wujiangensis sp. nov. Phytotaxa 2020, 455, 21–30. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Nylander, J.A.A. MrModeltest v2.2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Swden, 2004. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop 2010 (GCE), New Orleans, LA, USA, 14–15 November 2010. [Google Scholar]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef]
- Lu, Y.Z.; Liu, J.K.; Hyde, K.D.; Jeewon, R.; Kang, J.C.; Fan, C.; Boonmee, S.; Bhat, D.J.; Luo, Z.L.; Lin, C.G.; et al. A taxonomic reassessment of Tubeufiales based on multi-locus phylogeny and morphology. Fungal Divers. 2018, 92, 131–344. [Google Scholar] [CrossRef]
- Wendt, L.; Sir, E.B.; Kuhnert, E.; Heitkämper, S.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Luangsa-ard, J.J.; Srikitikulchai, P.; Peršoh, D.; et al. Resurrection and emendation of the Hypoxylaceae, recognised from a multigene phylogeny of the Xylariales. Mycol. Prog. 2018, 17, 115–154. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Voglmayr, H.; Aguirre-Hudson, M.B.; Wagner, H.G.; Tello, S.; Jaklitsch, W.M. Lichens or endophytes? The enigmatic genus Leptosillia in the Leptosilliaceae fam. nov. (Xylariales), and Furfurella gen. nov. (Delonicicolaceae). Persoonia 2019, 42, 228–260. [Google Scholar] [CrossRef]
- Daranagama, D.A.; Camporesi, E.; Tian, Q.; Liu, X.; Chamyuang, S.; Stadler, M.; Hyde, K.D. Anthostomella is polyphyletic comprising several genera in Xylariaceae. Fungal Divers. 2015, 73, 203–238. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Maharachchikumbura, S.; Hyde, K.D.; Bhat, J.D.; Jones, E.B.G.; McKenzie, E.H.C.; Dai, D.Q.; Daranagama, D.A.; Dayarathne, M.C.; Goonasekara, I.D.; et al. Towards unraveling relationships in Xylariomycetidae (Sordariomycetes). Fungal Divers. 2015, 73, 73–144. [Google Scholar] [CrossRef]
- Mirabolfathy, M.; Ju, Y.M.; Hsieh, H.M.; Rogers, J.D. Obolarina persica sp. nov. associated with dying Quercus in Iran. Mycoscience 2013, 54, 315–320. [Google Scholar] [CrossRef]
- Liu, F.; Bonthond, G.; Groenewald, J.; Cai, L.; Crous, P. Sporocadaceae, a family of coelomycetous fungi with appendage-bearing conidia. Stud. Mycol. 2019, 92, 287–415. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Voglmayr, H. Phylogenetic relationships of five genera of Xylariales and Rosasphaeria gen. nov. (Hypocreales). Fungal Divers. 2012, 52, 75–98. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNAbarcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Sun, Y.R.; Liu, N.G.; Samarakoon, M.C.; Jayawardena, R.S.; Hyde, K.D.; Wang, Y. Morphology and phylogeny reveal Vamsapriyaceae fam. nov. (Xylariales, Sordariomycetes) with two novel Vamsapriya species. J. Fungi 2021, 7, 891. [Google Scholar] [CrossRef]
- Thiyagaraja, V.; Senanayake, I.C.; Wanasinghe, D.N.; Karunarathna, S.C.; Worthy, F.R.; To-Anun, C. Phylogenetic and morphological appraisal of Diatrype lijiangensis sp. nov. (Diatrypaceae, Xylariales) from China. Asian J. Mycol. 2019, 2, 198–208. [Google Scholar] [CrossRef]
- Senwanna, I.C. Novel taxa of Diatrypaceae from Para rubber (Hevea brasiliensis) in northern Thailand; introducing a novel genus Allocryptovalsa. Mycosphere 2017, 8, 1835–1855. [Google Scholar] [CrossRef]
