
A New Genus of Short-Winged Flower Beetles with an Enlarged Antennal Scape in Mid-Cretaceous Amber from Northern Myanmar (Coleoptera: Kateretidae)


1
State Key Laboratory of Palaeobiology and Stratigraphy, Center for Excellence in Life and Paleoenvironment, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008, China
2
Nanjing College, University of Chinese Academy of Sciences, Nanjing 211135, China
3
School of Earth Sciences, University of Bristol, Life Sciences Building, Tyndall Avenue, Bristol BS8 1TQ, UK
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(1), 19; https://doi.org/10.3390/d15010019
Submission received: 25 November 2022
/
Revised: 20 December 2022
/
Accepted: 20 December 2022
/
Published: 22 December 2022
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The fossil records of Kateretidae from the Mesozoic are significant for understanding the origin and early evolution of the family. The recent discoveries of Cretaceous kateretids have provided valuable information about the palaeodiversity and morphological disparity of the taxa. Here, we establish a new genus and species of kateretid beetles, Scaporetes rectus gen. et sp. nov., based on a specimen preserved in mid-Cretaceous Kachin amber from northern Myanmar. The new species is characterized by its strongly enlarged scape and extremely small labrum. Our discovery highlights the morphological disparity of Kateretidae in the late Mesozoic.
1. Introduction
Kateretidae is a small family of beetles within the recently recognized superfamily Nitiduloidea, with a total of 106 described species in 23 genera. Most fossil kateretids are found in amber deposits, except for Amartus petrefactus Wickham, 1912, which is known from the Oligocene Florissant shales [1]. The oldest kateretid, Lebanoretes andelmani Kirejtshuk and Azar, 2008, was described from the Early Cretaceous Lebanese amber [2]. Eoceniretes yantaricus Kirejtshuk and Nel, 2008, and Heterhelus expressus Kirejtshuk and Nel, 2008, were described from Eocene Oise amber (France) [3]. The most diverse palaeofauna of Kateretidae was recorded from Mid-Cretaceous Myanmar (Kachin) amber, with seven species described to date, i.e., Furcalabratum burmanicum Poinar and Brown, 2018 [4], Polliniretes penalveri Peris and Jelínek, 2019 [5], Electrumeretes birmanicus Peris and Jelínek, 2019 [5], Cretaretes minimus Peris and Jelínek, 2020 [6], Eoceniretes antiquus Peris and Jelínek, 2020 [6], Pelretes vivificus Tihelka et al., 2021 [7], and Pelretes bicolor Zhao et al., 2022 [8].
Extant and Cretaceous species of kateretid beetles are potential pollinators of angiosperms. Some fossil species, such as El. birmanicus, C. minimus, and Eo. antiquus, have been suggested to pollinate a variety of gymnosperms, while Polliniretes penalveri was observed as a pollinator of Nymphaeaceae, an early branch of basal angiosperms [9]. Additionally, Tihelka et al. (2021) found pollen-laden coprolites of eudicots preserved with Pe. vivificus, which is direct evidence of pollen feeding behavior in Kateretidae [7]. Kateretids inhabit various biotopes, from forests to semideserts, most of which occur in the temperate and subtropical zones of both the Northern and Southern Hemispheres, excluding New Zealand [10].
Kateretidae was once classified into Cucujoidea, but recent phylogenomic studies have treated this family as a member of the newly defined ‘Nitidulidae group’, which consists of Nitidulidae, Smicripidae, and Kateretidae, in the superfamily Nitiduloidea [11,12]. Kateretidae can be distinguished from the other related families by the loose antenna club, lack of subantenna grooves, open procoxal cavities, distinct galea, and absence of outer lateral carinae on the tibae [13,14]. Among the fossil kateretids, a modified scape was observed in Eo. antiquus. In this species, the scape was enlarged in the female and strongly modified in the male, and this sexual dimorphism was thought to play a role in its mating behavior [6]. Here, we describe a new genus and a new species with an enlarged scape from Burmese amber.
2. Materials and Methods
The specimens examined in this study are housed in the Nanjing Institute of Geology and Palaeontology (NIGP200944), Chinese Academy of Sciences, Nanjing, China. The amber pieces were ground with sandpapers of different grit sizes and polished with diatomite mud [15]. Photographs were taken using three devices: a Zeiss Stereo Discovery V16 microscope system with an incident light and a transmitted light, a Zeiss Axio Imager 2 compound microscope with an AxioCam MRc 5 camera attached, and a Zeiss LSM 710 confocal laser scanning microscope (CLSM) with digital cameras attached [16,17]. The Zeiss Axio Imager 2 microscope was equipped with a mercury lamp and specific filters for DAPI, eGFP, and rhodamine. Photomicrographs with a green background were taken under the eGFP mode (Zeiss Filter Set 10; excitation/emission: 450–490/515–565 nm). The images were stacked using Helicon Focus 7.0.2 to achieve an extended depth of field. The figure plates were compiled and arranged in Adobe Photoshop 2021.
