
Peptides from Marine-Derived Fungi: Chemistry and Biological Activities †




1
H.E.J. Research Institute of Chemistry, International Center for Chemical and Biological Sciences, University of Karachi, Karachi 75270, Pakistan
2
Department of Advanced Medical and Surgical Sciences, University of Campania “Luigi Vanvitelli”, 80138 Naples, Italy
3
Laboratório de Química Orgânica e Farmacêutica, Departamento de Ciências Químicas, Faculdade de Farmácia, Universidade do Porto and CIIMAR, Rua de Jorge Viterbo Ferreira 228, 4050-313 Porto, Portugal
4
ICBAS—Instituto de Ciências Biomédicas Abel Salazar, Universidade do Porto and CIIMAR, Rua de Jorge Viterbo Ferreira 228, 4050-313 Porto, Portugal
*
Author to whom correspondence should be addressed.
†
Dedicated to the memory of Prof. Dr. José Augusto C. Pereira.
Mar. Drugs 2023, 21(10), 510; https://doi.org/10.3390/md21100510
Submission received: 3 August 2023
/
Revised: 16 September 2023
/
Accepted: 24 September 2023
/
Published: 26 September 2023
(This article belongs to the Section Structural Studies on Marine Natural Products)
Abstract
:Marine natural products are well-recognized as potential resources to fill the pipeline of drug leads to enter the pharmaceutical industry. In this circumstance, marine-derived fungi are one of the unique sources of bioactive secondary metabolites due to their capacity to produce diverse polyketides and peptides with unique structures and diverse biological activities. The present review covers the peptides from marine-derived fungi reported from the literature published from January 1991 to June 2023, and various scientific databases, including Elsevier, ACS publications, Taylor and Francis, Wiley Online Library, MDPI, Springer, Thieme, Bentham, ProQuest, and the Marine Pharmacology website, are used for a literature search. This review focuses on chemical characteristics, sources, and biological and pharmacological activities of 366 marine fungal peptides belonging to various classes, such as linear, cyclic, and depsipeptides. Among 30 marine-derived fungal genera, isolated from marine macro-organisms such as marine algae, sponges, coral, and mangrove plants, as well as deep sea sediments, species of Aspergillus were found to produce the highest number of peptides (174 peptides), followed by Penicillium (23 peptides), Acremonium (22 peptides), Eurotium (18 peptides), Trichoderma (18 peptides), Simplicillium (17 peptides), and Beauveria (12 peptides). The cytotoxic activity against a broad spectrum of human cancer cell lines was the predominant biological activity of the reported marine peptides (32%), whereas antibacterial, antifungal, antiviral, anti-inflammatory, and various enzyme inhibition activities ranged from 7% to 20%. In the first part of this review, the chemistry of marine peptides is discussed and followed by their biological activity.
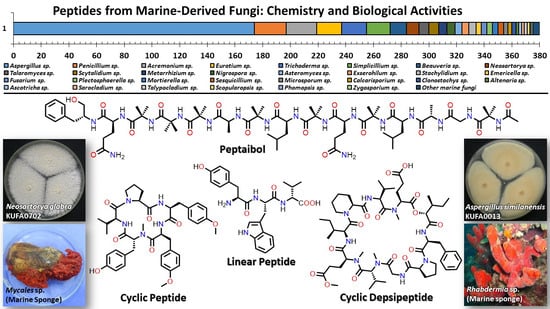
1. Introduction
Fungi are ubiquitous micro-organisms living symbiotically or endophytically with almost all viable resources. Marine fungal strains exist in various habitats and play a significant ecological role due to their capacity to produce a great variety of specialized metabolites such as alkaloids, polyketides, terpenoids, and peptides [1]. Peptides are a class of promising secondary metabolites with relevant pharmacological/biological activities due to their low toxicity and good affinity to a variety of important targets, such as G-protein-coupled receptors (GPCRs) or/and ion channels. These bioactive peptides consist of a series of well-ordered amino acids with approximate molecular weights of 500–5000 Da. Most significantly, bioactive marine-derived peptides exhibit a myriad of biological activities viz. antioxidant, antibacterial, antiviral, antifungal, antidiabetic, antiproliferative, anti-inflammatory, anticancer, anticoagulant, anti-hypertensive, antiobesity, anti-Alzheimer, and calcium-binding activities [2,3,4,5]. Due to their interesting bioactivities and specificity, peptides are considered to have a high potential for design, and even for modification, in order to obtain peptide-based drug candidates for clinical trial purposes. However, besides the expensive process of peptide synthesis, the main drawback of synthetic peptides is their poor physico-chemical property and enzymatic stability, resulting in less bioavailability and short circulating plasma half-life that limit their success in the body [6,7]. In terms of the production of marine peptides, there are three distinct categories including (i) naturally occurring peptides isolated directly from marine resources; (ii) naturally occurring peptides derived from the fermentation of marine-derived micro-organisms; and (iii) peptides derived from the hydrolysis of proteins obtained from marine organisms using various digestive enzymes, i.e., α-chymotrypsin, pepsin, papain, trypsin, and other proteases [8]. However, compared to naturally occurring human peptides, the structures of marine-derived peptides are different in either the backbone or side chains. Associated with unique conditions of the marine environment, marine-derived peptides are valuable candidates for drug design since they are mostly enzymatically and thermally stable [9]. The biosynthesis of marine-derived peptides and terrestrial peptides is a complex and varied process that can involve multiple enzyme systems, depending on the specific organisms and peptides being produced. For instance, some marine organisms produce peptides through nonribosomal peptide synthetases (NRPS) or nonribosomal peptide synthetase/polyketide synthase hybrids (NRPS/PKS), which are large, multi-domain enzymes that catalyze the assembly of amino acids into peptides. Similarly, some terrestrial organisms, such as fungi, produce peptides through NRPS. However, there are also examples of marine- and terrestrial-derived peptides that are produced by simpler biosynthetic pathways, such as ribosomal synthesis, which involve a direct translation of mRNA into a peptide chain by ribosomes, and, consequently, undergo post-translational modification [10]. Since ribosomally synthesized and post-translationally modified peptides (RiPPs) include only 20 unmodified proteinogenic amino acids, whereas nonribosomal peptides (NRPs) contain more than 100 non-coded non-proteinogenic amino acids, the chemical diversity of NRPs is higher than that of RiPPs. However, the RiPPs can better bind to the targets due to their restricted conformational flexibility originated during post-translational/co-translational modifications, thus making them appropriate for pharmaceutical applications [11]. Peptide-producing eukaryotic microbes generally reside in sponges, algae, dinoflagellates, and diatoms. Marine-derived fungi are among the micro-organisms of great interest and receive much attention because of their capacity to generate a great diversity of secondary metabolites with interesting biological and pharmacological properties [12,13].
To the best of our knowledge, there were several reviews covering only particular peptides from some specific fungal species [5,9,14,15,16]. Therefore, we hereby present a comprehensive review aiming to provide the information on structural diversity and biological/pharmacological activities of marine-derived peptides that have been reported from all the peptide-producing marine fungal strains from January 1991 to June 2023. Moreover, the stereochemistry of amino acids and non-amino acid moieties was briefly referred to in this review since this subject was thoroughly discussed in a recent review by Fernandes et al. [17]. The databases used to search for the keywords were SciFinder, Pubmed, Web of Science, Scopus, the Marine Pharmacology website, and Google Scholar.
2. The Importance of Marine-Derived Peptides in Drug Discovery
A large number of in vitro and in vivo investigations demonstrated the safety and efficacy of peptides. Naturally occurring and chemically modified marine peptides have demonstrated their potential in a myriad of therapeutic areas, acting as hormones, growth factors, neurotransmitters, ion channel ligands, antihypertensive, anti-inflammatory, and anti-infective agents, and continue to fuel the drug pipeline [4]. The best examples of the Food and Drug Administration (FDA)-approved marine-derived peptides, which are on the market, are ziconotide (Prilat®), brentuximab vedotin (Adcetris®), polatuzumab vedotin (Polivy®), enfortumab vedotin (PADCEVTM), disitamab vedotin (AidixiTM), tisotumab vedotin (TIVDAKTM), and plitidepsin (Aplidin®) (Figure 1) [18,19,20,21,22,23,24,25]. Ziconotide, a synthetic version of ω-conotoxin MVIIA, the cyclic peptide derived from the fish-eating marine cone snail Conus magus, is a N-type calcium channel blocker and is used as an analgesic for chronic pain [18]. Brentuximab vedotin, an antibody-drug conjugate medication used for the treatment of relapsed or refractory Hodgkin’s lymphoma (HL) and systemic anaplastic large cell lymphoma (ALCL), consists of monomethyl auristatin E (MMAE), which is a synthetic analogue of a marine-derived peptide, dolastatin 10, conjugated with a chimeric monoclonal antibody, cAC10, that targets CD30 protein which is expressed abundantly in cancer cells [19]. On the other hand, polatuzumab vedotin, which was approved by the FDA in 2019 for the treatment of non-Hodgkin’s lymphoma, chronic lymphovytic leukemia, and diffuse large B-cell lymphoma, consists of MMAE conjugated with a monoclonal antibody targeting the CD79b protein [20].
Enfortumab vedotin is an antibody and microtubule inhibitor conjugate designed to target nectin-4, a cell adhesion molecule found at high levels in urothelial cancer. This drug is directed at cancer cells, particularly in locally advanced or metastatic urothelial cancer cases where prior treatment involved a programmed death receptor-1 (PD-1) or programmed death ligand-1 (PD-L1) inhibitor and platinum-containing chemotherapy [21]. Disitamab vedotin is another antibody-drug conjugate that pairs a monoclonal antibody against human epidermal growth factor receptor 2 (HER2) with MMAE via a cleavable linker. This drug targets solid tumors like gastric cancer, HER2 overexpressing gastric carcinoma, urothelial carcinoma, and advanced breast cancer [22].
Tisotumab vedotin is an antibody-drug conjugate featuring a fully human monoclonal antibody (TF-011) directed at tissue factor, combined with MMAE to target tissue-factor-positive tumors. It is applied in the case of metastatic cervical cancer post-chemotherapy progression [23]. Plitidepsin (Aplidin®), a cyclic depsipeptide isolated from a Meditranean tunicate, Aplidium albicans, has passed the clinical trial studies as an anticancer drug for multiple melanoma, leukemia, and lymphoma [25].
Other marine peptides in the clinical trial phases include kahalalide F, elisidepsin (PM02734), tasidotin (ILX-651), glembatumumab vedotin (CDX-011), soblidotin (TZT-1027), E7974, HTI-286, XEN-2174, salinosporamide A, dalatazide (Shk-186), Vc1.1, RgIA, and RgIA4 (KCP-400) [9,26,27,28].
Kahalalide F (Figure 2), a synthetic version of a cyclic depsipeptide, isolated from a mollusk Elysia rufescens and its green algal diet Bryopsis sp., was being evaluated in a phase II clinical trial in malignant melanoma patients. Although kahalalide F was a well-tolerated and safe chemotherapy regimen, the trial was closed due to the lack of objective response in patients with malignant melanoma [29].
Elisidepsin (PM02734; Figure 2) is a synthetic analogue of kahalalide F, which has passed a phase I clinical study against malignant-solid tumors, and is in phase II clinical studies [30].
Tasidotin (ILX-651; Figure 2), an orally active synthetic microtubule-targeted derivative of dolastatin-15, is currently undergoing clinical evaluation for cancer treatment [31]. Glembatumumab vedotin (CDX-011) and soblidotin (TZT-1027), another dolastatin 10 analogues (Figure 2), are in phase I/II clinical trials for the treatment of breast cancer and soft tissue sarcoma, respectively [32,33].
E7974 and HTI-286 (Figure 2) are synthetic analogues of a cytotoxic tripeptide, hemiasterlin, which is derived from a marine sponge Hemiasterella minor. E7974 was in a phase I study for colorectal, prostate, and larynx carcinomas, while HTI-286 is preclinically applied in cases of metastatic prostate cancer [34,35].
XEN-2174 (Figure 2) is a synthetic peptide that binds specifically to the norepinephrine transporter, thus causing an inhibition of norepinephrine uptake. XEN-2174 has progressed to a phase IIb trial; however, it showed dose-limiting toxicity in pharmacodynamics and cerebrospinal fluid pharmacokinetics assays. Thus, it is unlikely that this conotoxin can be used for the treatment of acute pain in humans [36].
Shk-186 (Figure 2) is a 37-amino-acid synthetic peptide that is a specific inhibitor of the voltage-gated Kv1.3 potassium channel. It is a derivative of ShK, which was originally isolated from the venom of the sea anemone Stichodactyla helianthus. Shk-186 had a 100-fold improvement in selectivity for KV1.3 over the voltage-gated K+, KV1.1, KV1.4, and KV1.6, and was found to ameliorate autoimmune diseases such as multiple sclerosis and rheumatoid arthritis in human models. Interestingly, ShK-186 was found to have a long half-life at the site of injection, which produced sustained high pM levels in plasma, thus minimizing the need to improve its pharmacokinetic properties. An investigational new drug (IND) was filed by Kineta and approved by the FDA in 2012. ShK-186 has been allocated the generic name dalazatide, and completed phase 1a and 1b trials in 2016. The results of the phase Ib trial for psoriasis were reported recently, and showed that dalazatide was well-tolerated, without serious adverse events, and reduced psoriatic skin lesions. It is positioned to begin phase IIa trials [26].
Conotoxin Vc1.1 (Figure 2) is a modified synthetic form of the 16-amino-acid α-conotoxin Vc1a from the venom of Conus victoriae, which exhibited exceptional activity in animal models of neuropathic pain, causing long-lasting analgesia for at least 24 h following a single subcutaneous dose. Metabolic Pharmaceuticals started to develop Vc1.1 for the treatment of neuropathic pain and had progressed through double-blinded phase 1 clinical trials. However, a phase IIa clinical trial in patients with sciatic neuropathic pain was abandoned because Vc1.1 was significantly less potent on the human form of its presumed target, the α9α10 nicotinic acetylcholine receptor (nAChR), than had earlier been observed for the rat isoform. Therefore, further commercial development was halted [27].
RgIA (Figure 2) is a 13-residue two intramolecular disulfide peptide of the α-conotoxin class, originally isolated from a sea snail, Conus regius. Although a-conotoxin antagonists of α9α10 nAChRs have been proposed as potential analgesics for the treatment of neuropathic pain, they are less potent on human than rodent nAChRs, thus limiting their translational application. To overcome RgIA limitations, Kineta Inc., in collaboration with researchers from the University of Utah at Salt Lake City, has conducted SAR studies on RgIA which led to the development of RgIA4 (KCP-400; Figure 2), a peptide that exhibits high potency for both human and rodent α9α10 nAChRs, and was at least 1000-fold more selective for α9α10 nAChRs over all other molecular targets tested, including opioid and GABAB receptors [26]. Besides conducting safety and efficiency studies for KCP-400, Kineta Inc. also developed a non-opioid KCP-506 and has recently completed a single ascending dose (SAD) clinical study. KCP506 may potentially be an effective treatment for many types of chronic neuropathic pain including radiculopathy, chemotherapy-induced peripheral neuropathy, and diabetic neuropathy. KCP506 offers the potential for a disease-modifying therapy that may slow or halt the progression of chronic pain [28].
3. Peptides from Marine-Derived Fungi
A comprehensive literature survey of peptides covering the period of January 1991–June 2023 was undertaken. To facilitate the discussion of the reported peptides from marine-derived fungi, we classify them as linear peptides, cyclic peptides, cyclic depsipeptides, and, in some cases, more complicated structures [37].
3.1. Linear Peptides
Linear peptides are peptides whose amino acids are linked linearly in sequence. They have one free amino end and one free carboxyl end.
3.1.1. Linear Dipeptides
Liang et al. reported two acylated linear dipeptides, simplicilliumtides G (1) and H (2) (Figure 3), from the culture broth extract of a deep-sea-associated fungus, Simplicillium obclavatum EIODSF 020, collected in the Indian Ocean. Extensive 1D and 2D NMR spectral analysis revealed the presence of an acetyl group, Ile, and N-Me-Tyr residues in 1, while 2 contains the N-Me-Phe residue instead of the N-Me-Tyr residue. The absolute configurations of the Ile, N-Me-Tyr, and N-Me-Phe residues in 1 and 2 were determined by Marfey’s method as L-Ile, N-Me-D-Tyr, and N-Me-D-Phe, respectively [38].
Coniosulfide E (3) (Figure 3), a rare linear dipeptide containing an unusual cysteinol moiety, was isolated from the ethyl acetate (EtOAc) culture extract of a deep-sea shrimp-derived fungus, Aspergillus unguis IV17-109. In order to determine the absolute configuration at C-7, the authors synthesized 3 using farnesyl chloride with L- and D-Cys, and compared the optical rotations of the two obtained products with the isolated compound. Since the optical rotation of 3 ( −100; c 0.3, MeOH) was similar to that of the L-Cys-containing product ( −110; c 0.3, MeOH), the absolute configuration at C-7 in 3 was deduced as 7R. The E-configuration of Δ11 and Δ15 was assigned by NOESY correlations [39].
Chemical investigation of the culture extract of a marine-derived fungus, Penicillium sp. SCSIO 41512, afforded two undescribed linear dipeptides, penicamides A (4) and B (5) (Figure 3). Their structures were established by a combination of 1D and 2D NMR spectral and high-resolution electrospray ionization mass spectrometry (HRESI-MS) analyses. The absolute configurations of the stereogenic carbons were established by Marfey’s method and quantum chemical calculations [40].
3.1.2. Linear Tripeptides
The fermentation broth extract of S. obclavatum EIODSF 020 also furnished the linear acylated tripeptides, simplicilliumtides C-F (6–9) (Figure 4). HMBC correlations and Marfey’s analysis were used to identify their amino acid residues, amino acid sequences, and their absolute configurations, respectively, as Ac-L-allo-Ile-N-Me-D-Phe-L-Leu for 6; Ac-L-allo-Ile-N-Me-D-Tyr-L-Leu for 7; Ac-L-allo-Ile-N-Me-D-Tyr-L-Leu for 8 (replacing the COOH group of Leu with COOMe); and Ac-D/L-Val-N-Me-D-Phe-L-Leu for 9, which contains a mixture of D- and L-Val [38].
Another linear tripeptide, aspergillipeptide E (10) (Figure 4), was isolated from the culture broth extract of a marine-derived fungus, Aspergillus sp. SCSIO 41501, obtained from a gorgonian, Melitodes squamata, which was collected from the South China Sea. Then, 1D and 2D NMR data and Marfey’s method were used to establish the structure of 10 as D-Tyr-L-Trp-D-Val [41].
Two isomeric linear tripeptides, aspergillamides C (11) and D (12), together with the previously reported aspergillamides A (13) and B (14), cis-L-phenylalaninamide (15), and trans-L-phenylalaninamide (16) (Figure 4), were obtained from the mycelium extract of A. terreus SCSIO41008, isolated from a marine sponge, Callyspongia sp., which was collected from the seaside in Guangdong Province, China. The absolute configurations at C-12 and C-20 in 11 and 12 were established as 12S,20S, which are the same as those of C-12 and C-20 of 13–16, by a comparison of the calculated and experimental electronic circular dichroism (ECD) spectra [42].
A chemical investigation of the EtOAc extract of the mycelium of a marine-derived fungus, A. terreus LM.5.2, isolated from the leaves of a mangrove tree, Kandelia candel, which were collected from coast of Khanh Hoa province, South China Sea, Vietnam, resulted in the isolation of three unreported tripeptides containing a cinnamic acid (CA) moiety, asterripeptides A-C (17–19) (Figure 4). The structures of the amino acid residues of 17–19 were elucidated by 1D and 2D NMR spectral analysis and their sequences were determined by HMBC and ROESY correlations as Phe-Ile-Pro, Phe- Leu-Pro, and Phe-Val-Pro, respectively The absolute configurations of the amino acid residues were established by Marfey’s method and chiral HPLC as D-Phe-L-Ile-L-Pro, D-Phe-L-Leu-L-Pro, and D-Phe-L-Val-L-Pro [43].
Two unreported tripeptides with an unusual 5/6/5 heterocyclic scaffold bearing a N-trans-cinnamoyl moiety, talaropeptins A (20) and B (21) (Figure 4), were isolated from the EtOAc culture extract of a marine-derived fungus, Talaromyces purpureogenus CX11, isolated from the seaweed Grateloupia filicina C. Ag (Wulf.), which was collected from Zhoushan, Zhejiang province, China. The presence of a trans-cinnamoyl moiety (cin), Pro, Ala, and α-oxygenated Ser (α-O-Ser) in 20 was evidenced by 1D and 2D NMR spectral analyses. The HMBC and ROESY correlations showed the amino acid sequence as cin-Ser-Ala-Pro. The relative configurations of the stereogenic carbons in 20 were determined as 8R*,10R*,15S*,16S* by ROESY correlations while their absolute configurations were established as 8R,10R,15S,16S by Marfey’s analysis and comparison of the calculated and experimental ECD spectra [44].
On the other hand, the 1D and 2D NMR spectral analysis revealed that the structure of 21 is similar to 20 except for the presence of Ala instead of Ser. The absolute configurations of the stereogenic carbons in 21 were assumed to be the same as those of 20 due to their shared biosynthetic origin. This hypothesis was supported by a comparison of the calculated and experimental ECD spectra. The structures of 20 and 21 suggested that their biosynthesis is carried out predominantly by NRPS machinery [44].
Meyer et al. reported the isolation of penilumamide (22) (Figure 4), a lumazine peptide containing L-methionine sulfoxide and anthranilic acid (2-aminobenzoic acid, Abz) ester, from the fermentation broth extract of Penicillium sp. (strain CNL-338), isolated from a red alga, Laurencia sp., which was collected in the Bahamas Islands [45].
Penilumamides B–D (23–25) (Figure 4) were isolated, together with 22, from the mycelial and culture broth extracts of Aspergillus sp. SX-20090B15, isolated from the inner tissue of a fresh gorgonian, Muricella abnormaliz, which was collected from the Xisha Islands coral reef on the South China Sea. Interestingly, 22, 24, and 25 were isolated from the culture in a normal potato glucose liquid medium, whereas 23 was only isolated from the L-Met supplemented culture. The authors have found that 22 and 23 were unstable when exposed to air. Moreover, when 23 was exposed to air at room temperature, 22 was detected after a few days and 24 appeared several days later, suggesting that methionine sulfoxide in 22 and methionine sulfone in 24 are formed by the oxidation of Met in 23 [46].
The EtOAc extract of the mycelium of Aspergillus sp. (33241), isolated from the mangrove plant, Bruguiera sexangula var. rhynchopetala, collected from the South China Sea, also furnished another lumazine-containing peptide, aspergilumamide A (26) (Figure 4), in addition to 22. The stereochemistry of L-Glu in 26 was determined by Marfey’s analysis [47].
Further lumazine-containing peptides, terrelumamides A (27) and B (28) (Figure 4), were isolated from the culture broth extract of A. terreus FA009, which was obtained from a marine sediment collected off the shore of Jeju Island, Korea. Both 27 and 28 contain 1-methyllumazine-6-carboxylic acid and Abz methyl ester. The difference between 27 and 28 is the presence of Thr in the former and Ser in the latter. The configurations of both Thr and Ser were determined as L by the advanced Marfey’s method [48].
3.1.3. Linear Tetra- and Hexapeptides
Simplicilliumtides A (29) and B (30) (Figure 5), linear tetrapeptides containing Abz residue, were isolated from the combined mycelia and broth extracts of a deep-sea-derived fungus, S. obclavatum EIODSF 020, collected in the Indian Ocean. The amino acid sequences of both peptides were established by HMBC correlations, whereas Marfey’s analysis was used to determine the absolute configurations of the amino acid residues. Consequently, the structures of 29 and 30 were established as L-Val-N-Me-D-Phe-L-Leu-Abz, and L-allo-Ile-N-Me-D-Phe-L-Val-Abz, respectively [38].
Further chemical investigation of the same fungus led to the isolation of another linear tetrapeptide, simplicilliumtide I (31) (Figure 5); 1D and 2D NMR spectral analysis, in combination with low-resolution two-stage mass spectrometer equipped with electrospray ionization (LR-ESI-MS/MS), established the amino acid sequence as Ile-N-Me-Phe-Met sulfoxide [Met(O)], while Marfey’s method and HPLC established the configurations of the amino acids as L-allo-Ile, N-Me-D-Phe, and L-Met(O), respectively [49].
Ma et al. described the isolation of aspergillipeptides F (32) and G (33) (Figure 5) from the culture broth extract of a marine-derived fungus, Aspergillus sp. SCSIO 41501, isolated from a gorgonian, Melitodes squamata, which was collected from the South China Sea. HMBC correlations were used to establish the amino acid sequence of 32 as Tyr-Ala-Val-Val, and of 33 as Tyr-Val-Val-Ala. Since Marfey’s method revealed the presence of both D- and L-Val, it was not possible to identify which one is L and which one is D in both compounds [41].
Aspergillipeptides H–K (34–37) (Figure 5) were obtained from the extract of a solid rice culture of Aspergillus sp. SCSIO 41501, isolated from the gorgonian M. squamata Nutting, which was collected from the South China Sea. A detailed analysis of HMBC correlations established the sequence of the amino acid residues in 35–37 as N-Ac-Tyr-Trp-Val-Val, N-Ac-isopentenyl-Tyr-Ala-Val-Val, and N-acetyl-isopentenyl-Tyr-Phe-Val-Val, respectively. Since Marfey’s method was not able to completely assign the stereochemistry of the amino acids in 35–37, only D-Tyr, L-Val, and L-Val (for 35), L-Ala, L-Val, and L-Val (for 36), and L-Phe (for 37) were determined, while the configurations of the remaining amino acids were unassigned. Although the sequence of the amino acids in 34 was established as N-acetyl-isopentenyl-Tyr-Trp-Val-Val, their configurations were not assigned [50].
Chemical investigation of a semi-solid culture extract of A. ochraceopetaliformis, which was collected from an underwater sediment obtained off the coast of Jaeju-do Island, Korea, resulted in the isolation of FJ120DPB (38) (Figure 5). The structure of 38 was elucidated as N-Me-L-Phe-L-Ala-Ac-L-Thr-D-Val-L-Pro-L-Tyr by a combination of HMBC correlations and Mafrey’s analysis [51].
3.1.4. Linear Octapeptides
Two N-methylated linear octapeptides, RHM1 (39) and RHM2 (40) (Figure 6), were isolated from the culture extract of an atypical Acremonium sp. (UCSC coll. no. 021172 cKZ), isolated from a marine sponge, Teichaxinella sp. (coll. no. 02172), which was collected at Milne Bay, Papua New Guinea. The identification of amino acid residues and their sequences of both compounds were based on the assembly of various fragments whose structures were established by a combination of 2D NMR spectra and fragments ions from the electrospray ionization mass spectrum (ESIMS). The configurations of the amino acid residues in 39 were determined by Mafrey’s method. Thus, the structure of 39 was established as Ac-(R)-Gln-(2S,3S)-Ile-(S)-N-Me-Leu-(2S,3S)-Ile-(S)-N-Me-Val-(2S,3S)-N-Me-Ile-(2S,3S)-N-Me-Ile-(2S,3S)-N-Me-Ile-OH. On the other hand, the configuration of the amino acid residues in 40 were postulated to be the same as those in 40 on the basis of their biosynthetic origin which established the structure of 40 as Ac-(R)-Glu-(S)-Val-(S)-N-Me-Leu-(S)-N-Me-Val-(2S,3S)-Ile-(2S,3S)-N-Me-Ile-(2S,3S)-N-Me-Ile-(2S,3S)-N-Me-Ile-OH [52].
Further investigation of the same fungus by the same research group resulted in the isolation of another two N-methylated linear octapeptides, RHM3 (41) and RHM4 (42) (Figure 6). The absolute configurations of the amino acids in 41 and 42 were concluded to be the same as those of 39 and 40 on the basis of their biosynthetic origin [53].
3.1.5. Linear Nonapeptides
A chemical investigation of the fermentation extract of Trichoderma asperellum, obtained from a sediment of the Antarctic Penguin Island, furnished six peptaibols, asperelines A–F (43–48), containing nine amino acid residues (Figure 7). Peptaibols are characterized by featuring an abundance of α-aminoisobutyric acid (Aib) residue, the N-acyl terminus such as an acetyl group, and the C-terminus which contains an amino alcohol residue [54].
Compounds 43–46 and 48 contain various units of Aib, Val, Ile, Ala, and the amino alcohol protinol, while 47 contains Ser instead of Ala. The sequences of the amino acids in 43–48 were determined using 2D NMR, especially HMBC correlations together with the fragmentation ions from the ESIMS/MS analysis, while the configuration of each amino acid is determined by comparison of the 1H NMR spectra of the complex formed by each amino acid, obtained by hydrolysis of the peptaibol, with a chiral reagent, Ru(D4-Por*)CO, with the 1H NMR data of the complex between the L/D-standard amino acids. The configurations of Ala, Val, Ile, and Ser were established as L, while the configuration of the stereogenic carbon of prolinol was established as S [54].
3.1.6. Linear Undecapeptides
A 24-well microbioreactor cultivation analysis (MATRIX) using eleven different media and three phases (solid agar, as well as static and shaken broth), in combination with ultra-high-performance liquid chromatography with diode-array detection (UHPLC-DAD) and the ultra-high performance liquid chromatography-quadrupole time-of-flight tandem mass spectrometry (UHPLC-QTOF-MS/MS) analyses, resulted in the isolation of two N-methylated linear undecapeptides, talaropeptides A (49) and C (50) (Figure 8), from the culture extract of Talaromyces sp. CMB-TU011, isolated from an unidentified tunicate, which was collected in New South Wales, Australia. In the 1D NMR spectra of 49 and 50, the overlapping of resonances did not allow us to assign their amino acid residues. Although the partial amino acid sequences were determined by diagnostic HMBC and ROESY correlations, the linkage of amino acid fragments remained to be solved. A further diagnostic of the UHPLC-QTOF-MS/MS fragmentation patterns provided two consolidated partial sequences for 49 (i.e., N-Me-Ala1-N-Me-Val2-Val3-Thr4-N-Me-Val5 and N-Me-Val8-N-Me-Phe9-N-Me-Ile10-Leu11) and a consolidated partial sequence for 50 (i.e., N-Me-Val8-N-Me-Phe9-N-Me-Ile10-Leu11). Moreover, all the amino acid residues in 49 and 50 were determined as L-configured using C3 and C18 Marfey’s methods. The final assembly of the partial sequences of amino acids allowed us to reach the complete structures of 49 and 50 as N-Me-L-Ala1-N-Me-L-Val2-L-Val3-L-Thr4-N-Me-L-Val5-L-Pro6-N-Me-L-Val7-N-Me-L-Val8-N-Me-L-Phe9-N-Me-L-Ile10-L-Leu11 for 49, and N-Me-N-Ac-L-Ala1-L-Val2-L-Val3-L-Thr4-N-Me-L-Val5-L-Pro6-N-Me-L-Val7-N-Me-L-Val8-N-Me-L-Phe9-N-Me-L-Ile10-L-Leu11 for 50 [55].
The mycelial extract of a fungal strain K063, obtained from a red alga, Ceratodictyon spongiosum, which was collected off Seragaki Beach in Okinawa, furnished dictyonamide A (51), and its glycosylated analog, dictyonamide B (52) (Figure 8). The amino acid residues and their sequence in 51 and the presence of the Abz moiety were determined by a high-resolution fast atom bombardment mass spectrum (HRFAB-MS), and 1D and 2D NMR spectral analysis, as well as an amino acid analysis of its hydrolysate. Marfey’s method revealed that all the amino acid residues, except for N-Me-Thr, have an L-configuration. The configuration of N-Me-Thr was resolved as L by chiral HPLC analysis. Thus, the structure of 51 was determined as L-Ala-L-Thr- N-Me-L-Thr-N-Me-L-Val-L-Val-N-Me-L-Val-N-Me-L-Val-N-Me-L-Val-L-Ile-N-Me-L-Val-N-Me-Gly-Abz. The sequence of the amino acid residues and their configurations of 52 were identified by the same methods as those of 51, as well as by a comparison of the NMR spectra and HPLC retention time of its aglycone with those of 51. The presence of D-fructose was confirmed by gas chromatography (GC) analysis using the chiral column (Chirasil-Val) of the tetramethyl silyl (TMS) derivative of the methanolysis product of 52 [56].
A chemical investigation of the EtOAc extract of the culture of an endophytic fungus, Tolypocladium sp., isolated from a marine microalga, Spongomorpha arcta, collected from the shores at Green’s Point, L’Etete, NB, Canada, furnished two undescribed lipopeptaibols, tolypocaibols A (53) and B (54) (Figure 8). Extensive 1D and 2D NMR analysis, including COSY and HMBC correlations, revealed the identity of the amino acid residues, the Aib and decanoyl (Dc) moiety, while the existence of a valinol residue was confirmed by 1H–1H TOCSY and COSY correlations. HMBC and ROESY correlations established the sequence of the amino acids in 53 as Dc-Aib11-Pro10-Phe9-Aib8-Gln7-Gln6-Aib5-Aib4-Gln3-Leu2-Valol1, which was supported by the MS/MS fragmentation data. By using the same methodology, it was found that the structure of 54 is very similar to that of 53 except that one Pro (Pro10) in 53 was replaced by a 4-methyl-proline (4-Me-Pro10) residue in 54. Thus, the amino acid sequence in 54 was established as Dc-Aib11–4-Me-Pro10-Phe9-Aib8-Gln7-Gln6-Aib5-Aib4-Gln3-Leu2-Valol1 [57].
The absolute configurations of the amino acid and amino alcohol residues in 53 and 54 were determined by hydrolysis and the application of Marfey’s method which led to the determination of an L-configuration for all amino acids, and the stereochemistry of the 4-Me-Pro10 in 54 was 2S,4S. Therefore, the complete structures of 53 and 54 were determined as Dc-L-Aib11-L-Pro10-L-Phe9-L-Aib8-L-Gln7-L-Gln6-L-Aib5-L-Aib4-L-Gln3-L-Leu2-L-Valol1 and Dc-L-Aib11-(2S,4S)-4-Me-L-Pro10-L-Phe9-L-Aib8-L-Gln7-L-Gln6-L-Aib5-L-Aib4-L-Gln3-L-Leu2-L-Valol1, respectively [57].
3.1.7. Linear Dodecapeptides
Two N-methylated linear dodecapeptides, talaropeptides B (55) and D (56) (Figure 9), were also isolated, together with 49 and 50 (Figure 8), from the culture extract of Talaromyces sp. (CMB-TU011). The 1D and 2D NMR data revealed the presence of twelve amino acid residues in both 55 and 56, viz. Thr, Pro, Val (x2), Leu, N-Me-Ala, N-Me-Val (x4), N-Me-Phe, and N-Me-Ile. Similar to 49 and 50, the absolute configuration of all amino acids in 55 and 56 were determined as L by Marfey’s method. Diagnostic HMBC and ROESY correlations of 55 and 56 allowed us to assign the partial amino acid sequences; however, the connection of amino acids was established by diagnostic UHPLC-QTOF-MS/MS fragmentation patterns. A combination of the assembly of the fragments and HMBC correlations allowed us to elucidate the complete structures of 55 and 56 as N-Me-L-Ala1-N-Me-L-Val2-L-Val3-L-Thr4-L-Val*-N-Me-L-Val5-L-Pro6-N-Me-L-Val7-N-Me-L-Val8-N-Me-L-Phe9-N-Me-L-Ile10-L-Leu11, and N-Me-N-Ac-L-Ala1-N-Me-L-Val2-L-Val3-L-Thr4-L-Val*-N-Me-L-Val5-L-Pro6-N-Me-L-Val7-N-Me-L-Val8-N-Me-L-Phe9-N-Me-L-Ile10-L-Leu11, respectively [55].
3.1.8. Linear Pentadecapeptides
A linear pentadecapeptide, efrapeptin G (57) (Figure 10), was also reported, together with the undescribed linear pentadecapeptides, efrapeptins Eα (58) and H (59) (Figure 10), and the previously reported efrapeptins E (60) and F (61) (Figure 10), from the culture extract of an atypical Acremonium sp. (UCSC coll. no. 021172 cKZ) [52]. The structures of 58 and 59 were established by extensive MSn fragmentations using a linear quadrupole ion trap electrospray ionization mass spectrometry (LTQ ESI-MS) technique while the absolute configurations of the amino acid residues in 58 and 59 were found to be the same as those of the previously discussed peptides of the RHM (39–42) and efrapeptin (57, 60 and 61) families [53].
