
Three New Species of Jainus (Monogenea: Dactylogyridae) Parasitizing Gills of Brazilian Freshwater Fishes Supported by Morphological and Molecular Data †
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Host Sampling and Parasitological Procedures
2.2. Molecular and Phylogenetic Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monogenea Species | Host Species | Locality | Accession Numbers | References | |
|---|---|---|---|---|---|
| LSU rDNA | COI | ||||
| Dactylogyridae | |||||
| Ameloblastella chavarriai (Price, 1938) | Rhamdia quelen | Mexico | KP056251 | [6] | |
| Ameloblastella edentensis Mendoza-Franco, Mendoza-Palmero & Scholz, 2020 | Hypophthalmus edentatus | Peru | KP056255 | [6] | |
| Ameloblastella unapioides Mendoza-Franco, Mendoza-Palmero & Scholz, 2020 | Sorubim lima | Peru | KP056254 | [6] | |
| Aphanoblastella aurorae Mendoza-Palmero, Scholz, Mendoza-Franco & Kuchta, 2012 | Goeldiella eques | Peru | KP056239 | [6] | |
| Aphanoblastella magna Yamada, Acosta, Yamada, Scholz & Silva, 2018 | Pimelodella avanhandavae | Brazil | MH688484 | [32] | |
| Aphanoblastella travassosi (Price, 1938) | Rhamdia guatemalensis | Mexico | MK358458 | [9] | |
| Cacatuocotyle papilionis Zago, Franceschini, Müller & Silva, 2018 | Astyanax lacustris | Brazil | MG832889 | [33] | |
| Characithecium paranapanemense Zago, Franceschini, Abdallah, Müller, Azevedo & Silva, 2021 | Psalidodon paranae | Brazil | MZ408907 | [34] | |
| Cosmetocleithrum bifurcum Mendoza-Franco, Mendoza-Palmero & Scholz, 2000 | Hassar sp. | Peru | KP056217 | [6] | |
| Cosmetocleithrum bulbocirrus Kritsky, Thatcher & Boeger, 1986 | Pterodoras granulosus | Brazil | MG001326 | [9] | |
| Demidospermus prolixus Franceschini, Zago, Müller, Francisco, Takemoto & Silva, 2017 | Loricaria prolixa | Brazil | KY766955 | [5] | |
| Demidospermus anus Suriano, 1983 | Loricariichthys platymetopon | Brazil | KY766957 | [5] | |
| Demidospermus mortenthaleri Mendoza-Palmero, Scholz, Mendoza-Franco & Kuchta, 2012 | Brachyplatystoma juruense | Peru | KP056245 | [6] | |
| Demidospermus rhinelepisi Acosta, Scholz, Blasco-Costa, Alves & Silva, 2019 | Rhinelepis aspera | Brazil | MG001324 | [9] | |
| Demidospermus spirophallus Franceschini, Zago, Müller, Francisco, Takemoto & Silva, 2017 | Loricaria prolixa | Brazil | KY766954 | [5] | |
| Diaphorocleidus magnus Zago, Franceschini, Abdallah, Müller, Azevedo & Silva, 2021 | Astyanax lacustris | Brazil | MZ408903 | MZ408253 | [34] |
| Diaphorocleidus neotropicalis Zago, Franceschini, Abdallah, Müller, Azevedo & Silva, 2021 | Astyanax lacustris | Brazil | MZ408906 | MZ408254 | [34] |
| Diaphorocleidus petrosusi Mendoza-Franco, Aguirre-Macedo & Vidal-Martínez, 2007 | - | Panama | MF939878 | [35] | |
| Heteropriapulus anchoradiatus Acosta, Franceschini, Zago, Scholz & Silva, 2017 | Pterygoplichthys ambrosettii | Brazil | MF116371 | [7] | |
| Heteropriapulus heterotylus Acosta, Franceschini, Zago, Scholz & Silva, 2017 | Pterygoplichthys ambrosettii | Brazil | MF116370 | [7] | |
| Heteropriapulus simplex Acosta, Franceschini, Zago, Scholz & Silva, 2017 | Pterygoplichthys ambrosettii | Brazil | MF116372 | [7] | |
| Heteropriapulus sp. | Pterygoplichthys ambrosettii | Brazil | MF116373 | [7] | |
| Jainus beccus n. sp. | Leporinus friderici | Brazil | OQ833545 | Present study | |
| Jainus piava Karling, Bellay, Takemoto & Pavanelli, 2011 | Schizodon nasutus | Brazil | OQ843019 | OQ833543 | Present study |
| Jainus radixelongatus n. sp. | Leporinus striatus | Brazil | OQ843018 | OQ833544 | Present study |
| Nanayella aculeatrium Acosta, Mendoza-Palmero, Silva & Scholz, 2019 | Sorubim lima | Peru | KP056228 | [6] | |