The Burmese (Kachin) amber specimen was derived from an amber mine near the Noije Bum Hill summit site, 20 km southwest of Tanai, in the Hukawng Valley, Kachin Province, northern Myanmar [18,19]. According to paleontological and radioactive data, the age of the Myanmar amber has been constrained to the Mid-Cretaceous, Albian (around 98.79 ± 0.62 Ma) [20,21]. This published work and the nomenclatural acts it contains have been registered in ZooBank, the proposed online registration system for the International Code of Zoological Nomenclature (ICZN). The ZooBank LSID for this publication is urn:lsid:zoobank.org:pub:10AB1D52-455F-496D-B333-065EDC7C1844.
3. Systematic Palaeontology
Order: Coleoptera Linnaeus, 1758
Superfamily: Nitiduloidea Latreille, 1802
Family: Kateretidae Erichson in Agassiz, 1846
Genus Scaporetes gen. nov.
(ZooBank LSID urn:lsid:zoobank.org:act:7CEC934F-E318-40C2-B308-AE4F8574A918)
Type species. Scaporetes rectus sp. nov.
Etymology. The generic name is a combination of ‘Scapo’, from the Latin ‘scapus’ (scape), and ‘-retes’, a common suffix for generic names in Kateretidae.
Diagnosis. Body elongated, with long pubescence. Head prognathous, with distinct occipital ridge. Mandible distinct dorsally and unidentate. Antennal insertions concealed; subantennal grooves absent. Antennae 11-segmented, densely setose with 3-segmented club; scape straight, strongly elongate, and widened. Pygidium and preceding two abdominal tergums exposed. Tibiae with one row of long spines at outer edge; two tibial spurs present. Tarsi 5-5-5, sparsely setose.
Description. Body elongated, lightly convex dorsally and flattened ventrally, with long pubescence. Head prognathous. Labrum small but arcuately emarginate anteriorly. Mandible small dorsally, curved, and unidentate. Antennal insertions concealed; subantennal grooves absent. Antennae 11-segmented, densely setose with loosely 3-segmented club; scape straight, strongly elongate, and widened. Terminal maxillary palpomere cylindrical; terminal labial palpomere wider than preceding ones, nearly ovate.
Pronotum quadrate punctate with dense setae; sides paralleled. Prosternum transverse; prosternal process short and narrow. Procoxal cavities open. Scutellar shield developed, triangular. Pygidium and preceding two abdominal tergites exposed. Femora shallowly caniculate for reception of tibiae. Tibiae armed with one row of long spines at outer edge; two tibial spurs present. Tarsi 5-5-5, pretarsal claws simple. Abdomen densely setose ventrally with five ventrites.
Included species. Scaporetes rectus gen. et sp. nov.
Scaporetes rectus sp. nov.
(Figure 1, Figure 2 and Figure 3; ZooBank LSID urn:lsid:zoobank.org:act:095BBDE7-09A3-4142-A19C-29B75796C8B2)
Etymology. The specific epithet is from the Latin word ‘rectus’ (meaning straight), indicating the strongly straight and enlarged scape of the new species.
Diagnosis. Body elongated, lightly convex dorsally and flattened ventrally, with long pubescence. Antennal insertions concealed; subantennal grooves absent. Antennae 11-segmented, densely setose with 3-segmented club, with oval apex at terminal of last antennomere; scape straight, strongly elongate, and widened; pedicel half the width of the scape. Pronotum quadrate, as wide as elytra, punctate with dense setae. Prosternal process short and narrow. Procoxal cavities transverse and open. Posterior angles of pronotum acutely produced posteriorly. Pygidium and preceding two abdominal tergites visible in dorsal view. Tibiae with one row of long spines at outer edge; two tibial spurs present. Tarsi 5-5-5, sparsely setose; tarsomeres 1–3 bilobed, tarsomere 4 smallest, tarsomere 5 almost as long as combined length of tarsomeres 1–3.
Holotype. NIGP200944, likely male.
Type locality and horizon. Amber mine in the Hukawng Valley, Myitkyina District, Kachin State, Myanmar; Cenomanian to Late Albian (Mid-Cretaceous) [21]; acquired in April 2016.
Description. Body elongated, length around 1.6 mm (from terminal of abdomen to apex of mandibles) and width 0.6 mm (measured from median of pronotum), lightly convex dorsally and flattened ventrally (Figure 1A,B). Body covered with dense long pubescence (Figure 1A,B).