- Samarakoon, M.C.; Thongbai, B.; Hyde, K.D.; Brönstrup, M.; Beutling, U.; Lambert, C.; Miller, A.N.; Liu, J.K.; Promputtha, I.; Stadler, M. Elucidation of the life cycle of the endophytic genus Muscodor and its transfer to Induratia in Induratiaceae fam. nov. based on a polyphasic taxonomic approach. Fungal Divers. 2020, 101, 177–201. [Google Scholar] [CrossRef]
- Johnston, P.; Rogers, J.; Park, D.; Martin, N.A. Entalbostroma erumpens gen. et sp. nov. (Xylariaceae) from Phormium in New Zealand. Mycotaxon 2016, 131, 765–771. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Schumacher, R.K.; Akulov, A.; Bulgakov, T.S.; Carnegie, A.J.; Jurjević, Ž.; Decock, C.; Denman, S.; Lombard, L.; et al. New and interesting fungi. 3. Fungal Syst. Evol. 2020, 6, 157–231. [Google Scholar] [CrossRef]
- Crous, P.W.; Schumacher, R.K.; Akulov, A.; Thangavel, R.; Hernández-Restrepo, M.; Carnegie, A.; Cheewangkoo, R.; Wingfield, M.J.; Summerell, B.; Quaedvlieg, W.; et al. New and Interesting Fungi. 2. Fungal Syst. Evol. 2019, 3, 57–134. [Google Scholar] [CrossRef]
- Samarakoon, M.C.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Stadler, M.; Jones, E.B.G.; Promputtha, I.; Suwannarach, N.; Camporesi, E.; Bulgakov, T.S.; Liu, J.K. Taxonomy, phylogeny, molecular dating and ancestral state reconstruction of Xylariomycetidae (Sordariomycetes). Fungal Divers. 2022, 112, 1–88. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Fournier, J.; Rogers, J.D.; Voglmayr, H. Phylogenetic and taxonomic revision of Lopadostoma. Persoonia 2014, 32, 52–82. [Google Scholar] [CrossRef]
- Hernández-Restrepo, M.; Groenewald, J.; Crous, P. Taxonomic and phylogenetic re-evaluation of Microdochium, Monographella and Idriella. Persoonia Mol. Phylogeny Evol. Fungi 2016, 36, 57–82. [Google Scholar] [CrossRef] [Green Version]
- Konta, S.; Hyde, K.D.; Phookamsak, R.; Xu, J.C.; Maharachchikumbura, S.S.N.; Daranagama, D.A.; McKenzie, E.H.C.; Boonmee, S.; Tibpromma, S.; Eungwanichayapant, P.D.; et al. Polyphyletic genera in Xylariaceae (Xylariales): Neoxylaria gen. nov. and Stilbohypoxylon. Mycosphere 2020, 11, 2629–2651. [Google Scholar] [CrossRef]
- Rogers, J.D.; Ju, Y.M. The genus Stilbohypoxylon. Mycol. Res. 1997, 101, 135–138. [Google Scholar] [CrossRef]
- Petrini, L.E. A revision of the genus Stilbohypoxylon (Xylariaceae). Sydowia 2004, 56, 51–71. [Google Scholar]
- Takemoto, S.; Nakamura, H.; Sasaki, A.; Shimane, T. Species-specific PCRs differentiate Rosellinia necatrix from R. compacta as the prevalent cause of white root rot in Japan. J. Gen. Plant Pathol. 2011, 77, 107–111. [Google Scholar] [CrossRef]
- Sivanesan, A. New ascomycetes and some revisions. Trans. Br. Mycol. Soc. 1975, 65, 19. [Google Scholar] [CrossRef]
- Bahl, J.; Hyde, R. Phylogeny of Rosellinia capetribulensis sp. nov. and its allies (Xylariaceae). Mycologia 2005, 97, 1102–1110. [Google Scholar] [CrossRef]
- Petrini, L.E. Rosellinia and related genera in New Zealand. N. Z. J. Bot. 2003, 41, 71–138. [Google Scholar] [CrossRef]
- Læssøe, T.; Spooner, B.M. Rosellinia & Astrocystis (Xylariaceae): New Species and Generic Concepts. Kew Bull. 1993, 49, 1–70. [Google Scholar]
- Petrini, L.E.; Petrini, O. Morphological studies in Rosellinia (Xylariaceae): The first step towards a polyphasic taxonomy. Mycol. Res. 2005, 109, 569–580. [Google Scholar] [CrossRef]
- Rogers, J.D.; Miller, A.N.; Vasilyeva, L.N. Pyrenomycetes of the Great Smoky Mountains National Park. VI. Kretzschmaria, Nemania, Rosellinia and Xylaria (Xylariaceae). Fungal Divers. 2008, 29, 107–116. [Google Scholar]
Figure 1.