Van Bohemen et al., in their search for new peptaibols, described the isolation of five unreported 15-residue peptaibols, pentadecaibins I–V (62–66) (Figure 10) from the solid culture extract of a marine sediment-derived fungus, Trichoderma sp. MMS1255, belonging to the T. harzianum clade. The sequences of their amino acids were determined by the MS/MS fragmentation and extensive 1D and 2D NMR analysis as Ac-Aib1-Gly2-Ala3-Leu4-Aib5-Gln6-Aib7-Val8-Aib9-Ala10-Aib11-Aib12-Aib13-Gln14-Pheol15 for 62, Ac-Aib1-Gly2-Ala3-Leu4-Aib5-Gln6-Aib7-Leu8-Aib9-Ala10-Aib11-Aib12-Aib13-Gln14-Pheol15 for 63, Ac-Aib1-Gly2-Ala3-Leu4-Aib5-Gln6-Iva7-Val8-Aib9-Ala10-Aib11-Aib12-Aib13-Gln14-Pheol15 for 64, Ac-Aib1-Gly2-Ala3-Leu4-Aib5-Gln6-Iva7-Leu8-Aib9-Ala10-Aib11-Aib12-Aib13-Gln14-Pheol15 for 65, and Ac-Aib1-Gly2-Ala3-Leu4-Iva5-Gln6-Iva7-Val8-Aib9-Ala10-Aib11-Aib12-Aib13-Gln14-Pheol15 for 66 [58].
3.1.9. Linear Octadecapeptides
The EtOAc extract of the solid rice culture broth of a marine-derived fungus, Trichoderma sp. GXIMD 01001, isolated from a marine sponge, Haliclona sp., which was collected in Beibu Gulf, Guangxi, China, furnished seven unreported 18-residue peptaibols, trichorzins A–G (67–73) (Figure 11) [59].
A detailed 1D and 2D NMR spectral analysis, together with high-resolution electrospray ionization mass spectrum (HRESIMS) data, confirmed the presence of 17 amino acid residues, including two Ala, six Aib, two Gln, one Gly, one Pro, two Leu, two Iva, and one Val, as well as an N-acetyl terminus and a Trpol (tryptophanol) at the C-terminus in 67, while 68–73 contain the monosubstituted phenyl ring (Pheol) instead of indolyl (Trpol) at the C-terminus. The amino acid sequence was established by an examination of NOESY correlations from the NH proton at the acetyl terminus to the next NH and/or α-H of the adjacent amino acid residue, which ultimately gave a complete sequence of 67–73 as Ac-Aib1-Ala/Ser2-Ala3-Aib/Iva4-Iva5-Gln6-Aib/Iva7-Val/allo-Ile8-Aib9-Gly10-Leu11-Aib12-Pro13-Leu14-Aib15-Aib16-Gln17-Trpol/Pheol18 [59].
The absolute configurations of the stereogenic carbons of 68–73 were determined by Marfey’s method which indicated the presence of an L-configuration for Ala, Ser, Val, Leu, Pro, and Glu (resulting from the hydrolysis of Gln), as well as the characteristic nonproteinogenic peptaibol residues D-Iva, L-Trpol, and L-Pheol. However, the L-form of allo-Ile was observed for the Ile residue in 71. Moreover, the ECD spectra of 67–73 showed a positive maximum near 200 nm and two negative maxima at 207 and 225 nm, suggesting, therefore, a right-handed helical conformation [59].
3.1.10. Linear Lipopeptides
The antiviral-activity-guided isolation of the saline fermentation extract of a marine- derived fungus, Scytalidium sp., led to the isolation of five undescribed lipophilic linear hexapeptides, halovirs A–E (74–78) (Figure 12), which contain an unusual Aib-hydroxyproline (OH-Pro) dipeptide fragment, while a carboxyl terminus is reduced to a primary alcohol. In order to determine the absolute stereochemistry, the acidic hydrolysis products of 74–78 were analyzed by GC with a flame ionization detector (FID) using a chiral column (Chirasil-Val), followed by Mosher’s method. L-Leu, L-Val, and L-Glu were identified in 74 and L-Ala, L-Leu, and L-Glu were identified in 75, while L-Pro was found in 76. In addition, the absolute configuration at C-3 of the 3-hydroxyproline (3-OH-Pro) residue in 74 was determined as 3R by Mosher’s method. Moreover, detailed 1D and 2D NMR spectral analyses, in combination with matrix-assisted laser desorption/ionization Fourier-transform-MS (MALDI-FTMS) data, confirmed that 77 and 78 derived from 74 and 76, respectively [60].
3.1.11. Other Linear Peptides
Flavipesides A–C (79–81), unusual linear methylated dipeptides containing chlorinated xanthone and modified pyrazol residues (Figure 13), were isolated from the culture extract of A. flavipes 164013, obtained from a marine sponge Dysidea sp., which was collected off the Yongxing Island in the South China Sea. Although the structures of the subunits in 79–81 were clarified by 1D and 2D NMR correlations, the linkages between the substructures remained unsolved. The absolute configurations of two amino acids in 79 were determined as L, i.e., 23S and 32S by single-crystal X-ray analysis using Cu Kα radiation. For 80 and 81, the absolute configurations of the two amino acids were found to be the same as those of 79 since their specific rotations ( −14.7 for 80, and −15.6 for 81) were similar to that of 79 ( −11.5). Moreover, the cotton effects in the experimental ECD spectra of 80 and 81 were well-matched with that of 79. Compounds 79–81, possessing a N,N,N-trimethyl group to form an ammonium salt of Phe, are not common for natural products [61].
Two epimeric lipopeptidyl benzophenones, asperphenins A (82) and B (83) (Figure 13), were isolated from a culture broth extract of a marine-derived fungus, Aspergillus sp., collected from a marine-submerged decaying wood. Detailed NMR analysis revealed that the structures of 82 and 83 comprise three different motifs, including a hydroxy fatty acid, a tripeptide, and a trihydroxybenzophenone. Interestingly, the absolute configuration of the hydroxyl-bearing carbon, C-33, was determined as R by Mosher’s NMR method while the absolute configurations of the stereogenic centers, C-23 and C-28, in 82 and 83 were determined as L by Marfey’s analysis [62].
The culture extract of a marine fungus, Penicillium fellutanum Biourge, isolated from the gastrointestine of a marine fish, Apogon endekataenta Bleeker, furnished two undescribed linear cytotoxic lipopeptides, fellutamides A (84) and B (85) (Figure 13), which contain 3-hydroxydodecanoic acid (HDA), β-threo-hydroxy gluthamine (βHGln), and 2-amino-4-methylpentanal (leucinal). Detailed HMBC correlations, FAB-MS/MS, and chiral GC and HPLC analyses determined the sequences of the amino acids and their absolute stereochemistry. Therefore, 84 was elucidated as HDA-L-Asn-L-threo-βHGln-Leucinal, while 85 was identified as HDA-L-Asn-L-Gln-Leucinal. The absolute configuration at C-3 in HDA was determined as R by the sign of its optical rotation −17) [63].
3.2. Cyclic Peptides
Cyclic peptides containing 2 to 10 amino acid residues are biosynthesized by NRPS.
3.2.1. Cyclic Dipeptides
Scopel et al., in their search for novel compounds against anti-biofilm formation, reported the isolation of a cyclic dipeptide, cis-cyclo-(Leu-Tyr) (86) (Figure 14), from Penicillium sp. F37, which was obtained from a marine sponge, Axinella corrugata [64].
Cyclo-(trans-4-hydroxy-L-Pro-L-Leu) (87), cyclo-(trans-4-hydroxy-L-Pro-L-Phe) (88), cyclo-(L-Pro-L-Leu) (89 or 3-isobutylpyrrolopiperazine-1,4-dione), cyclo-(L-Pro-L-Val) (90), cyclo-(L-Pro-L-Phe) (91), and cyclo-(L-Pro-L-Tyr) (92) (Figure 14) were isolated from the culture extract of a marine sediment-derived fungus, A. niger BRF-074, collected from the Northeast Brazilian coast [65]. Compounds 89 and 92 were also isolated from the fermentation extracts of two Taiwanese sediment-derived fungi, Fusarium sp. RWS56-10 [66] and P. chrysogenum DXY-1, respectively [67].
Penicimutide (93), which contains an unusual amino acid, 4,5-didehydro-L-leu, and cyclo-(L-Ile-L-Pro) (94) (Figure 14) were isolated, together with 89–91 (Figure 14), from a neomycin-resistant mutant of a marine sediment-derived fungus, P. purpurogenum G59, collected at the tideland of Bohai Bay around Lujuhe in the Tanggu District of Tianjin, China [68].
A chemical investigation of a marine-derived fungus, Ascotricha sp. ZJ-M-5, furnished cyclo-(Pro-Ala) (95), cyclo-(Ile-Leu) (96), and cyclo-(Gly-Pro) (97) (Figure 14), in addition to 89 and 90 [69].
The EtOAc extract of a culture of Neosartorya glabra KUFA 0702, isolated from a marine sponge Mycale sp., collected at a depth of 15–20 m from the coral reef at Samaesarn Island in the Gulf of Thailand, furnished an unreported fellutanine A epoxide (98), in addition to aszonalenin (99), (3R)-3-(1H-indol-3-ylmethyl)-3,4-dihydro-1H-1,4-benzodiazepine-2,5-dione (100), takakiamide (101), (11aR)-2,3-dihydro-1H-pyrrolo[2,1-c][1,4] benzodiazepine-5,11(10H,11aH)-dione (102), and fellutanine A (103) (Figure 14). In the case of 98, the absolute configurations of the stereogenic carbons of the diketopiperazine ring (C-11 and C-11′) were proposed to be the same as those of the co-isolated 103, i.e., 11S,11′S due to the same biosynthetic precursor (L-Trp). The NOESY experiments and molecular dynamic simulations not only confirmed the 11S,11′S configurations of the diketopiperazine but also suggested the stereochemistry of the epoxide ring as 2′S,3′S [70].
Capon et al. isolated acetylaszonalenin (104) (Figure 14), a known terrestrial fungal metabolite, from the mycelial extract of an estuarine sediment-derived fungus, A. carneus (MST-MF156), collected in Tasmania, Australia [71]. May Zin et al. also reported the isolation of 97, 101, and 104 (Figure 14) from the EtOAc extract of a solid rice culture of an algicolous fungus, N. takakii KUFC 7898, collected from Samaesarn Island in the Gulf of Thailand [72].
The mycelium extract of a marine sponge-associated fungus, A. terreus SCSIO41008, isolated from the marine sponge, Callyspongia sp., which was collected from the seaside in Guangdong Province, China, yielded terretriones B (105) and C (106), and brevianamide F (107), together with 91 (Figure 14) [42].
A chemical investigation of the solid culture extract of Eurotium chevalieri MUT2316, isolated from the Atlantic sponge, Grantia compressa, led to the isolation of cyclo-(L-Trp-L-Ala) (108), together with two prenylated cyclic dipeptides, neoechinulin A (109) and echinulin (110) (Figure 14) [73]. In another study, 109 was also reported from a marine-derived fungus, Microsporum sp. strain MFS-YL, which was isolated from the surface of a marine red alga, Lomentaria catenata, collected at Guryongpo, Nam-Gu, PoHang in the Republic of Korea [74]. Smetanina et al. also reported the isolation of 110 from the mycelium extract of a marine-derived E. repens, isolated from a marine sponge, Suberites domuncula, collected near Zelenyi Island [75].
Zhang et al. reported the isolation of three cyclodipeptides, cyclo-(L-Trp-L-Ile) (111), cyclo-(L-Trp-L-Phe) (112), and cyclo-(L-Trp-L-Tyr) (113) (Figure 14), from the culture broth extract of A. niger EN-13, which was obtained from the inner tissue of a marine brown alga, Colpomenia sinuosa, collected from the Qingdao coastline, China [76].
Buttachon et al. reported the isolation of (3S,6S)-3,6-dibenzylpiperazine-2,5-dione (114) (Figure 14) from the culture extract of A. candidus KUFA0062, obtained from a marine sponge Epipolasis sp., which was collected from the coral reef at the Similan Island National Park, Phang-Nga province, Thailand [77].
A chemical investigation of the EtOAc extract of the fermentation of A. ochraceopetaliformis DSW-2, which was obtained from the sea water sample from Dongshan Island in Fujian Province of China, led to the isolation of mactanamide (115), in addition to 99 (Figure 14) [78].
A deep-sea sediment-derived fungus, Aspergillus sp. SCSIOW2, collected in the South China Sea, furnished an undescribed cyclic dipeptide, 14-hydroxy-cyclopeptine (116) (Figure 14). The 1H NMR spectrum of 116 in DMSO-d6, at room temperature, displayed two sets of proton signals. However, the two sets of signals began to merge when the temperature was raised to 50 °C and coalesced into single resonances at 85 °C, indicating the presence of two different stable conformational isomers of 116. This hypothesis was further confirmed by ROESY correlations. An extensive 1D and 2D NMR analysis revealed the presence of N-Me-Tyr and Abz. The L-configuration of N-Me-Tyr was established by Marfey’s method [79].
The oxepin-containing pyrazinopyrimidin-7-one derivatives, protuboxepins A (117) and B (118), and the pyrroloindolodiketopiperazine derivatives, protubonines A (119) and B (120) (Figure 14), were obtained from the EtOAc extract of a culture of a marine-derived fungus, Aspergillus sp. SF-50044, isolated from the intertidal sediment sample collected from Dadaepo Beach, Busan, Korea. Then, 1D and 2D NMR analysis revealed that 117 contained Leu and Phe residues, while 118 consisted of Val and Phe. Marfey’s method was used to identify the configuration of D-Phe in 117. Since the NOESY spectrum showed correlations from H-4 to H2-16 and H-18/H-22, these protons are cofacial. Therefore, the absolute configuration at C-1 and C-4 were determined as R and S, respectively, while the configuration at C-23 remained unassigned. For 119 and 120, an analysis of the COSY and HMBC correlations confirmed the presence of Leu and the diketopiperazine ring. Through a combination of NOESY correlations with the result of Marfey’s analysis, L-Leu was identified. Thus, the absolute configurations of the stereogenic carbons in 119 and 120 were determined as 3S,5aR,10bR,11aR [80].
Oxepinamide E (121) (Figure 14), an oxepin-containing pyrimidine alkaloid, was isolated together with 117, from the mycelial extract of a co-fermentation of the strains BM-05 and BM-05ML of Aspergillus sp., isolated from a marine brown alga Sargassum sp., which was collected off Helgoland, North Sea, Germany [81].
The EtOAc culture extract of a marine sediment-derived fungus, Aspergillus sp. (CMB-M081F), collected from the intertidal depth of 1 m near Shorncliffe, Queensland, Australia, yielded an undescribed shornephine A (122) and its methanolysis artifact of a solvolytically unstable product, seco-shornephine A methyl ester (122a), together with the previously described 15b-β-hydroxy-5-N-acetyladreemin (123), 5-N-acetyladreemin (124), and 15b-β-methoxy-5-N-acetyladreemin (125) (Figure 14). Detailed 2D NMR analysis, including COSY and HMBC correlations, established the planar structures of 122 and 122a. Further diagnostic of the ROESY correlations revealed that H-2, H-2′, H-3α, 4-OH, H3-16, and H3-17 are on the α face, thus establishing the relative configuration of 122 [82].
The EtOAc broth extract of Asteromyces cruciatus 763, obtained from an unidentified decaying green alga which was collected at La Jolla shore, San Diego, USA, yielded hyalodendrin (126), gliovictin (127), 1N-norgliovictin (128), and bis-N-norgliovictin (129) (Figure 14), and four previously described (3R,6R)-epipolythiopiperazinedione antibiotics [83].
The culture extract of a marine sediment-derived fungus, A. versicolor MF180151, which was collected from the Bohai Sea, China, furnished diketopiperazine derivatives, including (±)-7,8-epoxy-brevianamide Q ((±)-130), (±)-8-hydroxybrevianamide R ((±)-131), (±)-8-epihydroxybrevianamide R ((±)-132), and (±)-brevianamide R ((±)-133) (Figure 14). The planar structures of 130–132 were established by 1D and 2D NMR spectral analysis. ROESY correlations between 2-NH and H-13 confirmed the presence of a cis double bond between C-3 and C-10 in 130–132. Further ROESY correlations from OH-9 to H-7 and H-8 in 130, and from OCH3-9 to H-8 in 131, revealed their relative configurations. A comparison of the chemical shift values of C-9 in 131 and in 132 suggested that 132 was an epimer of 131. Circular dichroism (CD) experiments of 130–132 did not exhibit a significant absorption. However, the values of optical rotations, +3.0 (c 0.1, CH3OH) for (±)-130, +1.0 (c 0.1, CH3OH) for (±)-131, and +2.0 (c 0.1, CH3OH) for (±)-132, were inherent to the racemic nature of 130–132 [84].
The cytotoxic-activity-guided fractionation of the culture broth extract of Exserohilum rostratum (Drechsler) CNK-630, obtained from a marine cyanobacterial mat which was collected off the northwest corner of Lanai Island, Hawaii, furnished four unreported C2-symmetrical diketopiperazine disulfides, rostratins A–D (134–137) (Figure 14), together with exserohilone (138) (Figure 14). The symmetrical nature of 134–137 were determined by analyses of their 1D and 2D NMR spectra in combination with their molecular formulae. A comparison of the 1D and 2D NMR data of 134–137 with those of 138 revealed that 134–137 are formed by the cyclic dimerization of highly oxidized Phe residues. The absolute configurations of the stereogenic carbons in 134–137 were determined by a combination of NOESY correlations and regioselective acylation by Mosher’s reagents in the NMR tube at reduced temperatures. As a result, the absolute structures of 134–137 were elucidated as 2(2′)R,4(4′)S,5(5′)S,8(8′)S,9(9′)S-134, 2(2′)R,4(4′)S,8(8′)S,9(9′)R-135, 2(2′)R,4(4′)S,7(7′)S,8(8′)R,9(9′)R-136, and 2(2′)R,4(4′)S,7(7′)R,8(8′)R,9(9′)R-137 [85].
Following the bioassay-guided isolation approach, gliocladine C (139) (Figure 14) was isolated, together with 91, 97, and 108, from a fermentation broth extract of a marine sediment-derived fungus, Penicillium sp. WF-06 [86].
Wu et al., in their OSMAC (One Strain MAny Compounds) program, reported the isolation of a structurally unique hexacyclic dipeptide, azonazine (140) (Figure 14), from a Hawaiian marine sediment-associated fungus, A. insulicola. The structure of 140 was assembled by an analysis of the HRESI-MS, gCOSY, gHMBC, and NOE correlations. The absolute configurations of the stereogenic carbons in 140 were determined as 2R,10R,11S,19R by comparison of its calculated and experimental ECD spectra [87].
(11R,14S)-3-(1H-indol-3-ylmethyl)-6-isopropyl-2,5-piperazinedione (141), preechinulin (142), neoechinulin E (143), eurocristatine (144) (Figure 15), and echinulin (110) (Figure 14) were isolated from a mycelial extract of a mangrove endophytic fungus, E. chevalieri KUFA 0006, which was obtained from the healthy twig of a mangrove tree, Rhizophora mucronata Poir., collected at the Eastern part of Thailand [88].
A chemical investigation of the EtOAc soluble fraction of the crude extract of a marine sediment-derived fungus, A. versicolor (MF030), isolated from the Bohai Sea, China, furnished an unreported dimeric diketopiperazine, brevianamide S (145), three undescribed diketopiperazines, brevianamides T (146), U (147), and V (148), together with the previously reported brevianamides N (149) and K (150), and deoxybrevianamide E (151) (Figure 15). An analysis of the (−)-HRESI-MS, 1H and 13C NMR data of 145 revealed its dimeric and symmetrical nature. An analysis of COSY, HMBC, and ROESY correlations established the planar structure of 145. The 1H and 13C NMR spectra of 146–148 resembled those of 145; however, their (−)-HRESI-MS data revealed that they were monomers. The 1D and 2D NMR data showed that the structure of 146 is similar to the monomer of 145 but with a double bond between C-6 and C-7, while 148 contains Pro residue, i.e., with no double bond between C-8 and C-9. Marfey’s analysis revealed the presence of L-Pro in 148. On the other hand, 147 has two hydroxyl groups at C-8 and C-9 instead of a double bond; however, the absolute configurations at C-8 and C-9 in 147 were not determined [89].
Further diketopiperazines derived from a condensation of Pro and Trp, i.e., pairs of enantiomers of brevianamides Z ((±)-152) and Z1 ((±)-153), together with their analogues viz. brevianamides X ((±)-154), R ((±)-133), Q ((±)-155), K (150), V ((+)-148), and deoxy brevianamide E ((±)-151) (Figure 15), were isolated from the EtOAc extract of a culture of A. versicolor HBU-7, which was obtained from a sea mud sample collected from the coast of Bohai, China. The planar structures of (±)-152 and (±)-153 were elucidated by 1D and 2D NMR spectral analysis, especially COSY and HMBC correlations. The absolute configurations of the stereogenic centers in (+)-152 and (−)-152 were determined as 7S and 7R, respectively, by a comparison of their calculated and experimental ECD spectra. On the other hand, the absolute configurations of the stereogenic centers in (+)-153, (−)-153, (+)-154, and (−)-154 were determined using DP4+ data analysis of the unscaled shift and shielding tensor data. The absolute configurations of (+)-153 and (−)-153 were established as 8S,9R and 8R,9S, respectively, while the absolute configurations of (+)-154, and (−)-154 were determined as 8R,9R and 8S,9S, respectively [90].
An undescribed dioxopiperazine, 12-demethyl-12-oxo-eurotechinulin B (156), was isolated, together with previously reported variecolorin J (157), eurotechinulin B (158), variecolorin G (159), alkaloid E-7 (160), cryptoechinuline G (161), isoechinulin B (162), and 7-isopentenylcryptoechinuline D (163) (Figure 15), from a solid-rice culture extract of a mangrove endophytic fungus, E. rubrum G2, which was obtained from the inner tissue of the semi-mangrove plant, Hibiscus tiliaceus Linn., collected from Hainan Island, China [91].
By using an MS/MS-based molecular networking approach, Mao et al. isolated three unreported N-ethynyl diketopiperazine derivatives, sclerotioloid A (164), its seco-analog, sclerotioloids B (165), and sclerotioloids C (166), together with two previously described ones, gartryprostatin C (167) and speramide C (168) (Figure 15), from the EtOAc fermentation extract of A. sclerotiorum ST0501, isolated from the inner tissue of an unidentified marine sponge which was collected from the South China Sea, Guangdong, China. The structures of 164–166 were elucidated by the interpretation of 1D and 2D spectra. In the case of 165, its structure and the configuration of the double bond were confirmed by X-ray analysis. The absolute configuration of the sterogenic carbon, C-6, in 166 was established as 6S by comparison of its calculated and experimental ECD spectra [92].
The cytotoxic fraction of the EtOAc fermentation broth extract of Aspergillus sp. DY001, isolated from the internal tissue of a tunicate, Didemnum sp., collected off Jizan, at the Saudi Red Sea coast, furnished two undescribed cyclic dipeptides, asperopiperazines A (169) and B (170) (Figure 15). Detailed 1D and 2D NMR spectral analysis revealed the presence of disubstituted diketopiperazine which is derived from N-Me-Leu and N-Ac-Phe in 169. The absolute configurations of Leu and Phe were established as L by Marfey’s analysis. Therefore, the complete structure of 169 was elucidated as cyclo-(N-Me-L-Leu-N-Ac-L-Phe. On the other hand, the 1D and 2D NMR spectral analysis showed that 170 is also a trisubstituted diketopiperazine. An analysis of the structure of 170 revealed that it consists of 2-hydroxy Pro (2-OH-Pro) and Phe. The absolute configuration of Phe was determined as D-Phe by Marfey’s method while the absolute configuration of C-6 (of the Pro residue) was established as D-Pro by a comparison of the 13C NMR chemical shift of C-6 with the corresponding carbon of the previously described cyclo-(6-OH-D-Pro-L-Phe) and cyclo-(6-OH-L-Pro-L-Phe). Therefore, the structure of 170 was established as cyclo-(6-OH-D-Pro-L-Phe) [93].
A hemical investigation of the methanol extract of a slid rice culture of Aspergillus sp. FS445, isolated from a deep-sea sediment which was collected from the Indian Ocean, furnished four unreported cyclic dipeptides, aspechinulins A–D (171–174), together with eight known cyclic dipeptides, including neoechinulin B (175), isoechinulin A (176), tardioxopiperazine A (177) (Figure 16), 109, 110 (Figure 14), and 160–162 (Figure 15). Detailed analysis of 1D and 2D NMR spectra of 171–174 showed that they are prenylated indole diketopiperazine derivatives comprising of Trp and Ala. The cis configuration of the double bond between C-8 and C-9 in 171–173 was established based on the NOESY correlation between H-4 and NH-14. The absolute configuration of C-12 in 173 was established as 12S, based on the negative value of its optical rotation and a similarity of its ECD spectrum and the ECD spectrum of rubrumazines A/B (negative Cotton effect at 205 nm and positive Cotton effects at 230 nm) [94]. This was confirmed by a comparison of the calculated and experimental ECD spectra of 173 [95].
For 174, which showed a negative value of rotation similar to that of the previously described rubrumazine C, its H-9 and H-12 were deduced to be co-facial. Moreover, since the experimental ECD spectrum of 174 displayed a negative Cotton effect at 230 nm, the configuration at C-9 was suggested to be 9S, which was the same as that of rubrumazine C [93]. Thus, the absolute configurations of C-9 and C-12 in 174 were assigned to be 9S,12S. On the other hand, the absolute configuration of C-27 could not be assigned either by Mosher’s method or 13C calculations. Moreover, the calculated ECD spectrum exhibited mirrored Cotton effects which made the authors speculated that a hydrogen bond between the hydroxyl group at C-27 and NH-1, which formed a seven-membered ring, could influence the Cotton effect in the ECD spectrum. By a comparison of the calculated and experimental ECD spectra of the analogs of 174, prepared by replacing the hydroxyl group at C-27 by the methyl group, the authors were able to establish the absolute configuration of C-27 as 27R [95].
The EtOAc extract of a solid rice culture of A. chevalieri CS-122, isolated from a deep-sea cold seep sediment collected in the northeast of the South China Sea, furnished five unreported cyclic dipeptides, 24,25-dihydroxyvariecolorin G (178), 25-hydroxyrubrumazine B (179), 22-chloro-25-hydroxyrubrumazine B (180), 25-hydroxyvariecolorin F (181), and 27-epi-aspechinulin D (182), together with the known analogue, neoechinulin B (175) (Figure 16). Detailed analysis of 1D and 2D NMR spectral and (+)-HRESI-MS data revealed that 178–182 are also prenylated indole diketopiperazine derivatives, comprising prenylated Trp and Ala. The Z geometry of the double bond at C-8 was established based on the downfield chemical shift of H-8 caused by the deshielding effect of the carbonyl group. The absolute configuration of Ala was determined as L by chiral HPLC of the hydrolysis products of 178–182 [96].
The relative configurations of C-22 and C-23 in 179–182 were established by a comparison of the experimental 1H and 13C chemical shift values with those obtained from GIAO (gauge-including atomic orbitals) NMR calculations at the mPW1PW91/6-31+G(d,p) level with DP4+ probability analyses. The results showed that the 1H and 13C chemical shift values of 178 and 182 matched well with the 12S*,22S*,23R* configurations. Moreover, the relative configurations obtained by an analysis of the 13C chemical shift values and NOESY correlations of the key protons of the acetonide of 182 were in agreement with those obtained from DP4+ probability analysis. Thus, the absolute configurations of C-22 and C-23 were tentatively assigned as 22S,23R by correlating with the absolute configuration of C-12 which was unambiguously established as 12S [96].
A chemical investigation of the CH2Cl2 fraction of the EtOAc extract of a culture of A. niger, isolated from a marine sponge Hyrtios proteus, which was collected in the Dry Tortugas National Park, Florida, yielded a previously reported dimeric cyclo dipeptide, asperazine (183) (Figure 16) [97].
The in vitro screen for the indoleamine 2,3-dioxygenase (IDO) inhibition of the crude extract of the solid culture of a marine sediment-derived fungus, Plectosphaerella cucumerina, collected at −100 m depth in Barkley Sound, British Columbia, led to the isolation of the undescribed plectosphaeroic acids A–C (184–186), together with the previously reported T988 A (187) (Figure 16). The planar structures of 184–186 were established by extensive analysis of 1D and 2D NMR and (−)-HRESI-MS data. Compounds 184 and 185 consist of a bis-(methylthio)-diketopiperazine fused with 3-methyl-2,3-dihydro-1H-indole, while 186 contains a trisulfide bridge across the diketopiperazine ring. Biogenetically, the indolylmethyldiketopiperazine core in 184 and 186 is formed by the condensation of Trp and Ser, while that of 185 is formed from Trp and Ala. Compounds 184–186 also featured an indole and a phenoxazinone substituents. The absolute configurations of the stereogenic carbons in 184–187 were determined as 3S,5aR,10bR,11S,12S by a comparison of their CD spectra with the literature values for structurally related leptosins [98].
The EtOAc extract of a solid rice culture of A. violaceofuscus, which was isolated from the inner part of a marine sponge Reniochalina sp., collected from the Xisha Islands in the South China Sea, furnished an undescribed indolylmethyl diketopiperazine dimer (188) (Figure 16). The relative configuration of 188 was assigned by diagnostic NOESY correlations. The absolute configuration of the stereocenters in 188 were determined as 2R,3R,11S,15R by Marfey’s method [99].
Three pairs of racemic spirocyclic diketopiperazine enantiomers containing anthrone moiety, variecolortins A–C (189–191) (Figure 16), were isolated from a marine sediment-derived fungus, Eurotium sp. SCSIO F452, obtained from the South China Sea. While 189 possesses a highly functionalized seco-anthronopyranoid carbon skeleton featuring a 2-oxa-7-aza-bicyclo[3.2.1]octane moiety, 190 and 191 contain a 6/6/6/6 tetracyclic cyclohexene–anthrone moiety. The planar structures of 189–191 were established by extensive 1D and 2D NMR spectral analysis. In the case of 189, the X-ray analysis not only established the relative stereochemistry as 12R*,21S*,32R*, but also indicated its racemic nature from the P21/n space group, which was supported by the lack of optical activity. The separation of a racemic 189 by chiral HPLC led to the obtention of pure (+)-189 and (−)-189. The stereostructures of both enantiomers were determined by a comparison of their calculated and experimental ECD spectra, establishing (12R,21S,32R)-189 for (+)-189. On the other hand, the racemic nature of 190 and 191 was deduced from their baseline ECD curves, as well as barely measurable optical rotations. The separation of racemic 190 and 191 by chiral HPLC resulted in the obtention of pure (−)-190 and (+)-190, as well as pure (−)-191 and (+)-191. A comparison of the calculated and experimental ECD spectra established the absolute structure of (+)-190 as 12S,22R-190 and of (−)-191 as 12S,22R-191 [100].
3.2.2. Cyclic Tripeptides
Chemical investigation of the extract of a co-fermentation of the strains BM-05 and BM-05ML of Aspergillus sp., an endophytic fungus isolated from a marine brown alga Sargassum sp., also furnished a cyclic tripeptide, psychrophilin E (192) (Figure 17). Detailed analysis of 1D and 2D NMR spectra and HRESI-MS established the structure of 192 as cyclo-(N-Ac-Trp-Pro-Abz). A chiral-phased gas chromatographic (GC) analysis of methyl N-(trifluoroacetyl)-Pro derivative, obtained from the hydrolysis of 192, led to the identification of L-Pro, thus establishing the absolute configuration of C-20 as 20S. However, the absolute configuration at C-2 could not be determined due to the decomposition of N-Ac-Trp during acid hydrolysis [81].
At the same time, Peng et al. isolated cyclic tripeptides, psychrophilins F–H (193–195) (Figure 17), in addition to 192, from the solid rice culture extract of a marine mud-associated fungus, A. versicolor ZLN-60, by using the OSMAC approach. Interestingly, the authors were able to obtain a suitable crystal of 192 for X-ray analysis using Cu Kα radiation and established the absolute configurations of C-2 and C-20 as 2S,20S. For 193, L-Pro was identified by advanced Marfey’s method, thus establishing the absolute configuration of C-20 as 20S. The configuration of C-2 in 193 is the same as that of 192 since they have similar CD curves. The absolute configurations of C-2, C-3, and C-20 of both 194 and 195 were established as 2S,3R,20S by a comparison of the calculated and experimental ECD spectra [101].
Purification of the fermentation broth extract of a halotolerant fungus, A. sclerotiorum PT06-1, which was isolated from the salt sediments from the Putian Sea Salt Field, Fujian, China, led to the isolation of undescribed cyclic aspochracin-type tripeptides containing an unsaturated fatty acid side chain, sclerotiotides A–K (196–206) (Figure 17), together with two previously reported tripeptides, JBIR-15 (207) and aspochracin (208) (Figure 17) [99].
The chemical transformation of 207 and 208 revealed that 199–206 were artifacts, probably formed during the fermentation process or subsequent isolation steps. The amino acid sequences of the compounds were established by HMBC correlations, while the absolute configurations of the amino acid residues were determined by Marfey’s method as (2E,4E,6E)-cyclo-[(N-Me-L-Ala)-L-Val-(Nα-octa-2,4,6-trienoyl-L-Orn)] (196), (2E,4E)-cyclo-[(N-Me-L-Ala)-(N-Me-L-Val)-(Nα-hexa-2,4-dienoyl-L-Orn)] (197), (2E,4E,6E)-cyclo-[(N-Me-L-Ala)-(N-Me-L-Val)-(Nα-octa-2,4,6-trienoyl-L-Lys)] (198), (2E,4E,6Z)-cyclo-[L-Ala-(N-Me-L-Val)-(Nα-octa-2,4,6-trienoyl-L-Orn)] (199), (2E,4E,6Z)-cyclo-[(N-Me-L-Ala)-(N-Me-L-Val)-(Nα-octa-2,4,6-trienoyl-L-Orn)] (200), (2E,4E)-cyclo-[(N-Me-L-Ala)-(N-Me-L-Val)-(Nα-6-oxohexa-2,4-dienoyl-L-Orn)] (201), and (2E,4E)-cyclo-[(N-Me-L-Ala)-(N-Me-L-Val)-(Nα-7-hydroxy-6-oxoocta-2,4-dienoyl-L-Orn)] (202) [102].
Compounds 203–206 were stereoisomers. Since the NMR data of 203 and 204 were identical to those of 205 and 206, respectively, 203 and 204, and 205 and 206 were suggested to be enantiotopic in the fatty acid moiety. Since the 3J(H6′,H7′) of 203–206 were small, all four compounds displayed a gauche conformation in the side chain. Moreover, 203–206 were produced by air oxidation of 208. Therefore, the structures of 203 and 205, and 204 and 206, were determined as threo- and erythro-(2E,4E)-cyclo-[(N-Me-L-Ala)-(N-Me-L-Val)-(Nα-6,7-dihydroxyocta-2,4-dienoyl-L-Orn)], respectively [102].
Sclerotiotide L (209) (Figure 17), another aspochracin-type cyclic tripeptide, was isolated from the EtOAc extract of a solid rice culture of A. violaceofuscus, obtained from the marine sponge Reniochalina sp., which was collected from the Xisha Islands in the South China Sea. The structure of 209 was established as (2′E,4′E)-cyclo-[(N-Me-L-Ala)-(N-Me-L-Val)-Nα-6′-methoxy-7′-hydroxyocta-2′,4′-dienoyl-L-Orn)] by NMR spectral analysis and Mafrey’s method [99].
The EtOAc extract of a fermentation of A. ochraceopetaliformis DSW-2, isolated from the sea water from Dongshan Island in Fujian Province of China, yielded an undescribed cyclic tripeptide, sclerotiotide M (210) (Figure 17), in addition to 197 and 201 (Figure 17). An extensive analysis of 1D and 2D NMR spectra and HRMS revealed the presence of Ala and Val, both of which are N-methylated, in addition to Orn. The interpretation of COSY and HMBC correlations allowed us to establish the sequence of amino acids as cyclo-(N-Me-Ala-N-Me-Val-Orn). The relative configuration of the stereocenters in 210 were established as 2S*,6S*,12S* by the calculated 13C NMR chemical shifts coupled with the DP4+ statistical method. Moreover, the optical rotation of 210 showed the same sign as those of the previously reported 196, 197, 199–201, suggesting that they share the same absolute configuration. Thus, the structure of 210 was identified as (2′E)-cyclo-[(N-Me-L-Ala)-(N-Me-L-Val)-(Nα-4′,4′-dimethoxy-2′-butenoyl-L-Orn)] [78].
3.2.3. Cyclic Tetrapeptides
Endolides A (211) and B (212) (Figure 18), two N-methylated peptides containing a rare amino acid, 3-(3-furyl)-Ala (33FAla), were isolated from the culture extract of the fungus Stachylidium sp., obtained from a marine sponge, Callyspongia sp. sf. C. flammea, which was collected in Bare Island, New South Wales, Australia. Extensive analysis of COSY and HMBC correlations allowed us to determine the amino acid subunits, as well as their sequence in 211 as cyclo-[N-Me-33FAla-Leu-N-Me-33FAla-Val], and in 212 as cyclo-[N-Me-33FAla-N-Me-33FAla-N-Me-33FAla-Val]. The configurations of the amino acid residues in 211 and 212 were established as L by advanced Marfey’s method, except for 33FAla since it underwent a degradation during a process of acidic hydrolysis. However, the authors were able to obtain a suitable crystal of 211 for X-ray crystallographic analysis showing that all four α protons have the same relative configuration. Thus, a combination of the results of Marfey’s and X-ray analyses led to the conclusion that the absolute configuration of the two N-Me-33FAla residues in 211 also has to be L. However, the authors assumed that the configuration of N-Me-33FAla in 212 was also L on the basis of biogenetic considerations [103].