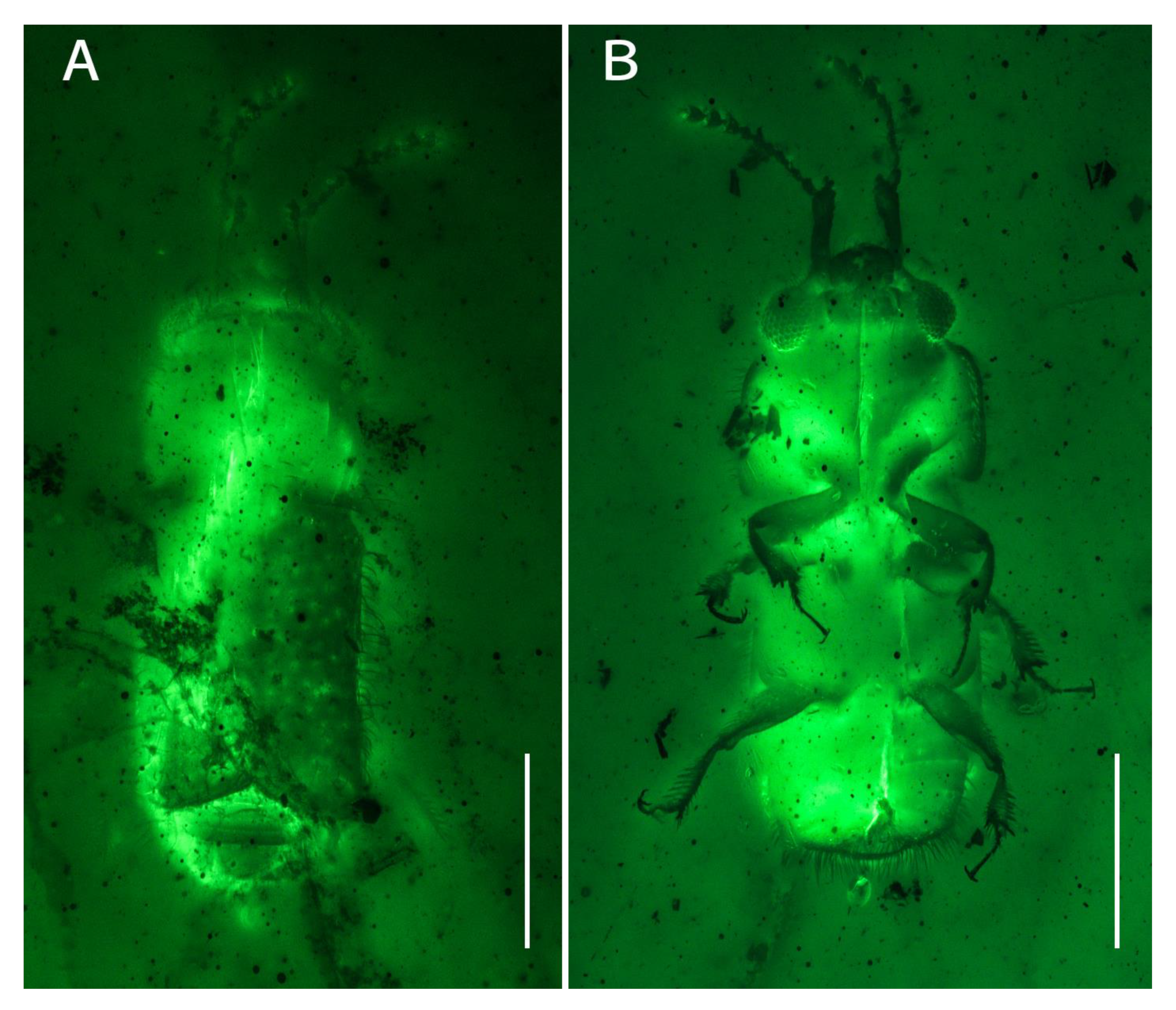
Figure 2.
Photomicrographs of Scaporetes rectus gen. et sp. nov. from mid-Cretaceous Burmese amber under green epifluorescence (holotype, NIGP200944): (A) dorsal view; (B) ventral view. Scale bars: 0.5 mm.
Figure 2.
Photomicrographs of Scaporetes rectus gen. et sp. nov. from mid-Cretaceous Burmese amber under green epifluorescence (holotype, NIGP200944): (A) dorsal view; (B) ventral view. Scale bars: 0.5 mm.