| Nanayella amplofalcis Acosta, Mendoza-Palmero, Silva & Scholz, 2019 | Hemisorubim platyrhynchos | Brazil | MG001325 | [9] | |
| Nanayella fluctuatrium Acosta, Mendoza-Palmero, Silva & Scholz, 2019 | Sorubim lima | Brazil | MG001327 | [9] | |
| Nanayella megorchis Acosta, Mendoza-Palmero, Silva & Scholz, 2019 | Sorubim lima | Peru | MK367407 | [9] | |
| Nanayella processusclavis Acosta, Mendoza-Palmero, Silva & Scholz, 2019 | Hemisorubim platyrhynchos | Brazil | MG001328 | [9] | |
| Parasciadicleithrum octofasciatum Mendoza-Palmero, Blasco-Costa, Hernández-Mena & Pérez-Ponce de León, 2017 | Rocio octofasciata | Mexico | KY305885 | [36] | |
| Sciadicleithrum meekii Mendoza-Palmero, Blasco-Costa, Hernández-Mena & Pérez-Ponce de León, 2017 | Thorichthys meeki | Mexico | KY305889 | [36] | |
| Sciadicleithrum mexicanum Mendoza-Palmero, Blasco-Costa, Hernández-Mena & Pérez-Ponce de León, 2017 | Rocio octofasciata | Mexico | KY305886 | [36] | |
| Sciadicleithrum panamensis Mendoza-Franco, Aguirre-Macedo & Vidal-Martínez, 2007 | - | Panama | MF939864 | [35] | |
| Sciadicleithrum splendidae Kritsky, Vidal-Martínez & Rodriguez-Canul, 1994 | Parachromis friedrichsthalii | Mexico | KY305890 | [36] | |
| Trinigyrus anthus Franceschini, Acosta, Zago, Müller & Silva, 2020 | Hypostomus regani | Brazil | MN947622 | MN916719 | [37] |
| Trinigyrus carvalhoi Franceschini, Acosta, Zago, Müller & Silva, 2020 | Hypostomus ancistroides | Brazil | MN947608 | MN922321 | [37] |
| Trinigyrus peregrinus Nitta & Nagasawa, 2016 | Pterygoplychthys ambrosettii | Brazil | MN944890 | MN913212 | [37] |
| Trinigyrus peregrinus Nitta & Nagasawa, 2016 | Pterygoplichthys disjunctivus | Japan | LC104308 | [38] | |
| Unibarra paranoplatensis Suriano & Incorvaia, 1995 | Aguarunichthys torosus | Peru | KP056219 | [6] | |
| Unilatus unilatus Mizelle & Kritsky, 1967 | Pterygoplichthys ambrosettii | Brazil | MF102106 | [7] | |
| Urocleidoides cultellus Mendoza-Franco & Reina, 2008 | - | Panama | MF939848 | [35] | |
| Urocleidoides digitabulum Zago, Yamada, Yamada, Franceschini, Bongiovani & Silva, 2020 | Megaleporinus elongatus | Brazil | MT556796 | [39] | |
| Urocleidoides indianensis Oliveira, Silva, Vieira & Acosta, 2020 | Parodon nasus | Brazil | OK482868 | [40] | |
| Urocleidoides paradoni Oliveira, Silva, Vieira & Acosta, 2020 | Parodon nasus | Brazil | OK482867 | [40] | |
| Urocleidoides paradoxus Kritsky, Thatcher & Boeger, 1986 | Leporinus friderici | Brazil | MT556795 | [39] | |
| Urocleidoides sinus Zago, Yamada, Yamada, Franceschini, Bongiovani & Silva, 2020 | Schizodon nasutus | Brazil | MT556799 | [39] | |
| Urocleidoides tenuis Zago, Yamada, Yamada, Franceschini, Bongiovani & Silva, 2020 | Parodon nasus | Brazil | OK465455 | [40] | |
| Urocleidoides uncinus Zago, Yamada, Yamada, Franceschini, Bongiovani & Silva, 2020 | Gymnotus inaequilabiatus | Brazil | MT556798 | MT594473 | [39] |
| Vancleaveus janauacaensis Kritsky, Thatcher & Boeger, 1986 | Pterodoras granulosus | Peru | KP056247 | [6] | |
| Walteriella conica Mendoza-Palmero, Mendoza-Franco, Acosta & Scholz, 2019 | Brachyplatystoma juruense | Peru | MK834513 | [41] | |
| Walteriella ophiocirrus Mendoza-Palmero, Mendoza-Franco, Acosta & Scholz, 2019 | Platystomatichthys sturio | Peru | MK834515 | [41] | |
| Diplectanidae | |||||
| Murraytrema pricei Caballero, Bravo & Grocott, 1955 | Nibea albiflora | China | DQ157672 | [42] | |
| Pseudorhabdosynochus epinepheli (Yamaguti, 1938) | Epinephelus brunneus | China | AY553622 | [43] | |
| Pseudorhabdosynochus lantauensis (Beverley-Burton & Suriano, 1981) | Epinephelus brunneus | China | AY553624 | [43] |