Phylogeny of Xylariales obtained from a Maximum Likelihood analysis of the combined ITS, LSU, rpb2 and tub2 using RAxML-HPC BlackBox software online. Furfurella nigrescens (CE1) and F. stromatica (CE4) were taken as outgroup taxa. Strains or specimen numbers were followed by their names. Type and authority strains are marked in bold. Bayesian posterior probabilities ≥0.95 and bootstrap support values for maximum likelihood (ML) higher than ≥70% are marked above the nodes; an en-dash (“-”) indicates a value < 0.95 (PP) or <70% (BS).
Figure 1.
Phylogeny of Xylariales obtained from a Maximum Likelihood analysis of the combined ITS, LSU, rpb2 and tub2 using RAxML-HPC BlackBox software online. Furfurella nigrescens (CE1) and F. stromatica (CE4) were taken as outgroup taxa. Strains or specimen numbers were followed by their names. Type and authority strains are marked in bold. Bayesian posterior probabilities ≥0.95 and bootstrap support values for maximum likelihood (ML) higher than ≥70% are marked above the nodes; an en-dash (“-”) indicates a value < 0.95 (PP) or <70% (BS).

Figure 2.
Rhizomaticola guizhouensis (GZUH0101,holotype) (A) Material; (B,C) Stromata on the surface of host; (D,E) Sections of stromata; (F,G) Ascus apex with a J+, apical ring (stained in Melzer’s reagent); (H–J) Asci with ascospores; (K–O) Ascospores. Scale bars: (A) = 0.5 cm, (B–E) = 200 µm, (F,G) = 5 µm, (H–J) = 10 µm, (K–O) = 5 µm.
Figure 2.
Rhizomaticola guizhouensis (GZUH0101,holotype) (A) Material; (B,C) Stromata on the surface of host; (D,E) Sections of stromata; (F,G) Ascus apex with a J+, apical ring (stained in Melzer’s reagent); (H–J) Asci with ascospores; (K–O) Ascospores. Scale bars: (A) = 0.5 cm, (B–E) = 200 µm, (F,G) = 5 µm, (H–J) = 10 µm, (K–O) = 5 µm.

Figure 3.
Dematophora necatrix (GZUH0139) (A,B) Stromata on the host; (C) Vertical section of a stroma; (D–F) Asci; (G,H) Urn–shaped J+ apical rings (stained in Melzer’s reagent); (I–L) Ascospores. Bars: (A) = 2 mm, (B) = 500 μm, (C) = 200 μm, (D) = 20 μm, (E–H) = 20 μm, (I–L) = 10 μm.
Figure 3.
Dematophora necatrix (GZUH0139) (A,B) Stromata on the host; (C) Vertical section of a stroma; (D–F) Asci; (G,H) Urn–shaped J+ apical rings (stained in Melzer’s reagent); (I–L) Ascospores. Bars: (A) = 2 mm, (B) = 500 μm, (C) = 200 μm, (D) = 20 μm, (E–H) = 20 μm, (I–L) = 10 μm.

Figure 4.