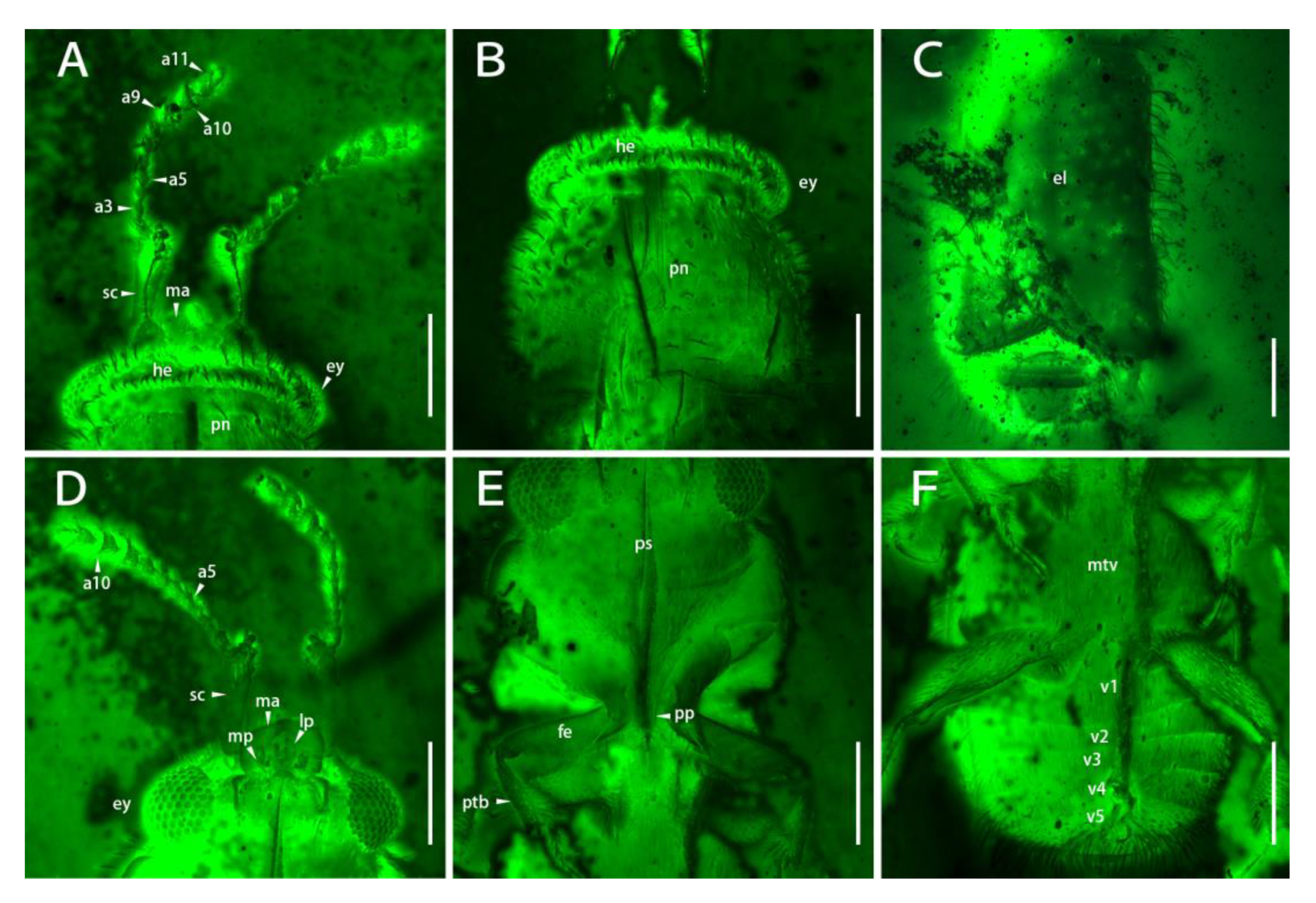
Head prognathous, with distinct occipital ridge (Figure 1A, Figure 2A, and Figure 3A,B). Eyes moderately convex, crossing temple strongly (Figure 1A,B and Figure 3A,D). Labrum free, transverse, small, but arcuately emarginate anteriorly, with emarginate anterior margin of head (Figure 1B). Mandible small dorsally, moderately curved mesally, and unidentate (Figure 1A,B, Figure 2B, and Figure 3A,D). Antennal insertions concealed; subantennal grooves absent (Figure 1B and Figure 3D). Antennae 11-segmented, densely setose with loosely 3-segmented club; antennomere 1 (scape) straight, strongly elongate, and widened, around five times as long as antennomere 2 (pedicel), with pedicel attached at terminal lateral sides; antennomere 2 half the width of the scape; antennomeres 3–8 subequal in length, nearly half of the pedicel and gradually wider; antennomeres 9–11 trapezoid, approximately 1.5 times as long and wide as preceding antennomeres, with oval apex at terminal of last antennomere (Figure 3A,D and Figure 4B). Terminal maxillary palpomere nearly cylindrical; terminal labial palpomere wider than preceding ones and nearly ovate (Figure 3D and Figure 4A).
Figure 3.
Morphological details of Scaporetes rectus gen. et sp. nov. from mid-Cretaceous Burmese amber (holotype, NIGP200944) under confocal laser scanning microscopy (CLSM): (A) dorsal view of the head; (B) pronotum; (C) elytra; (D) ventral view of the head; (E) prosternum; (F) abdomen. Abbreviations: a1–11, antennomeres 1–11; el, elytra; ey, eye; fe, femur; he, head; lp, labial palp; ma, mandible; mp, maxillary palp; mtv, metaventrite; pn, pronotum; pp, prosternal process; ps, prosternum; ptb, protibiae; sc, scape; v1–5, ventrites 1–5. Scale bars: 200 μm.