3. Results
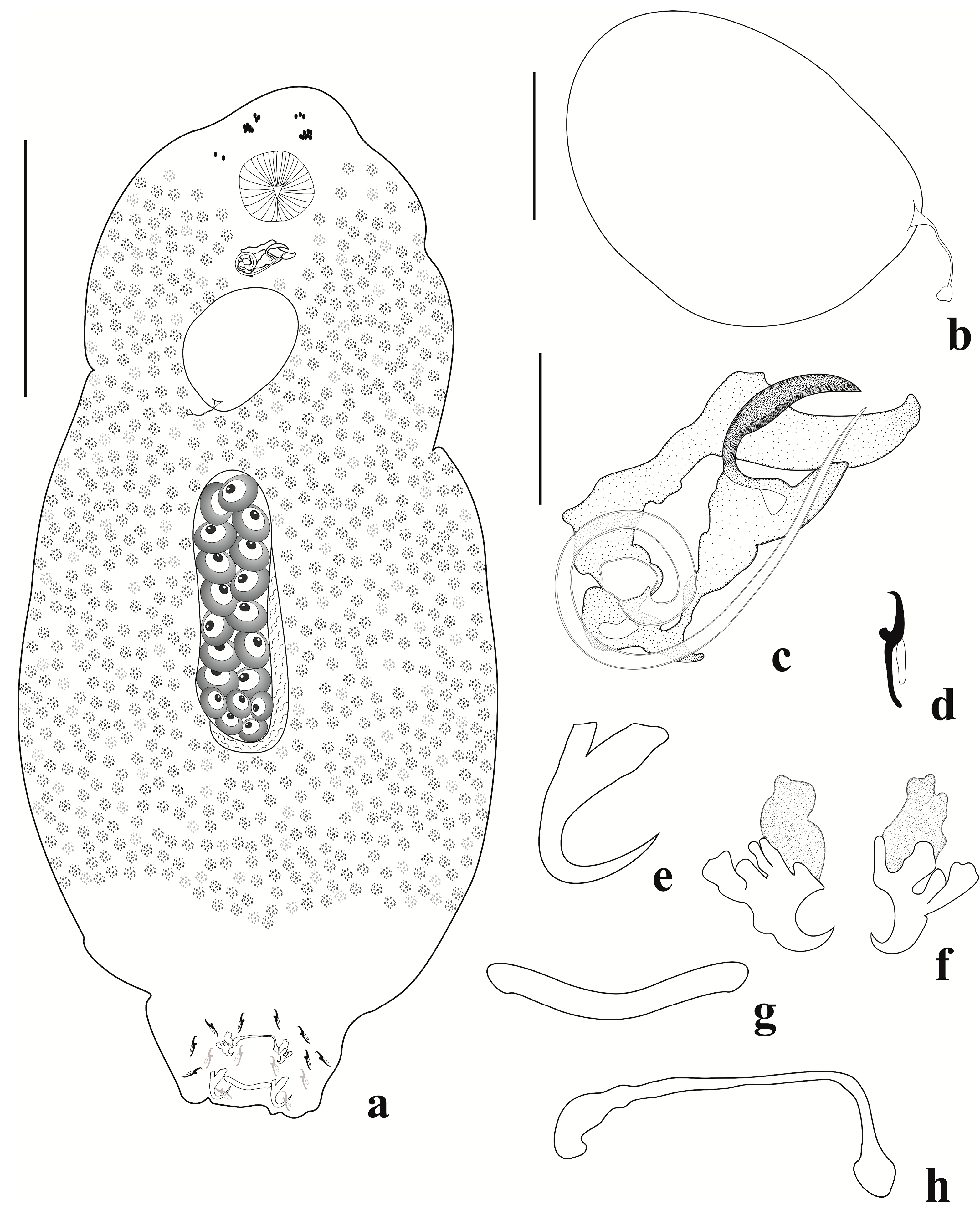
3.1. Description of Jainus beccus n. sp.
3.1.1. Taxonomic Summary
3.1.2. Description (Based on 30 Specimens: 15 Mounted in Gray and Wess’s Medium, and 15 Mounted in Hoyer’s Medium)
3.1.3. Remarks
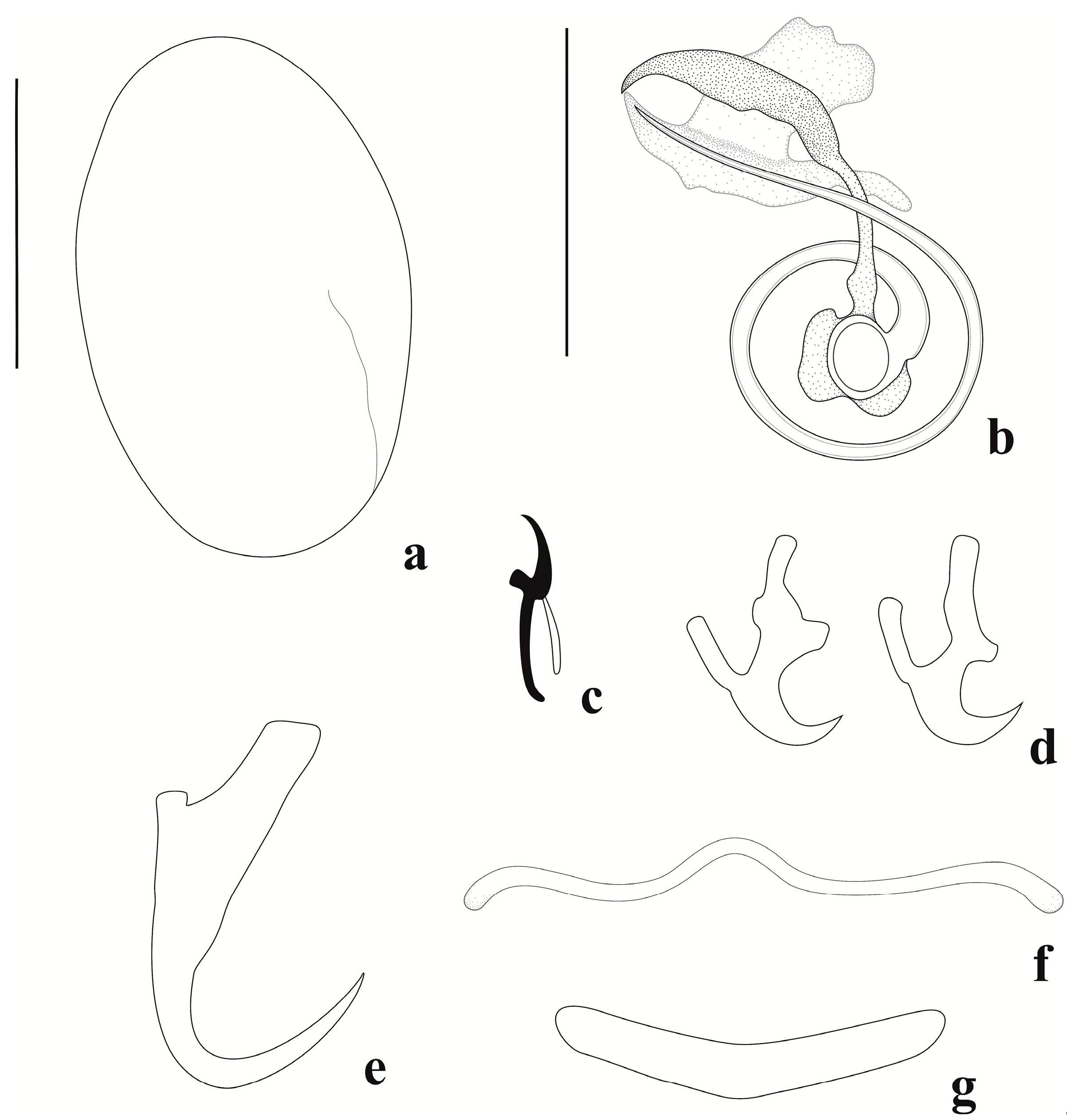
3.2. Description of Jainus ornatus n. sp.
3.2.1. Taxonomic Summary
3.2.2. Description (Based on 23 Specimens Mounted in Hoyer’s Medium)
3.2.3. Remarks
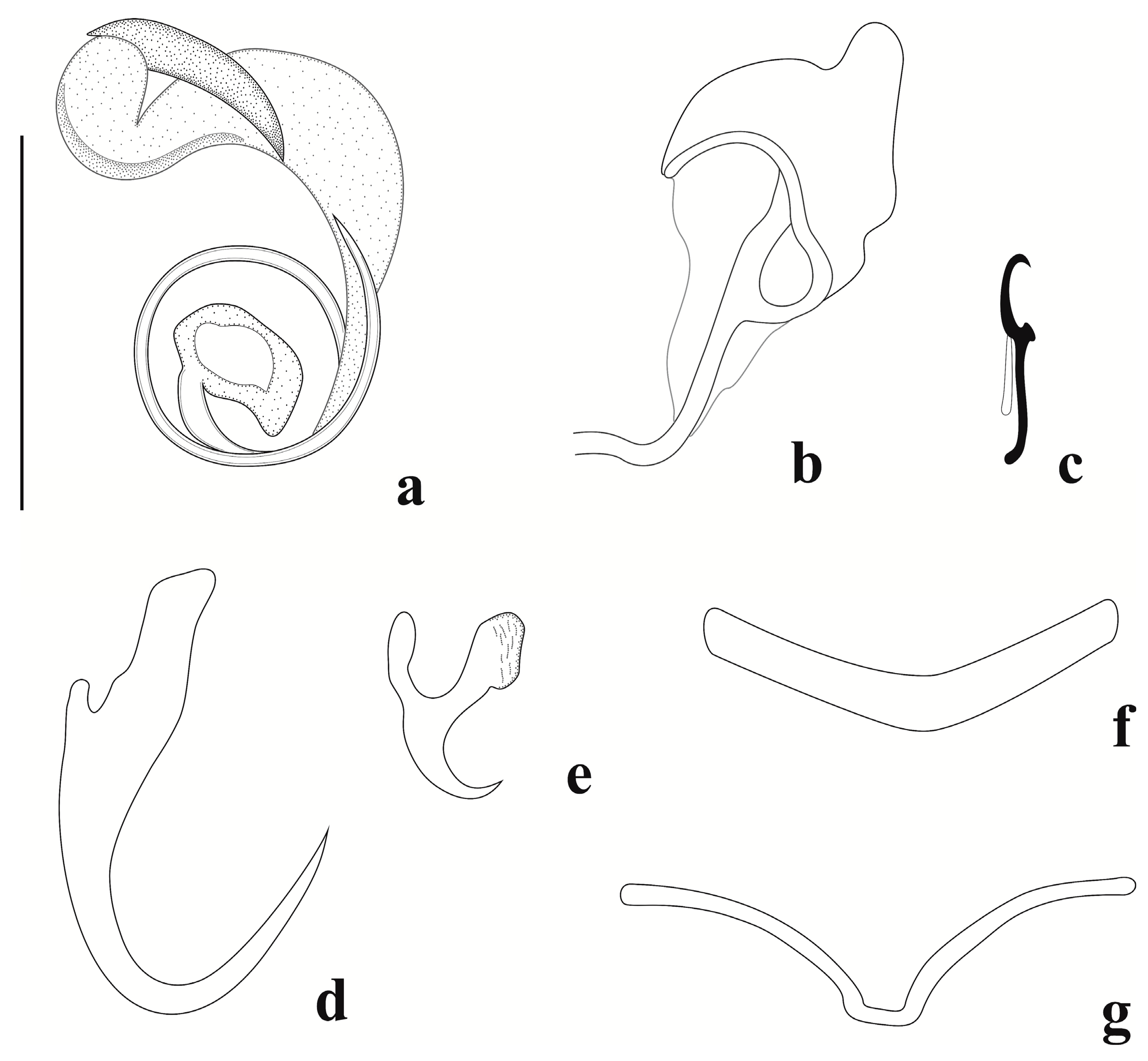
3.3. Description of Jainus radixelongatus n. sp.
3.3.1. Taxonomic Summary
3.3.2. Description (Based on Nine Specimens Mounted in Gray and Wess’s Medium)
3.3.3. Remarks
3.4. Supplementary Observations of Jainus piava Karling, Bellay, Takemoto & Pavanelli, 2011
3.4.1. Taxonomic Summary
3.4.2. Supplementary Observations (Based on 30 Specimens Mounted in Gray and Wess’s Medium)
3.4.3. Remarks
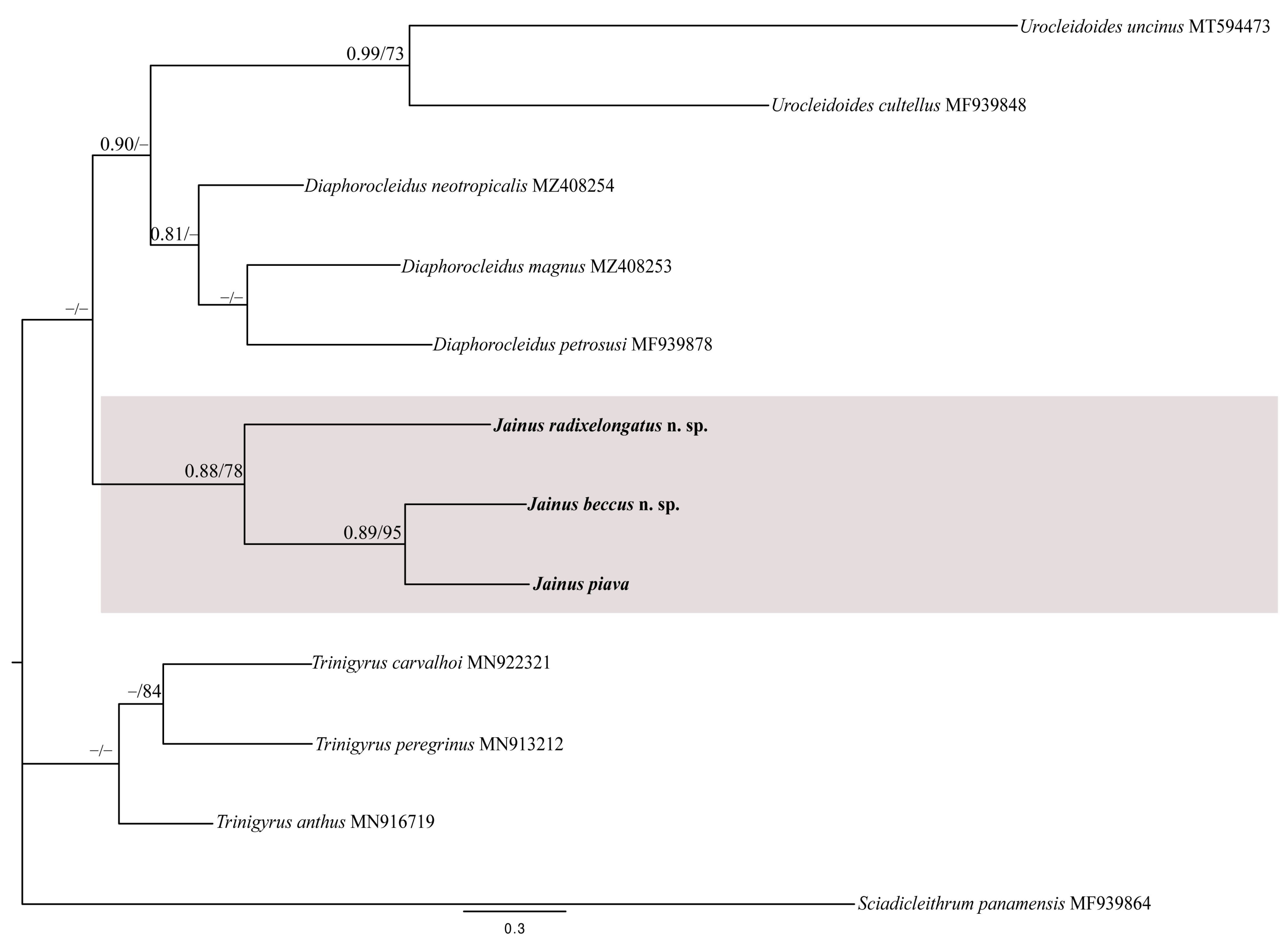
3.5. Molecular Analyses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eiras, J.C.; Takemoto, R.M.; Pavanelli, G.C.; Adriano, E.A. About the biodiversity of parasites of freshwater fish from Brazil. Fish Pathol. 2011, 31, 161–168. [Google Scholar]
- Kritsky, D.C.; Boeger, W.A.; Jegu, M. Neotropical Monogenoidea. 28. Ancyrocephalinae (Dactylogyridae) of Piranha and Their Relatives (Teleostei, Serrasalmidae) from Brazil and French Guiana: Species of Notozothecium Boeger and Kritsky, 1988, and Mymarothecium gen. n. J. Helminthol. 1996, 63, 153–175. [Google Scholar]
- Almeida, K.S.S.; Cohen, S.C. Diversidade de Monogenea (Platyhelminthes) parasitos de Astyanax altiparanae do reservatório da Usina Hidrelétrica de Itaipu. Saúde Ambiente em Revista. 2011, 6, 31–41. [Google Scholar]
- Cohen, S.C.; Kohn, A.; Justo, M.C.N. South American Monogenoidea Parasites of Fishes, Amphibians and Reptiles; Oficina de Livros: Rio de Janeiro, Brazil, 2013; 663p. [Google Scholar]
- Franceschini, L.; Zago, A.C.; Müller, M.I.; Francisco, C.J.; Takemoto, R.M.; Silva, R.J. Morphology and molecular characterization of Demidospermus spirophallus n. sp., D. prolixus n. sp. (Monogenea: Dactylogyridae) and a redescription of D. anus in siluriform catfish from Brazil. J. Helminthol. 2017, 92, 228–243. [Google Scholar] [CrossRef]