Dematophora populi (GZUH0116) (A–C) Stromata on the host; (D–F) Asci.; (G,H) Urn-shaped J+ apical rings (stained in Melzer’s reagent); (I–L) Ascospores with short cell appendage on the ends. Bars: (A) = 1000 μm, (B) = 500 μm, (C) = 200 μm, (D–L) = 10 μm.
Figure 4.
Dematophora populi (GZUH0116) (A–C) Stromata on the host; (D–F) Asci.; (G,H) Urn-shaped J+ apical rings (stained in Melzer’s reagent); (I–L) Ascospores with short cell appendage on the ends. Bars: (A) = 1000 μm, (B) = 500 μm, (C) = 200 μm, (D–L) = 10 μm.

Figure 5.
Rosellinia cainii (GZUH0119). (A,B) Stromata on the host; (C) Longitudinal section of stroma; (D,E) Urn–shaped J+ apical rings (stained in Melzer’s reagent); (F) Paraphyses; (G,H) Asci. (I–K) Ascospores. Bars: (A) = 5 mm, (B) = 1 mm, (C) = 200 μm, (D–H) = 10 μm, (I–K) = 5 μm.
Figure 5.
Rosellinia cainii (GZUH0119). (A,B) Stromata on the host; (C) Longitudinal section of stroma; (D,E) Urn–shaped J+ apical rings (stained in Melzer’s reagent); (F) Paraphyses; (G,H) Asci. (I–K) Ascospores. Bars: (A) = 5 mm, (B) = 1 mm, (C) = 200 μm, (D–H) = 10 μm, (I–K) = 5 μm.

Figure 6.
Rosellinia thailandica (MFLU12-2136, holotype). (A,B) Stromata of on the host; (C) Cross-section of a stroma; (D–F) Asci; (G,H) Asci apical rings (stained in Melzer’s reagent); (I–K) Ascospore with a germ slit and end-sheaths; (I,J stained in Melzer’s reagent; K, stained in ink). Bar: (B,C) = 500 µm, (D–K) = 10 µm.
Figure 6.
Rosellinia thailandica (MFLU12-2136, holotype). (A,B) Stromata of on the host; (C) Cross-section of a stroma; (D–F) Asci; (G,H) Asci apical rings (stained in Melzer’s reagent); (I–K) Ascospore with a germ slit and end-sheaths; (I,J stained in Melzer’s reagent; K, stained in ink). Bar: (B,C) = 500 µm, (D–K) = 10 µm.

Figure 7.
Rosellinia vitis (GZUH0123). (A–C) Stromata on the host; (D–G) Asci (H–J) Ascospores; (K) Urn-shaped J+ apical ring (stained in Melzer’s reagent). Bars: (B,C) = 200 μm, (D–J) = 20 μm, (K) = 10 μm.
Figure 7.
Rosellinia vitis (GZUH0123). (A–C) Stromata on the host; (D–G) Asci (H–J) Ascospores; (K) Urn-shaped J+ apical ring (stained in Melzer’s reagent). Bars: (B,C) = 200 μm, (D–J) = 20 μm, (K) = 10 μm.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, Q.; Long, S.; Lin, Y.; Liu, L.; Shen, X.; Kang, J. Rhizomaticola guizhouensis gen. et sp. nov. and Five Rosellinia Like Species Isolated from Decaying Wood. Diversity 2022, 14, 703. https://doi.org/10.3390/d14090703
AMA Style
Li Q, Long S, Lin Y, Liu L, Shen X, Kang J. Rhizomaticola guizhouensis gen. et sp. nov. and Five Rosellinia Like Species Isolated from Decaying Wood. Diversity. 2022; 14(9):703. https://doi.org/10.3390/d14090703
Chicago/Turabian StyleLi, Qirui, Sihan Long, Yan Lin, Lili Liu, Xiangchun Shen, and Jichuan Kang. 2022. "Rhizomaticola guizhouensis gen. et sp. nov. and Five Rosellinia Like Species Isolated from Decaying Wood" Diversity 14, no. 9: 703. https://doi.org/10.3390/d14090703
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.