Figure 3.
Morphological details of Scaporetes rectus gen. et sp. nov. from mid-Cretaceous Burmese amber (holotype, NIGP200944) under confocal laser scanning microscopy (CLSM): (A) dorsal view of the head; (B) pronotum; (C) elytra; (D) ventral view of the head; (E) prosternum; (F) abdomen. Abbreviations: a1–11, antennomeres 1–11; el, elytra; ey, eye; fe, femur; he, head; lp, labial palp; ma, mandible; mp, maxillary palp; mtv, metaventrite; pn, pronotum; pp, prosternal process; ps, prosternum; ptb, protibiae; sc, scape; v1–5, ventrites 1–5. Scale bars: 200 μm.

Pronotum quadrate, nearly 1.9 times as wide as long, widest near middle, almost as wide as elytra; punctate with dense setae; sides paralleled; anterior angles round, posterior angles acutely produced posteriorly (Figure 1A, Figure 2A, and Figure 3B). Prosternum transverse; prosternal process short and narrow, slightly across anterior of mesoventrite (Figure 1B and Figure 3E). Procoxal cavities transverse and open (Figure 3E).
Scutellar shield developed, triangular (Figure 1A). Elytra length around 0.3 mm, pygidium and preceding two abdominal tergites exposed, separately rounded apically; disc of elytra irregularly punctate and strigose (Figure 1A, Figure 2A and Figure 3C).
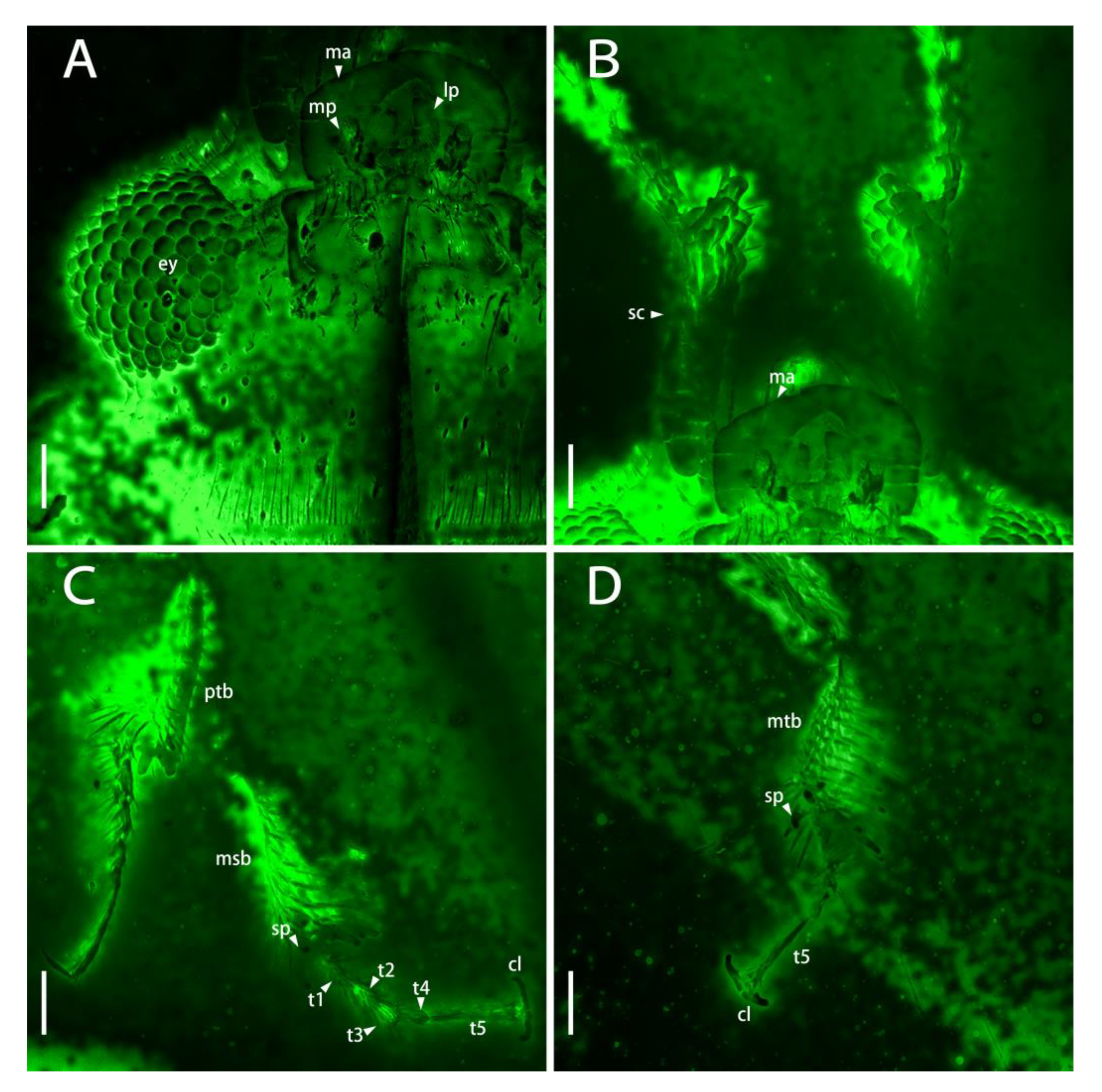
Meso- and metacoxae clearly separated (Figure 3E,F). Femora setose, shallowly caniculate for reception of tibiae (Figure 3F). Tibiae dilated apically, armed with one row of long spines at outer edge; two tibial spurs present (Figure 4C,D). Tarsi 5-5-5, sparsely setose; tarsomeres 1–3 bilobed, subequal in length and width, tarsomere 4 smallest, tarsomere 5 almost as long as combined length of tarsomeres 1–3; pretarsal claws simple (Figure 4C,D).