- Mendoza-Palmero, C.A.; Blasco-Costa, I.; Scholz, T. Molecular phylogeny of Neotropical monogeneans (Platyhelminthes: Monogenea) from catfishes (Siluriformes). Parasites Vectors 2015, 8, 1–11. [Google Scholar] [CrossRef]
- Acosta, A.A.; Franceschini, L.; Zago, A.C.; Scholz, T.; Silva, R.J. Six new species of Heteropriapulus (Monogenea: Dactylogyridae) from South American fishes with an amended diagnosis to the genus. Zootaxa 2017, 4290, 459–482. [Google Scholar] [CrossRef]
- Acosta, A.A.; Scholz, T.; Blasco-Costa, I.; Alves, P.V.; Silva, R.J. A new genus and two new species of dactylogyrid monogeneans from gills of Neotropical catfishes (Siluriformes: Doradidae and Loricariidae). Parasitol. Int. 2018, 67, 4–12. [Google Scholar] [CrossRef]
- Acosta, A.A.; Mendoza-Palmero, C.A.; Silva, R.J.; Scholz, T. A new genus and four new species of dactylogyrids (Monogenea), gill parasites of pimelodid catfishes (Siluriformes: Pimelodidae) in South America and the reassignment of Urocleidoides megorchis Mizelle et Kritsky, 1969. Folia Parasitol. 2019, 66, 1–12. [Google Scholar] [CrossRef]
- Poulin, R. The evolution of monogenean diversity. Int. J. Parasitol. 2002, 37, 245–254. [Google Scholar] [CrossRef]
- Braga, M.P.; Araújo, S.B.L.; Boeger, W.A. Patterns of interaction between Neotropical freshwater fishes and their gill Monogenoidea (Platyhelminthes). Parasitol. Res. 2014, 113, 481–490. [Google Scholar] [CrossRef]
- Mizelle, J.D.; Kritsky, D.C.; Crane, J.W. Studies on Monogenetic Trematodes. XXXVIII. Ancyrocephalinae from South America with the proposal of Jainus gen. n. Am. Midl. Nat. 1968, 80, 186–198. [Google Scholar] [CrossRef]
- Kritsky, D.C.; Thatcher, V.E.; Kayton, R.J. Neotropical Monogenea. 3. Five new species from South America with the proposal of Tereancistrum gen. n. and Trinibaculum gen. n. (Dactylogyridae: Ancyrocephalinae). Acta Amaz. 1980, 10, 411–417. [Google Scholar] [CrossRef]
- Karling, L.C.; Bellay, S.; Takemoto, R.M.; Pavanelli, G.C. A new species of Jainus (Monogenea), gill parasite of Schizodon borellii (Characiformes, Anostomidae) from the upper Paraná river floodplain, Brazil. Acta Sci. Biol. Sci. 2011, 33, 227–231. [Google Scholar] [CrossRef]