In order to investigate the metabolic origin of the 33FAla moiety in the Stachylidium sp. peptides by using isotope-labeled precursors, the major metabolite 211 was targeted. Because the peptides are not produced in liquid media, solid biomalt sea-salt-containing agar media (BMS) were used for the investigation. Various 13C-labeled biosynthetic precursors such as L-[1-13C]-Phe, fully labeled [U-13C] glycerol, D-[1-13C] glucose, [1-13C] sodium acetate, and L-[Me-13C] Met were added to the solid BMS medium culture of the marine-derived fungus Stachylidium sp. 293 K04, which was isolated from the sponge Callyspongia sp. cf. C. flammea, collected from the coral reef in Bare Island, New South Wales, Australia. The results revealed that the exogenous Phe was not incorporated in the N-Me-33FAla residue. On the other hand, the biosynthesis of the N-Me-33FAla moiety involves both phosphoenolpyruvate and erythrose-4-phosphate as precursor molecules and the shikimate pathway is the biosynthetic route for the 33FAla moiety in 211, with Met as a provider of the N-Me group. Moreover, during the course of this study, two structurally unprecedented 33FAla-containing cyclic N-methylated peptide analogues, endolides C (213) and E (214) (Figure 18), and the previously described hirsutide (215) (Figure 18) were also isolated. The HMBC correlations established the structure of 213 as cyclo-(N-Me-Phe-Val-Phe-33FAla), and of 214 as cyclo-(N-Me-33FAla-Val-N-Me-33FAla-Phe). The absolute configurations of Val and Phe were established as L by the advanced Marfey’s method while the L-configuration of 33FAla was based on biogenetic considerations as well as from the results obtained from the X-ray analysis of 211 [104].
The EtOAc extract of a culure of A. violaceofuscus, isolated from the marine sponge Reniochalina sp., which was collected from the Xisha Islands in the South China Sea, yielded an unreported cyclic tetrapeptide, violaceotide A (216) (Figure 18). The amino acid sequence of 216 was determined as cyclo-(Thr-O-Me-Tyr-N-Me-Ala-Ile) by detailed HMBC, NOESY, and mass fragmentation analyses using a quadrupole-time-of-flight tandem mass spectrometer (Q-TOF-MS/MS). Marfey’s method was used to established the L-configuration of all amino acids in 216 [99].
The EtOAc culture extract of a marine-derived fungus, N. glabra KUFA 0702, isolated from a marine sponge Mycale sp., collected at a depth of 15–20 m from the coral reef at Samaesarn Island in the Gulf of Thailand, furnished two undescribed cyclic tetrapeptides, sartoryglabramides A (217) and B (218) (Figure 18). Extensive analysis of COSY and HMBC correlations were used to identify their amino acid residues and their sequence of 217 as cyclo-(Abz-Phe-Phe-Pro), and of 218 as cyclo-(Abz-Trp-Phe-Pro). The absolute configurations of the amino acids in both compounds were determined by X-ray analysis and confirmed by chiral HPLC analyses of their acidic hydrolysate, thus establishing complete structures of 217 and 218 as cyclo-(Abz-L-Phe-L-Phe-L-Pro) and cyclo-(Abz-L-Trp-L-Phe-L-Pro), respectively [70].
Li et al. reported a new cyclic tetrapeptide, cyclo-(L-Leu-trans-4-OH-L-Pro-D-Leu-trans-4-OH-L-Pro) (219) (Figure 18), from the co-culture broth extract of Phomopsis sp. K38 and Altenaria sp. E55, isolated from mangrove which was collected from Leizhou Peninsula, Guangdong Province, China. The amino acid residues and their sequence were determined by 1D and 2D NMR spectral analysis while Marfey’s method was used to determine the absolute configuration of the amino acids [105].
Mycelial and broth extracts of a mangrove endophytic fungus, Penicilluim sp. GD6, isolated from the stem bark of a mangrove tree, Bruguiera gymnorrhiza, which was collected in Zhanjiang, China, yielded an undescribed cyclic tetrapeptide 5,5ʹ-epoxy-MKN-349A (220) (Figure 18). The amino acid residues, i.e., two Pro, one Leu, and one Ile, were identified by extensive analysis of 1D and 2D NMR spectra. The amino acid sequence of 220 was achieved by HMBC correlations. The structure of 220 was found to resemble that of MKN-349A, a cyclic tetrapeptide previously reported from a marine-derived bacterium, Nocardiopsis sp. [106]. Due to a negligible amount of 220, the absolute configuration of its amino acid residues could not be determined by Marfey’s method. However, the authors proposed the L-configuration for all the amino acid residues on the basis of biogenic considerations [107].
The EtOAc extract of the mycelium of A. terreus SCSGAF0162, which was isolated from the tissue of a gorgonian, Echinogorgia aurantiaca, collected from Sanya, Hainan Province, China, furnished asperterrestide A (221) (Figure 18). Detailed analysis of 1D and 2D spectral data and the loss of fragments from the ESIMS of 221 led to the identification of Abz and other amino acid units, including 3-OH-N-Me-Phe, Ile, and Ala. The amino acid sequence of 221 was deduced as cyclo-(3-OH-N-Me-Phe-Abz-Ala-Ile) from NOESY correlations between α-protons and the protons of the NH groups of neighboring residues, as well as from HMBC correlations. The absolute configurations of Ala and Ile residues were determined as D by Marfey’s method, while the absolute configuration at C-10 was unambiguously assigned as S by a modified Mosher’s method in the NMR tube. Consequently, the structure of 221 was elucidated as cyclo-(Abz-D-Ala-D-Ile-3(S)-OH-N-Me-Phe) [108].
The EtOAc extract of a culture broth of A. flavipes, obtained from the gut of a marine isopod, Ligia oceanica, which was collected in Zhoushan, Zhejiang province of China, yielded a cyclic tetrapeptide 222 (Figure 18) whose structure consists of Pro, Ile, and two unusual amino acids, 5-methoxyanthranillic acid (5-OMe-Abz), and aminoacrylic acid. The identification of the amino acid residues was based on the interpretation of 1D and 2D NMR spectr, a while the amino acid sequence, cyclo-(5-OMe-Abz-Pro-Ile-aminoacrylic acid), was deduced from HMBC correlations and the major ESI-MSn fragment ion peaks. However, the configurations of Pro and Ile were not determined [109].
In a search for new metabolites that inhibit cancer cell proliferation from cultured marine fungi, Gu et al. reported the isolation of two undescribed cyclic tetrapeptides microsporins A (223) and B (224) (Figure 18), 12-membered ring cyclic peptides containing all α amino acids, from the culture extracts of the Microsporum cf. gypseum strain CNL-629, obtained from a marine bryozoan Bugula sp., which was collected in the U.S. Virgin Islands. Then, 1D and 2D NMR spectral analysis, including 1D-TOCSY and a combination of 1D-TOCSY with HMBC, revealed the presence of one Ala, one Phe, and one pipercolic acid (Pip) residues, and an unusual amino acid, 2-amino-8-oxo-decanoic acid (Aoda) in 223. The sequence of the four amino acids was established as cyclo-(Pip-Phe-Ala-Aoda) by HMBC correlations from the carbonyl carbon of each amino acid with the corresponding NH proton of the adjacent amino acid. By the same way, one Ala, one Phe, one Pip residues, and 2-amino-8-hydroxydecanoic acid, a hydroxyl derivative of Aoda were identified in 224. The configuration of Ala, Phe, and Pip were established as D-Pip, L-Phe, and D-Ala for 224. The S configuration of the stereogenic carbon of Aoda (C-22) in 223 was established based on NOE correlations between the α-H of Ala and the α-H of Aoda with Ala NH. However, the absolute configuration at the C-28 (the hydroxy-bearing carbon of the 2-amino-8-hydroxydecanoic acid residue) of 224 was not determined due to an insufficient amount of the compound to perform Mosher’s analysis [110].
3.2.4. Cyclic Pentapeptides
The solid-phase culture extract of Aspergillus sp. MEXU27854, isolated from a sample of sand collected from the intertidal zone of Caleta Bay in Acapulco, Guerreo, Mexico, furnished a new N-methyl cyclic pentapeptide, caletasin (225) (Figure 19). A combination of 1D and 2D NMR spectra, especially the TOCSY spectrum and MS/MS analysis, established the preliminary structure of 225 as cyclo-(Tyr-N-Me-Phe-Leu-Pro-Val). Marfey’s method was used to determine the absolute configuration of the amino acid residues; however, it was not possible to achieve a full separation of all the amino acids present in the mixture of the derivatized standards under any of the experimental conditions, especially between L-Leu and D-Val. A comparison of the mass profiles and mass values from HRESI-MS data for each amino acid confirmed the presence of L-Leu. Thus, the complete structure of 225 was established as cyclo-(L-Tyr-N-Me-L-Phe-L-Leu-L-Pro-L-Val) [111].
Asperpeptide A (226) (Figure 19), was isolated from the mycelial and broth extracts of Aspergillus sp. SX-20090B15, which was obtained from the inner tissue of a fresh gorgonian Muricella abnormaliz. Extensive analysis of HMBC and COSY correlations revealed the presence of one Tyr, two Ala, one Pro, and one 5-OH-Abz. The amino acid sequence was established as cyclo-(Pro-Ala-Ala-Tyr-5-OH-Abz) by HMBC correlations and the interpretation of fragment ions in the ESIMS/MS spectrum. The configurations of all the amino acid residues were determined as L by Marfey’s method. Thus, the complete structure of 226 was established as cyclo-(L-Pro-L-Ala-L-Ala-L-Tyr-5-OH-Abz) [46].
Fremlin et al. described the isolation of cotteslosins A (227) and B (228) (Figure 19) from the fermentation extract of A. versicolor (MST-MF495), which was obtained from a beach sand sample collected at a low tide from Cottesloe, Western Australia. The (+)-HRESI-MS and NMR spectral analysis of 227 revealed the presence of five amino acid residues. By using C3 Marfey’s analysis, the amino acids were identified as L-Tyr, N-Me-L-Tyr, L-Pro, L-Val, and L-Val. HMBC correlations were used to establish the sequence L-Tyr-N-Me-L-Tyr-L-Val-L-Val-L-Pro, and required cyclization from the L-Pro nitrogen to the L-Tyr carboxyl. By a comparison of the NMR spectra and (+)-HRESI-MS of 228 with those of 227, and with the support of C3 Marfey’s analysis, the sequence of 228 was established as L-Tyr-N-Me-L-Tyr-L-allo-Ile-L-Val-L-Pro. Thus, 228 is the L-Val1 to L-allo-Ile variant of 227 [112].
Lajollamide A (229) (Figure 19) was isolated from the culture broth extract of a marine-derived fungus, Asteromyces cruciatus 763, which was obtained from an unidentified decaying green alga, collected at La Jolla Shores, San Diego, USA. Then, a 1D and 2D NMR interpretation of 229 identified the individual amino acids as one Val, three Leu, and one N-Me-Leu residues. HMBC correlations from NH and NCH3 signals to the carbonyl groups of the adjacent amino acids were used to determine the amino acid sequence as N-Me-Leu-Leu-Leu-Leu-Val. The cyclic nature of 229 was confirmed by the degrees of unsaturation and HMBC correlations from the NCH3 signal of the N-Me-Leu residue to the carbonyl of Val. Chiral HPLC analysis of the acid hydrolysate of 229 resulted in the identification of the configurations of the amino acid residues as N-Me-L-Leu, two L-Leu, one D-Leu, and L-Val. However, the position of D-Leu could not be identified by the NMR data. A comparison of the NMR data of the three synthetic diastereomers of 229 with those of naturally occurring 229 to localize the position of D-Leu led to the conclusion that the D-Leu residue present in the hydrolysate was a result of the epimerization of the original L-Leu during the process of hydrolysis. Thus, the structure of 229 was established as cyclo-(N-Me-L-Leu-L-Leu-L-Leu-L-Leu-L-Val) [83].
JG002CPA (230) and JG002CPB (231) (Figure 19) were also isolated from the solid rice culture extract of a marine-derived fungus, A. allahabadii (strain number JG002), which was obtained from the underwater sediment collected off the coast of Jaeju-do Island, Korea. The 1D and 2D NMR spectral analysis, especially COSY and HMBC correlations, led to the identification of the amino acid residues and their sequences which were confirmed by high-resolution LC-MS/MS analysis, while the L-configuration of the amino acids was determined by the advanced Marfey’s method. Consequently, the complete structures of 230 and 231 were established as cyclo-(N-Me-L-Phe-L-Tyr-L-Pro-L-Leu-L-Ala) and cyclo-(N-Me-L-Phe-L-Tyr-L-Pro-L-Val-L-Ala), respectively [51].
The EtOAc extract of a solid rice culture of a marine-derived fungus, A. flocculosus 16D-1, isolated from the inner tissue of a marine sponge Phakellia fusca, which was collected from Yongxing Island, China, furnished an undescribed N-methylated pentapeptide, asperflomide (232) (Figure 19). An analysis of the 1D and 2D NMR spectra revealed the pentapeptide nature of 232. An analysis of the TOCSY spectrum revealed the presence of two Val (Val1 and Val2), one N-Me-Tyr, one Pro, and one Phe residues. An analysis of the COSY, HMBC, and TOCSY correlations established the amino acid sequence as Val1-N-Me-Tyr-Val2-Pro-Phe. The ROESY correlations were used to connect the final sequence as cyclo-(Val1-N-Me-Tyr-Val2-Pro-Phe), which was confirmed by MS/MS fragmentation. The absolute configuration of the amino acids in 232 was established by Marfey’s method. Thus, the complete structure of 232 was elucidated as cyclo-(L-Val1-N-Me-L-Tyr-L-Val2-L-Pro-L-Phe) [113].
The CH2Cl2-soluble fraction of a salt water culture of A. niger, isolated from a Caribbean sponge Hyrtiosproteus sp., furnished the previously reported cyclic pentapeptide, malformin C (233) (Figure 19) [97]. Later on, Uchoa et al. isolated the previously described malformin A1 (234) (Figure 19), together with malformin C (233) from a liquid culture extract of a marine sediment-derived fungus, A. niger BRF-074, collected from the Northeast Brazilian coast [65]. Compound 234 was also reported from the culture extract of a marine sediment-derived fungus Aspergillus sp. SCSIOW2 [114].
A chemical investigation of the culture broth extract of a marine-derived fungus, Aspergillus sp. SCSIO 41501, which was isolated from the gorgonian Melitodes squamata, collected from the South China Sea, furnished an undescribed aspergillipeptide D (235) (Figure 19). A detailed 1D and 2D NMR analysis, followed by Marfey’s analysis of the hydrolysate of 235, established the sequence of the amino acid residues and their absolute configurations as cyclo-(L-Val-N-Me-D-Tyr-O-Me-L-Tyr-O-Me-L-Tyr-L-Pro) [41].
The EtOAc extract of a culture of A. versicolor strain ZLN-60, which was isolated from the mud, at a 20 m depth, of the Yellow Sea, China, afforded two undescribed isomeric cyclic pentapeptides, versicotides A (236) and B (237) (Figure 19). An analysis of COSY and HMBC correlations of 236 and 237 revealed the presence of one Ala, two Abz, and two N-Me-Ala residues. Marfey’s method was used to determine the absolute configurations of the amino acid residues while HMBC correlations were used to establish the amino acid sequence. Consequently, the structures of 236 and 237 were established as cyclo-(L-Ala-N-Me-L-Ala-Abz-N-Me-L-Ala-Abz) and cyclo-(N-Me-L-Ala-N-Me-L-Ala-Abz-L-Ala-Abz), respectively [115].
Ding et al. used the LC-MS/MS-based molecular networking approach to assist the isolation of seven undescribed cyclopentapeptides, pseudoviridinutans A-G (238–244) (Figure 19), which feature a rare amino acid residue, O,β-dimethyltyrosine (O,β-diMeTyr), from the solid rice culture extract of A. pseudoviridinutans TW58-5, isolated from a hydrothermal vent sediment collected in Kueishantao, Taiwan. The planar structures of 238–244 were elucidated by a detailed analysis of HR ESI-QTOF MS, and 1D and 2D NMR spectroscopic data. The 1D and 2D NMR analysis of 238 revealed the presence of three common amino acids, viz. Ile, Gly, and homoleucine (Hleu), as well as two rare moieties, i.e., O,β-diMeTyr and (3-OH-Abz). HMBC correlations of α-H of one amino acid to the carbonyl of a neighboring amino acid established the amino acid sequence of 238 as cyclo-[(O,β-diMeTyr)-Ile-Gly-Hleu-3-OH-Abz]. The absolute configurations of Hleu and Ile were established as L-Hleu and D-allo-Ile, respectively, by Marfey’s analysis. Due to the unavailability of standard O,β-diMeTyr, the absolute configurations of its stereogenic carbons (Cα and Cβ) could not be determined by Marfey’s method. Since 238 was obtained in a suitable crystal for X-ray analysis, the absolute configurations of Cα and Cβ were determined as S and R, respectively. Thus, the complete structure of 238 is cyclo-[(αS, βR)(O,β-diMeTyr)-D-allo-Ile-Gly-L-Hleu-3-OH-Abz-] [116].
Compound 239 is similar to 238 except for the presence of Leu in 239 instead of Ile in 238. The amino acid sequence in 239 was established by the interpretation of HMBC correlations as cyclo [(O,β-diMeTyr)-Leu-Gly-Hleu-3-OH-Abz], and this sequence was supported by its fragmentation pattern in MS/MS. The absolute configurations of the Hleu and Leu residues in 239 were determined as L-Hleu and D-Leu, respectively, by Marfey’s method. Since 239 displayed the same coupling constant of α-H with β-H (Jα,β = 4.1 Hz) for the O,β-diMeTyr residue, as observed in 238, it, thus, supports the same relative configurations of Cα and Cβ. The absolute configurations of Cα and Cβ in 239 were suggested to be the same as those of Cα and Cβ in 238 based on the NMR data and biosynthetic origin [116].
By using the same methods, the amino acid residues, their stereochemistry, and sequence in 240 were established as cyclo-[-(αS, βR)(O,β-diMeTyr)-D-Val-Gly-L-Hleu-3-OH-Abz], while the structures of 241 was established as cyclo-[-(αS, βR)(O,β-diMeTyr)-D-allo-Ile-Gly-L-Phe-3-OH-Abz], of 242 as cyclo-[-(αS, βR)(O,β-diMeTyr)-D-Val-Gly-L-Phe-3-OH-Abz], of 243 as cyclo-[-(αS, βR)(O,β-diMeTyr)-D-Leu-Gly-D-Phe-3-OH-Abz], and of 244 as cyclo-[-(αS, βR)(O,β-diMeTyr)-L-Leu-Gly-D-Ile-3-OH-Abz] [116].
3.2.5. Cyclic Hexapeptides
An undescribed centrally symmetrical cyclohexapeptide, aspersymmetide A (245) (Figure 20), was isolated from the solid rice culture extract of a marine-derived fungus, A. versicolor (TA01-14), obtained from a gorgonian coral Carijoa sp. (GX-WZ-2010001), which was collected from the Weizhou coral reefs in the South China Sea. Detailed analysis of 1D and 2D NMR spectral data allowed us to identify the presence of one Pro, one Phe, and one Abz residues, which corresponded to half of the proposed molecular formula obtained from (+)-HRESI-MS, suggesting 245 as a symmetrical dimer. An analysis of HMBC correlations from the signals of NH proton of one amino acid to the carbonyl of the adjacent amino acid led to the deduction of their sequence as Phe-Abz-Pro. The HMBC correlation from α-H of Pro to the carbonyl of Phe connected the two half sequences to establish the whole cyclic structure of 245 as cyclo-(CO-Phe–Abz–Pro–Phe–Abz–Pro-N). The L-configuration of Pro and Phe was established by Marfey’s analysis. Thus, the complete structure of 245 was elucidated as cyclo-(L-Phe–Abz–L-Pro–L-Phe–Abz–L-Pro) [117].
Liang et al. described the isolation of four unreported cyclohexapeptides, simplicilliumtides J-M (246–249), together with the previously reported verlamelins A (250) and B (251) (Figure 20), from a fermentation broth extract of a deep-sea-derived fungus, S. obclavatum EIODSF 020, which was isolated from a marine sediment sample collected in the East Indian Ocean. Detailed 1D and 2D NMR analysis, especially COSY and HMBC correlations, suggested the presence of a long chain fatty acid, 5-hydroxytetradecanoic acid (5HTA), in 246, 250, and 251, while 247 and 248 contained 5-hydroxy-13-ketotetradecanoic acid (5H13KTA) and 5-hydroxy-12-ketotetradecanoic acid (5H12KTA), respectively. Marfey’s and Mosher’s methods were used to establish the absolute configurations of amino acids and the hydroxy-bearing carbons on long chain fatty acids, respectively. The sequences of amino acids were achieved by an analysis of the HMBC correlations of the α-proton of one amino acid to the carbonyl carbon of the next one. Thus, the complete structures of 246–249 were established as cyclo-(L-allo-Ile-D-Tyr-L-Gln-L-Pro-D-Ala-D-allo-Thr-5(S)HTA), cyclo-(L-Val-D-Tyr-L-Gln-L-Pro-D-Ala-D-allo-Thr-5(S)H13KTA), cyclo-(L-Val-D-Tyr-L-Gln-L-Pro-D-Ala-D-allo-Thr-5(S)H12KTA), and cyclo-(L-Val-D-Tyr-L-Gln-L-Pro-D-Ala-D-allo-Thr), respectively [49].
Later on, the same group has described the isolation of two undescribed cyclohexapeptides, simplicilliumtides N (252) and O (253) (Figure 20), together with 246, 250, and 251, from the combined broth and mycelia extracts of the same fungus. The identity and the sequence of the amino acids were established by a detailed analysis of the 1D and 2D NMR spectra, especially HMBC correlations, while the absolute configurations of the amino acid residues were determined by Marfey’s analysis. Thus, the structure of 252 was elucidated as cyclo-(L-Val-D-Tyr-L-Glu-L-Pro-D-Ala-D-allo-Thr-5HTA). The structure of 253 is similar to that of 252 except for the presence of a methyl ester of L-Glu in 253 instead of L-Glu in 252. Thus, 253 is a methyl ester of 252. The configuration of the 5HTA moiety in both 252 and 253 was inferred to be the same as that of 5HTA in 246, 250, and 251 [118].
Versicotide C (254) (Figure 20) was also isolated from the culture extract of a marine mud-derived fungus, A. versicolor ZLN-60. Detailed analysis of 1D and 2D NMR spectra, together with the molecular formula obtained from HRESI-MS, allowed us to identify six amino acid residues, i.e., two Ala, two N-Me-Ala, and two Abz residues. The sequence of the six amino acids was established, by observing HMBC correlations from α-H and N-Me protons, as well as amide NH signals to the carbonyl carbons of the connecting amino acid, as cyclo-(Me-Ala-Ala-Abz-Me-Ala-Ala-Abz), while Marfey’s analysis was used to identify the configuration of L-Ala and N-Me-L-Ala. Thus, the complete structure of 254 was elucidated as cyclo-(N-Me-L-Ala-L-Ala-Abz-N-Me-L-Ala-L-Ala-Abz). Interestingly, 254 displayed two sets of NMR signals of the two N-Me, indicating different configurations of the two corresponding amide bonds that are formed from N-Me-Ala. Thus, the cis/trans conformation was proposed for 254, and this was confirmed by variable-temperature 1H NMR experiments, which showed a significant decrease in the number of 1H NMR resonances measured at 80 °C compared to that measured at 25 °C [101].
The fermentation extract of A. sclerotiorum PT06-1, isolated from the Putian Sea Salt Field, Fujian, China, and cultured in a nutrient-limited hypersaline medium containing 10% NaCl, yielded two undescribed cyclic hexapeptides, sclerotides A (255) and B (256) (Figure 20). Detailed analysis of 1D and 2D NMR spectra, in combination with the molecular formula of 255 obtained from (+)-HRESI-MS, led to the identification of six amino acid residues viz. Thr, Ala, Phe, Ser, Abz, and dehydrotryptophan (∆-Trp). The amino acid sequence of 255 was established as cyclo-(Thr-Ala-Phe-Ser-Abz-∆-Trp) by HMBC correlations from the carbonyl carbon of one amino acid residue to the amide protons of the neighboring residue [119].
Compound 256, an isomer of 255, was found to have the same amino acid residues and sequence as those in 255. By the observation of the chemical shift values of H-29, H-38 and the amide proton of ∆-Trp, it was found that 256 is a geometric isomer of 255, differing only in the geometry of the double bond between C-28 and C-29 of ∆-Trp, being Z in 255 and E in 256. The absolute configurations of the amino acid residues in 255 and 256 were determined by chiral HPLC using a Crownpak CR(+) column. Thus, the structures of 255 and 256 were identified as cyclo-(L-Thr-L-Ala-D-Phe-D-Ser-Abz-Z-Δ-Trp) and cyclo-(L-Thr-L-Ala-D-Phe-D-Ser-Abz-E-Δ-Trp), respectively [119].
Compounds 255 and 256 were stable in the dark, but changed into each other in light, indicating that 255 and 256 are photointerconvertible. HPLC analysis showed that the ratio of 255 and 256 was 87:13 in the equilibrium, and both temperature and solvent had little influence on the photoequilibrium. All the evidences suggested that the photointerconversion proceeded through a radical mechanism and the Z-isomer of 255 is more stable. Therefore, it is most probable that 255 was really produced by A. sclerotiorum, while 256 was formed from the photoreaction of 255 during the fermentation or subsequent isolation steps [119].
Further three undescribed cyclic hexapeptides of the sclerotide family, sclerotides C-E (257–259) (Figure 20), were isolated together with 255 (Figure 20), from the extract of a solid rice culture of A. sclerotiorum SCSIO 41031, obtained from a soft coral which was collected in Beihai, Guangxi Province, China. Detailed analysis of 1D and 2D NMR spectra and HRESI-MS spectrum were used to identify the amino acid residues in these compounds, while their amino acid sequences were established by HMBC correlation of the α-H and NH signals of one amino acid to the carbonyl carbon of the connecting amino acid. Thus, the amino acid sequence of 257 was established as cyclo-(Thr-Ala-Phe-butanoic acid serine ester (BASE)-Abz-Δ-Trp). The geometry of the double bond between C-28 and C-29 in the Δ-Trp residue was the same as that in 255, i.e., Z-geometry. X-ray analysis not only confirmed the structure of 257 but also determined the configuration of its amino acid residues. Thus, the complete structure of 257 was elucidated as cyclo-(L-Thr-L-Ala-D-Phe-D-BASE-Abz-Z-Δ-Trp) [120].
An analysis of the 1D and 2D NMR and HRESI-MS spectra of 258 revealed that it had the same carbon backbone as that of 255, and absolute configurations of the amino acid residues were identified as L-Ser, L-Ala, D-Phe, and D-Ser by Marfey’s method. Thus, the structure of 258 was established as cyclo-(L-Ser-L-Ala-D-Phe-D-Ser-Abz-Z-∆-Trp). For 259, the 1D and 2D NMR and HRESI-MS spectral analysis revealed the presence of 2-(3-acetyl-2,6-dihydroxyphenyl)-N-∆-acetyltryptamine (ADPAT), in addition to Thr-Ala-Phe-Ser-Abz. Key HMBC correlations and the ESIMS/MS spectrum allowed us to establish the connectivity of the six amino acid residues, while Marfey’s analysis was used to determine the absolute configuration of the amino acids. Thus, the complete structure of 259 was established as cyclo-(L-Thr-L-Ala-D-Phe-D-Ser-Abz-Z-ADPAT) [120].
Similanamide (260) (Figure 20) is a cyclohexapeptide isolated from the EtOAc extract of a solid rice culture of A. similanensis KUFA0013, which was obtained from the marine sponge Rhabdermia sp., collected from the coral reef of the Similan Islands, Phang Nga Province, Thailand. Detailed analysis of the 1D and 2D NMR and HRESI-MS spectra was used to determine the six amino acid residues and their sequence as cyclo-(Abz-Val-Leu-Ala-N-Me-Leu-Pip). Chiral HPLC analysis of the acid hydrolysate of 260 led to the identification of the absolute configurations of the amino acids. Thus, the complete structure of 260 was elucidated as cyclo-(Abz-L-Val-D-Leu-L-Ala-N-Mel-L-Leu-D-Pip) [121].
The culture extract of Acremonium persicinum SCSIO115, which was obtained from a marine sediment collected from the South China Sea, resulted in the isolation of four undescribed cyclic hexapeptide siderophores, acremonpeptides A–D (261–264) (Figure 20), together with the aluminium complex of 264, Al(III)-acremonpeptide D (265) (Figure 18). An analysis of the 1D and 2D NMR and HRESI-MS spectra revealed that 261–265 all contain three units of 2-amino-5-(N-hydroxyacetamido)pentanoic acid (N5-OH-N5-Ac-L-Orn), which is able to chelate metal ions, besides other common amino acids. Marfey’s method was used to determine the absolute configurations of the amino acids in 261–264, and their sequences were established by an analysis of HMBC correlations from the α-H and NH of one amino acid to the carbonyl carbon of the connecting amino acid. Thus, the structures of 261–264 were elucidated as cyclo-(L-Phe-L-Leu-L-Ser-L-AcN(OH)-Orn1-L-AcN(OH)-Orn2-L-AcN(OH)-Orn3), cyclo-(L-Phe-L-Leu-L-Ala-L-AcN(OH)-Orn1-L-AcN(OH)-Orn2-L-AcN(OH)-Orn3), cyclo-(L-Phe-L-Leu-L-Phe-L-AcN(OH)-Orn1-L-AcN(OH)-Orn2-L-AcN(OH)-Orn3), and cyclo-(L-Phe-L-Leu-L-Trp-L-AcN(OH)-Orn1-L-AcN(OH)-Orn2-L-AcN(OH)-Orn3), respectively. The amino acid sequence of 265 was found to be the same as that of 264. However, the 13C NMR chemical shifts of the methyl carbons of the three acetyl groups were found to appear at lower frequencies, at δC 15.8, 16.3, and 16.7, when compared to those of the methyl carbons of the three acetyl groups in 264 (δC 20.5), which is consistent with the aluminium ion chelation by 264 to form 265 [122].
Tang et al. described the isolation of three unreported petrosamides A-C (266–268) (Figure 20) from the culture extract of Aspergillus sp. 151304, obtained from the inner tissue of a marine sponge Petrosia sp., which was collected from Yongxing Island, China. The six amino acid residues (one Pro, one Tyr, one Ac-Thr, one N-Me- Tyr, and two Val residues in 266; one Tyr, one N-Me-Tyr, one Pro, two Val, and one propylated Thr for 267; and two Tyr, two Val, one Pro and one Ac-Thr in 268) were identified by a detailed analysis of the 1D and 2D NMR spectra and HRESI-MS data, while the sequences of the amino acids were established using HMBC and ROESY correlations from NH or α-H of one amino acid to the carbonyl carbon of the connecting amino acid. The absolute configurations of the amino acid residues were determined by advanced Marfey’s analysis. Thus, the structures of 266–268 were elucidated as cyclo-(L-Tyr-N-Me-L-Tyr-L-Val2-L-Pro-L-Val1-N-Ac-Thr), cyclo-(L-Tyr-N-Me-L-Tyr-L-Val2-L-Pro-L-Val1-N-Pr-Thr), and cyclo-(L-Tyr2-L-Tyr1-D-Val2-L-Pro-L-Val1-N-Ac-Thr), respectively. The trans geometry for the proline amide bond in the three compounds was evident from the ΔδCβ-Cγ values, which is 5.0 ppm in 266, 5.1 ppm in 267, and 4.7 ppm in 268. The amino acid sequences were further confirmed by the corresponding ESIMS/MS fragments [123].
3.2.6. Cyclic Heptapeptides
By integrating molecular networking (untargeted HPLC-MS/MS) and 1H-NMR techniques, Shao’s group isolated three unreported cycloheptapeptides, asperversiamides A-C (269–271) (Figure 21), from the EtOAc extract of the coral-associated fungus A. versicolor (CHNSCLM-0063), collected from the South China Sea. Interpretation of 1D and 2D NMR spectra, together with the molecular formula obtained from HRESI-MS, led to the conclusion that 269 is a cyclic peptide containing one Phe, one Trp, one Ala, two Val, and two Ser residues. The sequence of the seven amino acids was deduced by HMBC correlations from the signals of NH to its respective adjacent carbonyl carbons, as cyclo-(Ser2-Trp-Ser1-Ala-Val1-Phe-Val2), which was confirmed by key ESI-MS/MS fragment ions. The same methods were applied to 270 and whose amino acid backbone was deduced as cyclo-(Ser-Trp-Ala2-Ala1-Val1-Phe-Val2) [124].
The configurations of amino acids were established by Marfey’s method which confirmed the presence of D-Ser, D-Trp, L-Ser, D-Ala, D-Val, and L-Phe in 269, and D-Ser, D-Trp, L-Ala, D-Ala, D-Val, and L-Phe in 270, respectively. However, the locations of D-Ser and L-Ser in 269, and L-Ala and D-Ala in 270 could not be confirmed. The problem was solved by performing a total synthesis of 269 and 270 and their corresponding isomers. By a comparison of the 1H NMR data and the retention times of the synthetic and naturally occurring counterparts, the structures of 269 and 270 were confirmed as cyclo-(D-Ser2–D-Trp–L-Ser1–D-Ala1–D-Val1–L-Phe–D-Val2) and cyclo-(D-Ser2–D-Trp–L-Ala2–D-Ala1–D-Val1–L-Phe–D-Val2), respectively. The amino acid residues of 271 were the same as those of 270, differing only in their connection sequence. HMBC correlations and Marfey’s method elucidated the structure of 271 as cyclo-(D-Ala2–D-Trp–L-Ser–D-Ala1–D-Val1–L-Phe–D-Val2), which was also supported by ESI-MS/MS key fragmentations [124].
In a further study, Shao’s group also used the ESI-MS/MS-based molecular networking strategy to isolate another four unreported cyclopeptides, asperheptatides–D (272–275) (Figure 21), in addition to 269–271, from the solid rice culture extract of the same fungus, obtained from the marine gorgonian coral Rumphella aggregate, which was collected from Nansha Islands in the South China Sea. Based on 1H and 13C NMR spectral analysis, the molecular formula obtained from the HRESI-MS, HMBC correlations, and key ESI-MS/MS fragment ions, the planar structures of 272 and 273 were established as cyclo-(Ser2-Trp-Ser1-Gly-Val1-Phe-Val2) and cyclo-(Ala2-Trp-Ala1-Ala3-Val1-Phe-Val2), respectively. Marfey’s method confirmed the presence of L-Phe, D-Trp, D-Val, D-Ser, and L-Ser in 272, and D-Ala, L-Ala, L-Phe, D-Trp, and D-Val in 273. The locations of L-Ser and D-Ser in 272, and L-Ala and D-Ala in 273 were determined based on their shared biogenesis with 269–271. Thus, the complete structures of 272 and 273 were elucidated as cyclo-(D-Ser2-D-Trp-L-Ser1-Gly-D-Val1-L-Phe-D-Val2) and cyclo-(D-Ala2-D-Trp-L-Ala1-D-Ala3-D-Val1-L-Phe-D-Val2), respectively. Since 274 and 275 were isolated in trace amounts, their planar structures were proposed as cyclo-(Ser-Trp-Met-Phe-Val-Phe-Val) and cyclo-(Ser-Trp-Ala-Gly-Val-Phe-Val) by ESI-MS/MS fragmentation experiments [125].
The unreported cordyheptapeptides C–E (276–278) (Figure 21) were isolated from a liquid culture extract of A. persicinum SCSIO115, which was obtained from a marine sediment sample collected at a depth of 205 m in the South China Sea. Detailed analysis of 1D and 2D NMR spectra and HRESI-MS of 276 revealed the presence of N-Me-Tyr, Phe, N-Me-Gly, Pro, N-Me-Phe, Leu, and Val. The sequence of the amino acids was determined by HMBC correlations and confirmed by analyses of the ESIMS/MS data of the quasi-molecular ion. The cyclic nature of the peptides was deduced by the degrees of unsaturation. Thus, the planar structure of 276 was established as cyclo-(N-Me-Tyr-Phe-N-Me-Gly-Pro-N-Me-Phe-Leu-Val). The absolute configuration of the amino acid residues was determined by X-ray analysis using Cu Kα radiation. Thus, the complete structure of 276 was established as cyclo-(N-Me-L-Tyr-L-Phe-N-Me-Gly-L-Pro-N-Me-D-Phe-L-Leu-L-Val). This was in agreement with the results obtained from Marfey’s analysis. The planar structure of 277 was established as cyclo-(N-Me-Tyr-Phe-N-Me-Gly-Pro-N-Me-Tyr-Leu-Val). By using Marfey’s analysis to determine the absolute configuration of the amino acids, the complete structure of 277 was elucidated as cyclo-(N-Me-L-Tyr1-L-Phe-N-Me-Gly-L-Pro-N-Me-D-Tyr2-L-Leu-L-Val) [126].
Detailed analyses of the COSY, HMQC, and HMBC correlations of 278 revealed that its structure was similar to that of 277. The 1H and 13C NMR data of 278 apparently suggested the presence of Ileu instead of Val in 278. However, detailed analysis of the 1H and 13C chemical shifts suggested the presence of L-allo-Ile instead of L-Ile. Although Marfey’s analysis was able to establish the absolute configurations of the amino acid residues, it was unable to distinguish between L-Ile and L-allo-Ile. Fortunately, a chiral-phase HPLC analysis of the acid hydrolysate of 278 was able to clarify that the amino acid was, in fact, L-allo-Ile. Thus, the complete structure of 278 was identified as cyclo-(N-Me-L-Tyr1-L-Phe-N-Me-Gly-L-Pro-N-Me-D-Tyr2-L-Leu-L-allo-Ile) [126].
The culture extract of Talaromyces sp. (CMB-TU011), isolated from an unidentified marine tunicate which was collected near Tweed Heads, NSW, Australia, furnished a hydroxamic acid-containing cyclic heptapeptide, talarolide A (279) (Figure 21). (+)-HRESI-MS and C3 and C18 Marfey’s analyses allowed the identification of six of the proposed amino acid residues as N-Me-L-Tyr, D-allo-Ile, N-Me-D-Ala, N-Me-D-Leu, L-Ala, and D-Ala, while NMR data revealed the presence of N-OH Gly residue. MS-MS fragmentations confirmed the amino acid sequence deduced by 2D NMR data, except for the location of L-Ala and D-Ala which was solved by C3 Marfey’s method. Thus, the structure of 279 was elucidated as cyclo-(N-Me-L-Tyr-N-Me-D-Ala-L-Ala-N-OH-Gly-N-Me-D-Leu-D-Ala-D-allo-Ile) [127].