Abdomen densely setose ventrally with five ventrites (Figure 3F). Ventrites 1–5 gradually narrow; ventrite 1 longest, shorter than combined length of following three ventrites; ventrites 2–5 nearly subequal in length (Figure 3F).
Figure 4.
Photomicrographs of head and tibiae of Scaporetes rectus gen. and sp. nov. from Mid-Cretaceous Burmese amber (holotype, NIGP200944) under confocal laser scanning microscopy (CLSM): (A) ventral view of head; (B) ventral view of scape; (C) protibiae and mesotibiae; (D) metatibiae. Abbreviations: cl, claw; ey, eye; lp, labial palp; ma, mandible; mp, maxillary palp; msb, mesotibiae; mtb, metatibiae; ptb, protibiae; sc, scape; sp, spur; t1–5, tarsomeres 1–5. Scale bars: 50 μm.
Figure 4.
Photomicrographs of head and tibiae of Scaporetes rectus gen. and sp. nov. from Mid-Cretaceous Burmese amber (holotype, NIGP200944) under confocal laser scanning microscopy (CLSM): (A) ventral view of head; (B) ventral view of scape; (C) protibiae and mesotibiae; (D) metatibiae. Abbreviations: cl, claw; ey, eye; lp, labial palp; ma, mandible; mp, maxillary palp; msb, mesotibiae; mtb, metatibiae; ptb, protibiae; sc, scape; sp, spur; t1–5, tarsomeres 1–5. Scale bars: 50 μm.

4. Discussion
Nitidulidae is the closest sister family of Kateretidae as compared to other beetle families, so there are some similar features observed between kateretids and nitidulids, such as the elytra being short with at least the pygidium exposed; the antenna being 11-segmented with a 3-segmented club; the 5-5-5 tarsal pattern, with tarsomere 4 being smallest; and other detailed characteristics [13,14]. Given the above description, the new species described here can be classified into one of the two families. Kateretidae are distinguished from most of Nitidulidae based on characteristics such as the loosely 3-segmented antennal club, absence of antennal grooves, and lack of an outer lateral carinae on the meso- and metatibiae [13,14]. Among the above three characteristics, the absence of antennal grooves can be found in the tribe Mystropini in Nitidulidae, but there are no other features similar to Kateretids in Mystropini [22], while these features can all be found in the new species. Additionally, the open procoxal cavities in Scaporetes rectus is a trait that differs from most extant nitidulids except Cryptarchinae [13,14]. The presence of a free labrum and three exposed abdominal segments rules it out from the subfamily Cryptarchinae. Overall, Scaporetes retus is more likely to be classified into Kateretidae.
Eoceniretes is the only genus reported to have a special stalk scape. The most conspicuous difference between the two genera Eoceniretes and Saporetes is the overall shape of the scape, which is straight and large in Saporetes, while Eoceniretes has a scape with an inner flat extension behind the insertion of the pedicel. Moreover, the relative proportion of wings to the body is different. An additional key distinction is the meso- and metatibiae armed by one row of spines on the outer edge in Scaporetes, while two rows of spines on the outer ledge of the tibiae are observed in Eoceniretes.