- Cohen, S.C.; Kohn, A.; Boeger, W.A. Neotropical Monogenoidea. 57. Nine new species of Dactylogyridae (Monogenoidea) from the gill of Salminus brasiliensis (Characidae, Characiformes) from the Paraná River, State of Paraná, Brazil. Zootaxa 2012, 3049, 57–68. [Google Scholar] [CrossRef]
- Abdallah, V.D.; Azevedo, R.K.; Luque, L.F. Three new species of Monogenea (Platyhelminthes) parasites of fish in the Guandu river, southeastern Brazil. Acta Sci. Biol. Sci. 2012, 34, 483–490. [Google Scholar] [CrossRef]
- Kritsky, D.C.; Thatcher, V.E.; Boeger, W.A. Neotropical Monogenea. 8. Revision of Urocleidoides (Dactylogyridae, Ancyrocephalinae). Proc. Helminthol. Soc. Wash. 1986, 53, 1–37. [Google Scholar]
- Kritsky, D.C.; Boeger, W.A.; Thatcher, V.E. Neotropical Monogenea. 7. Parasites of the Pirarucu, Arapaima gigas (Cuvier), with descriptions of two new species and redescription of Dawestrema cycloancistrium Price and Nowlin, 1967 (Dactylogyridae: Ancyrocephalinae). Proc. Biol. Soc. Wash. 1985, 98, 321–331. [Google Scholar]
- Mizelle, J.D. New species of trematodes from the gills of Illinois fishes. Am. Midl. Nat. 1936, 17, 785–806. [Google Scholar] [CrossRef]
- Bush, A.O.; Laferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitolgy meets ecology on its own terms: Margolis revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Pleijel, F.; Jondelius, U.; Norlinder, E.; Nygren, A.; Oxelman, B.; Schander, C.; Sundberg, P.; Thollesson, M. Phylogenies without roots? A plea for the use of vouchers in molecular phylogenetic studies. Mol. Phylogenet. Evol. 2008, 48, 369–371. [Google Scholar] [CrossRef]
- Plaisance, L.; Rousset, V.; Morand, S.; Littlewood, T.D.J. Colonization of Pacific islands by parasites of low dispersal ability: Phylogeography of two monogenean species parasitizing butterfly fishes in the South Pacific Ocean. J. Biogeogr. 2008, 35, 65–87. [Google Scholar] [CrossRef]
- Lockyer, A.E.; Olson, P.D.; Littlewood, D.T.J. Utility of complete large and small subunit rRNA genes in resolving the phylogeny of the Neodermata (Platyhelminthes): Implications and a review of the cercomer theory. Biol. J. Linn. Soc. 2003, 78, 155–171. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Miller, M.A.; Schwartz, T.; Pickett, B.E.; He, S.; Klem, E.B.; Scheuermann, R.H.; Passarotti, M.; Kaufman, S.; O’Leary, M.A. A RESTful API for Access to Phylogenetic Tools via the CIPRES Science Gateway. Evol. Bioinform. 