The undescribed mortiamides A-D (280–283) (Figure 21) were isolated from the solid agar culture extract of a novel marine-derived fungus Mortierella sp. RKAG 110, isolated from a marine sediment at low tide at a depth of 30 cm in Frobisher Bay, Nunavut, Canada. The 1D and 2D NMR spectra, especially TOCSY experiments, were used to identify the amino acids, whereas HMBC and ROESY correlations were used to establish the amino acid sequences, which were confirmed by MS-MS fragmentation. The absolute configuration of the amino acids was determined by Mafrey’s method. When the peptides contained the D/L pair of amino acid such as 280 which contained L-Phe and D-Phe, their locations were determined by a comparison of the retention times from the LC-HRMS of the product obtained by the derivatization of a dipeptide fragment generated by partial hydrolysis with N-(5-fluoro-2,4-dinitrophenyl-5)-L-alaninamide (FDAA) with the retention times of the products of synthesized dipeptide standard derived in the same manner. Thus, the structures of 280–283 were elucidated as cyclo-(D-Phe-L-Phe-D-Val-D-Val-D-Val-L-Leu-D-Val), cyclo-(D-Phe-L-Phe-D-Val-D-Val-D-Val-L-Phe-D-Val), cyclo-(D-Ala-L-Ile-D-Ile-D-Val-D-Ile-L-Phe-D-Ile), and cyclo-(D-Leu-L-Phe-D-Ala-D-Ile-D-Ile-L-Phe-D-Val), respectively [128].
Unguisins A (284) and B (285) (Figure 21), two cyclic heptapeptides containing γ-aminobutyric acid (GABA), were obtained from the mycelia extract of a marine-derived fungus, Emericella unguis, which was isolated from a medusa, Stomolopus meliagris, collected in Venezuelan waters. The amino acid residues and GABA in both compounds were identified by FABMS, and 1D and 2D NMR analysis, while the amino acid sequences were established by an analysis of NOESY correlations. Marfey’s analysis was used to determine the absolute configuration of the amino acids. Consequently, the structures of 284 and 285 were elucidated as cyclo-(D-Val-D-Ala-D-Try-GABA-D-Ala-D-Val-L-Phe) and cyclo-(D-Val-D-Ala-D-Try-GABA-D-Ala-D-Val-L-Leu), respectively [129]. Compound 284 was also isolated from the chemically induced fermentation of a seaweed-derived fungus, A. unguis DLEP2008001 [130], and from the EtOAc extract of the deep-sea shrimp-associated fungus, A. unguis IV17-109 [39].
Further undescribed cycloheptapeptide incorporating GABA, unguisin E (286) (Figure 21), was isolated from the fermentation extract of Aspergillus sp. AF119, which was obtained from the soil collected in Xiamen Beach, China. The structure of 286 was quite similar to that of 284. The only difference is the presence of β-Me-Phe in 286 instead of Phe in 284. The sequence and relative stereochemistry of the amino acids were established by an analysis of NOESY correlations. Thus, the structure of 286 was elucidated as cyclo-(Ala-Trp-GABA-Ala-Val-β-Me-Phe-Val) [131].
A liquid culture extract of a marine-derived fungus, Scytalidium sp. CNC-310, isolated from a surface of a marine green alga Halimedia sp., which was collected from a patch reef at a depth of 15 m from the northern end of Long Island, the Bahamas, yielded two undescribed Aib-containing cyclic heptapeptides, scytalidamides A (287) and B (288) (Figure 21). The structures and sequences of amino acids were elucidated by detailed 1D and 2D NMR spectral analysis. The absolute configurations of the amino acid residues were determined by the advanced Marfey’s method. The structures of 287 and 288 were then established as cyclo-(L-Leu-L-Phe1-N-Me-L-Phe-L-Phe2-Aib-N-Me-L-Leu-L-Pro) and cyclo-(L-Leu-L-Phe1-N-Me-L-Phe-L-Phe2-Aib-N-Me-L-Leu-3-Me-L-Pro), respectively. The trans conformation of the Pro amide bond in 287 was determined by a small (3.5 ppm) differential value of 13C chemical shifts of Cβ and Cγ (∆βγ), while the absolute stereochemistry of (2S,3S)-3-Me-Pro in 288 was determined by the advanced Marfey’s method [132].
The combined acetone and EtOAc extracts of a solid rice culture of Aspergillus sp. SCSIO 41501, isolated from the marine gorgonian M. squamata Nutting, collected from the South China Sea, furnished three undescribed cyclic lipopeptides, maribasins C–E (289–291) (Figure 21), together with the previously reported maribasins A (292), B (293), and marihysin A (294) (Figure 21). The structures of 289–291 were tentatively established as cyclo-(D-Pro-L-Gln-L-Asn-L-Ser-D-Asn-D-Tyr-D-Asn-D-β-aminoisohexadecanoic acid), cyclo-(D-Pro-L-Gln-L-Asn-L-Ser-D-Asn-D-Tyr-D-Asn-D-β-aminohexadecanoic acid), and cyclo-(D-Pro-L-Gln-L-Asn-L-Ser-D-Asn-D-Tyr-D-Asn-D-β-aminoanteisoheptadecanoic acid), respectively, by a combination of a detailed analysis of 1D and 2D NMR spectra, especially HMBC and NOESY correlations, and Marfey’s analysis [50].
3.2.7. Cyclic Nonapeptides
Adachi et al. reported the isolation of two cyclic peptides containing highly N-methylated amino acids, clonostachysins A (295) and B (296) (Figure 22), from the EtOAc extract of a culture of a marine-derived fungus, Clonostachys rogersoniana (HJK9), obtained from a marine sponge Halicondria japonica, which was collected in Numazu, Japan. Extensive 1H and 13C NMR and 2D NMR spectral analysis, especially COSY, TOCSY, gHSQC, and gHMBC correlations, revealed the identity and the sequence of the amino acid residues in 295 and 296 as N-Me-Gly1-N-Me-Leu2-Pro3-N-Me-Tyr(OMe)4-Ala5-N-Me-Val6-N-Me-Leu7-N-Me-Ile8-N-Me-Ala9 and N-Me-Gly1-N-Me-Leu2-Pro3-N-Me-Tyr(OMe)4-Ala5-N-Me-Ile6-N-Me-Leu7-N-Me-Ile8-N-Me-Ala9, respectively. These sequences were further confirmed by LC-ESI MS/MS analyses. Marfey’s analysis revealed that all amino acids in 295 and 296 had the L-configuration. Moreover, N-Me-Ile was determined to be N-Me-L-Ile and not N-Me-L-allo-Ile, by a comparison of the HPLC retention time with those of the authentic samples [133].
3.2.8. Cyclic Decapeptides
The EtOAc extract of a culture of a marine-derived fungus, Sesquicillium microsporum RKAG 186, isolated from a marine sediment collected from the intertidal zone of Frobisher Bay, Nunavut, Canada, furnished four cyclic decapeptides, auyuittuqamides A–D (297–300) (Figure 23). The amino acid residues and their connectivity in these peptides were established by extensive 1D and 2D NMR spectral analysis, especially COSY, TOCSY, HSQC, HMBC, and ROESY correlations, while Marfey’s method revealed the L-configuration of all the amino acid residues in 297–300, except for the N-Me-Thr residue whose commercial standard of N-Me-D-allo-Thr was not available. As the LC-HRMS using a reversed-phase C18 column could not separate L-Ile from L-allo-Ile, the configuration at the β-carbon remained unassigned. Thus, the structures of 297–300 were elucidated as cyclo-(N-Me-Thr-L-Val-N-Me-L-Phe-Gly-Ile-L-Ser-L-Val-N-Me-L-Val-Gly-Leu), cyclo-(N-Me-Thr-L-Val-N-Me-Phe-Gly-L-Val-L-Ser-L-Val-N-Me-L-Val-Gly-Leu), cyclo-(N-Me-Thr-Ile-N-Me-L-Phe-Gly-Ile-L-Ser-L-Val-N-Me-L-Val-Gly-Leu), and cyclo-(N-Me-Thr-Ile-N-Me-L-Phe-Gly-Ile-L-Ser-Ile-N-Me-L-Val-Gly-Leu), respectively [134].
3.3. Depsipeptides
Depsipetides are a group of peptides whose amide group is replaced by an ester group of the carboxylic acid of the amino acid [37].
3.3.1. Linear Depsipeptides
The culture extract of a marine-derived fungus, Aspergillus sp. CMB-W031, isolated from an internal tissue of a mud dauber wasp Sceliphron sp., furnished a nitro depsitetrapeptide diketopiperazine, waspergillamide A (301) (Figure 24). Diagnostic 1H and 13C NMR signals, in combination with COSY and HMBC correlations, revealed the presence of Δ2,3-Leu, 3-OH-Val, Gly, and p-nitrobenzene residues. Marfey’s analysis revealed the D-configuration of the 3-OH-Val residue [135].
3.3.2. Cyclic Depsipeptides
The EtOAc extract of a culture of a marine-derived fungus, Sarocladium kiliense HDN11-112, obtained from the rhizosphere soil of the mangrove plant, Avicennia marina, furnished two depsipeptides, saroclides A (302) and B (303) (Figure 25). An analysis of the 1D and 2D NMR spectra suggested the presence of 7-hydroxy-4-methyl-3-oxo-dec-4-enoic acid (HMODA), in addition to Pro and Phe residues. Marfey’s analysis revealed the presence of D-Phe and L-Pro. The NOESY correlations between H2-6 and H3-11 established the E geometry of the double bond between C-4 and C-5 in HMODA. A single-crystal X-ray analysis of 302 established the absolute configurations of the stereogenic carbons as 7R,13S,18R and the trans conformations of the amid bonds between Pro and Phe [136].
An analysis of 1D and 2D NMR spectra revealed that 302 and 303 shared the same planar structure. However, Marfey’s analysis revealed the presence of D-Pro in 303 instead of L-Pro in 302. NOESY correlations not only established that 303 has the same E-geometry of the double bond between C-4 and C-5 as in 302 but also deduced the same absolute configuration at C-7 as in 302, i.e., 7R. Thus, 302 and 303 are a pair of epimers differing in the configuration of the Pro units, i.e., L-Pro and D-Pro, respectively. Interestingly, 302 was also isolated from the culture extract of S. lamellicola strain HDN13-430, which was obtained from a marine sediment collected in Antarctic Pritz Bay [136].
A chemical investigation of the mycelial extract of a solid fermentation of a marine-derived fungus, Acremonium sp. (MST-MF588a), isolated from an estuarine sediment sample collected from the Huon River near Franklin in Tasmania, Australia, led to the isolation of two major lipodepsipeptides, acremolides A (304) and B (305), together with two minor co-metabolites, acremolides C (306) and D (307) (Figure 25). A preliminary analysis of the 1H NMR (DMSO-d6) spectra of 304–307 revealed resonance doubling, at a ratio of 2:1, that coalesced at an elevated temperature, suggesting the presence of equilibrating isomers [137].
The NMR spectra of 304 (in DMSO-d6) exhibited not only the presence of both major and minor isomers but also the presence of Phe and Pro residues, which were confirmed as L-Pro and D-Phe by C3 Marfey’s analysis. A key gHMBC correlation between these residues (H2-5′ to C-1″) confirmed the amide linkage. Through 13C NMR chemical shift differences between Cβ and Cγ of Pro, the equilibrating isomers of 304 were attributed to cis and trans prolinyl amide rotamers. An analysis of 1D and 2D NMR spectra revealed the presence of the L-Pro-D-Phe-NH dipeptide substructure and that the remaining structural feature of 304 was a substituted fatty acid which is attached to D-Phe by an amide bond. (+)-ESIMS analysis of a mild basic hydrolysis product of 304, in combination a with NMR spectral analysis of the products obtained from the oxidation of 304 with pyridinium dichromate, revealed the structure of the fatty acid moiety. However, since the hydroxyl groups of the secondary alcohol of the hydroxy fatty acid portion did not react with the (S)-Mosher’s reagent, the absolute configurations of these stereogenic carbons could not be determined [137].
The HRESI-MS and 1D and 2D NMR analysis and C3 Marfey’s analysis of 305 revealed that its structure contained the same amino acid residues as 304, i.e., L-Pro and D-Phe. The only difference is the presence of a ketone group at C-11, instead of an OH group, in the fatty acid portion. The value of ∆βγ revealed the same 2:1 ratio of major cis versus minor trans rotamers as 304 [137].
The structures of 306 and 307 differ from those of 304 and 305 in that the D-Phe residue in 304 and 305 was replaced by an Ile and a Val residue, respectively. An analysis of Marfey’s 2,4-dinitrophenyl-5-L-alanine amide (DNP) derivatives obtained from 306 and 307 by C3 Marfey’s method unambiguously identified L-Pro and D-Ile, and L-Pro and D-Val, respectively. Moreover, in contrast to 304 and 305, an analysis of the 13C NMR data of 306 and 307 confirmed a preference for major trans versus minor cis prolinyl amide bond conformers, suggesting that the replacement of the bulky D-Phe (in 304 and 305) residue with either D-Ile (306) or D-Val (307) is responsible for an inversion of the preferred cis/trans balance in acremolide macrocycles [137].
The culture broth extract of the Calcarisporium sp. strain KF525, isolated from a water sample collected from the German Wadden Sea, yielded three undescribed cyclic depsipeptides, calcaripeptides A–C (308–310) (Figure 25). An analysis of HRESI-MS and 1D and 2D NMR data of 308 revealed the presence of a dipeptide consisting of the Pro and Phe residues, as well as a nonpeptidic portion that formed a 16-membered microcycle. The advanced Marfey’s analysis identified the configuration of Pro and Phe as L-Pro and L-Phe. Detailed analysis of the nonpeptidic portion revealed the presence of three secondary methyls, one tertiary methyl, one conjugated ketone carbonyl, and one E-double bond. The X-ray analysis allowed the determination of the relative configurations of 308. Through a combination of the results of the advanced Marfey’s analysis and X-ray analysis, the absolute configurations of the stereogenic carbons in 308 were established as 2S,6R,9R,2′S,2″S, as well as the cis conformation of the Pro amide bond [138].
Compound 309, whose NMR spectra were similar to those of 308, was elucidated by a detailed analysis of HRESI-MS and 1D and 2D NMR data as an analog of 308, but without the methyl group on C-2. The absolute configurations of its stereogenic carbons were not determined but were proposed to be the same as those in 308 [138].
On the other hand, the structure of 310 was very similar to that of 308 except for the lack of the E-double bond. Therefore, 310 consists of the L-Pro-L-Phe dipeptide and a nonpeptidic portion to form a 14-membered macrocycle. Like 309, the absolute configurations of the stereogenic carbons in 310 were not determined but were proposed to be the same as those in 308 [138].
The EtOAc extract of a liquid culture of an endophytic fungus, A. terreus (no. GX7-3B), obtained from a branch of a mangrove tree, Bruguiera gymnoihiza (Linn.) Savigny, which was collected from the salt coastline of the South China Sea in Guangxi province, China, furnished the previously reported cyclic hexadepsipeptide, beauvericin (311) (Figure 25) [139]. The structure of 311 consists of three D-α-hydroxyisovaleryl and three N-Me-L-Phe residues in alternating sequence, i.e., cyclo-(D-α-hydroxyisovaleryl-N-Me-L-Phe)3 [140]. Compound 311 was also isolated from a culture extract of an endophytic fungus, Fusarium sp. (No.DZ27), obtained from the bark of a mangrove tree, Kandelia cande (L.) Druce, which was collected from the Dongzhai mangrove, Hainan, China [141].
Two undescribed cyclic depsipeptides, guangomides A (312) and B (313), together with homodestcardin (314), a member of the destruxin family (Figure 23), were isolated from the EtOAc extract of a liquid culture of an unidentified marine-derived fungus, strain No. 001314c, obtained from a yellow fan-shaped sponge, which was collected at the coast of Guango, Papua New Guinea. Detailed analysis of 1D and 2D NMR spectra and HRESI-MS data of 312 revealed the presence of 2-hydroxyisocaproic acid (Hic), three Ala residues, one Phe residue, and a 2,3-dihydroxyisovaleric acid (Hiv) subunit. The connectivity of the amino acid residues and two hydroxy carboxylic acids were established by HMBC correlations. The absolute configuration of N-Me-D-Phe was determined by Marfey’s method while the relative configurations of the stereogenic carbons were established by X-ray analysis. Through a combination of the absolute configuration of N-Me-D-Phe and the relative configurations from the X-ray data, the final absolute stereostructure of 312 was assigned as 2S,9S,13S,19S,24R,28R [142].
Since HRESI-MS data displayed the molecular formula of 313 with one oxygen atom less than that of 312, and the 1H NMR showed diastereotopic H3-15 and H3-16 as doublets, it was suggested that Hiv in 312 was replaced by 2-hydroxyisovaleric acid, which was confirmed by 2D NMR analysis. The absolute configurations of the stereogenic carbons in 313 were proposed to be the same as those in 312 on the basis of biogenetic origin [142].
The 1H and 13C NMR spectra of 314 were very similar to that of homodestruxin B [143] and roseocardin [144]. Detailed analysis of the 2D NMR data of 314 revealed the presence of β-Me-Pro and N-Me-Leu residues instead of a Pro residue in homodestruxin B and N-Me-Val residue in roseocardin, respectively. Moreover, 314 and homodestruxin B displayed identical shifts of the chiral centers C-6, C-11, C-12, C-19, and C-20. On the other hand, the chiral centers C-26, C-27, and C-33 of 314 showed the same chemical shifts as the corresponding chiral centers of roseocardin. Moreover, the significant NOE observed between H-26 and H-28 was also consistent with these stereochemical proposal [142].
The mycelium extract of a marine-derived fungus, Fusarium sp., collected from the surface of a seagrass, Halodule wrightii, which was obtained in the inner lagoon of Little San Salvador Island, Bahamas, furnished a cyclic pentadepsipeptide, sansalvamide A (315) (Figure 25). Detailed interpretation of 1D and 2D NMR spectral data and HRESI-MS, revealed the presence of Val, Phe, two Leu, and leucic acid (O-Leu) residues. Chiral capillary GC analysis of the hydrolysate of 315 using the authentic standards for comparison established the L-configuration for the four amino acid residues in 315. NOESY correlations between α-H of Leu1 and α-H of O-Leu suggested that both protons are on the same (α) face; therefore, the O-Leu residue also possesses the L-configuration [145].
The culture extract of Fusarium sp. CNL-619, isolated from a green alga, Avrainvillea sp., which was collected from a mangrove at a depth of 2 m in St. Thomas at Bovoni Cay, United States Virgin Islands, furnished N-methylsansalvamide (316) (Figure 25). The structure of 316 is closely related to 315. An analysis of 1D-TOCSY data established the presence of Val, Leu, and Phe residues. Moreover, 1D-TOCSY, HMQC, and HMBC data further revealed the presence of one N-Me-Leu and one O-Leu residues. The amino acid sequence of the cyclic peptide was established by an analysis of HMBC correlations from the NH proton of the amino acid to the carbonyl carbon of the connecting amino acid. Marfey’s method revealed that all the amino acids in 316 have an L-configuration. The absolute configuration at C-2 in O-Leu was determined as 2S by the hydrolysis of 316, followed by methylation to give a linear peptide with free hydroxyl group at C-2 for the preparation of Mosher’s ester [146].
An undescribed cyclic pentadepsipeptide, acremonamide (317) (Figure 25), was reported from a culture broth extract of a marine-derived fungus, Acremonium sp. CNQ-049, which was isolated from a sediment collected at a depth of 25 m from the coast of La Jolla, California. Detailed analysis of 1D and 2D NMR and HRESI-MS spectra led to the identification of N-Me-Phe, N-Me-Ala, Val, Phe, and Hiv. The stereochemistry of the four amino acids was determined as L-configured by the advanced Marfey’s method. The absolute configuration of the stereogenic carbon in Hiv was determined as R by GC-MS analysis, by a comparison of the retentime of the O-pentafluoropropionylated (−)-menthyl ester of Hiv in 317 with those of the the O-pentafluoropropionylated (−)-menthyl ester of the standard S-Hiv and R-Hiv [147].
An undescribed zygosporamide (318) (Figure 25) was isolated from the fermentation broth extract of a marine-derived fungus, Zygosporium masonii (CNK458), obtained from a marine cyanobacterium, collected off the Island of Maui, Hawaii. A combination of HRESI-MS data and 1D and 2D NMR spectral analysis revealed the presence of two Leu, two Phe, and one O-Leu residues. Marfey’s analysis indicated that all the amino acids in 318 have an L-configuration, except for one Leu residue which has a D-configuration. The absolute configuration at C-32 of O-Leu was determined as 32S by Mosher’s method of the linear peptide obtained by the hydrolysis of 318 [148].
Alternaramide (319) (Figure 25) was reported from the EtOAc culture extract of a marine-derived fungus, Altenaria sp. SF-5016, isolated from a sediment sample collected from the shoreline in the Masam Bay area, Korea. The 1D and 2D NMR spectral analysis revealed the presence of two Phe, two Pro, and one Hiv residues. Marfey’s analysis identified two D-Phe and two L-Pro residues. The absolute configuration at C-2 of Hiv was determined as 2S by Mosher’s method of a linear peptide with free OH group obtained from hydrolysis of 319. Thus, the overall absolute configurations of 319 were established as 2S,7S,12R,21S,26R [149].
Boot et al. described the isolation of six previously reported cyclic depsipeptides belonging to the destruxin class, viz. destruxins A (320), B (321), B2 (322), desmethyldestruxin B (323), destruxin E chlorohydrin (324), and destruxin E2 chlorohydrin (325) (Figure 25) [150], from a culture extract of a marine sponge-associated fungus, Metarrhizium sp. (strain number 001103), obtained from a marine sponge, Pseudoceratina purpurea (coll. no. 00103), which was collected in Fiji [53].
Capon et al., in their search for new antiparasitic compounds, have isolated aspergillicins A–E (326–330) (Figure 25), depsipeptides that incorporated the unusual N-Me-L-Tyr-O-Me residue from the mycelia extract of A. carneus (MST-MF156), obtained from an estuarine sediment which was collected in Tasmania, Australia. The 1H and 13C NMR spectra of 326 indicated the presence of six amino acid residues, including two Pro, one Val, one Ile, and both N- and O-methylated Tyr. The degree of unsaturation revealed that 326 was a cyclic peptide. ESI-MS analysis of the mixture of amino acids obtained from acid hydrolysis of 326 confirmed the presence Pro, Val, Thr, Ile, and N-Me-Tyr. The assignment of the absolute stereochemistry of 326 was achieved by the advanced Marfey’s method. Thus, the structure of 326 comprises of L-Val, N-Ac-L-Thr, D-Ile, two L-Pro, and N-Me-L-Tyr-O-Me. In this case, the author has used the Marfey’s derivatives on a chiral HPLC column to distinguish between D-Ile and D-allo-Ile. Therefore, the complete structure of 326 was elucidated as cyclo-(N-Me-L-Tyr-O-Me-L-Pro-L-Pro-D-allo-Ile-O-Ac-L-Thr-L-Val). Moreover, the small values of the difference in 13C NMR shifts between C-12 and C-13, as well as between C-17 and C-18, confirmed the trans configuration of the amide bonds of the two Pro residues [71].
A Combination of the molecular formula, obtained by HRESI-MS, 1H NMR spectral analysis, ESI-MS analysis of the acid hydrolysate, and HPLC analysis of the Marfey’s derivatised acid hydrolysate confirmed that the structure of 327 differed from that of 326 by a replacement of the D-allo-Ile residue by a D-nor-Val residue. Compounds 328 and 329 were minor components and were isolated as a mixture. Despite this, an analysis of 1H NMR and ESIMS data proposed that 328 and 329 differed from 326 in the presence of the N-Me-L-Tyr-O-Me residue instead of the N-Me-L-Phe and L-Phe residues, respectively. The 1H NMR spectrum of the minor co-metabolite 330 was very similar to that of 326. However, 330 showed more 14 atomic mass units than 326. Marfey’s method with chiral HPLC analysis confirmed that Val in 326 was replaced by L-allo-Ile in 330 [71].
The mycelial extract of a marine-derived fungus, Scopulariopsis brevicaulis NCPF 2177, isolated from the inner tissue of a marine sponge, Tethya aurantium, which was collected in the Limski Fjord, Croatia, furnished two undescribed cyclohexadepsipeptides, scopularides A (331) and B (332) (Figure 25). Detailed analysis of HRESI-MS, 1D and 2D NMR spectra and HMBC correlations of 331 revealed the presence of Gly, Phe, Ala, Val, Leu, and 3-hydroxy-4-methyldecanoic acid (HMDA). The amino acid sequence could not be established unambiguously with NMR methods. However, with the aid of MS/MS fragment ions, the amino acid sequence was established as cyclo-(HMDA-Gly-Val-Leu-Ala-Phe). The configurations of the amino acid residues were established by the derivatization of the acid hydrolysis product of the peptide with Nα-(2,4-dinitro-5-fluorophenyl)-L-valinamide (L-FDVA), and HPLC analysis of the derivatives. A comparison with the amino acid standards derived with L- and D-FDVA showed that the Val, Ala, and Phe residues all have the L-configuration, while Leu was D-configured. The absolute configuration of the stereogenic center C-3 of HMDA was established as 3S by Mosher’s method. Thus, the complete structure of 331 was elucidated as cyclo-[(S)-HMDA-Gly-L-Val-D-Leu-L-Ala-L-Phe)]. The HRMS and NMR data of 332 revealed that it had the same amino acid residues as 331. However, it contains a 3-hydroxy-4-methyloctanoic acid (HMOA) instead of HMDA in 331 [151]. Compounds 331 and 332 were also reported from the EtOAc extract of the culture of A. flavus, isolated from the soft coral Sarcophyton ehrenbergi, collected from the Red Sea, Egypt [152].
Three undescribed cyclohexadepsipeptides, oryzamides A–C (333–335), were isolated, together with two isolation artifacts oryzamides D (336) and E (337), and 331 (Figure 25), from the mycelia extract of a marine sponge-associated fungus, Nigrospora oryzae PF18, which was obtained from a marine sponge, Phakellia fusca, collected from Yongxing Island. An analysis of HRESI-MS, 1D and 2D NMR spectral data of 333 led to the identification of one Ala, one Gly, one Val, two Leu residues, and HMDA. HMBC correlations allowed us to establish the amino acid sequence as Leu1-Ala-Leu2-Val-Gly-HMDA, which was further confirmed by the MS/MS fragment ion series. Given the degree of unsaturation, it was suggested that the Leu1 residue was connected to the HMDA moiety by an ester linkage. The absolute configuration of 333 was establish by a single-crystal X-ray diffraction analysis using Cu Kα radiation, allowing the assignments of the amino acid residues as L-Leu1, L-Ala, D-Leu2, and L-Val, as well as the configuration the stereogenic centers in the HMDA moiety as 3S,4S. Thus, the complete structure of 333 was elucidated as cyclo-[(L-Leu1-L-Ala-D-Leu2-L-Val-Gly-(3S,4S)-HMDA)] [153].
The 1H and 13C NMR spectra of 334 were similar to those of 333 with some differences in the aromatic region. An analysis of the 2D NMR spectra of 334 led to the conclusion that the Leu1 residue in 333 was replaced by the Tyr residue in 334. HMBC correlations allowed us to establish the connectivity as cyclo-(Tyr-Ala-Leu-Val-Gly-HMDA). By using Marfey’s analysis, it was found that all the amino acid residues have the L configuration except Leu whose configuration is D. The absolute configurations of C-3 and C-4 of HMDA were proposed to be the same as those in 333 on the basis of biogenetic origin. Thus, the complete structure of 334 was elucidated as cyclo-[(L-Tyr-L-Ala-D-Leu-L-Val-Gly-(3S,4S)-HMDA)] [153].
An analysis of the HRESI-MS, 1D and 2D NMR spectra of 335 revealed that it contains Met instead of Leu1 residue in 333. Thus, the planar structure of 335 was identified as cyclo-(Met-Ala-Leu-Val-Gly-HMDA), which was supported by the analysis of the ESIMS/MS fragments. The advanced Marfey’s method established the configurations of the amino acids as L-Met, L-Ala, D-Leu, and L-Val. Thus, the complete structure of 335 was elucidated as cyclo-[(L-Met-L-Ala-D-Leu-L-Val-Gly-(3S,4S)-HMDA] [153].
The two minor metabolites, 336 and 337, are isomers as they showed the same molecular formula. Their molecular formula have one oxygen atom more than that of 335. The 1H and 13C NMR spectra of 336 and 337 were almost identical to each other and very similar to those of 335. A careful analysis of the 1D and 2D NMR data of 336 and 337 revealed that they are two diastereomers at a methionine sulfoxide residue resulting from oxidation of 335. The absolute configurations of 336 and 337 were partially determined as L-Ala, L-Val, and D-Leu by the advanced Marfey’s method. In the case of the previously reported 331 whose stereocenter C-4 of the HMDA moiety had not been assigned for its absolute configuration [144], the X-ray analysis of 331 was able to establish the absolute configuration at C-4 of HMDA as 4S [153].
The extract of a solid rice culture of Beauveria feline EN-135, isolated from an unidentified marine bryozoan, which was collected from Huiquan Gulf of the Yellow Sea, furnished three undescribed cyclohexadepsipeptides of the isaridin family, isaridin G (338), desmethylisaridin G (339), desmethylisaridin C1 (340) (Figure 26), together with three previously reported congeners, isaridins A (341), B (342), and E (343) (Figure 26) [154,155,156], all containing 2-hydroxy-4-methylpantanoic acid (HMPA) unit. An analysis of the HRESI-MS, 1H, and 13C NMR spectra revealed that 338 is a cyclic hexadepsipeptide with a Tyr residue. An analysis of COSY and HMBC correlations revealed the presence of HMPA, and five amino acid residues, viz. Pro, Tyr, N-Me-Val, N-Me-Leu, and β-Ala. The sequence of the amino acids was preliminarily established by observing HMBC correlations from the H-α and amide NH signals to the carbonyl of the connecting amino acid, as cyclo-(HMPA1-Pro2-Tyr3-N-Me-Val4-N-Me-Leu5-β-Ala6). Single-crystal X-ray diffraction using Cu Kα radiation established the S configuration for the HMPA1 and L configuration for all amino acid residues. Thus, the structure of 338 was established as cyclo-(S-HMPA1-L-Pro2-L-Tyr3-N-Me-L-Val4-N-Me-L-Leu5-L-β-Ala6) [157].
The molecular formula of 339, determined by HRESI-MS, suggested that 399 might be a demethylated congener of 338. Further interpretation of COSY, HSQC, and HMBC spectra confirmed the replacement of N-Me-Leu5 in 338 by a Leu5 residue in 339. X-ray analysis of 339 confirmed the presence of S-HMPA and the L configuration of all the amino acid residues. Thus, the structure of 339 was elucidated as cyclo-(S-HMPA1-L-Pro2-L-Tyr3-N-Me-L-Val4-L-Leu5-L-β-Ala6) [157].
Detailed analysis of HRESI-MS and 1D and 2D NMR spectra of 340 identified its planar structure as cyclo-(HMPA1-Pro2-Phe3-N-Me-Val4-Leu5-β-Ala6). X-ray analysis of 340 established the S configuration for HMPA and the L-configuration for all amino acids. Therefore, the complete structure of 340 was established as cyclo-(S-HMPA1-L-Pro2-L-Phe3-N-Me-L-Val4-L-Leu5-β-L-Ala6). A conformation study of 338–340 using X-ray diffraction revealed that the HMPA1-Pro2 and N-Me-Val4-N-Me-Leu5 amide bonds in 338 showed cis conformations, while those in 339 and 340 showed trans conformations [157].
Hou et al., by using an integration of the LC-MS/MS-dependent molecular network and 1H NMR techniques, were able to target seven unreported cyclohexadepsipeptides, chrysogeamides A–G (344–350), in addition to the previously reported 331 and 332 [151], and nodupetide (351) [158] (Figure 26), from a solid rice-potato culture extract of an endophytic fungus, P. chrysogenum (CHSCLM-0003), isolated from a gorgonian coral, Carijoa sp. (GX-WZ-2010001), collected from Weihou coral reefs, South China Sea. An analysis of HRESI-MS, 1D and 2D NMR spectra of 344 led to the identification of one Gly, one Ala, one Leu, and two Val residues, in addition to a HMOA unit. HMBC correlations from the amide NH protons to their adjacent carbonyls established the connectivity of the amino acids in 344 as cyclo-(Val2-Ala-Leu-Val1-Gly-HMOA) and oxygen on C-24 should be part of an ester group between HMOA and Val2. Marfey’s analysis established the configuration of the amino acids as L-Ala, L-Val, and D-Leu. Thus, the structure of 344 was fully elucidated as cyclo-(L-Val2-L-Ala-D-Leu-L-Val1-Gly-HMOA) [159].
The NMR spectra of 345 were similar to those of 344. However, a careful analysis of 1D and 2D NMR spectra of 345 revealed the presence of an HMDA moiety instead of HMOA. HMBC correlations, ESI-MS/MS, and Marfey’s method elucidated the full structure of 345 as cyclo-(L-Val-L-Ala-D-Leu-L-Val-Gly-HMDA) [159].
Detailed analysis of 1D and 2D spectra of 346 revealed the presence one Gly, two Leu, one Ala, one Phe residues, and HMDA. HMBC correlations, ESI-MS/MS, and Marfey’s method elucidated the full structure of 346 as cyclo-(L-Phe-L-Ala-D-Leu-D-Leu-Gly-HMDA) [159].
Compounds 347 and 349 were isolated as a 1:0.5 mixture. A combination of careful analysis of the 1H and 13C NMR data, molecular formula, HMBC correlations, and Marfey’s analysis confirmed the structures of 348 and 350 as cyclo-(L-Phe-L-Ala-D-Leu-D-Leu-Gly-HMOA) and cyclo-(L-Phe-L-Ala-D-Leu-L-Pro-Gly-HMDA), respectively [159].
FJ120DPA (352) (Figure 26) was also isolated from a semi-solid rice culture extract of a marine sediment-derived fungus, A. ochraceopetaliformis. Detailed analysis of HRESI-MS, 1D and 2D NMR spectra to identify the amino acids and HMBC correlations to establish the amino acid connectivity, as well as Marfey’s analysis, led to the elucidation of the structure of 352 as cyclo-(N-Me-L-Phe-L-Ala-Ac-L-Thr-D-Val-L-Pro-L-Tyr) [51].
The extract of a co-culture of a marine-derived fungus, Emericella sp. (strain CNL-878), isolated from the surface of a green alga Halimeda sp., collected at Madang Bay, in Papua New Guinea, and a marine actinomycete, Salinispora arenicola (strain CNH-665), isolated from a sediment sample collected from the Bahamas, resulted in the isolation of the unreported depsipeptides, emericellamides A (353) and B (354) (Figure 26) [160].
Detailed analysis of 1D and 2D NMR spectra including the HSQC, DEPT, COSY, and 1H-1H TOCSY of 353 led to the identification of one Gly, two Ala, one Val, one Leu, and 3-hydroxy-2,4-dimethyldecanoic acid (HDMD). HMBC and ROESY correlations allowed the determination of the connectivity of the amino acid sequence and the location of HDMD as cyclo-(Ala1-Ala2-Leu-Val-Gly-HDMD). Marfey’s analysis determined the L-configuration of the amino acids. The relative configurations of the stereogenic carbons of HDMD were determined by a J-based configurational analysis using 3JHH, 3JCH, and NOE correlations as (21R*,22R*,23S*). Mosher’s method determined the absolute configuration at C-22 as 22R. Therefore, the absolute configurations of the stereogenic carbons of HDMD were established as 21R,22R,23S. Thus, the complete structure of 353 was elucidated as cyclo-[L-Ala1-L-Ala2-L-Leu-L-Val-Gly-(21R,22R,23S)-HDMD] [160].
By using the same approach, 354 was found to contain the same amino acids as 353, i.e., two L-Ala, one L-Val, and one L-Leu; however, the side chain was identified as 3-hydroxy-2,4,6-trimethyldodecanoic acid (HTMD). The relative configurations of C-21, C-22, and C-23 were proposed to be the same as those in 353, i.e., 21R*,22R*,23S*, based on the high degree of similarity of their analogous 1H coupling constants and 1D NOE correlations. Since ROESY correlations between H-2 and H-22, and between 19-NH and H-22 in 354, were similar to those of 353, the authors suggested that it was highly indicative of the same absolute configuration at C-22 for both compounds. On the basis of consistent ROESY correlations, the absolute stereochemistry of HTMD was established as 21R,22R,23S,25S. It is worth mentioning that the marine actinomycete S. arenicola induced the culture of Ermericella sp. to produce ca. 100 times more than the single culture [160].
The solid rice culture extract of a soft coral-derived fungus, A. sclerotiorum SCSIO 41031, also furnished a lipodepsipeptide, scopularide I (355) (Figure 26). The HRESI-MS and 1D and 2D NMR analysis revealed the presence of one Val, two Ala, and one Leu residues, in addition to the 24-hydroxy-25-methyllauric acid (HMLA) moiety. The complete structure of 355 was confirmed by single-crystal X-ray analysis using Cu Kα radiation as cyclo-[L-Val-L-Ala1-D-Leu-L-Ala2-(24S,25S)-HMLA] [120].
An undescribed N-methylated depsipeptide, asperflosamide (356) (Figure 26), was also isolated from the fermentation extract of a marine sponge-associated fungus, A. flocculosus 16D-1. An analysis of the 1H, 13C NMR, COSY, HSQC, and TOCSY spectra of 356 revealed the presence of Tyr, N-Ac-Thr, Val, Pro, Leu, and N-Me-Phe residues. HMBC and ROESY correlations were used to determine the sequence of six amino acids as Tyr-N-Ac-Thr-Val-Pro-Leu-N-Me-Phe. Considering the degree of unsaturation, the cyclic nature of 356 was proposed, suggesting that the N-Ac-Thr residue was attached to the moiety by an ester bond. Thus, the planar structure of 356 was established as cyclo-(Tyr-N-Ac-Thr-Val-Pro-Leu-N-Me-Phe). The stereochemistry of the amino acid residues was determined as an L configuration by Marfey’s method. Therefore, the complete structure of 356 was assigned as cyclo-(L-Tyr-N-Ac-L-Thr-L-Val-L-Pro-L-Leu-N-Me-L-Phe). Moreover, the values of ΔδCβ-Cγ of the L-Pro residue indicated a trans geometry for the Pro amide bond [113].