Scaporetes rectus is most similar to Eoceniretes antiquus as compared to other fossil kateretids from Cretaceous ambers due to the particular scape, 3-segmented club with a subacute apex, open procoxal cavities, and absence of antennal grooves. However, there are still some distinct differences between Sc. rectus and Eo. antiquus [6]. Sc. rectus shows a strongly enlarged scape, being nearly 1.2 times as long as its head, while Eo. antiquus has a scape with an inner flat extension behind the insertion of the pedicel and a shovel-like reticulate with a dorsal border [6]. In addition, the antenna insertions are concealed by the anterior angles of the frons, and the angles are stout and projecting too strongly to make the head dorsally emarginate deeply in Sc. rectus. In contrast, the projecting anterior angles of the frons are absent, and the antennal insertions are just concealed by the smooth anterolateral border of the head in Eo. antiquus [6]. In Sc. rectus, the pronotum is quadrate with the parallel sides being slightly bordered, and the elytra is relatively long with a 1:2 length ratio between the elytra and body. However, the pronotum has arcuate and distinctly narrow explanate sides and short elytra, being one-third the length of the body in Eo. antiquus [6]. In Sc. rectus, the meso- and metatibiae are armed by one row of spines on the outer edge and the first three tarsomeres are bilobed, but Eo. antiquus shows two rows of spines on the lateral sides of the tibiae and simply cylindrical tarsi [6]. Additionally, the following features of Sc. rectus are all different in Eo. antiquus: an elongate body densely covered by a long pubescence, the absence of a longitudinal coxal line on the metaventrite, and a narrow prosternal process projecting across the anterior margin of the mesoventrite.
5. Conclusions
The fossil records of Kateretidae are few, and only one species, Eoceniretes antiquus, possesses a particular scape among these fossil kateretids. Here, we described a new kateretid species Scaporetes rectus sp. nov. characterized by its extremely enlarged scape, strongly projecting anterior angles to cover the antennal insertion, meso- and metatibiae armed by one row of spines, and body covered by a long pubescence. Our new species Scaporetes rectus is distinguished from Eo. antiquus by the morphological differences of the scape and anterior angles concealing the antennal fossa, the different number of spines on the tibiae, and the differing lengths of the elytra. Our discovery is the only fossil kateretid except Eo. antiquus with a special scape and provides important evidence revealing the high diversity of the morphology, especially regarding the scape, in Kateretidae in the late Mesozoic.
Author Contributions
Methodology, Q.Z.; software, Q.Z.; writing—original draft preparation, Q.Z.; writing—review and editing, C.C. and D.H.; supervision, C.C.; project administration, C.C.; funding acquisition, C.C. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support was provided by the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB26000000), the Second Tibetan Plateau Scientific Expedition and Research project (2019QZKK0706), and the National Natural Science Foundation of China (42288201, 42222201).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated during this study are included in this published article. The type specimen (NIGP200944) is housed in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China.
Acknowledgments
We are grateful to three anonymous reviewers for constructive comments on the early version of this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wickham, H.F. A Report on Some Recent Collections of Fossil Coleoptera from the Miocene Shales of Florissant. In Bulletin from the Laboratories of Natural History of the State University of Iowa; University of Iowa: Iowa City, IA, USA, 1912; pp. 3–38. [Google Scholar]
- Kirejtshuk, A.G.; Azar, D. New taxa of beetles (Insects, Coleoptera) from Lebanese amber with evolutionary and systematic comments. Alavesia 2008, 2, 15–46. [Google Scholar]
- Kirejtshuk, A.G.; Nel, A. New beetles of the suborder Polyphaga from the Lowermost Eocene French amber (Insecta: Coleoptera). Ann. Société Entomol. Fr. 2008, 44, 419–442. [Google Scholar] [CrossRef]
- Poinar, G., Jr.; Brown, A.E. Furcalabratum burmanicum gen. et sp. nov., a short-winged flower beetle (Coleoptera: Kateretidae) in mid-Cretaceous Myanmar amber. Cretac. Res. 2018, 84, 240–244. [Google Scholar] [CrossRef]
- Peris, D.; Jelínek, J. Atypical short elytra in Cretaceous short-winged flower beetles (Coleoptera: Kateretidae). Palaeoentomology 2019, 2, 505–514. [Google Scholar] [CrossRef]
- Peris, D.; Jelínek, J. Syninclusions of two new species of short-winged flower beetle (Coleoptera: Kateretidae) in mid-Cretaceous Kachin amber (Myanmar). Cretac. Res. 2020, 106, 104264. [Google Scholar] [CrossRef]