2015, 16, 43–48. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1.4. Molecular Evolution, Phylogenetics and Epidemiology. 2012. Available online: https://tree.bio.ed.ac.uk/software/figtree/ (accessed on 8 August 2022).
- Yamada, F.H.; Acosta, A.A.; Yamada, P.O.F.; Scholz, T.; Silva, R.J. A new species of Aphanoblastella Kritsky, Mendoza-Franco and Scholz, 2000 (Monogenea, Dactylogyridae) parasitic on heptapterid catfish (Siluriformes) in the Neotropical region. Acta Parasitol. 2018, 63, 772–780. [Google Scholar] [CrossRef]
- Zago, A.C.; Franceschini, L.; Müller, M.I.; Silva, R.J. A new species of Cacatuocotyle (Monogenea, Dactylogyridae) parasitizing Astyanax spp. (Characiformes, Characidae) from Brazil, including molecular data and a key to species identification. Acta Parasitol. 2018, 63, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Zago, A.C.; Franceschini, L.; Abdallah, V.D.; Müller, M.I.; Azevedo, R.K.; Silva, R.J. Morphological and molecular data of new species of Characithecium and Diaphorocleidus (Monogenea: Dactylogyridae) from Neotropical characid fishes. Parasitol. Int. 2021, 84, 102406. [Google Scholar] [CrossRef] [PubMed]
- Alda, F.; Reina, R.G.; Mendoza, E.; Rios, E.; Torchin, M.E. Freshwater Fish Parasite Barcoding Project (Panama. Museum of Natural Science); Louisiana State University: Baton Rouge, LA, USA, 2017. [Google Scholar]
- Mendoza-Palmero, C.A.; Blasco-Costa, I.; Hernandez-Mena, D.; Perez-Ponce de Leon, G. Parasciadicleithrum octofasciatum n. gen., n. sp. (Monogenoidea: Dactylogyridae), parasite of Rocio octofasciata (Regan) (Cichlidae: Perciformes) from Mexico characterised by morphological and molecular evidence. Parasitol. Int. 2017, 66, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, L.; Acosta, A.A.; Zago, A.C.; Müller, M.I.; Silva, R.J. Trinigyrus spp. (Monogenea: Dactylogyridae) from Brazilian catfishes: New species, molecular data and new morphological contributions to the genus. J. Helminthol. 2020, 94, E126. [Google Scholar] [CrossRef] [PubMed]