By integrating genomics and metabolomics approaches to discover new cyclopeptides from the EtOAc extract of a solid rice culture of a marine-derived fungus, B. feline SYSU-MS7908, isolated from a marine ascidian sample (Styela plicata) collected from the North Atoll of the Xisha Islands in the South China, Jiang et al. described the isolation of six undescribed cyclohexadepsipeptides of the isaridin family, isaridins I–N (357–362) (Figure 26), together with the previously reported isaridin E (343). The 1D and 2D NMR spectra, including COSY, NOESY HSQC, HMBC, and TOCSY, revealed the presence of HMPA in all the isolated compounds, in addition to common amino acids. The amino acid sequences were established by HMBC correlations from H-α or amide NH signals to the carbonyl of the connecting amino acid. The absolute configurations of the amino acid residues were determined by Marfey’s analysis while the absolute configuration of the stereogenic carbon (C-1α) of HMPA was established as S by an inspection of NOESY correlation. The absolute configuration of C-5α in N-Me-Thr in 357 was established as S by NOESY correlations and confirmed by a modified Mosher’s method [161].
In the case of 358, the structure and absolute configurations were confirmed by X-ray analysis. Thus, the structures of 357–362 were elucidated as cyclo-[(O)-HMPA1-L-Pro2-L-Phe3-NMe-L-Val4-NMe-L-Thr5-β-Ala6 (CO)], cyclo-[(O)-HMPA1-L-Pro2-L-Tyr3-NMe-Val4-NMe-Val5-β-Ala6 (CO)], cyclo-[(O)-(1αS)-HMPA1-L-Pro2-L-Phe3-NMe-L-Val4-(5αS)-NMe-Abu5-β-Ala6 (CO)], cyclo-[(O)-(1αS)-HMPA1-L-Pro2-L-Phe3-NMe-L-Ala4-(5αS)-NMe-Val5-β-Ala6 (CO)], cyclo-[(O)-(1αS)-HMPA1-L-Pro2-L-Phe3-NMe-L-Ile4-NMe-Val5-β-Ala6 (CO)], and cyclo-[(O)-(1αS)-HMPA1-L-Pro2-L-Phe3-L-Val4-L-Val5-β-Ala6 (CO)], respectively [161].
The mycelium extract of Clonostachys sp. ESNA-A009, isolated from an unidentified marine sponge collected in Japan, afforded IB-01212 (363), a cyclodepsipeptide featuring C2 symmetry (Figure 26). The symmetrical dimeric nature of 363 was based on the fact that its molecular formula has twice the number of the carbons and protons observed in the NMR spectra. An analysis of the COSY, gHSQC, and gHMBC spectra led to the identification of the natural amino acid Ser, and the unnatural N-Me-Leu, N,N-Me2-Leu, and N-Me-Phe, while gHMBC experiments were used to unequivocally establish the amino acid sequence and the site of dimerization. The stereochemistry of Ser, N-Me-Leu, and N-Me-Phe was determined by Marfey’s method, which confirmed the L configuration for all of them. The configuration of N,N-Me2-Leu was determined by derivatization of the hydrolyzate of 363 with 1-mental, and we compared its HPLC retention times with those of the derivatized standards. In this way, the configuration of N,N-Me2-Leu was established as L. Moreover, the structure of 363 was also confirmed by a comparison of its NMR data and HPLC retention times with those of the synthetic counterpart [162].
The undescribed cyclodecadepsipeptides, phaeosphamides A (364) and B (365), were isolated together with a previously reported congener Sch 217048 (366) (Figure 26) [163], from the extract of the solid rice culture of the fungus, Phaeosphaeriopsis sp. Esf-30, isolated from a rhizosphere sediment of a mangrove plant, Bruguiera gymnorhiza, which was collected from Techeng Isle, Zhanjiang, Guangdong Province, China [164].
An analysis of the 1D and 2D NMR spectra of 364 revealed the presence of nine proteinogenic amino acids, viz. Phe, Pro, Gly, N-Me Val, N-Me Gln, Ile, Pip, Val, and N-O-dimethylglutamic acid (N-O-DiMe Glu), in addition to 2-hydroxy-3-methylpentanoic acid (HMP). The amino acid sequence in 364 was established by HMBC and NOESY correlations as cyclo-(N-O-DiMe-Glu-Val-Pip-Ile-N-Me-Gln-N-Me-Val-Gly-Pro-Phe-HMP). Therefore, the structure of 364 differs from 366 only the presence of N-O-DiMe Glu in 364 instead of N-Me Glu in 366. The absolute stereochemistry of 364 was determined by a comparison of its 1H NMR spectra and optical rotation with those of the methylated 366. Since, the 1H NMR spectra and optical rotations of 364 and the methylated 366 are nearly identical, it was concluded that they are the same compound. Therefore, the complete structure of 364 was elucidated as cyclo-(N-O-DiMe L-Glu-L-Val-L-Pip-L-Ile-N-Me L-Gln-N-Me D-Val-Gly-L-Pro-L-Phe-(2R,3S)-HMP) [164].
An analysis of the 1D and 2D NMR spectra of 365 revealed the presence of N-Me-Glu instead of N-Me-Gln in 366. The amino acid sequence of 365 was identical to that of 366 on the basis of HMBC and NOESY correlations. The absolute configurations of the amino acids were determined by the advanced Marfey’s method. Therefore, the complete structure of 365 was established as cyclo-(N-Me L-Glu-L-Val-L-Pip-L-Ile-N-O-DiMe L-Glu-NMe D-Val-Gly-L-Pro-L-Phe-(2R,3S)-Hmp) [164].
4. Biological and Pharmacological Activities
Marine-derived peptides are another group of marine natural products that exhibit a vast array of biological and pharmacological activities. For practical purposes and to facilitate the readers, the activities are classified as follows:
4.1. Cytotoxic Activity
Compounds 1, 2 (Figure 3), 8 (Figure 4), and 29 (Figure 5), isolated from the culture broth extract of a deep-sea-derived fungus, S. obclavatum EIODSF 020, were assayed against human leukemia cell lines, HL-60 ((human acute promyelocytic leukemia) and K562 (chronic myelogenous leukemia cells). Compounds 1 and 29 exhibited weak cytotoxicity against HL-60 cells, with IC50 values of 100 and 64.7 μM, respectively, while 2 and 8 showed cytotoxicity toward K562 cells with IC50 values of 73.5 and 39.4 μM, respectively [38].
Compounds 17–19 (Figure 4), isolated from a mangrove endophytic fungus, Aspergillus terreus LM.5.2, showed weak cytotoxicity against human cancer cell lines, such as DLD-1 (colorectal adenocarcinoma), MCF-7 (breast adenocarcinoma), and PC-3 (prostatic adenocarcinoma), with IC50 values ranging from 58.3 to 96.8 μM, in the MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide] assay. Moreover, 17–19 exhibited cytotoxic activity toward rat non-malignant cardiomyocyte H9c2 cells, with IC50 higher than 76.7 μM [43].
Compounds 27 and 28 (Figure 4), isolated from a marine sediment-derived fungus, A. terreus FA009, were examined for their potential as anticancer agents or gene expression modulators by exploring their DNA-binding properties by fluorescence spectroscopy. The results showed that the fluorescence of 27 decreased upon adding double-stranded DNA oligomers. Based on the observed stoichiometry, 27 appeared to have a combined binding mode, i.e., intercalation by 27 along with groove binding by the peptide backbone [48].
Compounds 39, 40 (Figure 6) and 57 (Figure 10), isolated from the marine sponge- associated Acremonium sp., were tested for the in vitro anticancer activity. Compounds 39 and 40 mildly inhibited the growth of the murine L1210 (lymphocytic leukemia) cancer cell line in a disk diffusion soft agar colony-forming assay, with relative potencies of 4 and 1. Compound 57 showed potent cytotoxic activity against L1210 cell line with relative potency of 3300 and against HCT-116 (human colorectal carcinoma) cell line, with IC50 value of 3.5 ng/mL [52].
Compounds 41, 42 (Figure 6), and 57–61 (Figure 10), isolated from a marine sponge- associated fungus, Acremonium sp. 021172, were subject to the in vitro soft agar disk diffusion screening assay on various cancer cell lines, such as NCl-H125 (human non-small-cell lung carcinoma), HCT-116, CEM (human lymphocytic leukemia), C38 (murine colon adenocarcinoma), and CFU-GM (murine bone marrow). However, only 57, 58, and 61 exhibited similar cytotoxicity against the NCl-H125 cancer cell line, with similar IC50 values of 1.3 nM, suggesting that the methyl groups on R1, R3, and R4 have little impact on their cytotoxicity. A colonogenic study indicated that a therapeutic effect of 57 against NCl-H125 cells would require a chronic exposure (168 h) at a concentration of 2.0 ng/mL or higher, or a bolus exposure (24 h) at a concentration of 300 ng/mL or higher [53].
Compounds 62–66 (Figure 10), isolated from a marine sediment-derived fungus, Trichoderma sp. MMS1255, were evaluated for cytotoxic activity against KB (human epithelial carcinoma) cell line, using the MTT assay. All the tested compounds showed strong cytotoxicity, with IC50 values of 2.4, 4.3, 0.8, 0.8, and 0.7 μM, respectively. The positive control, alamethicin F50/5, showed an IC50 of 9 μM [58].
Compounds 67–73 (Figure 11), isolated from a marine sponge-associated fungus, Trichoderma sp. GXIMD 01001, displayed cytotoxicity against four human cancer cell lines, including A549 (lung adenocarcinoma), H1299 (non-small cell lung cancer), SW480 (colorectal cancer), and SW1990 (pancreatic adenocarcinoma), with IC50 values ranging from 0.46 to 4.7 μM. The positive control, cisplatin, showed IC50 values between 6.2 and 34 μM. Furthermore, the most potent compound, 72, showed antiproliferative activity against SW480 cells and deformation of the colony. Compound 72 was found to induce apoptosis in SW480 cancer cells [59].
Compounds 82 and 83 (Figure 13), isolated from the culture broth extract of a marine-derived fungus, Aspergillus sp., showed significant antiproliferative activity against a panel of cancer cell lines, viz. RKO (colon carcinoma), SNU638 (gastric carcinoma), SK-HEP-1 (hepatic adenocarcinoma), and MDA-MB (breast cancer), with IC50 (72h) values of 0.8, 4.8, 2.9, and 7.0 μM for 82, and 1.1, 8.0, 3.5, and 9.7 μM for 83, respectively. The positive control, etoposide, showed the IC50 values of 3.3, 0.3, 0.4, and 10.1 μM, respectively [62].
Compounds 84 and 85 (Figure 13), isolated from a marine fish-possessing-derived fungus, P. fellutanum Biourge, showed potent cytotoxicity against three cancer cell lines, i.e., murine leukemia P388 (IC50 values of 0.2 and 0.1 μg/mL, respectively), L1210 (IC50 values of 0.8 and 0.7 μg/mL, respectively), and KB (IC50 values of 0.5 and 0.7 μg/mL, respectively) [63].
Compound 93 (Figure 14), isolated from a neomycin-resistant mutant of a marine sediment-associated fungus, P. purpurogenum G59, selectively inhibited the growth of HeLa (human cervical carcinoma) cells with an inhibition rate of 39.4% at a concentration of 100 μg/mL, which was comparable with the positive control, 5-fluorouracil (41.4% inhibition at a concentration of 100 μg/mL) [68].
Compound 109 (Figure 14), isolated from a marine-derived fungus, Microsporum sp., displayed cytotoxic activity against HeLa cells. Furthermore, 109 induced apoptosis in HeLa cells through the downregulation of Bcl-2 expression, upregulation of Bax expression, and activation of the caspase-3 and p53 pathways [74].
Compound 117 (Figure 14), isolated from a marine-derived fungus, Aspergillus sp. SF-5044, exhibited weak cytotoxic activity against HL-60, MDA-MB-231 (triple-negative human breast cancer adenocarcinoma), Hep3B (hepatocellular carcinoma), 3Y1 (rat fibroblast), and K562, with IC50 values of 75, 130, 150, 180, and 250 μM, respectively [80].
Compounds 134–137 (Figure 14), isolated from a marine cyanobacterium-associated fungus, E. rostratum (Drechsler) CNK-630, exhibited in vitro cytotoxicity against HCT-116 cell line, with IC50 values of 8.5, 1.9, 0.76, and 16.5 μg/mL (9, 4.4, 1.6, and 35 μM), respectively. The authors suggested that the increased potency of 135 when compared to 136 might be due to the ketone functionality at C-5 (C-5′); however, the reduced potency in 137 was not in agreement with this simplistic analysis [85].
Compound 139 (Figure 14), isolated from a marine sediment-derived Penicillium sp. WF-06, was shown to inhibit the growth of HepG2 (hepatoblastoma) cell line, with an IC50 value of 19.6 μmol/L [86].
Compound (+)-148 (Figure 15), isolated from the marine-derived fungus, A. versicolor HBU-7, showed cytotoxicity against HGC-27 (human gastric carcinoma) cell line, with an IC50 value of 4.54 μM. The positive control, cisplatin, showed IC50 = 8.63 μM [90].
Compounds 156, 159, and 160 (Figure 15), isolated from a semi-mangrove endophytic fungus, E. rubrum G2, were assayed against seven human cancer cell lines, including MCF-7, SW1990, HepG2, NCl-H460 (non-small-cell lung carcinoma), SMMC-7721 (hepatocarcinoma), HeLa, and Du145 (prostatic carcinoma). Compound 156 selectively inhibited the growth of the SMMC-7721 cell line with IC50 = 30 μg/mL. Compound 159 showed cytotoxicity against HepG2, NCI-H460, and HeLa cell lines with IC50 values of 20, 22, and 20 μg/mL, respectively, while 160 was active against MCF-7, SW1990, SMMC-7721, and HeLa cells with IC50 = 20, 20, 20, and 30 μg/mL, respectively [91].
Compounds 169 and 170 (Figure 15), isolated from the tunicate-derived fungus, Aspergillus sp. DY001, were evaluated for cytotoxicity, by MTT assay, against MDA-MB-231, HCT-116, and HeLa. Compounds 169 and 170 showed cytotoxicity against MDA-MB-231 and HCT-116 cell lines, with IC50 values of 24.3 and 15.1 μM (for 169) and 26.3 and 16.2 μM (for 170), but were inactive against HeLa cells. The IC50 values of the positive control, 5-FU, were 13.0 and 4.6 μM, against MB-231 and HCT-116 cells, respectively [93].
Compounds 190 and 191 (Figure 16), isolated from a marine sediment-derived fungus, Eurotium sp. SCSIO F452, exhibited moderate cytotoxicity against the SF-268 (human glioblastoma) cell line, with IC50 values of 12.5 and 15.0 μM, respectively, and weak cytotoxicity against the HepG2 cancer cell line, with IC50 values of 30.1 and 37.3 μM, respectively. The positive control, taxol, showed IC50 values of 6.0 and 11.1 μM toward SF-268 and HepG2, respectively [100].
Compound 192 (Figure 17), isolated from a brown alga-derived fungus, Aspergillus sp. (BM-05 and BM-05ML), was assessed, by a microplate MTT assay, for the anti-proliferative activity against four human tumor cell lines, including K562, HCT-116, A2780 (ovarian cancer), and A2780CisR (cisplatin-resistant mutant in ovarian cancer). Compound 192 showed cytotoxicity against all the tested cell lines, with IC50 values of 67.8, 28.5, 27.3, and 49.4 μM, respectively. The positive control, cisplatin, showed IC50 values of 7.8, 33.4, 0.8, and 8.4 μM, respectively [81].
Compounds 92, 115 (Figure 14), 197, 201, and 210 (Figure 17), isolated from a sea water-derived fungus, A. ochraceopetaliformis DSW-2, showed weak cytotoxicity against HPAC (pancreatic ductal adenocarcinoma) and BxPC3 (pancreatic carcinoma) cell lines, with IC50 values higher than 20 μM [78].
Compound 221 (Figure 18), isolated from a marine gorgonian-associated A. terreus SCSGAF0162, was tested for its cytotoxicity against U937 (pro-monocytic, human myeloid leukaemia), K562, BGC823 (gastric carcinoma), MOLT-4 (acute lymphoblastic leukemia), MCF-7, and A549 cell lines by the MTT method. However, 221 only showed cytotoxicity against the U937 and MOLT-4 cell lines, with IC50 = 6.4 and 6.2 μM, respectively. Taxol was used as a positive control against the U937, K562, BGC-823, MOLT-4, Mcf7, and A549 cell lines, with IC50 values of 1.9, 4.9, 3.5, 1.8, 5.0, and 3.6 nM, respectively [108].
Compounds 223 and 224 (Figure 18), isolated from a marine bryozoan-associated fungus, Microsporum cf. gypseum, were assayed for in vitro cytotoxicity against human HCT-116 cancer cell line. Compound 223 (IC50 = of 0.6 μg/mL) exhibited better cytotoxicity than 224 (IC50 = of 8.5 μg/mL) toward HCT-116, suggesting the importance of the ketone carbonyl group in the side chain for biological activity. Compound 223 also showed cytotoxicity against a 60-cell line panel of National Cancer Institute (NCI), with the mean IC50 = 2.7 μg/mL. Etoposide and DMSO (solvent) were used as positive and negative controls, respectively [110].
Compound 227 (Figure 19), isolated from a marine-derived fungus, A. versicolor (MST-MF495), exhibited weak cytotoxicity toward some human tumor cell lines, including MM418c5 (malignant melanoma), DU145, and T47D (breast cancer cell line resistant to tamoxifen), with the median effective concentration (EC50) values of 66, 90, and 94 μg/mL, respectively [112].
Compound 245 (Figure 20), isolated from a marine gorgonian coral-derived fungus, A. versicolor (TA01-14), at a concentration of 10 μM, showed weak cytotoxicity toward human NCI-H292 (pulmonary carcinoma) and A-431 (skin squamous carcinoma) cell lines, by the MTT assay, with an inhibition ratio of 53.84% and 63.6%, respectively. The reference drug, adriamycin, at a concentration of 1 μM, showed inhibition of 93.4% and 91.0%, respectively [117].
Compound 256 (Figure 20), isolated from a marine-derived halotolerant fungus, A. sclerotiorum PT06-1, displayed weak growth inhibition of the HL-60 cell line, with an IC50 value of 56.1 μg/mL [119].
Compound 260 (Figure 20), isolated from a marine sponge-associated fungus, A. similanensis KUFA0013, showed weak cytotoxicity against MCF-7, NCl-H460, and A373 (melanoma), in the sulforhodamine B (SRB) assay, with GI50 values of 125, 117, and 115, respectively. Doxorubicin, a positive control, showed GI50 for MCF-7 = 42.8 nM; MDA-MB-231 = 10.86 nM; NCI-H460 = 94.0 nM; SF-268 = 94.0; and UACC-62 = 94.0 nM [121].
Compounds 276–278 (Figure 21), isolated from a marine sediment-derived A. persicinum SCSIO115, displayed cytotoxicity, in a SRB assay, against SF-268 (anaplastic gliblastoma), MCF-7, and NCI-H460, with IC50 values of 3.7, 3.0, and 11.6 μM (for 276), 45.6, 82.7, and >100 μM (for 277), and 3.2, 2.7, and 4.5 μM (for 278), respectively. The IC50 values of cisplatin, a positive control, were 4.6, 10.2, and 1.6 μM [126].
Compounds 287 and 288 (Figure 21), isolated from a marine alga-derived fungus, Scytalidium sp. CNC-310, exhibited moderate cytotoxic activity against HCT-116 cells, with IC50 = 2.7 and 11.0 μM, respectively. In the NCI 60 cell-line panel, both compounds showed the mean GI50 values of 7.9 and 4.1 μM, respectively. Interestingly, the most sensitive cell lines were MOLT-4 (IC50 = 3.0 μM for 287) and UACC-257 (melanoma) (IC50 = 1.2 μM for 288) [132].
Compound 311 (Figure 25), isolated from the culture extract of a mangrove endophytic fungus, A. terrus (no. GX7-3B), exhibited cytotoxicity against MCF-7, A549, HeLa, and KB, with IC50 values of 2.02, 0.82, 1.14, and 1.10 μM, respectively. The IC50 values of epirubicin, the positive control, were 1.07, 0.79, 0.42, and 0.05 μM, respectively [140]. Compound 311 also inhibited the growth of KB and KBv200 (multidrug-resistant KB) cell lines, with IC50 values of 5.76 and 5.34 μM, respectively. Further cell cycle studies revealed that the mitochondrial pathway, such as the reduction of reactive oxygen species (ROS) production, a decrease of mitochondrial membrane functionality, the release of cytochrome c, caspases-9 and -3 activation, and poly (ADP-ribose) polymerase (PARP) cleavage, are involved in the apoptosis induction. On the other hand, the regulation of either Bcl-2 or Bax was not involved in the apoptosis induction by 311 in the KB and KBv200 cell lines [141].
Compound 315 (Figure 25), isolated from a mycelia extract of a marine-derived Fusarium sp., showed the in vitro selective cytotoxicity against COLO 205 (colon cancer) and SK-MEL-2 (melanoma) cell lines, with IC50 values of 3.5 and 5.9 μg/mL, respectively. The positive control, mitomycin C, showed IC50 = 5.3 μg/mL against both cancer cell lines. Interestingly, 315 exhibited IC50 = 9.8 μg/mL against HCT-116 cell line, while the corresponding linear peptide was inactive, suggesting that the cyclization is vital for cytotoxic activity [145]. Compounds 315 and 316 (Figure 25) also exhibited moderate cytotoxicity, with mean GI50 values of 8.3 and 3.6 μM, respectively, in the NCI’s human tumor cell line screen [146].
Compound 318 (Figure 25), isolated from a marine cyanobacterium-associated fungus, Z. masonii (CNK458), showed the median GI50 value of 9.1 μM in the cytotoxicity assay against 60 cancer cell lines in the NCI. Interestingly, 318 exhibited a selective cytotoxic activity toward SF-268 and RXF 393 (renal cancer) cell lines, with GI50 values of 6.5 and 5.0 nM, respectively [148].
Compounds 320–325 (Figure 25), isolated from a marine sponge-associated fungus, Metarrhizium sp., were tested against H125, HCT-116, and CEM, as well as C38 (mouse colon carcinoma) and mouse CFU-GM (colony-forming unit for granulocytes and macrophages), using a soft agar disk diffusion method. Compounds 324 and 325 selectively inhibited the murine C38 cells at the lowest concentration of 0.136 and 0.150 nM, respectively, but no inhibition of CFU-GM cells was observed. Compounds 320, 322–325 inhibited all the solid tumors, with 324 having the best inhibitory activity at a concentration of 34 μM. Compound 324 showed an IC50 value of 160 nM against HCT-116 cells. The colonogenic study of 324 indicated that only a chronic exposure (168 h) of 190 ng/mL or higher would display the therapeutic effect toward HCT-116 cells. The maximum tolerated dose in mice (MTD) for 324 was established as 0.125 mg/mouse (6.25 mg/kg) [53].
Compounds 331 and 332 (Figure 25), isolated from a marine sponge-associated fungus, S. brevicaulis NCPF 2177, showed significant inhibition on growth of Colo357 (metastatic pancreatic adenocarcinoma), Panc89 (pancreatic ductal adenocarcinoma), and HT29 (colorectal adenocarcinoma) cell lines at a final concentration of 10 μg/mL. The cell viability of Colo357, Panc89, and HT29 was decreased by 36%, 42%, and 37% (for 331) and 26%, 49%, and 24% (for 332), respectively [151].
Compounds 331 and 332 (Figure 25), isolated from a gorgonian coral-derived fungus, P. chrysogenum (CHSCLM-0003), were also examined for their cytotoxicity against MCF-7 by the MTT method, and BEL-7402 (human hepatocellular carcinoma), HepG2, and SMMC7721 (human hepatocellular carcinoma) by the SRB method. Compounds 331 and 332 exhibited strong and selective cytotoxic activity against HepG2 and SMMC7721, with IC50 values of 26.81 and 33.75 μM, respectively. The IC50 values of the positive controls, 5-FU, were >100 and 15.79 μM, and of cisplatin were 28.27 and 21.81 μM [159].
Compounds 353 and 354 (Figure 26), isolated from a co-culture of the algicolous fungus, Emericella sp. (strain CNL-878), and a marine actinomycete, S. arenicola (strain CNH-665), displayed weak cytotoxicity against HCT-116, with IC50 values of 23 and 40 μM, respectively [160].
Compound 355 (Figure 26), isolated from a soft coral-associated fungus, A. sclerotiorum SCSIO 41031, showed cytotoxicity against human nasopharyngeal carcinoma cell lines, HONE1 and HONE1-EBV, with IC50 values of 13.0 and 10.1 μM, respectively [120].
Compound 356 (Figure 26), isolated from a marine sponge-associated fungus, Clonostachys sp. ESNA-A009, was assayed against a panel of 14 human tumor cell lines. However, 356 showed the most potent cytotoxicity toward LN-caP (prostate cancer), SK-BR3 (breast cancer), HT29, and HeLa cell lines, with the same growth inhibition (GI50) of 10−8 M [162].
Compound 364 (Figure 26), isolated from the sediment-derived fungus, Phaeosphaeriopsis sp. Esf-30, was evaluated, by the MTT method, against five cancer cell lines, viz. AGS (human gastric adenocarcinoma), BEL-7402 (human hepatocarcinoma), HepG2, BIU87 (human bladder cancer), and B16 (mouse melanoma). Compound 364 exhibited selective cytotoxicity against AGS cells at 20 μM, which was equivalent to the positive control, docetaxel, at 10 μM. Furthermore, the IC50 value of 364 against the proliferation of AGS cells was 5.14 μM. Flow cytometry analysis showed that 364 treatment caused AGS cell arrest in the G2 phase in a dose-dependent manner, leading to the increase in apoptotic induction at the concentration ranging from 2.5 to 10 μM [164].
4.2. Antibacterial and Antibiofilm Activities
Most of marine organisms biosynthesize a unique and distinct class of peptides with antibacterial and antimicrobial properties against a broad spectrum of Gram-positive and Gram-negative bacteria, which are known as antimicrobial peptides (AMPs). However, these peptides are also able to inhibit the detrimental fungi, viruses, and parasites with the lowest possibility of developing resistance [165]. The majority of AMPs are produced ribosomally and then undergo post-translational modification, whereas some of the AMPs are biosynthesized by a multiple-enzyme system [9].
The antibacterial activity of 39, 40 (Figure 6), and 57 (Figure 10), isolated from the marine sponge-associated Acremonium sp., were assayed against Staphylococcus epidermis. Compounds 39 and 57 were active and exhibited minimum inhibition concentration (MIC) values ranging from 25–80 μg/mL, while 40 showed no antibacterial activity. The positive control, vancomycin, showed MIC = 0.63 μg/mL [52].
Compounds 49 (Figure 8) and 55 (Figure 9), isolated from a culture extract of the marine tunicate-associated Talaromyces sp. (CMB-TU011), showed strong antibacterial activity against a Gram-positive bacterium, Bacillus subtilis ATCC 6633, with IC50 = 1.5 and 3.7 μM, respectively. Rifampicin was used as a positive control (40 mg/mL in 10% DMSO) [55].
Compounds 53 and 54 (Figure 8), isolated from the marine microalgal-derived fungus, Tolypocladium sp., were evaluated against ten Gram-negative, seven Gram-positive, and two acid-fast bacterial strains, by a broth microdilution antibacterial assay. Both compounds selectively showed weak to moderate antibacterial activity against six Gram-positive bacteria, including B. subtilis, methicillin-susceptible S. aureus (MSSA), methicillin-resistant S. aureus (MRSA), Listeria ivanovii, Enterococcus faecium, and E. faecalis (MIC values ranging from 4 to 32 μM), and against two acid-fast bacteria, i.e., Mycobacterium tuberculosis and M. smegmatis (MIC values ranging from 20 to 80 μM) [57].
Compounds 62, 64, and 66 (Figure 10), isolated from a marine sediment-derived fungus, Trichoderma sp. MMS1255, exhibited the same antibacterial activity against S. aureus (CIP53.156), with MIC value of 25 μg/mL. Gentamycin (100% inhibition of S. aureus at 40 μg/mL in 1% MeOH) was used as a positive control [58].
Compounds 67–73 (Figure 11), obtained from a sponge-associated fungus, Trichoderma sp. GXIMD 01001, exhibited weak and moderate antibacterial activity against S. aureus, with MIC values between 23 and 45 μg/mL. The positive control, chloromycetin, showed MIC = 3.1 μg/mL [59].
Compound 86 (Figure 14), isolated from a marine sponge-associated fungus, Penicillium sp. F37, inhibited a biofilm formation of the pathogenic S. epidermidis ATCC 35984 in a range of 0.25, 0.5, and 1.0 mg/mL (60%, 65%, and 85%, respectively) without interference on bacterial growth, whereas, at 2.0 mg/mL, it was capable of inhibiting bacterial growth, and, at 0.125 mg/mL, no significant inhibition of a biofilm formation was observed [64].
Compound 92 (Figure 14), isolated from a marine sediment-derived fungus, P. chrysogenum DXY-1, at a sub-minimum inhibitory concentration, decreased the production of quorum sensing (QS)-regulated violacein of Chromobacterium violaceum by 79%, and the production of QS-mediated pyocyanin, proteases, and elastase activities of P. aeruginosa PA01 by 41%, 20%, and 32%, respectively. Moreover, 92 also destroyed the biofilm formation and decreased the QS gene expression of P. aeruginosa PA01. A molecular docking study showed that 92 blocked the effects of QS autoinducers via competitive binding to the same pocket of the receptor proteins [67].
Compounds 108 and 110 (Figure 14), isolated from a marine sponge-associated E. chevalieri MUT2316, were assayed for antibacterial activity against marine bacteria. Compound 108 showed inhibitory activity against Halomonas aquamarina ATCC 14400 and Vibrio aesturianus ATCC 35048, with the same low observable effect concentration (LOEC) value of 0.001 μg/mL, while 110 inhibited the growth of Pseudoalteromonas citera ATCC 29720 and Polaribacter irgensii ATCC 700398, with LOEC values of 0.01 and 1 μg/mL, respectively. Compound 110 also inhibited the adhesion of only two bacterial strains, P. citera and Roseobacter littoralis ATCC 49566, with LOEC values of 0.001 and 0.1 μg/mL, respectively [73].
Compound 145 (Figure 15), isolated from a marine sediment-derived fungus, A. versicolor (MF030), displayed significant antibacterial activity against Bacille Calmette-Guerin (BCG) with an MIC value of 6.25 μg/mL, which is modest when compared to the positive control, isoniazid (MIC value of 0.05 μg/mL), but no significant antibacterial activity against the Gram-positive bacteria S. aureus ATCC 6538 and B. subtilis ATCC 6633, the Gram-negative bacteria P. aeruginosa (PAO1) and Escherichia coli (ATCC 25922), or yeast, Candida albicans (SC 5314) [89].
Compound 226 (Figure 19), isolated from a marine gorgonian-associated fungus, Aspergillus sp. XS-20090B15, showed antibacterial activity toward B. cereus and S. epidermis with the same MIC value of 12.5 μM. Ciprofloxacin was used as a positive control [46].
Compounds 129 (Figure 14) and 229 (Figure 19), isolated from an unidentified decaying green alga-derived fungus, Asteromyces cruciatus 763, were assayed for their antibacterial activity. Compound 229 showed weak antibacterial activity against B. subtilis and S. epidermidis with growth inhibition of 61% and 30%, at a concentration of 100 μM. Compound 129, at a concentration of 50 μg level, exhibited total inhibition of 2.5 and 7 mm against E. coli and B. megaterium in the agar diffusion assay [83].
Compounds 169 and 170 (Figure 15), isolated from the tunicate-associated fungus, Aspergillus sp. DY001, was evaluated for antibacterial activity by a disc diffusion assay, at a concentration of 50 μg/disc, against two pathogens, including E. coli ATCC 25922 and S. aureus ATCC 25923. Both compounds moderately inhibited the bacterial strains with inhibition zones ranging from 16 to 22 mm, and MIC values ranging from 4 to 8 μM. The positive control, ciprofloxacin, showed inhibition zones of 30 and 22 mm, respectively [93].
Compounds 175 and 178–182 (Figure 16), isolated from a deep-sea cold seep-derived fungus, A. chevalieri CS-122, were evaluated against two human pathogenic bacteria, E. coli and Micrococcus luteus, and three aquatic pathogenic bacteria, Vibrio harveyi, Edwardsiella tarda, and Aeromonas hydrophilia, by a serial dilution assay using 96-well microtiter plates. Compounds 179 and 182 moderately inhibited all pathogens, with MIC values of 16–32 μg/mL, while 178, 180, and 181 selectively showed antibacterial activity against E. coli and V. harveyi, with MIC values ranging from 4 to 32 μg/mL. Compound 175 inhibited the growth of A. hydrophilia and E. coli with MIC values of 4 and 8 μg/mL, respectively. Chloramphenicol was used as a positive control, showing MIC values ranging from 2 to 8 μg/mL. Given the significant antibacterial activity of 175 against A. hydrophilia (MIC = 4 μg/mL), a further antibacterial mechanism was assessed using scanning electron microscope (SEM) before and after treatment with 175. In addition to obvious cell deformation and serious damage to the cell membrane, 175 also caused deep grooves and clear pores on the bacterial surface leading to sufficient bacteriolysis and death of A. hydrophilia [96].
Compound 256 (Figure 20), isolated from a marine-derived halotolerant fungus, A. sclerotiorum PT06-1, selectively inhibited the growth of P. aeruginosa with an MIC value of 35.3 μM [119].
Compounds 269–271 (Figure 21), isolated from the coral-associated A. versicolor (CHNSCLM-0063), displayed antibacterial activity against Mycobacterium marinum, with MIC values of 23.4, 81.2, and 87.5 μM, respectively. The positive controls, rifampin, streptomycin, and isoniazid, showed MIC values of 19.0, 20.1, and 88.5 μM, respectively. In a further assay against M. tuberculosis H37Rv, only 270 showed a weak activity, with MIC = 100 μM. Compared to 270, it was suggested that the hydroxyl group at the R1 position of 269 considerably reduces the anti-M. tuberculosis activity [124].
Compounds 272 and 273 (Figure 21), also isolated from the marine gorgonian coral-derived A. versicolor (CHNSCLM-0063), showed anti-M. tuberculosis H37Rv activity, with MIC values of 50 and 100 μM, respectively. The MIC values of the positive controls, rifampin and isoniazid, were 0.00625 and 0.0125 μM, respectively [125].
Compounds 284 and 285 (Figure 21), isolated from a marine-derived fungus, E. unguis, showed moderate antibacterial activity against, S. aureus [129]. Compound 284, also inhibited the aquatic pathogen, Vibrio parahemolyticus, with an inhibition zone of 9.0 mm, which was the same as the positive control, ampicillin [130].
Compounds 312 and 313 (Figure 25), isolated from a sponge-associated fungal strain No. 001314c, exhibited weak antibacterial activity against S. epidermis ATTC 12228 and Enterococcus durans ATTC 11576, in a micro broth dilution assay, with the same MIC value of 100 μg/mL [144].
Compound 319 (Figure 25), isolated from a marine sediment-derived fungus, Altenaria sp. SF-5016, showed antibacterial activity against B. subtilis KCTC1021 and S. aureus KCTC1928, with inhibition zones of 8 and 13 mm, respectively. The positive control, gentamicin, showed inhibition zones of 16 and 13 mm, respectively [149].
Compounds 338–340, and 343 (Figure 26), isolated form a marine bryozoan-associated fungus, B. feline EN-135, showed antibacterial activity, in the disk diffusion and double-dilution methods, against E. coli with MIC values of 64, 64, 8, and 16 μg/mL, respectively. The MIC value of chloramphenicol, the positive control, was 4 μg/mL [157].
4.3. Antifungal Activity
Compounds 20 and 21 (Figure 4), isolated from a marine-derived fungus, T. purpureogenus CX11, were assayed for their antifungal activity against phytopathogenic fungus, Fusarium oxysporum ATCC 28856, and human pathogenic fungi Candida albicans SC5314 and Cryptococcus neoformans GOYJ11988. The MIC values against F. oxysporum ranged from 12.5 to 25 μg/mL, and against the two remaining fungi ranged from 40 to 80 μg/mL [44].
Compound 129 (Figure 14), isolated from an unidentified decaying green alga-derived fungus, A. cruciatus 763, showed antifungal activity at a concentration of 50 μg, against Mycotypha microspora, Euritium rubrum, and Microbotryum violaceum, with total inhibition zones of 13.5, 4, and 13 mm, in the agar diffusion assay. The positive control, miconazole exhibited total inhibition zones of 6–10 mm [83].
Compounds 169 and 170 (Figure 15), isolated from a tunicate-associated fungus, Aspergillus sp. DY001, was evaluated for antifungal activity against C. albicans ATCC 14053, using a disc diffusion assay at a concentration of 50 μg/disc. Both compounds weakly inhibited the growth of C. albicans, with inhibition zones of 11 and 12 mm, and exhibited the same MIC values of 16 μM. The inhibition zone and MIC value of ketoconazole, the reference drug, were 30 mm and 0.26 μM [93].
Compounds 196, 197, 201, 204, and 207 (Figure 17), isolated from a marine halotolerant fungus, A. sclerotiorum PT06-1, selectively inhibited the growth of C. albicans, in an agar dilution method, with MIC values of 7.5, 3.8, 30, 6.7, and 30 μM, respectively [102].
Compound 219 (Figure 18), isolated from a co-culture of mangrove endophytic fungi, Phomopsis sp. K38 and Altenaria sp. E55, displayed antifungal activity against four crop-threatening fungi, viz. Gaeumannomyces graminis, Rhizoctonia cerealis, Helminthosporium sativum, and Fusarium graminearum, with MIC values of 220, 160, 130, and 250 μg/mL, respectively. The positive control, tridimefon, showed MIC values of 150, 100, 120, and 150 μg/mL, respectively [105].
Compounds 246, 250, and 251 (Figure 20), isolated from a deep-sea-derived fungus, S. obclavatum EIODSF 020, exhibited significant antifungal activity against A. versicolor and Curvularia australiensis, with MIC values of 0.625, and 0.156 μM for 246, 0.625, and 0.156 μM for 250, and 1.562, and 0.156 μM for 251, respectively. The MIC values of the positive controls, ketoconazole and amphotericin, were 0.625, 0.156 μM against A. versicolor, and 1.562 and 0.156 μM against C. australiensis, respectively [49].