- Tihelka, E.; Li, L.Q.; Fu, Y.Z.; Su, Y.T.; Huang, D.Y.; Cai, C.Y. Angiosperm pollinivory in a Cretaceous beetle. Nat. Plants 2021, 7, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Huang, D.Y.; Cai, C.Y. Pelretes bicolor sp. nov., a new short-winged flower beetle (Coleoptera: Kateretidae) from mid-Cretaceous amber of northern Myanmar. Cretac. Res. 2022, 140, 105330. [Google Scholar] [CrossRef]
- Peris, D.; Labandeira, C.C.; Barron, E.; Delclos, X.; Rust, J.; Wang, B. Generalist pollen-feeding beetles during the mid-Cretaceous. iScience 2020, 23, 100913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hisamatsu, S. A review of the Japanese Kateretidae fauna (Coleoptera: Cucujoidea). Acta Entomol. Musei Natl. Pragae 2011, 51, 551–585. [Google Scholar] [CrossRef]
- McKenna, D.D.; Shin, S.; Ahrens, D.; Balke, M.; Beza-Beza, C.; Clarke, D.J.; Donath, A.; Escalona, H.E.; Friedrich, F.; Letsch, H.; et al. The evolutionand genomic basis of beetle diversity. Proc. Natl. Acad. Sci. USA 2019, 116, 24729–24737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.Y.; Tihelka, E.; Giacomelli, M.; Lawrence, J.F.; Slipinski, A.; Kundrata, R.; Yamamoto, S.H.; Thayer, M.K.; Newton, A.F.; Leschen, R.A.; et al. Integrated phylogenomics and fossil data illuminate the evolution of beetles. R. Soc. Open Sci. 2022, 9, 211771. [Google Scholar] [CrossRef] [PubMed]
- Jelinek, J.; Cline, A.R. Chapter 10.25 Kateretidae Erichson in Agassiz, 1846. In Handbook of Zoology, Arthropoda: Insecta, Coleoptera, Beetles Morphology and Systematics; Leschen, R.A.B., Beutel, R.G., Lawrence, J.F., Eds.; De Gruyter: Berlin/Heidelberg, Germany, 2010; pp. 386–390. [Google Scholar] [CrossRef]
- Jelinek, J.; Carlton, C.; Cline, A.R.; Leschen, R.A.B. Chapter 10.26 Nitidulidae Latreille, 1802. In Coleoptera, Beetles, Volume 2, Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim); De Gruyter: Berlin/Heidelberg, Germany, 2011; pp. 390–407. [Google Scholar] [CrossRef]
- Sidorchuk, E.A.; Vorontsov, D.D. Preparation of small-sized 3D amber samples: State of the technique. Palaeoentomology 2018, 1, 80–90. [Google Scholar] [CrossRef]
- Cai, C.Y.; Huang, D.Y. The oldest micropepline beetle from Cretaceous Burmese amber and its phylogenetic implications (Coleoptera: Staphylinidae). Naturwissenschaften 2014, 101, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.Z.; Li, Y.D.; Su, Y.T.; Cai, C.Y.; Huang, D.Y. Application of confocal laser scanning microscopy to the study of amber bioinclusions. Palaeoentomology 2021, 4, 266–278. [Google Scholar] [CrossRef]
- Cruickshank, R.; Ko, K. Geology of an amber locality in the Hukawng Valley, northern Myanmar. J. Asian Earth Sci. 2003, 21, 441–455. [Google Scholar] [CrossRef]
- Yin, Z.W.; Cai, C.Y.; Huang, D.Y. Last major gap in scydmaenine evolution filled (Coleoptera: Staphylinidae). Cretac. Res. 2018, 84, 62–68. [Google Scholar] [CrossRef]
- Shi, G.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.C.; Lei, W.Y.; Li, Q.L.; Li, X.H. Age constraint on Burmese amber based on U-Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Mao, Y.Y.; Liang, K.; Su, Y.T.; Li, J.G.; Rao, X.; Zhang, H.; Xia, F.Y.; Fu, Y.Z.; Cai, C.Y.; Huang, D.Y. Various amberground marine animals on Burmese amber with discussions on its age. Palaeoentomology 2018, 1, 91–103. [Google Scholar] [CrossRef]
- Kirejtshuk, A.G.; Jelinek, J. Preliminary review of genera of the tribe Mystropini with redescriptions and new descriptions of some genera, subgenera and species (Coleoptera: Nitidulidae: Nitidulinae). Folia Heyrovskyana 2000, 8, 171–192. [Google Scholar]
Figure 1.
Photomicrographs of Scaporetes rectus gen. et sp. nov. from mid-Cretaceous Burmese amber under normal reflected light (holotype, NIGP200944): (A) dorsal view; (B) ventral view. Scale bars: 0.5 mm.
Figure 1.
Photomicrographs of Scaporetes rectus gen. et sp. nov. from mid-Cretaceous Burmese amber under normal reflected light (holotype, NIGP200944): (A) dorsal view; (B) ventral view. Scale bars: 0.5 mm.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, Q.; Huang, D.; Cai, C. A New Genus of Short-Winged Flower Beetles with an Enlarged Antennal Scape in Mid-Cretaceous Amber from Northern Myanmar (Coleoptera: Kateretidae). Diversity 2023, 15, 19. https://doi.org/10.3390/d15010019
AMA Style
Zhao Q, Huang D, Cai C. A New Genus of Short-Winged Flower Beetles with an Enlarged Antennal Scape in Mid-Cretaceous Amber from Northern Myanmar (Coleoptera: Kateretidae). Diversity. 2023; 15(1):19. https://doi.org/10.3390/d15010019
Chicago/Turabian StyleZhao, Qian, Diying Huang, and Chenyang Cai. 2023. "A New Genus of Short-Winged Flower Beetles with an Enlarged Antennal Scape in Mid-Cretaceous Amber from Northern Myanmar (Coleoptera: Kateretidae)" Diversity 15, no. 1: 19. https://doi.org/10.3390/d15010019
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.