- Nitta, M.; Nagasawa, K. Four Alien Monogeneans, Including Trinigyrus peregrinus n. sp., Parasitic on the Invasive Armored Catfish Pterygoplichthys disjunctivus (Siluriformes: Loricariidae) from Okinawa-jima Island, Okinawa Prefecture, Japan. Spec. Div. 2016, 21, 95–104. [Google Scholar] [CrossRef]
- Zago, A.C.; Yamada, F.H.; Yamada, P.O.F.; Franceschini, L.; Bongiovani, M.F.; Silva, R.J. Seven new species of Urocleidoides (Monogenea: Dactylogyridae) from Brazilian fishes supported by morphological and molecular data. Parasitol. Res. 2020, 119, 3255–3283. [Google Scholar] [CrossRef]
- Oliveira, M.S.B.; Santos-Neto, J.F.; Tavares-Dias, M.; Domingues, M.V. New species of Urocleidoides (Monogenoidea: Dactylogyridae) from the gills of two species of Anostomidae from the Brazilian Amazon. Braz. J. Vet. Parasitol. 2020, 29, e007820. [Google Scholar] [CrossRef]
- Mendoza-Palmero, C.A.; Mendoza-Franco, E.F.; Acosta, A.A.; Scholz, T. Walteriella n. g. (Monogenoidea: Dactylogyridae) from the gills of pimelodid catfishes (Siluriformes: Pimelodidae) from the Peruvian Amazonia based on morphological and molecular data. Syst. Parasitol. 2019, 96, 441–452. [Google Scholar] [CrossRef]
- Wu, X.Y.; Zhu, X.Q.; Xie, M.Q.; Li, A.X. The radiation of Haliotrema (Monogenea: Dactylogyridae: Ancyrocephalinae): Molecular evidence and explanation inferred from LSU rDNA sequences. Parasitology 2006, 132, 659–668. [Google Scholar] [CrossRef]
- Wu, X.Y.; Li, A.X.; Zhu, X.Q.; Xie, M.Q. Description of Pseudorhabdosynochus seabassi sp. n. (Monogenea: Diplectanidae) from Lates calcarifer and revision of the phylogenetic position of Diplectanum grouperi (Monogenea: Diplectanidae) based on rDNA sequence data. Folia Parasitol. (Praha). 2005, 52, 231–240. [Google Scholar] [CrossRef]
- Kritsky, D.C.; Leiby, P.D. Dactylogyridae (Monogenea) from the Freshwater Fish, Astyanax fasciatus (Cuvier), in Costa Rica, with Descriptions of Jainus hexops sp. n., Urocleidoides costaricensis, and U. heteroancistrium combs. n. Proc. Helminthol. Soc. Wash. 1972, 39, 227–230. [Google Scholar]







| Host Species | Locality | Sampled Hosts | SL (cm) ± SD (Range) |
|---|---|---|---|
| Leporinus friderici | Jurumirim Reservoir | 116 | 16.67 ± 5.79 (8.90–27) |
| Sapucaí-Mirim River | 40 | 23.12 ± 4.59 (12.70–31.80) | |
| Leporinus octofasciatus | Jurumirim Reservoir | 15 | 14.65 ± 3.77 (9.80–20.90) |