Compounds 246, 250–253 (Figure 20), isolated from a marine sediment-derived fungus, S. obclavatum EIODSF 020, showed antifungal activity against phytopahogenic fungi, Altenaria solani and Colletotricum asianum, with MIC values ranging from 0.195 to 6.250 μg/disc, and mild antifungal activity against Pyricullaria oryzae. The positive controls, amphotericin B and ketoconazole, showed MIC values of 6.250, 0.195, 25 μg/disc, and 12.5, 25 and 100 μg/disc, respectively, against A. solani, C. asianum and P. oryzae [118].
Compounds 255 and 256 (Figure 20), isolated from a marine-derived halotolerant fungus, A. sclerotiorum PT06-1, exhibited moderate antifungal activity, in an agar dilution method, toward C. albicans with MIC values of 7.0 and 3.5 μM, respectively [119].
Compounds 289–293 (Figure 21), isolated from a marine gorgonian-associated fungus, Aspergillus sp. SCSIO 41501, showed antifungal activiy against phytopathogenic fungal strains, Fusarium oxysporum, C. australiensis, P. oryzae, A. solani, and Colletotrichum gloeosporioiles, with MIC values ranging from 3.12 to 50 μg/disc. The MIC of the reference drug, actidione, ranged between 6.25 and 12.5 μg/disc. The difference in the β-amino fatty acid side chain can effectively influence the antifungal activity of these cyclic lipopeptides [50].
Compounds 358 and 360 (Figure 26), isolated from a marine-derived fungus, B. feline SYSU-MS7908, showed significant inhibition of the mycelial growth of Geotrichum citri-aurantii, with EC50 values of 56.8, and ca. 90.0 μg/mL, in the agar dilution method. The positive control, triadimefon, showed EC50 values of 146.4 μg/mL. In order to understand the effect of 358, the morphology and membrane permeability of G. citri-aurantii caused by 358, at a concentration of 60 μg/mL, were examined by SEM which showed a morphology change through the spare growth, collapse, deformation, peeling, and shrinkage of the mycelia of G. citri-aurantii. Additionally, treatment with a 60 μg/mL concentration disturbed its cell membrane integrity [161].
4.4. Antiviral Activity
Compounds 74–78 (Figure 12), isolated from a marine-derived fungus, Scytalidium sp., were assayed against herpes simplex virus type 1 and type 2 (HSV-1 and HSV-2). Compounds 74–78 were added to Vero cells infected with HSV-1 for one hour and the virus-induced cytopathic effects were measured for 5 days. Compounds 74–78 showed ED50 values of 1.1, 3.5, 2.2, 2.0, and 3.1 μM, respectively. The experiments showed that the antiviral efficacy of 74 is greater when it is incubated with HSV-1 prior to the exposure to Vero cells. In a standard plaque reduction assay, 74 also inhibited the replication of both HSV-1 and HSV-2 with the same ED50 values of 280 nM [60].
Compound 221 (Figure 18), isolated from a marine gorgonian-derived A. terreus SCSGAF0162, showed antiviral activity against the influenza virus strains A/WSN/33 (H1N1) and A/Hong Kong/8/68 (H3N2), with IC50 values of 15 and 8.1 μM, respectively. RIBA was used as a positive control against H1N1 and H3N2, showing IC50 values of 20.2 and 0.41 μM, respectively [108].
Compounds 10 (Figure 4) and 235 (Figure 19), isolated from a gorgonian-derived fungus, Aspergillus sp. SCSIO 41501, were assayed against HSV-1 using a plaque reduction assay. Compounds 10 and 235 exhibited antiviral activity, under their non-cytotoxic concentration (TC0), toward a Vero cells, with IC50 = 19.8 and 9.5 μM, respectively. The TC0 and TC50 values against Vero cells were 153.2 and 346.0 μM (for 10), and 81.9 and 204.4 μM (for 235). The positive control, acyclovir, showed the IC50 and TC0 values of 3.0 and >1000 μM, respectively. Interestingly, 235 displayed, at a concentration of 12.5 μM, antiviral activity against acyclovir-resistant clinical isolates of HSV-1-106 and HSV-1-153, with about a 50% inhibition rate [41].
Compounds 246, 250, and 251 (Figure 20), isolated from a deep-sea-derived fungus, S. obclavatum EIODSF 020, showed, under their non-cytotoxic concentration (TC0) toward a Vero cell, antiviral activity against HSV-1 in a plaque reduction assay, with IC50 values of 14.0, 16.7, and 15.6 μM, respectively. The TC0 values were 25.1, 57.2, and 49.4 μM, respectively. The IC50 and TC0 values of the positive control, acyclovir, were 3.0 and >1000 μM, respectively [49].
Compounds 261–265 (Figure 20), isolated from the marine sediment-derived fungus, A. persicinum SCSIO115, showed antiviral activity against HSV-1, with EC50 values of 16, 8.7, 27, 24, and 14 μM, respectively. The EC50 value of the positive control, ganciclovir, was 0.025 μM. Compounds 261, 262, and 265 act through an inhibition of UL42 gene transcription in HSV, which is an essential polypeptide for viral DNA replication [122].
4.5. Antiparasite Activity
Compound 356 (Figure 26), isolated from a marine sponge-associated Clonostachys sp., showed leishmanicidal activity at a low micromolar range of concentrations on promastigote and amastigote forms of the parasite. The mechanism of action of 356 to kill Leishmania is through an apoptosis-like process which involves intracellular targets [166].
4.6. Anti-Dinoflagellate Activity
Dinoflagellates are the most important contributors to the harmful algal bloom (red tide). Compounds 295 and 296 (Figure 20), isolated from a sponge-associated fungus, Clonostachys rogersoniana (HJK9), caused an inhibition of the motility of a dinoplagellate, Prorocentrum micans, at a concentration of 30 μM [133].
4.7. Algicidal Activity
The algicidal activity of 108–110 (Figure 14), isolated from a marine sponge-associated fungus, E. chevalieri MUT2316, was investigated against marine microalgae including Cylindrotheca closterium (AC170), Exanthemachrysis gayraliae (AC15), Halamphora coffeaeformis (AC713), Phaeodactylum tricornutum (AC171), and Prophyridium purpureum (AC122). Interestingly, 108 inhibited the growth and adhesion of all microalgae with LOEC values ranging from 0.001 to 100 μg/mL. Compound 109 inhibited the growth of C. closterium, H. coffeaeformis, and P. tricornutum, with LOEC values of 0.001, 0.01, and 1 μg/mL, respectively, and inhibited the adhesion of C. closterium, E. gayraliae, H. coffeaeformis, and P. tricornutum, with LOEC values of 1, 0.01, 1, and 0.001 μg/mL, respectively. Compound 110 only showed inhibitory activity against the growth of H. coffeaeformis and P. purpureum, with LOEC values of 0.1 and 0.001 μg/mL, respectively. The LOEC values of 110 against the adhesion of E. gayraliae and P. tricornutum were the same, i.e., 0.01 μg/mL [73].
Compound 234 (Figure 19), isolated from a marine sediment-derived fungus, Aspergillus sp. SCSIOW2, exhibited algicidal activity against two strains of noxious red tide algae, Akashiwo sanguinea and Chattonella marina. Compound 234 showed high 2 h inhibition rates of 58% and 36%, at a concentration of 50 μM, respectively. The authors found that 234 inhibited the growth of examined algae cells through a significant increase in ROS level, which was caused by the damage of superoxide dismutase (SOD) activity, and, consequently, generated high malondialdehyde (MDA) levels [114].
4.8. Enzyme Inhibitory Activity
4.8.1. Inhibition of Indoleamine 2,3-Dioxygenase (IDO) Activity
Indoleamine 2,3-dioxygenase (IDO) catalyzes the cleavage of the 2,3-indole bond of L-Trp, converting it to N-formylkynurenine in the first and rate-limiting step of the kynurenine catabolic pathway. IDO-expressing cancer cells use this transformation to reduce Trp concentrations in their microenvironments to levels that prevent T-lymphocyte activation and proliferation, thus rendering the ability of tumors to escape the T-lymphocyte-based immune response. A growing body of clinical data shows that many primary tumor cell lines obtained from patients overexpress IDO. Therefore, IDO has emerged as a promising molecular target for the development of a new class of therapeutic agents for treating cancer that work by modulating an extrinsic property of tumor cells [85].
Compounds 184–186 (Figure 16), isolated from a marine sediment-derived fungus, P. cucumerina, inhibited the purified, recombinant human IDO activity in vitro with IC50 values of ca. 2 μM. It was suggested that some portion of the phenoxazinone fragment C represents a new IDO inhibitory pharmacophore [98].
4.8.2. Inhibition of Acethylcholinesterase (AChE) Activity
Compound 311 (Figure 25), isolated from a mangrove endophytic fungus, A. terreus (no. GX7-3B), showed anti-AChE activity with an IC50 value of 3.09 μM. The positive control, huperazine A, had IC50 = 0.003 μM [140].
Compound 355 (Figure 26), isolated from a soft coral-associated fungus, A. sclerotiorum SCSIO41031, inhibited the activity of AChE with IC50 = 15.6 μM. A molecular docking study in the 2D model showed that the alkyl chain of 355 formed a hydrophobic interaction with the active site residues such as Trp84, Asp72, Try70, and Trp279. Moreover, the NH proton of Gly formed a hydrogen bond with the active site residue Tyr334 [120].
4.8.3. Inhibition of Histone Deacethylase Activity
Histone deacetylases (HDACs) comprise a family of enzymes that catalyze the removal of acetyl groups from lysine residues of histones and mediate chromatin remodeling and gene expression. Due to the capacity of HDAC inhibitors to suppress cell proliferation in a variety of transformed cells in culture and in tumor-bearing animals, they have great promise as new cancer drugs [167].
Compound 223 (Figure 18), isolated from a marine bryozoan-associated fungus, Microsporum cf. gypseum, showed stronger inhibitory activity against a mixture of both histone deacethylases, (HDACs) and HDAC8, than suberoylaniline hydroxamic acid (SAHA), a known HDAC inhibitor antitumor agent, with IC50 values of 0.14 and 0.55 μM (for 223) and 0.3 and 0.78 μM (for SAHA), respectively [110].
4.8.4. Inhibition of Sortase A (SrtA) and Isocitrate Lyase (ICL) Activities
Sortase A (SrtA) is an important enzyme found in the membrane of S. aureus, which is directly related to the virulence of bacteria. By inhibiting SrtA activity, promising antibacterial agents can be expected. Compounds 18 and 19 (Figure 4), isolated from a mangrove endophytic fungus, Aspergillus terreus LM.5.2, at a concentration of 80 μM, inhibited Srt A activity by more than 20%, while 17 (Figure 4) was void of activity [43].
Compounds 38 (Figure 5), 230, 231 (Figure 19), and 252 (Figure 26), isolated from marine-derived fungi, A. allahabadii and A. ochraceopetaliformis, were assayed against the key anti-virulence targets in pathogenic micro-organisms such as SrtA and isocitrate lyase (ICL). Compounds 38, 230, 231, and 252 exhibited moderate inhibition of S. aureus SrtA with IC50 values of 77.0, 70.0, 53.1, and 131.9 μM, respectively. The positive controls, berberine chloride and curcumin, showed the IC50 values 104.3 and 47.8 μM, respectively. Compound 231 was further assayed for C. albicans ICL and showed an inhibition of ICL with an IC50 = 116.8 μM. The 3-Nitropropanoic acid, the positive control for ICL, showed IC50- = 18.5 μM [51].
4.8.5. Inhibition of Pancreatic Lipase (PL) Activity
The enzyme pancreatic lipase (PL) hydrolyzes triglyceride so that it could be absorbed by human intestines. Therefore, PL inhibitors can prevent hyperlipidemia and obesity by decreasing the absorption of triglycerides [168].
Compounds 79–81 (Figure 13), isolated from a marine sponge-associated fungus, A. flavipes 164013, were found to potently inhibit PL activity, with IC50 values of 0.23, 0.07, and 0.14 μM, respectively. The positive control, kaempferol, showed IC50 = 1.5 μM. The position of chlorine was important in the bioactivity since 80 (2-chlorinated) was more active than 81 (4-chlorinated) and 79 (2,4-dichlorinated) [61].
Compounds 266–268 (Figure 20), isolated from a marine sponge-associated fungus, Aspergillus sp. 151304, showed a significant and dose-dependent inhibition of PL activity, with IC50 values of 7.6, 1.8, and 0.5 μM, respectively. Although kinetic studies revealed noncompetitive inhibition of PL by 268, the molecular dynamics simulation showed that 268 could bind PL at the entrance of the catalytic pocket. The corresponding positive controls, isoginkhetin and orlistat, had the IC50 values of 2.90 μM and 0.75 nM, respectively [123].
4.8.6. Inhibition of Tankyrase1/2 Activity
Tankyrases are proteins with poly (ADP-ribose) polymerase activity. Human tankyrases post-translationally modify multiple proteins involved in processes including maintenance of telomere length, sister telomere association, and trafficking of glut4-containing vesicles. The tankyrase protein family consists of tankyrases 1 and 2, which share 85% amino acid sequence identity [169].
4.8.7. Inhibition of Protein Tyrosine Phosphatase 1B (PTP1B) Activity
Protein tyrosine phosphatase 1B (PTP1B) has been shown to be involved in the negative regulation of both insulin and leptin action and was suggested to play an important role in insulin signalling and possibly in obesity in humans [170].
4.8.8. Inhibition of Cyclin-Dependent Kinase 4 (CDK-4) Activity
4.9. Anti-Fouling Activity
Compounds 1, 2 (Figure 3), 6–9 (Figure 4), 29, and 30 (Figure 5), isolated from the deep-sea-derived fungus, S. obclavatum EIODSF 020, were assayed against a settlement of Bugula neritina larvae. Compound 7 was found to show strongest activity, inhibiting the B. neritina larvae settlement with 77.8%, 69.9%, 68.1%, 56.5%, and 25.4% at concentrations of 25.0, 12.5, 6.25, 3.125, and 1.562 μg/mL, respectively (EC50 = 7.8 μg/mL and LC50/EC50 > 100), indicating that 7 is a potential antifouling candidate. Compounds 1, 2, 6, 8, 9, 29, and 30 displayed, at concentration of 25 μg/mL, 48.7%, 42.3% 40.2%, 12.5%, 23.6%, 19.9%, and 20.1% inhibition against B. neritina larvae settlement. Wells containing only filtered sea water with DMSO served as the controls [38].
4.10. Lipid-Lowering Activity
Compound 194 (Figure 17), isolated from a marine mud-derived fungus, A. versicolor ZLN-60, exhibited a strong lipid-lowering effects in HepG2 cells at a dose of 10 μM in the oil-red O staining assay [101].
Compound 302 (Figure 25), isolated from a mangrove endophytic fungus, S. kiliense HDN11-112, significantly reduced oleic acid (OA)-elicited lipid accumulation, as determined from the oil-red O staining, as well as intracellular cholesterol and triglyceride. Compound 302 did not show any cytotoxicity on the tested HepG2 cells at 20 μM, indicating that the lipid-lowering activity was not due to cytotoxicity [136].
4.11. Anti-Angiogenesis Activity
Compounds 344, 345, and 351 (Figure 26), isolated from a gorgonian coral-associated fungus, P. chrysogenum (CHSCLM-0003), were assayed for their pro-angiogenic activity using Tg(kdrl:EGFP) transgenic zebrafish line. All the tested compounds induced angiogenesis at concentration of 1.0 μg/mL against zebrafish embryo by promoting the cavity of blood vessels. These compounds were also non-toxic in embryonic zebrafish at a concentration of 100 μg/mL [159].
4.12. Anti-Inflammatory Activity
Compound 116 (Figure 14), isolated from a deep-sea sediment-derived fungus, Aspergillus sp. SCSIOW2, exhibited an inhibitory activity against a nitric oxide (NO) production, with IC50 = 40.3 μg/mL (68.0 μM), in a lipopolysaccharide (LPS) and recombinant mouse interferon-γ (INF-γ)-activated macrophage-like cell line, RAW 264.7. Compound 116 did not show any cytotoxic effect at the tested dose range (30–100 mg/mL). Quercetin was used as a positive control which showed stronger NO inhibitory effect (inhibitory rate 97.4% at 100 mg/mL) [79].
Compound 140 (Figure 14), isolated from a marine sediment-derived fungus, A. insulicola, showed anti-inflammatory activity through inhibition of NF-κB luciferase and nitrite production, with IC50 values of 8.37 and 13.70 μM, respectively. The corresponding positive control, celastrol, showed an IC50 value of 0.3 μM [87].
Compounds 164–166 (Figure 15), isolated from a marine sponge-associated fungus, A. sclerotiorum ST0501, were assayed for their capacity to inhibit NO production induced by LPS. Compounds 164 and 165 inhibited the NO production better than dexamethasone, the reference drug, with inhibition percentages of 28.92%, 26.31%, and 25.87%, respectively. However, 166 showed a weak inhibition rate at 21.08% [92].
Compounds 109, 110 (Figure 14), 160–162 (Figure 15), and 171–176, 179 (Figure 16), obtained from a deep-sea sediment-derived fungus, Aspergillus sp. FS445, were assayed for the inhibition of NO production in LPS-induced mouse macrophage RAW264.7 cells. Compound 175 showed the most significant inhibition of NO production, followed by 173, 162, 161, 176, and 172, with IC50 values of 16, 20, 25, 37, 57, and 89 μM, respectively. The positive control, aminoguanidine, showed the IC50 value of 23 μM [95].
Compounds 188 (Figure 16), 209 (Figure 17), and 216 (Figure 18), isolated from a marine sponge-associated fungus, A. violaceofuscus, were investigated for their inhibitory activities against the production of four cytokines (IL-6, IL-10, MCP-1, and TNF-α) in the serum of human acute monocytic leukemia cell line, THP-1, by using the human inflammation cytometric bead array (CBA) assay. Pretreatment of THP-1 cells with 188 and 216 exhibited a significant reduction in the LPS-induced expression of IL-10 with an inhibitory rate of 84.3% and 78.1% (p < 0.01), respectively, while 209 showed only a 23.6% decrease. The inhibition of the levels of IL-6, MCP-1, and TNF-α, ranged from 28.0% to 64.2% [99].
Compounds 238–244 (Figure 19), isolated from the hydrothermal vent sediment-derived fungus, A. pseudoviridinutans TW58-5, were tested for their capacity to inhibit NO production in LPS-induced murine macrophage RAW264.7 cells. Compounds 238–244, particularly 243, inhibited the NO production at a concentration of 20 μM. Compound 243 also showed the dose-dependent inhibition of NO production. Moreover, 243 effectively reduced the expression of iNOS (the enzyme responsible for NO production) and NLRP3 (an inflammatory mediator), suggesting that 243 could be a promising compound for the development of an anti-inflammatory agent [116].
4.13. Inhibition of G-Protein-Coupled Receptors
Compounds 211 and 212 (Figure 18), isolated from a marine sponge-associated fungus, Stachylidium sp., were examined for their inhibition of G-protein-coupled receptors (GPCRs) using a radioligand binding assays. Compound 211 showed an affinity to the vasopressin receptor 1A with Ki of 7.04 μM, while 212 was found to have a selective affinity to the serotonin receptor 5H2b, with Ki of 0.77 μM. It is important to note that 5HT2b receptors have been reported to play an important role in cardiac, intestinal, and central levels, and in bone marrow formation and growth [103].
4.14. Inhibition of Transporter Proteins
To tackle the resistance of multiple chemotherapeutic agents, targeting the transporter proteins such as P-glycoprotein (P-gp) is vital since it is one of the various factors that contribute to the multidrug resistant (MDR) factors. Thus, P-gp inhibitors that offer the prospect of reversing the MDR phenotype deserve to be investigated. Compound 122 (Figure 14), isolated from a marine sediment-derived fungus, Aspergillus sp. (CMB-M081F), was evaluated for P-gp inhibitory activity using a Calcein AM assay. This assay is based on the fact that, in the presence of a P-gp inhibitor, calcein AM efflux is blocked and calcein AM (nonfluorescent) undergoes hydrolysis to calcein (fluorescent), leading to an increase in intracellular fluorescence. Intracellular calcein fluorescence is quantified by cell flow cytometry to arrive at a fluorescence arbitrary ratio (FAR), which measures intracellular calcein fluorescence in cells exposed to a putative P-gp inhibitor, with that from cells not exposed to an inhibitor. Thus, the larger the FAR value, the more effective the P-gp inhibitor. Compound 122 inhibited the P-gp with FAR = 35.5. The positive control, verapamil, exhibited FAR = 72.5 [82].
4.15. Anti-Diabetic Activity
In order to examine the insulin sensitivity, the levels of adiponectin production in the adipogenesis model of human bone marrow mesenchymal stem cells (hBM-MSCs) are used. In a cell-based assay, 27 and 28 (Figure 4), isolated from a marine sediment-derived fungus, A. terreus FA009, were found to increase the adiponectin production during adipogenesis in hBM-MSCs, with EC50 values of 37.1 and 91.9 μM, respectively. The positive controls, glibenclamide and aspirin, showed the EC50 values of 3.47 and 145.6 μM, respectively. It is important to note that pharmacological mechanisms for improving the insulin sensitivity of glibenclamide and aspirin are different. The antidiabetic activity of glibenclamide is through binding with sulfonylurea receptor 1 (SUR1), which inhibits the conductance of the adenosine triphosphate (ATP)-dependent potassium (KATP) channel, whereas aspirin inhibits the serine kinase IKKβ. Although aspirin significantly increased the production of adiponectin, the maximum effect achieved by aspirin in the hBM-MSCs was less than that of glibenclamide. Interestingly, 27 induced the maximum increase in adiponectin levels by 56.9% relative to that produced by glibenclamide [48].
4.16. Antioxidant Activity
Compound (±)-189 (Figure 16), isolated from a marine sediment-derived fungus, Eurotium sp. SCSIO F452, scavenged the DPPH radicals with an IC50 = 58.4 μM, while the pure (−)-189 showed DPPH radicals scavenging activity with IC50 = 159.2 μM. The positive control, ascorbic acid, showed IC50 value of 45.8 μM [100].
4.17. Wound-Healing Activity
Compound 317 (Figure 25), isolated from a marine sediment-derived fungus, Acremonium sp. CNQ-049, did not show cytotoxicity against cancer (Caco2) and non-cancer [HaCaT (a spontaneously immortalized keratinocyte) and NIH3T3 (fibroblast)] cell lines at concentrations below 50 μM. Since 317 enhanced cell viability and increased effectively the mortality of the HaCaT and NIH3T3 cell lines, Kim et al. proposed that 317 might be beneficial for wound healing in the skin of mice by testing three genes related to human wound healing, including actin alpha (ACTC1), collagen type I alpha 2 (COL1A2), and matrix metallopeptidase 1 (MMP1). Quantitative real-time polymerase chain reaction (qRT-PCR) experiments showed that 317 significantly induced the expression of ACTC1 and COL1A2 in both the HaCaT and NIH3T3 cell lines. The increase in COL1A2 expression in both HaCaT and NIH3T3 cells could promote tissue growth and the remodeling phase, thereby potentiating wound closure. On the other hand, ACTC1, an isoform of actin alpha, can play an important role in wound repair by mediating contraction, the recruitment of repair machinery, and cell migration [147].
4.18. Larvicidal Activity
Compounds 331 and 332 (Figure 25), isolated from the fermentation extract of A. flavus, were tested for larvicidal activity against third-instar larvae of Culex pipiens mosquitoes, at concentrations of 10, 50, 100, 200, and 300 ppm. The mortality rates were recorded at various time intervals, i.e., 24, 48, 72, and 96 h. After 96 h of treatment, the highest mortality rates were detected for 331 (ranging from 26.66% to 82.66%), followed by 332 (ranging from 25.33% to 80.0%). Interestingly, 331 exhibited the most potent larvicidal activity, with LC50 (72h) and LC50 (96h) = 69.96 and 58.96 ppm, respectively. The corresponding positive control, toosenedanin, showed LC50 (72h) and LC90 (72h) = 2.66 and 10.0 ppm, respectively. In silico molecular docking studies of 331 and 332 for larvicidal chitinase enzyme activity were performed. Upon docking, the ligand showed the −(C-docker interaction energy) of −49.23 kcal/mol, and six essential hydrogen bonds with Glu177, Tyr245, Asp246, Arg301, and Glu322. On the other hand, 331 and 332 showed relatively better–(C-docker interaction energy) than the ligand, with 65.85 and −59.25 kcal/mol, respectively, The most active conformer of 331 displayed interaction with chitinase through three hydrogen bonds with Arg301, Trp137, and Ser250, while 332 showed further three hydrogen bonds with Asp246, Arg301, and Glu322 [152].
To facilitate the readers, the names and numbers of marine-derived peptides (according to their classes), the producing organisms, the sources from which the fungi were isolated, biological/pharmacological activities of the bioactive peptides, and the corresponding references are listed in Table 1.
5. Concluding Remarks and Future Perspectives
With a growing body of research on bioactive peptides, coupled with an accelerating pace of drug development with peptides, nowadays, more than 170 peptides and their derivatives are are in active clinical development [4]. Therefore, efforts to concentrate on marine fungal peptides, which are less considered, deserve further investigations.
A systematic investigation revealed a great potential for marine peptides to modulate critical biological events and pathways. Thus, peptides can serve as valuable models for designing novel lead compounds. Intriguingly, most of the reported biological/pharmacological activities of marine peptides are performed in vitro. Therefore, further investigations in vivo are needed to shed light on the potential of this class of compounds for further structure modifications or semi-synthesis to obtain analogs with better efficacy and stability in vivo. However, when performing in vivo studies, the dynamics shift significantly due to the emergence of peptide bioavailability concerns. For instance, when peptides are administered orally, they face challenges such as gastric acid exposure and enzymatic degradation in the stomach. This necessitates protective strategies to ensure the preservation of peptide integrity. Consequently, ensuring the presence of bioactive peptides at the intended target site demands a reassurance that traditional administration methods may not provide. This is where innovative approaches of pharmaceutical technology like nanoencapsulations, nanoemulsions, and nanotubes for drug delivery come into play. Moreover, the technology of linking bioactive peptides to chimeric antibodies to produce peptide-based drug-conjugates has shown to be an effective way to direct bioactive peptides to the targets. Despite the relevant activities of peptides, safety, tolerability, and efficacy profiles in the human body are fundamental.
Therefore, marine peptides provide salient opportunities not only for expanding the pharmacological profiles but also for designing the synthesis of unique peptides. In recent years, we have witnessed increasing numbers of FDA-approved marine peptides, which reflect the potential of these metabolites for drug discovery and development.
Hence, the importance of marine peptides is being highlighted since more than 36,000 marine natural products possessing various biological activities have been reported so far [171]. Finally, but yet importantly, in order to explore the potential of bioactive peptides, evaluations of less-considered biological/pharmacological targets are also strongly recommended.
To the best of our knowledge, this review is the first comprehensive review of marine-derived peptides obtained from 30 marine-derived fungal genera, which discusses their chemistry and biological activities. Overall, 366 peptides belonging to various classes are presented (Figure 27). The variation in the amino acid sequences, the chemical features, and a myriad of biological/pharmacological activities, such as cytotoxic activity against various human cancer cell lines, antibacterial and antibiofilm, antifungal, antiviral, antiparasitic, enzyme inhibitory activity, anti-angiogenesis, and anti-inflammatory activity, the inhibition of G-protein-coupled receptors, the inhibition of transporter proteins, anti-diabetic properties, and wound healing make marine peptides an interesting group of compounds to be explored for their potential in drug discovery and other health applications. It is worth mentioning also that a majority of marine peptides were isolated from marine-derived Aspergillus species (174 peptides from 48 species), but, to lesser extent, from other fungal genera such as Acremonium, Penicillium, Eurotium, Talaromyces, Trichoderma, and others. Moreover, the cytotoxic activity against various human cancer cell lines is the most reported bioactivity for these metabolites. Some of the reported cyclic heptapeptides and cyclic depsipeptides also showed antibacterial and antifungal activities (Figure 28).
Unlike several peptides isolated from marine invertebrates, none of the peptides isolated from marine-derived fungi have made their way to the advanced phase of clinical trial, except for halimide (a naturally occurring diketopiperazine derivative derived from the marine fungus Aspergillus sp. CNC139) whose synthetic analog, plinabulin (NPI-2358), was in a complete Phase III clinical trial by BeyondSpring Inc. in development for use in combination with granulocyte colony-stimulating factor for the prevention of chemotherapy-induced neutropenia [172]. Thus, new approaches for optimizing the potential of marine fungal peptides should be considered. The first approach is the optimization and modification of the culture media to stimulate the fungal silence genes to produce novel compounds. In this aspect, although the OSMAC (One Strain MAny Compounds) approach has been used to produced new compounds, so far this technique has not yet provided any concrete example of compounds for clinical trials. Another approach that should be considered is the addition of synthetic pharmacophores that contain functional groups that can link peptides to the culture media to explore the possibility of the production of novel bioactive peptides.
Author Contributions
S.H.G. and A.K. engaged in the conceptualization and design of the manuscript; S.H.G. and F.T. conducted the literature survey, designed and drew the structures, and prepared the draft of the manuscript; E.S. and C.F. gave suggestions and helped in revising the manuscript; A.K. revised and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work is partially supported by national funds through the FCT—Foundation for Science and Technology with the scope of UIDB/04423/2020 and UIDP/04423/2020, and by the structured program of the R&D&I ATLANTIDA Platform for the monitoring of the North Atlantic Ocean and tools for the sustainable exploitation of marine resources (reference NORTE-01-0145-FEDER-000040), supported by the North Portugal Regional Operational Program (NORTE2020) through the European Regional Development Fund (ERDF).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable.
Acknowledgments
The authors thank the Instituto de Ciências Biomédicas Abel Salazar of the University of Porto for their logistical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hafez Ghoran, S.; Taktaz, F.; Ayatollahi, S.A.; Kijjoa, A. Anthraquinones and their analogues from marine-derived fungi: Chemistry and biological activities. Mar. Drugs 2022, 20, 474. [Google Scholar] [CrossRef]
- Hafez Ghoran, S.; Kijjoa, A. Marine-derived compounds with anti-Alzheimer’s disease activities. Mar. Drugs 2021, 19, 410. [Google Scholar] [CrossRef] [PubMed]
- Giordano, D.; Costantini, M.; Coppola, D.; Lauritano, C.; Pons, L.N.; Ruocco, N.; di Prisco, G.; Ianora, A.; Verde, C. Biotechnological applications of bioactive peptides from marine sources. Adv. Microb. Physiol. 2018, 73, 171–220. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, N.; Zhang, W.; Cheng, X.; Yan, Z.; Shao, G.; Wang, X.; Wang, R.; Fu, C. Therapeutic peptides: Current applications and future directions. Signal Transduct. Target. Ther. 2022, 7, 48. [Google Scholar] [CrossRef] [PubMed]
- Youssef, F.S.; Ashour, M.L.; Singab, A.N.B.; Wink, M. A comprehensive review of bioactive peptides from marine fungi and their biological significance. Mar. Drugs 2019, 17, 559. [Google Scholar] [CrossRef] [PubMed]
- Lamers, C. Overcoming the shortcomings of peptide-based therapeutics. Future Drug Discov. 2022, 4, FDD75. [Google Scholar] [CrossRef]
- Liang, X.; Luo, D.; Luesch, H. Advances in exploring the therapeutic potential of marine natural products. Pharmacol. Res. 2019, 147, 104373. [Google Scholar] [CrossRef]
- Sridhar, K.; Inbaraj, B.S.; Chen, B.H. Recent developments on production, purification and biological activity of marine peptides. Food Res. Inter. 2021, 147, 110468. [Google Scholar] [CrossRef]
- Sable, R.; Parajuli, P.; Jois, S. Peptides, peptidomimetics, and polypeptides from marine sources: A wealth of natural sources for pharmaceutical applications. Mar. Drugs 2017, 15, 124. [Google Scholar] [CrossRef]
- McIntosh, J.A.; Donia, M.S.; Schmidt, E.W. Ribosomal peptide natural products: Bridging the ribosomal and nonribosomal worlds. Nat. Prod. Rep. 2009, 26, 537–559. [Google Scholar] [CrossRef]
- Hetrick, K.J.; van der Donk, W.A. Ribosomally synthesized and post-translationally modified peptide natural product discovery in the genomic era. Curr. Opin. Chem. Biol. 2017, 38, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Proksch, P.; Putz, A.; Ortlepp, S.; Kjer, J.; Bayer, M. Bioactive natural products from marine sponges and fungal endophytes. Phytochem. Rev. 2010, 9, 475–489. [Google Scholar] [CrossRef]
- Ruiz-Ruiz, F.; Mancera-Andrade, E.I.; Iqbal, H.M.N. Marine-derived bioactive peptides for biomedical sectors: A review. Protein Pep. Lett. 2017, 24, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Pavlicevic, M.; Maestri, E.; Marmiroli, M. Marine bioactive peptides—An overview of generation, structure and application with a focus on food sources. Mar. Drugs 2020, 18, 424. [Google Scholar] [CrossRef] [PubMed]
- Gogineni, V.; Hamann, M.T. Marine natural product peptides with therapeutic potential: Chemistry, biosynthesis, and pharmacology. Biochim. Biophys. Acta Gen. Sub. 2018, 1862, 81–196. [Google Scholar] [CrossRef] [PubMed]
- Helmy, N.M.; Parang, K. Cyclic peptides with antifungal properties derived from bacteria, fungi, plants, and synthetic sources. Pharmaceuticals 2023, 16, 892. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.; Ribeiro, R.; Pinto, M.; Kijjoa, A. Absolute Stereochemistry Determination of bioactive marine-derived cyclopeptides by liquid chromatography methods: An update review (2018–2022). Molecules 2023, 28, 615. [Google Scholar] [CrossRef] [PubMed]
- Pope, J.E.; Deer, T.R. Ziconotide: A clinical update and pharmacologic review. Expert Opin. Pharmacother. 2013, 14, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.J. Brentuximab vedotin: A review in CD30-positive Hodgkin lymphoma. Drugs 2017, 77, 435–445. [Google Scholar] [CrossRef]
- Malecek, M.K.; Watkins, M.P.; Bartlett, N.L. Polatuzumab vedotin for the treatment of adults with relapsed or refractory diffuse large B-cell lymphoma. Expert Opin. Biol. Ther. 2021, 21, 831–839. [Google Scholar] [CrossRef]
- Chang, E.; Weinstock, C.; Zhang, L.; Charlab, R.; Dorff, S.E.; Gong, Y.; Hsu, V.; Li, F.; Ricks, T.K.; Song, P.; et al. FDA approval summary: Enfortumab vedotin for locally advanced or metastatic urothelial carcinoma. Clin. Cancer Res. 2021, 27, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Deeks, E.D. Disitamab Vedotin: First approval. Drugs 2021, 81, 1929–1935. [Google Scholar] [CrossRef] [PubMed]
- Markham, A. Tisotumab Vedotin: First approval. Drugs 2021, 81, 2141–2147. [Google Scholar] [CrossRef] [PubMed]
- Approved Marine Drugs. Available online: https://www.marinepharmacology.org/approved (accessed on 27 August 2023).
- van Andel, L.; Fudio, S.; Rosing, H.; Munt, S.; Miguel-Lillo, B.; González, I.; Tibben, M.M.; de Vries, N.; de Vries Schultink, A.H.M.; Schellens, A.H.M.; et al. Pharmacokinetics and excretion of 14C–Plitidepsin in patients with advanced cancer. Investig. New Drugs 2017, 35, 589–598. [Google Scholar] [CrossRef]
- Pennington, M.W.; Czerwinski, A.; Norton, R.S. Peptide therapeutics from venom: Current status and potential. Bioorg. Med. Chem. 2018, 26, 2738–2758. [Google Scholar] [CrossRef]
- Poth, A.G.; Chiu, F.C.K.; Stalmans, S.; Hamilton, B.R.; Huang, Y.-H.; Shackleford, D.M.; Patil, R.; Le, T.T.; Kan, M.-W.; Durek, T.; et al. Effects of backbone cyclization on the pharmacokinetics and drug efficiency of the orally active analgesic conotoxin cVc1.1. Med. Drug Discov. 2021, 10, 100087. [Google Scholar] [CrossRef]