| Sapucaí-Mirim River | 17 | 18.50 ± 3.09 (13.50–22.20) | |
| Leporinus striatus | Jurumirim Reservoir | 26 | 10.12 ± 0.85 (8.70–11.70) |
| Megaleporinus elongatus | Jurumirim Reservoir | 30 | 13.79 ± 3.97 (9.50–28.20) |
| Sapucaí-Mirim River | 22 | 25.05 ± 4.26 (12–33.50) | |
| Schizodon nasutus | Jurumirim Reservoir | 30 | 16.18 ± 4.23 (9.60–24) |
| Sapucaí-Mirim River | 40 | 20.11 ± 0.73 (9.50–28.80) |
| Gene/Region | Primers and Sequences 5′–3′ | Cycling Conditions | Product (bp) | Source |
|---|---|---|---|---|
| COI | COI_Mono_5 5′-TAA TWG GTG GKT TTG GTAA-3′ COI_Mono_3 5′-TAA TGC ATM GGA AAA AAA CA-3′ | Initial denaturation of 3 min at 94 °C; 40 cycles of: 94 °C for 30 s, 47 °C for 30 s, 72 °C for 60 s; final extension of 7 min at 72 °C. | ~650 | [22] |
| LSU | U178 5′-GCA CCC GCT GAA YTT AAG-3′ L1642 5′-CCA GCG CCA TCC ATT TTC A-3′ 6F 5′-CGG CGA GTG AAC GGA GAT TA-3′ 382F 5′-AGC TGG TGG AGT CAA GCT TC-3′ 1289R 5′-TGC TCA CGT TTG ACG ATC GA-3′ | Initial denaturation of 5 min at 95 °C; 40 cycles of: 95 °C for 30 s, 56 °C for 30 s, 72 °C for 2 min; final extension of 10 min at 72 °C. | ~1800 | [23] |
| Present study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamada, P.d.O.F.; Müller, M.I.; Zago, A.C.; Yamada, F.H.; Ebert, M.B.; Franceschini, L.; da Silva, R.J. Three New Species of Jainus (Monogenea: Dactylogyridae) Parasitizing Gills of Brazilian Freshwater Fishes Supported by Morphological and Molecular Data. Diversity 2023, 15, 667. https://doi.org/10.3390/d15050667
Yamada PdOF, Müller MI, Zago AC, Yamada FH, Ebert MB, Franceschini L, da Silva RJ. Three New Species of Jainus (Monogenea: Dactylogyridae) Parasitizing Gills of Brazilian Freshwater Fishes Supported by Morphological and Molecular Data. Diversity. 2023; 15(5):667. https://doi.org/10.3390/d15050667
Chicago/Turabian StyleYamada, Priscilla de Oliveira Fadel, Maria Isabel Müller, Aline Cristina Zago, Fabio Hideki Yamada, Mariana Bertholdi Ebert, Lidiane Franceschini, and Reinaldo José da Silva. 2023. "Three New Species of Jainus (Monogenea: Dactylogyridae) Parasitizing Gills of Brazilian Freshwater Fishes Supported by Morphological and Molecular Data" Diversity 15, no. 5: 667. https://doi.org/10.3390/d15050667