- Kineta Announces First Participant Dosed in Phase 1 Multiple Ascending Dose Clinical Trial of KCP506. Available online: https://kinetabio.com/2021/07/14/kineta-announces-first-participant-dosed-in-phase-1-multiple-ascending-dose-clinical-trial-of-kcp506/ (accessed on 24 August 2023).
- Martín-Algarra, S.; Espinosa, E.; Rubió, J.; López, J.J.L.; Manzano, J.L.; Carrión, L.A.; Plazaola, A.; Tanovic, A.; Paz-Ares, L. Phase II study of weekly kahalalide F in patients with advanced malignant melanoma. Eur. J. Cancer 2009, 45, 732–735. [Google Scholar] [CrossRef]
- Ratain, M.J.; Geary, D.; Undevia, S.D.; Coronado, C.; Alfaro, V.; Iglesias, J.L.; Schilsky, R.L.; Miguel-Lillo, B. First-in-human, phase I study of elisidepsin (PM02734) administered as a 30-min or as a 3-hour intravenous infusion every three weeks in patients with advanced solid tumors. Investig. New Drugs 2015, 33, 901–910. [Google Scholar] [CrossRef]
- Mayer, A.M.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef]
- Vaklavas, C.; Forero, A. Management of metastatic breast cancer with second-generation antibody–drug conjugates: Focus on glembatumumab vedotin (cdx-011, cr011-vcmmae). BioDrugs 2014, 28, 253–263. [Google Scholar] [CrossRef]
- Patel, S.; Keohan, M.L.; Saif, M.W.; Rushing, D.; Baez, L.; Feit, K.; DeJager, R.; Anderson, S. Phase II study of intravenous TZT-1027 in patients with advanced or metastatic soft-tissue sarcomas with prior exposure to anthracycline-based chemotherapy. Cancer 2006, 107, 2881–2887. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Lima, C.M.; Bayraktar, S.; MacIntyre, J.; Raez, L.; Flores, A.M.; Ferrell, A.; Rubin, E.H.; Poplin, E.A.; Tan, A.R.; Lucarelli, A.; et al. A phase 1 trial of E7974 administered on day 1 of a 21-day cycle in patients with advanced solid tumors. Cancer 2012, 118, 4262–4270. [Google Scholar] [CrossRef] [PubMed]
- Hadaschik, B.A.; Ettinger, S.; Sowery, R.D.; Zoubeidi, A.; Andersen, R.J.; Roberge, M.; Gleave, M.E. Targeting prostate cancer with HTI-286, a synthetic analog of the marine sponge product hemiasterlin. Int. J. Cancer 2008, 122, 2368–2376. [Google Scholar] [CrossRef]
- Bordon, K.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From animal poisons and venoms to medicines: Achievements, challenges and perspectives in drug discovery. Front. Pharmacol. 2020, 11, 132. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Phat, C.; Hong, S.C. Structural diversity of marine cyclic peptides and their molecular mechanisms for anticancer, antibacterial, antifungal, and other clinical applications. Peptides 2017, 95, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, X.Y.; Nong, X.H.; Wang, J.; Huang, Z.H.; Qi, S.H. Eight linear peptides from the deep-sea-derived fungus Simplicillium obclavatum EIODSF 020. Tetrahedron 2016, 72, 3092–3097. [Google Scholar] [CrossRef]
- Anh, C.V.; Yoon, Y.D.; Kang, J.S.; Lee, H.S.; Heo, C.S.; Shin, H.J. Nitrogen-containing secondary metabolites from a deep-sea fungus Aspergillus unguis and their anti-inflammatory activity. Mar. Drugs 2022, 20, 217. [Google Scholar] [CrossRef]
- Yao, F.H.; Liang, X.; Qi, S.H. Two new linear peptides from the marine-derived fungus Penicillium sp. SCSIO 41512. J. Asian Nat. Prod. Res. 2023, 25, 941–948. [Google Scholar] [CrossRef]
- Ma, X.; Nong, X.H.; Ren, Z.; Wang, J.; Liang, X.; Wang, L.; Qi, S.H. Antiviral peptides from marine gorgonian-derived fungus Aspergillus sp. SCSIO 41501. Tetrahedron Lett. 2017, 58, 1151–1155. [Google Scholar] [CrossRef]
- Xiao-Wei, L.; Yun, L.; Yong-Jun, L.; Xue-Feng, Z.; Yong-Hong, L. Peptides and polyketides isolated from the marine sponge-derived fungus Aspergillus terreus SCSIO 41008. Chin. J. Nat. Med. 2019, 17, 149–154. [Google Scholar] [CrossRef]
- Girich, E.V.; Rasin, A.B.; Popov, R.S.; Yurchenko, E.A.; Chingizova, E.A.; Trinh, P.T.H.; Ngoc, N.T.D.; Pivkin, M.V.; Zhuravleva, O.I.; Yurchenko, A.N. New tripeptide derivatives asterripeptides A–C from Vietnamese mangrove-derived fungus Aspergillus terreus LM. 5.2. Mar. Drugs 2022, 20, 77. [Google Scholar] [CrossRef]
- Zhou, C.; Cao, X.; Ge, Y.; Wu, X.; Zhang, Z.; Ma, Y.; Dickschat, J.S.; Wu, B. Talaropeptins A and B, tripeptides with an N-trans-cinnamoyl moiety from the marine-derived fungus Talaromyces purpureogenus CX11. J. Nat. Prod. 2022, 85, 2620–2625. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.W.; Mordhorst, T.F.; Lee, C.; Jensen, P.R.; Fenical, W.; Köck, M. Penilumamide, a novel lumazine peptide isolated from the marine-derived fungus, Penicillium sp. CNL-338. Org. Biomol. Chem. 2010, 8, 2158–2163. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Shao, C.L.; Fu, X.M.; Kong, C.J.; She, Z.G.; Wang, C.Y. Lumazine peptides penilumamides B–D and the cyclic pentapeptide asperpeptide A from a gorgonian-derived Aspergillus sp. fungus. J. Nat. Prod. 2014, 77, 1601–1606. [Google Scholar] [CrossRef]
- Zheng, C.J.; Wu, L.Y.; Li, X.B.; Song, X.M.; Niu, Z.G.; Song, X.P.; Chen, G.Y.; Wang, C.Y. Structure and absolute configuration of aspergilumamide A, a novel lumazine peptide from the mangrove-derived fungus Aspergillus sp. Helv. Chim. Acta 2015, 98, 368–373. [Google Scholar] [CrossRef]
- You, M.; Liao, L.; Hong, S.H.; Park, W.; Kwon, D.I.; Lee, J.; Noh, M.; Oh, D.-C.; Oh, K.-B.; Shin, J. Lumazine peptides from the marine-derived fungus Aspergillus terreus. Mar. Drugs 2015, 13, 1290–1303. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Nong, X.H.; Huang, Z.H.; Qi, S.H. Antifungal and antiviral cyclic peptides from the deep-sea-derived fungus Simplicillium obclavatum EIODSF 020. J. Agr. Food Chem. 2017, 65, 5114–5121. [Google Scholar] [CrossRef]
- Yao, F.H.; Liang, X.; Cheng, X.; Ling, J.; Dong, J.D.; Qi, S.H. Antifungal peptides from the marine gorgonian-associated fungus Aspergillus sp. SCSIO41501. Phytochemistry 2021, 192, 112967. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.Y.; Lee, J.H.; Park, S.C.; Lee, J.; Oh, D.C.; Oh, K.B.; Shin, J. New peptides from the marine-derived fungi Aspergillus allahabadii and Aspergillus ochraceopetaliformis. Mar. Drugs 2019, 17, 488. [Google Scholar] [CrossRef]
- Boot, C.M.; Tenney, K.; Valeriote, F.A.; Crews, P. Highly N-methylated linear peptides produced by an atypical sponge-derived Acremonium sp. J. Nat. Prod. 2006, 69, 83–92. [Google Scholar] [CrossRef]
- Boot, C.M.; Amagata, T.; Tenney, K.; Compton, J.E.; Pietraszkiewicz, H.; Valeriote, F.A.; Crews, P. Four classes of structurally unusual peptides from two marine-derived fungi: Structures and bioactivities. Tetrahedron 2007, 63, 9903–9914. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Xue, C.; Tian, L.; Xu, M.; Chen, J.; Deng, Z.; Proksch, P.; Lin, W. Asperelines A−F, peptaibols from the marine-derived fungus Trichoderma asperellum. J. Nat. Prod. 2009, 72, 1036–1044. [Google Scholar] [CrossRef] [PubMed]
- Dewapriya, P.; Khalil, Z.G.; Prasad, P.; Salim, A.A.; Cruz-Morales, P.; Marcellin, E.; Capon, R.J. Talaropeptides A-D: Structure and biosynthesis of extensively N-methylated linear peptides from an Australian marine tunicate-derived Talaromyces sp. Front. Chem. 2018, 6, 394. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, K.; Shigemori, H.; Kobayashi, J. Dictyonamides A and B, new peptides from marine-derived fungus. J. Org. Chem. 2001, 66, 6189–6192. [Google Scholar] [CrossRef] [PubMed]
- Morehouse, N.J.; Flewelling, A.J.; Liu, D.Y.; Cavanagh, H.; Linington, R.G.; Johnson, J.A.; Gray, C.A. Tolypocaibols: Antibacterial lipopeptaibols from a Tolypocladium sp. endophyte of the marine macroalga Spongomorpha arcta. J. Nat. Prod. 2023, 86, 1529–1535. [Google Scholar] [CrossRef] [PubMed]
- van Bohemen, A.-I.; Ruiz, N.; Zalouk-Vergnoux, A.; Michaud, A.; du Pont, T.R.; Druzhinina, I.; Atanasova, L.; Prado, S.; Bodo, B.; Meslet-Cladiere, L.; et al. Pentadecaibins I–V: 15-residue peptaibols produced by a marine-derived Trichoderma sp. of the Harzianum clade. J. Nat. Prod. 2021, 84, 1271–1282. [Google Scholar] [CrossRef]
- Lin, X.; Tang, Z.; Gan, Y.; Li, Z.; Luo, X.; Gao, C.; Zhao, L.; Chai, L.; Liu, Y. 18-Residue peptaibols produced by the sponge-derived Trichoderma sp. GXIMD 01001. J. Nat. Prod. 2023, 86, 994–1002. [Google Scholar] [CrossRef]
- Rowley, D.C.; Kelly, S.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Halovirs A–E, new antiviral agents from a marine-derived fungus of the genus Scytalidium. Bioorg. Med. Chem. 2003, 11, 4263–4274. [Google Scholar] [CrossRef]
- Jiao, W.H.; Xu, Q.H.; Ge, G.B.; Shang, R.Y.; Zhu, H.R.; Liu, H.Y.; Cui, J.; Sun, F.; Lin, H.W. Flavipesides A–C, PKS-NRPS hybrids as pancreatic lipase inhibitors from a marine sponge symbiotic fungus Aspergillus flavipes 164013. Org. Lett. 2020, 22, 1825–1829. [Google Scholar] [CrossRef]
- Liao, L.; Bae, S.Y.; Won, T.H.; You, M.; Kim, S.H.; Oh, D.C.; Lee, S.K.; Oh, K.B.; Shin, J. Asperphenins A and B, lipopeptidyl benzophenones from a marine-derived Aspergillus sp. fungus. Org. Lett. 2017, 19, 2066–2069. [Google Scholar] [CrossRef]
- Shigemori, H.; Wakuri, S.; Yazawa, K.; Nakamura, T.; Sasaki, T.; Kobayashi, J. Fellutamides A and B, cytotoxic peptides from a marine fish-possessing fungus Penicillium fellutanum. Tetrahedron 1991, 47, 8529–8534. [Google Scholar] [CrossRef]
- Scopel, M.; Abraham, W.R.; Henriques, A.T.; Macedo, A.J. Dipeptide cis-cyclo (Leucyl-Tyrosyl) produced by sponge associated Penicillium sp. F37 inhibits biofilm formation of the pathogenic Staphylococcus epidermidis. Bioorg. Med. Chem. Lett. 2013, 23, 624–626. [Google Scholar] [CrossRef]
- Uchoa, P.K.S.; Pimenta, A.T.A.; Braz-Filho, R.; de Oliveira, M.C.F.; Saraiva, N.N.; Rodrigues, B.S.F.; Pfenning, L.H.; Abreu, L.M.; Wilke, D.V.; Florêncio, K.G.; et al. New cytotoxic furan from the marine sediment-derived fungi Aspergillus niger. Nat. Prod. Res. 2017, 31, 2599–2603. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wei, J.; Tang, J.; Wu, B. Two new prenylated glycine derivatives from the marine-derived fungus Fusarium sp. TW56-10. Chem. Biodivers. 2022, 19, e202100899. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, L.; Sun, S.; Chang, A.; Dai, X.; Li, H.; Wang, Y.; Zhu, H. A cyclic dipeptide from marine fungus Penicillium chrysogenum DXY-1 exhibits anti-quorum sensing activity. ACS Omega 2021, 6, 7693–7700. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Cui, C.B.; Li, C.W. A new cyclic dipeptide penicimutide: The activated production of cyclic dipeptides by introduction of neomycin-resistance in the marine-derived fungus Penicillium purpurogenum G59. Arch. Pharm. Res. 2016, 39, 762–770. [Google Scholar] [CrossRef]
- Xie, L.R.; Li, D.Y.; Wang, P.L.; Hua, H.M.; Wu, X.; Li, Z.L. A new 3,4-seco-lanostane triterpenoid from a marine-derived fungus Ascotricha sp. ZJ-M-5. Yao Xue Xue Bao 2013, 48, 89–93. [Google Scholar]
- May Zin, W.W.; Buttachon, S.; Dethoup, T.; Fernandes, C.; Cravo, S.; Pinto, M.M.; Gales, L.; Pereira, J.A.; Silva, A.M.S.; Sekeroglu, N.; et al. New cyclotetrapeptides and a new diketopiperzine derivative from the marine sponge-associated fungus Neosartorya glabra KUFA 0702. Mar. Drugs 2016, 14, 136. [Google Scholar] [CrossRef]
- Capon, R.J.; Skene, C.; Stewart, M.; Ford, J.; Richard, A.; Williams, L.; Lacey, E.; Gill, J.H.; Heiland, K.; Friedel, T. Aspergillicins A–E: Five novel depsipeptides from the marine-derived fungus Aspergillus carneus. Org. Biomol. Chem. 2003, 1, 1856–1862. [Google Scholar] [CrossRef]
- May Zin, W.W.; Buttachon, S.; Buaruang, J.; Gales, L.; Pereira, J.A.; Pinto, M.M.; Silva, A.M.S.; Kijjoa, A. A new meroditerpene and a new tryptoquivaline analog from the algicolous fungus Neosartorya takakii KUFC 7898. Mar. Drugs 2015, 13, 3776–3790. [Google Scholar] [CrossRef]
- Bovio, E.; Fauchon, M.; Toueix, Y.; Mehiri, M.; Varese, G.C.; Hellio, C. The sponge-associated fungus Eurotium chevalieri MUT 2316 and its bioactive molecules: Potential applications in the field of antifouling. Mar. Biotechnol. 2019, 21, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Wijesekara, I.; Li, Y.-X.; Vo, T.S.; Ta, Q.V.; Ngo, D.H.; Kim, S.K. Induction of apoptosis in human cervical carcinoma HeLa cells by neoechinulin A from marine-derived fungus Microsporum sp. Process Biochem. 2013, 48, 68–72. [Google Scholar] [CrossRef]
- Smetanina, O.F.; Kalinovskii, A.I.; Khudyakova, Y.V.; Slinkina, N.N.; Pivkin, M.V.; Kuznetsova, T.A. Metabolites from the marine fungus Eurotium repens. Chem. Nat. Comp. 2007, 43, 395–398. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.M.; Feng, Y.; Wang, B.G. Phenethyl-α-pyrone derivatives and cyclodipeptides from a marine algous endophytic fungus Aspergillus niger EN–13. Nat. Prod. Res. 2010, 24, 1036–1043. [Google Scholar] [CrossRef]
- Buttachon, S.; Ramos, A.A.; Inácio, Â.; Dethoup, T.; Gales, L.; Lee, M.; Costa, P.M.; Silva, A.M.S.; Sekeroglu, N.; Rocha, E.; et al. Bis-indolyl benzenoids, hydroxypyrrolidine derivatives and other constituents from cultures of the marine sponge-associated fungus Aspergillus candidus KUFA0062. Mar. Drugs 2018, 16, 119. [Google Scholar] [CrossRef]
- Liao, Y.; Hong, X.; Yang, J.; Qin, J.J.; Zhang, B.; Lin, J.; Shao, Z.; Wang, W. Structure elucidation of a novel cyclic tripeptide from the marine-derived fungus Aspergillus ochraceopetaliformis DSW-2. Nat. Prod. Res. 2022, 36, 3572–3578. [Google Scholar] [CrossRef]
- Zhou, X.; Fang, P.; Tang, J.; Wu, Z.; Li, X.; Li, S.; Wang, Y.; Liu, G.; He, Z.; Gou, D.; et al. A novel cyclic dipeptide from deep marine-derived fungus Aspergillus sp. SCSIOW2. Nat. Prod. Res. 2016, 30, 52–57. [Google Scholar] [CrossRef]
- Lee, S.U.; Asami, Y.; Lee, D.; Jang, J.H.; Ahn, J.S.; Oh, H. Protuboxepins A and B and protubonines A and B from the marine-derived fungus Aspergillus sp. SF-5044. J. Nat. Prod. 2011, 74, 1284–1287. [Google Scholar] [CrossRef]
- Ebada, S.S.; Fischer, T.; Hamacher, A.; Du, F.Y.; Roth, Y.O.; Kassack, M.U.; Wang, B.G.; Roth, E.H. Psychrophilin E, a new cyclotripeptide, from co-fermentation of two marine alga-derived fungi of the genus Aspergillus. Nat. Prod. Res. 2014, 28, 776–781. [Google Scholar] [CrossRef]
- Khalil, Z.G.; Huang, X.; Raju, R.; Piggott, A.M.; Capon, R.J. Shornephine A: Structure, chemical stability, and P-glycoprotein inhibitory properties of a rare diketomorpholine from an Australian marine-derived Aspergillus sp. J. Org. Chem. 2014, 79, 8700–8705. [Google Scholar] [CrossRef]
- Gulder, T.A.M.; Hong, H.; Correa, J.; Egereva, E.; Wiese, J.; Imhoff, J.F.; Gross, H. Isolation, structure elucidation and total synthesis of lajollamide A from the marine fungus Asteromyces cruciatus. Mar. Drugs 2012, 10, 2912–2935. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Li, Z.; Gao, J.; He, H.; Dai, H.; Xia, X.; Liu, C.; Zhang, L.; Song, F. New diketopiperazines from a marine-derived fungus strain Aspergillus versicolor MF180151. Mar. Drugs 2019, 17, 262. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.X.; Jensen, P.R.; Williams, P.G.; Fenical, W. Isolation and structure assignments of rostratins A− D, cytotoxic disulfides produced by the marine-derived fungus Exserohilum rostratum. J. Nat. Prod. 2004, 67, 1374–1382. [Google Scholar] [CrossRef] [PubMed]
- Hong, R.; Yang, Y. Antitumor metabolites from marine sediment derived Penicillium sp. WF-06. World Notes Antibiots 2011, 2, 86–98. [Google Scholar]
- Wu, Q.X.; Crews, M.S.; Draskovic, M.; Sohn, J.; Johnson, T.A.; Tenney, K.; Valeriote, F.A.; Yao, X.J.; Bjeldanes, L.F.; Crews, P. Azonazine, a novel dipeptide from a Hawaiian marine sediment-derived fungus, Aspergillus insulicola. Org. Lett. 2010, 12, 4458–4461. [Google Scholar] [CrossRef]
- May Zin, W.W.; Buttachon, S.; Dethoup, T.; Pereira, J.A.; Gales, L.; Inácio, Â.; Costa, P.M.; Lee, M.; Sekeroglu, N.; Silva, A.M.S.; et al. Antibacterial and antibiofilm activities of the metabolites isolated from the culture of the mangrove-derived endophytic fungus Eurotium chevalieri KUFA 0006. Phytochemistry 2017, 141, 86–97. [Google Scholar] [CrossRef]
- Song, F.; Liu, X.; Guo, H.; Ren, B.; Chen, C.; Piggott, A.M.; Yu, K.; Gao, H.; Wang, Q.; Liu, M.; et al. Brevianamides with antitubercular potential from a marine-derived isolate of Aspergillus versicolor. Org. Lett. 2012, 14, 4770–4773. [Google Scholar] [CrossRef]
- Li, L.; Chang, Q.H.; Zhang, S.S.; Yang, K.; Chen, F.L.; Zhu, H.J.; Cao, F.; Liu, Y.F. (±)-Brevianamides Z and Z1, New diketopiperazine alkaloids from the marine-derived fungus Aspergillus versicolor. J. Mol. Struct. 2022, 1261, 132904. [Google Scholar] [CrossRef]
- Yan, H.J.; Li, X.M.; Li, C.S.; Wang, B.G. Alkaloid and anthraquinone derivatives produced by the marine-derived endophytic fungus Eurotium rubrum. Helv. Chim. Acta 2012, 95, 163–168. [Google Scholar] [CrossRef]
- Mao, J.Q.; Zheng, Y.Y.; Wang, C.Y.; Liu, Y.; Yao, G.S. Sclerotioloids A–C: Three new alkaloids from the marine-derived fungus Aspergillus sclerotiorum ST0501. Mar. Drugs 2023, 21, 219. [Google Scholar] [CrossRef]
- Youssef, D.T.; Shaala, L.A.; Genta-Jouve, G. Asperopiperazines A and B: Antimicrobial and cytotoxic dipeptides from a tunicate-derived fungus Aspergillus sp. DY001. Mar. Drugs 2022, 20, 451. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.-H.; Du, F.Y.; Li, X.M.; Pedpradab, P.; Xu, G.M.; Wang, B.G. Rubrumazines A–C, indolediketopiperazines of the isoechinulin class from Eurotium rubrum MA-150, a fungus obtained from marine mangrove-derived rhizospheric soil. J. Nat. Prod. 2015, 78, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, Y.; Li, S.; Hu, C.; Liu, H.; Zhang, W. Indole diketopiperazine alkaloids from the deep-sea-derived fungus Aspergillus sp. FS445. Nat. Prod. Res. 2022, 36, 5213–5221. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.H.; Du, F.Y.; Li, X.M.; Yang, S.Q.; Wang, B.G.; Li, X. Antibacterial indole diketopiperazine alkaloids from the deep-sea cold seep-derived fungus Aspergillus chevalieri. Mar. Drugs 2023, 21, 195. [Google Scholar] [CrossRef]
- Varoglu, M.; Crews, P. Biosynthetically diverse compounds from a saltwater culture of sponge-derived Aspergillus niger. J. Nat. Prod. 2000, 63, 41–43. [Google Scholar] [CrossRef]
- Carr, G.; Tay, W.; Bottriell, H.; Andersen, S.K.; Mauk, A.G.; Andersen, R.J. Plectosphaeroic acids A, B, and C, indoleamine 2,3-dioxygenase inhibitors produced in culture by a marine isolate of the fungus Plectosphaerella cucumerina. Org. Lett. 2009, 11, 2996–2999. [Google Scholar] [CrossRef]
- Liu, J.; Gu, B.; Yang, L.; Yang, F.; Lin, H. New anti-inflammatory cyclopeptides from a sponge-derived fungus Aspergillus violaceofuscus. Front. Chem. 2018, 6, 226. [Google Scholar] [CrossRef]
- Zhong, W.; Wang, J.; Wei, X.; Chen, Y.; Fu, T.; Xiang, Y.; Huang, X.; Tian, X.; Xiao, Z.; Zhang, W.; et al. Variecolortins A–C, three pairs of spirocyclic diketopiperazine enantiomers from the marine-derived fungus Eurotium sp. SCSIO F452. Org. Lett. 2018, 20, 4593–4596. [Google Scholar] [CrossRef]
- Peng, J.; Gao, H.; Zhang, X.; Wang, S.; Wu, C.; Gu, Q.; Guo, P.; Zhu, T.; Li, D. Psychrophilins E–H and versicotide C, cyclic peptides from the marine-derived fungus Aspergillus versicolor ZLN-60. J. Nat. Prod. 2014, 77, 2218–2223. [Google Scholar] [CrossRef]
- Zheng, J.; Xu, Z.; Wang, Y.; Hong, K.; Liu, P.; Zhu, W. Cyclic tripeptides from the halotolerant fungus Aspergillus sclerotiorum PT06-1. J. Nat. Prod. 2010, 73, 1133–1137. [Google Scholar] [CrossRef]
- Almeida, C.; Maddah, F.E.; Kehraus, S.; Schnakenburg, G.; König, G.M. Endolides A and B, vasopressin and serotonin-receptor interacting N-methylated peptides from the sponge-derived fungus Stachylidium sp. Org. Lett. 2016, 18, 528–531. [Google Scholar] [CrossRef] [PubMed]
- El Maddah, F.; Kehraus, S.; Nazir, M.; Almeida, C.; König, G.M. Insights into the biosynthetic origin of 3-(3-furyl) alanine in Stachylidium sp. 293 K04 tetrapeptides. J. Nat. Prod. 2016, 79, 2838–2845. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, J.; Luo, C.; Ding, W.; Cox, D.G. A new cyclopeptide with antifungal activity from the co-culture broth of two marine mangrove fungi. Nat. Prod. Res. 2014, 28, 616–621. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Seo, Y.; Lee, H.S.; Rho, J.R.; Mo, S.J. A new cyclic peptide from a marine-derived bacterium of the genus Nocardiopsis. J. Nat. Prod. 2003, 66, 883–884. [Google Scholar] [CrossRef]
- Zhou, Z.F.; Yang, X.H.; Liu, H.L.; Gu, Y.C.; Ye, B.P.; Guo, Y.W. A new cyclic peptide and a new steroid from the fungus Penicillium sp. GD6 isolated from the mangrove Bruguiera gymnorrhiza. Helv. Chim. Acta 2014, 97, 1564–1570. [Google Scholar] [CrossRef]
- He, F.; Bao, J.; Zhang, X.Y.; Tu, Z.C.; Shi, Y.M.; Qi, S.H. Asperterrestide A, a cytotoxic cyclic tetrapeptide from the marine-derived fungus Aspergillus terreus SCSGAF0162. J. Nat. Prod. 2013, 76, 1182–1186. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, S.; Yang, X. A new cyclopeptide metabolite of marine gut fungus from Ligia oceanica. Nat. Prod. Res. 2014, 28, 994–997. [Google Scholar] [CrossRef]
- Gu, W.; Cueto, M.; Jensen, P.R.; Fenical, W.; Silverman, R.B. Microsporins A and B: New histone deacetylase inhibitors from the marine-derived fungus Microsporum cf. gypseum and the solid-phase synthesis of microsporin A. Tetrahedron 2007, 63, 6535–6541. [Google Scholar] [CrossRef]
- Aparicio-Cuevas, M.A.; González, M.C.; Raja, H.A.; Rivero-Cruz, I.; Kurina, S.J.; Burdette, J.E.; Oberlies, N.H.; Figueroa, M. Metabolites from the marine-facultative Aspergillus sp. MEXU 27854 and Gymnoascus hyalinosporus MEXU 29901 from Caleta Bay, Mexico. Tetrahedron Lett. 2019, 60, 1649–1652. [Google Scholar] [CrossRef]
- Fremlin, L.J.; Piggott, A.M.; Lacey, E.; Capon, R.J. Cottoquinazoline A and cotteslosins A and B, metabolites from an Australian marine-derived strain of Aspergillus versicolor. J. Nat. Prod. 2009, 72, 666–670. [Google Scholar] [CrossRef]
- Jiao, F.R.; Zhu, H.R.; Gu, B.B.; Wu, Y.; Sun, F.; Wang, S.P.; Zhang, A.; Jiao, W.H.; Xu, S.H.; Lin, H.W. Asperflomide and asperflosamide, new N-methylated cyclopeptides from the marine sponge-derived fungus Aspergillus flocculosus 16D-1. Tetrahedron 2022, 109, 132579. [Google Scholar] [CrossRef]
- Li, X.; Xia, Z.; Wang, B.; Lai, L.; Wang, J.; Jiang, L.; Li, T.; Wu, J.; Wang, L. Malformin C, an algicidal peptide from marine fungus Aspergillus species. Ecotoxicology 2021, 30, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.N.; Gao, H.Q.; Cai, S.X.; Zhu, T.J.; Gu, Q.Q.; Li, D.H. Two new cyclic pentapeptides from the marine-derived fungus Aspergillus versicolor. Helv. Chim. Acta 2011, 94, 1065–1070. [Google Scholar] [CrossRef]
- Ding, W.; Tian, D.; Chen, M.; Xia, Z.; Tang, X.; Zhang, S.; Wei, J.; Li, X.; Yao, X.; Wu, B.; et al. Molecular networking-guided isolation of cyclopentapeptides from the hydrothermal Vent sediment derived fungus Aspergillus pseudoviridinutans TW58-5 and their anti-inflammatory effects. J. Nat. Prod. 2023, 86, 1919–1930. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.M.; Zhang, Y.H.; Hai, Y.; Zheng, J.Y.; Gu, Y.C.; Wang, C.Y.; Shao, C.L. Aspersymmetide A, a new centrosymmetric cyclohexapeptide from the marine-derived fungus Aspergillus versicolor. Mar. Drugs 2017, 15, 363. [Google Scholar] [CrossRef]
- Karim, F.; Liang, X.; Qi, S.-H. Bioassay-guided isolation of antifungal cyclopeptides from the deep-sea-derived fungus Simplicillium obclavatum EIODSF 020. Phytochem. Lett. 2022, 48, 68–71. [Google Scholar] [CrossRef]
- Zheng, J.; Zhu, H.; Hong, K.; Wang, Y.; Liu, P.; Wang, X.; Peng, X.; Zhu, W. Novel cyclic hexapeptides from marine-derived fungus, Aspergillus sclerotiorum PT06-1. Org. Lett. 2009, 11, 5262–5265. [Google Scholar] [CrossRef]
- Long, J.; Chen, Y.; Chen, W.; Wang, J.; Zhou, X.; Yang, B.; Liu, Y. Cyclic peptides from the soft coral-derived fungus Aspergillus sclerotiorum SCSIO 41031. Mar. Drugs 2021, 19, 701. [Google Scholar] [CrossRef]
- Prompanya, C.; Fernandes, C.; Cravo, S.; Pinto, M.M.M.; Dethoup, T.; Silva, A.M.S.; Kijjoa, A. A new cyclic hexapeptide and a new isocoumarin derivative from the marine sponge-associated fungus Aspergillus similanensis KUFA 0013. Mar. Drugs 2015, 13, 1432–1450. [Google Scholar] [CrossRef]
- Luo, M.; Zang, R.; Wang, X.; Chen, Z.; Song, X.; Ju, J.; Huang, H. Natural hydroxamate-containing siderophore acremonpeptides A–D and an aluminum complex of acremonpeptide D from the marine-derived Acremonium persicinum SCSIO 115. J. Nat. Prod. 2019, 82, 2594–2600. [Google Scholar] [CrossRef]
- Tang, W.Z.; Liu, J.T.; Hu, Q.; He, R.J.; Guan, X.Q.; Ge, G.B.; Han, H.; Yang, F.; Lin, H.-W. Pancreatic lipase inhibitory cyclohexapeptides from the marine sponge-derived fungus Aspergillus sp. 151304. J. Nat. Prod. 2020, 83, 2287–2293. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.M.; Liang, T.M.; Guo, Z.Y.; Wang, C.Y.; Shao, C.L. Discovery, absolute assignments, and total synthesis of asperversiamides A–C and their potent activity against Mycobacterium marinum. Chem. Comm. 2019, 55, 1104–1107. [Google Scholar] [CrossRef] [PubMed]
- Chao, R.; Hou, X.M.; Xu, W.F.; Hai, Y.; Wei, M.Y.; Wang, C.Y.; Gu, Y.C.; Shao, C.L. Targeted isolation of asperheptatides from a coral-derived fungus using LC-MS/MS-based molecular networking and antitubercular activities of modified cinnamate derivatives. J. Nat. Prod. 2020, 84, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Song, Y.; Chen, Y.; Huang, H.; Zhang, W.; Ju, J. Cyclic heptapeptides, cordyheptapeptides C–E, from the marine-derived fungus Acremonium persicinum SCSIO 115 and their cytotoxic activities. J. Nat. Prod. 2012, 75, 1215–1219. [Google Scholar] [CrossRef]
- Dewapriya, P.; Prasad, P.; Damodar, R.; Salim, A.A.; Capon, R.J. Talarolide A, a cyclic heptapeptide hydroxamate from an Australian marine tunicate-associated fungus, Talaromyces sp. (CMB-TU011). Org. Lett. 2017, 19, 2046–2049. [Google Scholar] [CrossRef]
- Grunwald, A.L.; Berrue, F.; Robertson, A.W.; Overy, D.P.; Kerr, R.G. Mortiamides A–D, cyclic heptapeptides from a novel Mortierella sp. obtained from Frobisher Bay. J. Nat. Prod. 2017, 80, 2677–2683. [Google Scholar] [CrossRef]
- Malmstrøm, J. Unguisins A and B: New cyclic peptides from the marine-derived fungus Emericella unguis. J. Nat. Prod. 1999, 62, 787–789. [Google Scholar] [CrossRef]
- Yang, W.C.; Bao, H.Y.; Liu, Y.Y.; Nie, Y.Y.; Yang, J.M.; Hong, P.Z.; Zhang, Y. Depsidone derivatives and a cyclopeptide produced by marine fungus Aspergillus unguis under chemical induction and by its plasma induced mutant. Molecules 2018, 23, 2245. [Google Scholar] [CrossRef]
- Liu, S.; Shen, Y. A new cyclic peptide from the marine fungal strain Aspergillus sp. AF119. Chem. Nat. Comp. 2011, 47, 786–788. [Google Scholar] [CrossRef]
- Tan, L.T.; Cheng, X.C.; Jensen, P.R.; Fenical, W. Scytalidamides A and B, new cytotoxic cyclic heptapeptides from a marine fungus of the genus Scytalidium. J. Org. Chem. 2003, 68, 8767–8773. [Google Scholar] [CrossRef]
- Adachi, K.; Kanoh, K.; Wisespongp, P.; Nishijima, M.; Shizuri, Y. Clonostachysins A and B, new anti-dinoflagellate cyclic peptides from a marine-derived fungus. J. Antibiot. 2005, 58, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Grunwald, A.L.; Cartmell, C.; Kerr, R.G. Auyuittuqamides A–D, cyclic decapeptides from Sesquicillium microsporum RKAG 186 isolated from frobisher bay sediment. J. Nat. Prod. 2020, 84, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Quezada, M.; Shang, Z.; Kalansuriya, P.; Salim, A.A.; Lacey, E.; Capon, R.J. Waspergillamide A, a nitro depsi-tetrapeptide diketopiperazine from an Australian mud dauber wasp-associated Aspergillus sp. (CMB-W031). J. Nat. Prod. 2017, 80, 1192–1195. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Wang, S.; Li, N.; Li, F.; Zhu, T.; Gu, Q.; Guo, P.; Li, D. Saroclides A and B, cyclic depsipeptides from the mangrove-derived fungus Sarocladium kiliense HDN11-112. J. Nat. Prod. 2018, 81, 1050–1054. [Google Scholar] [CrossRef] [PubMed]
- Ratnayake, R.; Fremlin, L.J.; Lacey, E.; Gill, J.H.; Capon, R.J. Acremolides A−D, lipodepsipeptides from an Australian marine-derived fungus, Acremonium sp. J. Nat. Prod. 2008, 71, 403–408. [Google Scholar] [CrossRef]
- Silber, J.; Ohlendorf, B.; Labes, A.; Näther, C.; Imhoff, J.F. Calcaripeptides A–C, cyclodepsipeptides from a Calcarisporium strain. J. Nat. Prod. 2013, 76, 1461–1467. [Google Scholar] [CrossRef]
- Wang, Q.; Xu, L. Beauvericin, a bioactive compound produced by fungi: A short review. Molecules 2012, 17, 2367–2377. [Google Scholar] [CrossRef]
- Deng, C.M.; Liu, S.X.; Huang, C.H.; Pang, J.Y.; Lin, Y.C. Secondary metabolites of a mangrove endophytic fungus Aspergillus terreus (No. GX7-3B) from the South China Sea. Mar. Drugs 2013, 11, 2616–2624. [Google Scholar] [CrossRef]
- Tao, Y.W.; Lin, Y.C.; She, Z.G.; Lin, M.T.; Chen, P.X.; Zhang, J.Y. Anticancer activity and mechanism investigation of beauvericin isolated from secondary metabolites of the mangrove endophytic fungi. Anti-Cancer Agents Med. Chem. 2015, 15, 258–266. [Google Scholar] [CrossRef]
- Amagata, T.; Morinaka, B.I.; Amagata, A.; Tenney, K.; Valeriote, F.A.; Lobkovsky, E.; Clardy, J.; Crews, P. A chemical study of cyclic depsipeptides produced by a sponge-derived fungus. J. Nat. Prod. 2006, 69, 1560–1565. [Google Scholar] [CrossRef]
- Krasnoff, S.B.; Gibson, D.M.; Belofsky, G.N.; Gloer, K.B.; Gloer, J.B. New destruxins from the entomopathogenic fungus Aschersonia sp. J. Nat. Prod. 1996, 59, 485–489. [Google Scholar] [CrossRef]
- Tsunoo, A.; Kamijo, M.; Taketomo, N.; Sato, Y.; Ajisaka, K. Roseocardin, a novel cardiotonic cyclodepsipeptide from Trichothecium roseum TT103. J. Antibiot. 1997, 50, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- Belofsky, G.N.; Jensen, P.R.; Fenical, W. Sansalvamide: A new cytotoxic cyclic depsipeptide produced by a marine fungus of the genus Fusarium. Tetrahedron Lett. 1999, 40, 2913–2916. [Google Scholar] [CrossRef]
- Cueto, M.; Jensen, P.R.; Fenical, W. N-Methylsansalvamide, a cytotoxic cyclic depsipeptide from a marine fungus of the genus Fusarium. Phytochemistry 2000, 55, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, C.W.; Park, S.Y.; Asolkar, R.N.; Kim, H.; Kim, G.J.; Oh, S.J.; Kim, Y.; Lee, E.Y.; Oh, D.C.; et al. Acremonamide, a cyclic pentadepsipeptide with wound-healing properties isolated from a marine-derived fungus of the genus Acremonium. J. Nat. Prod. 2021, 84, 2249–2255. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.C.; Jensen, P.R.; Fenical, W. Zygosporamide, a cytotoxic cyclic depsipeptide from the marine-derived fungus Zygosporium masonii. Tetrahedron Lett. 2006, 47, 8625–8628. [Google Scholar] [CrossRef]
- Kim, M.Y.; Sohn, J.H.; Ahn, J.S.; Oh, H. Alternaramide, a cyclic depsipeptide from the marine-derived fungus Alternaria sp. SF-5016. J. Nat. Prod. 2009, 72, 2065–2068. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.F.; Pan, W.; Ong, G.T.; Chiou, A.J.; Chuang, C.C.; Chiou, S.H.; Wu, S.H. Study of structure–activity correlation in destruxins, a class of cyclodepsipeptides possessing suppressive effect on the generation of hepatitis B virus surface antigen in human hepatoma cells. Biochem. Biophys. Res. Comm. 1996, 229, 65–72. [Google Scholar] [CrossRef]
- Yu, Z.; Lang, G.; Kajahn, I.; Schmaljohann, R.; Imhoff, J.F. Scopularides A and B, cyclodepsipeptides from a marine sponge-derived fungus, Scopulariopsis brevicaulis. J. Nat. Prod. 2008, 71, 1052–1054. [Google Scholar] [CrossRef]
- Singab, A.N.B.; Mostafa, N.M.; Elkhawas, Y.A.; Al-Sayed, E.; Bishr, M.M.; Elissawy, A.M.; Elnaggar, M.S.; Fawzy, I.M.; Salama, O.M.; Tsai, Y.H. Cyclodepsipeptides: Isolation from endophytic fungi of Sarcophyton ehrenbergi and verification of their larvicidal activity via in-vitro and in-silico studies. Mar. Drugs 2022, 20, 331. [Google Scholar] [CrossRef]
- Ding, L.J.; Yuan, W.; Liao, X.J.; Han, B.N.; Wang, S.P.; Li, Z.Y.; Xu, S.H.; Zhang, W.; Lin, H.W. Oryzamides A–E, cyclodepsipeptides from the sponge-derived fungus Nigrospora oryzae PF18. J. Nat. Prod. 2016, 79, 2045–2052. [Google Scholar] [CrossRef] [PubMed]
- Langenfeld, A.; Blond, A.; Gueye, S.; Herson, P.; Nay, B.; Dupont, J.; Prado, S. Insecticidal cyclodepsipeptides from Beauveria felina. J. Nat. Prod. 2011, 74, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Ravindra, G.; Ranganayaki, R.S.; Raghothama, S.; Srinivasan, M.C.; Gilardi, R.D.; Karle, I.L.; Balaram, P. Two novel hexadepsipeptides with several modified amino acid residues isolated from the fungus Isaria. Chem. Biodiver. 2004, 1, 489–504. [Google Scholar] [CrossRef] [PubMed]
- Sabareesh, V.; Ranganayaki, R.; Raghothama, S.; Bopanna, M.; Balaram, H.; Srinivasan, M.; Balaram, P. Identification and characterization of a library of microheterogeneous cyclohexadepsipeptides from the fungus Isaria. J. Nat. Prod. 2007, 70, 715–729. [Google Scholar] [CrossRef]
- Du, F.Y.; Zhang, P.; Li, X.M.; Li, C.S.; Cui, C.M.; Wang, B.G. Cyclohexadepsipeptides of the isaridin class from the marine-derived fungus Beauveria felina EN-135. J. Nat. Prod. 2014, 77, 1164–1169. [Google Scholar] [CrossRef]
- Wu, H.M.; Lin, L.P.; Xu, Q.L.; Han, W.B.; Zhang, S.; Liu, Z.W.; Mei, Y.N.; Yao, Z.J.; Tan, R.X. Nodupetide, a potent insecticide and antimicrobial from Nodulisporium sp. associated with Riptortus pedestris. Tetrahedron Lett. 2017, 58, 663–665. [Google Scholar] [CrossRef]
- Hou, X.M.; Li, Y.Y.; Shi, Y.W.; Fang, Y.W.; Chao, R.; Gu, Y.C.; Wang, C.Y.; Shao, C.L. Integrating molecular networking and 1H NMR to target the isolation of chrysogeamides from a library of marine-derived Penicillium fungi. J. Org. Chem. 2019, 84, 1228–1237. [Google Scholar] [CrossRef]
- Oh, D.C.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Induced production of emericellamides A and B from the marine-derived fungus Emericella sp. in competing co-culture. J. Nat. Prod. 2007, 70, 515–520. [Google Scholar] [CrossRef]
- Jiang, M.; Chen, S.; Lu, X.; Guo, H.; Chen, S.; Yin, X.; Li, H.; Dai, G.; Liu, L. Integrating genomics and metabolomics for the targeted discovery of new cyclopeptides with antifungal activity from a marine-derived fungus Beauveria felina. J. Agri. Food Chem. 2023, 71, 9782–9795. [Google Scholar] [CrossRef]
- Cruz, L.J.; Insua, M.M.; Baz, J.P.; Trujillo, M.; Rodriguez-Mias, R.A.; Oliveira, E.; Giralt, E.; Albericio, F.; Canedo, L.M. IB-01212, a new cytotoxic cyclodepsipeptide isolated from the marine fungus Clonostachys sp. ESNA-A009. J. Org. Chem. 2006, 71, 3335–3338. [Google Scholar] [CrossRef]
- Hegde, V.R.; Puar, M.S.; Chan, T.M.; Dai, P.; Das, P.R.; Patel, M. Sch 217048: A novel cyclodepsipeptide with neurokinin antagonist activity. J. Org. Chem. 1998, 63, 9584–9586. [Google Scholar] [CrossRef]
- Niu, S.; He, J.; Huang, S.; Wu, S.; Zeng, L.; Wang, J.; Hong, B.; Chen, Z. Phaeosphamides A and B, cytotoxic cyclodecadepsipeptides from the mangrove-derived fungus Phaeosphaeriopsis sp. S296. Mar. Drugs 2022, 20, 591. [Google Scholar] [CrossRef] [PubMed]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [PubMed]
- Luque-Ortega, J.R.; Cruz, L.J.; Albericio, F.; Rivas, L. The antitumoral depsipeptide IB-01212 kills Leishmania through an apoptosis-like process involving intracellular targets. Mol. Pharm. 2010, 7, 608–1617. [Google Scholar] [CrossRef]
- Dokmanovic, M.; Clarke, C.; Marks, P.A. Histone deacetylase inhibitors: Overview and perspectives. Mol. Cancer Res. 2007, 5, 981–989. [Google Scholar] [CrossRef]
- Birari, R.B.; Bhutani, K.K. Pancreatic lipase inhibitors from natural sources: Unexplored potential. Drug Discov. Today 2007, 12, 879–889. [Google Scholar] [CrossRef]
- Chiang, Y.J.; Hsiao, S.J.; Yver, D.; Cushman, S.W.; Tessarollo, L.; Smith, S.; Hodes, R.J. Tankyrase 1 and tankyrase 2 are essential but redundant for mouse embryonic development. PLoS ONE 2008, 3, e2639. [Google Scholar] [CrossRef]
- Johnson, T.O.; Ermolieff, J.; Jirousek, M.R. Protein tyrosine phosphatase 1B inhibitors for diabetes. Nat. Rev. Drug Discov. 2002, 1, 696–709. [Google Scholar] [CrossRef]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef]
- Blayney, D.W.; Zhang, Q.; Feng, J.; Zhao, Y.; Bondarenko, I.; Vynnychenko, I.; Kovalenko, N.; Nair, S.; Ibrahim, E.; Udovista, D.P.; et al. Efficacy of plinabulin vs pegfilgrastim for prevention of chemotherapy-induced neutropenia in adults with non–small cell lung cancer: A phase 2 randomized clinical trial. JAMA Oncol. 2020, 6, e204429. [Google Scholar] [CrossRef]
Figure 1.
FDA-approved marine peptide-based drugs [24].
Figure 1.
FDA-approved marine peptide-based drugs [24].

Figure 2.
Marine-derived peptides and their derivatives which are in clinical trials.

Figure 3.
Structures of 1–5.

Figure 4.
Structures of 6–28.

Figure 5.
Structures of 29–38.

Figure 6.
Structures of 39–42.

Figure 7.
Structures of 43–48.

Figure 8.
Structures of 49–54.

Figure 9.
Structures of 55 and 56.

Figure 10.
Structures of 57–66.

Figure 11.
Structures of 67–73.

Figure 12.
Structures of 74–78.

Figure 13.
Structures of 79–85.

Figure 14.
Structures of 86–140.

Figure 15.
Structures of 141–170.

Figure 16.
Structures of 171–191.

Figure 17.
Structures of 192–210.

Figure 18.
Structures of 211–224.

Figure 19.
Structures of 225–244.

Figure 20.
Structures of 245–268.

Figure 21.
Structures of 269–294.

Figure 22.
Structures of 295 and 296.

Figure 23.
Structures of 297–300.

Figure 24.
Structure of 301.

Figure 25.
Structures of 302–337.

Figure 26.
Structures of 338–366.

Figure 27.
The number of isolated peptides from marine-derived fungal resources.

Figure 28.
Mode of biological activities of peptides isolated from marine-derived fungal resources and their percentages.
Figure 28.
Mode of biological activities of peptides isolated from marine-derived fungal resources and their percentages.


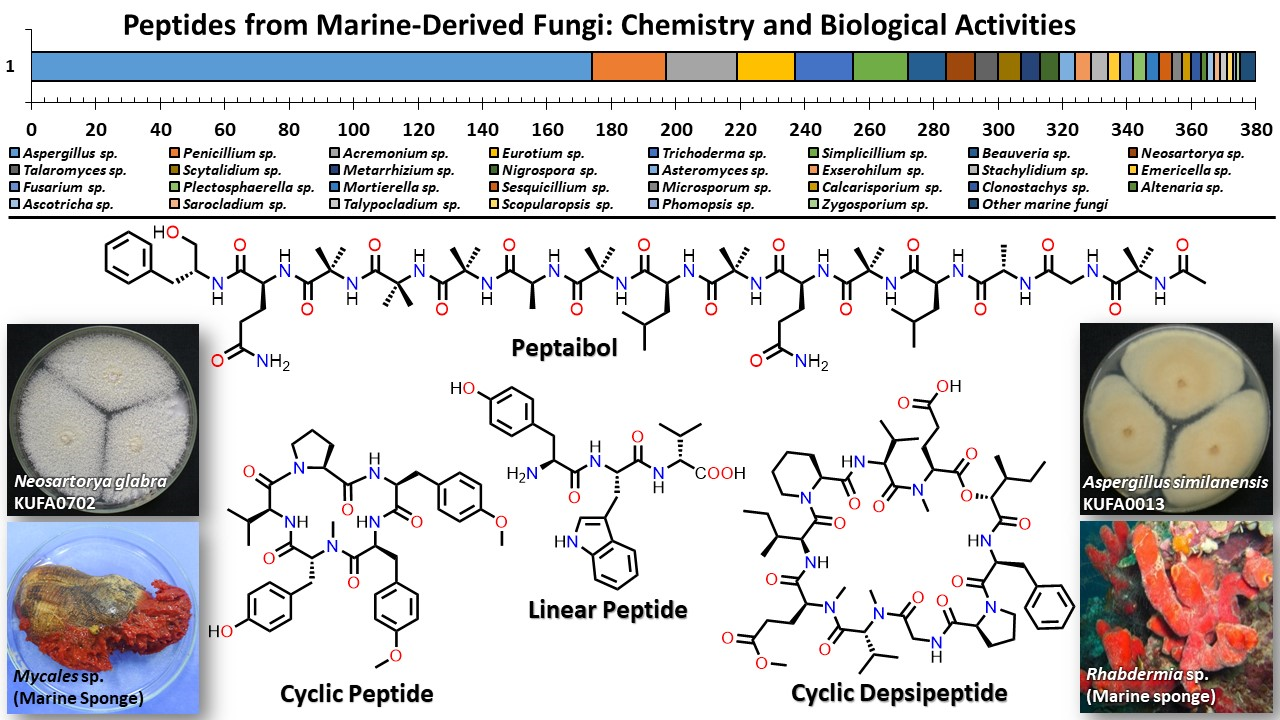{kind=link}
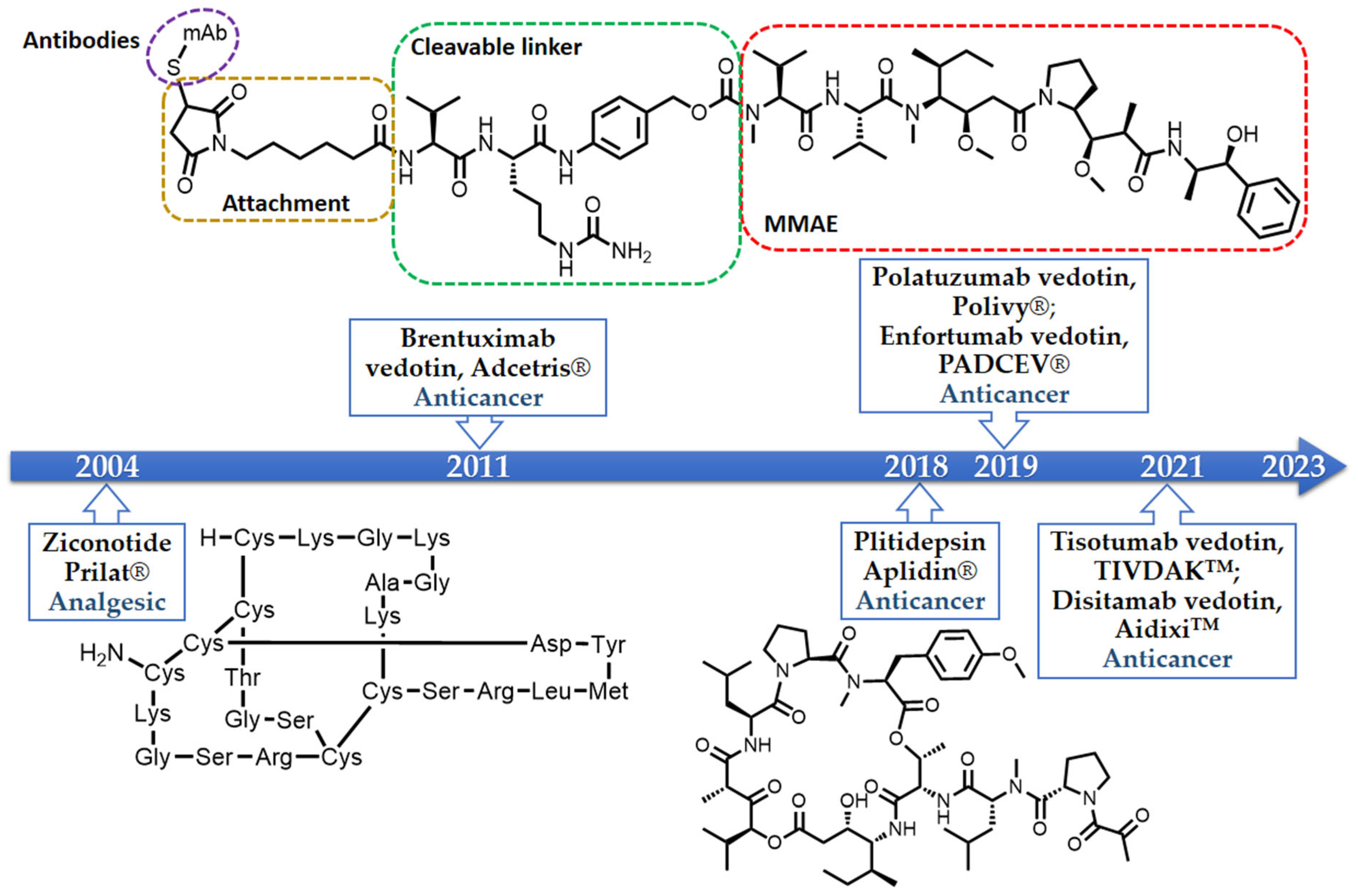{kind=link}
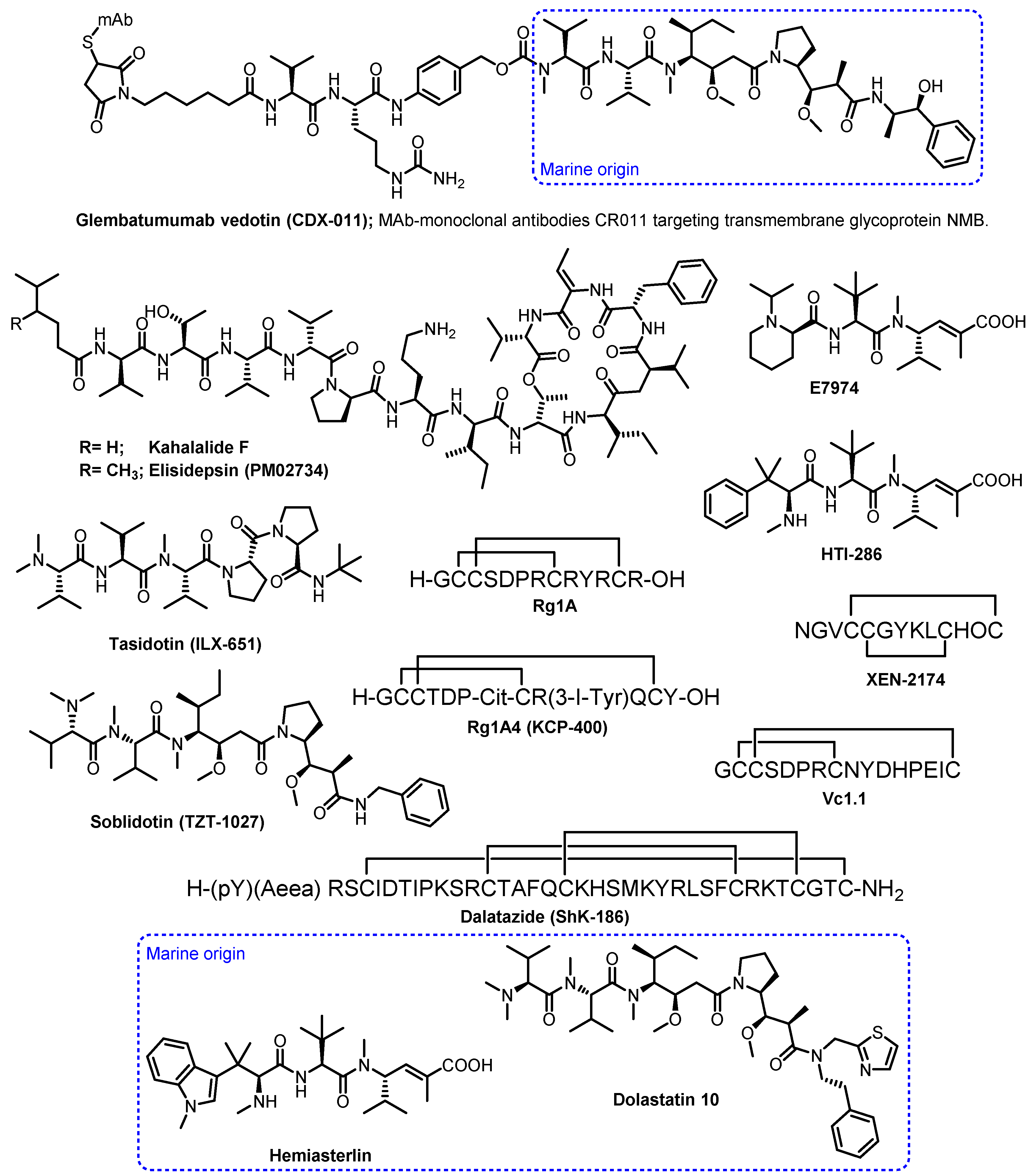{kind=link}
{kind=link}
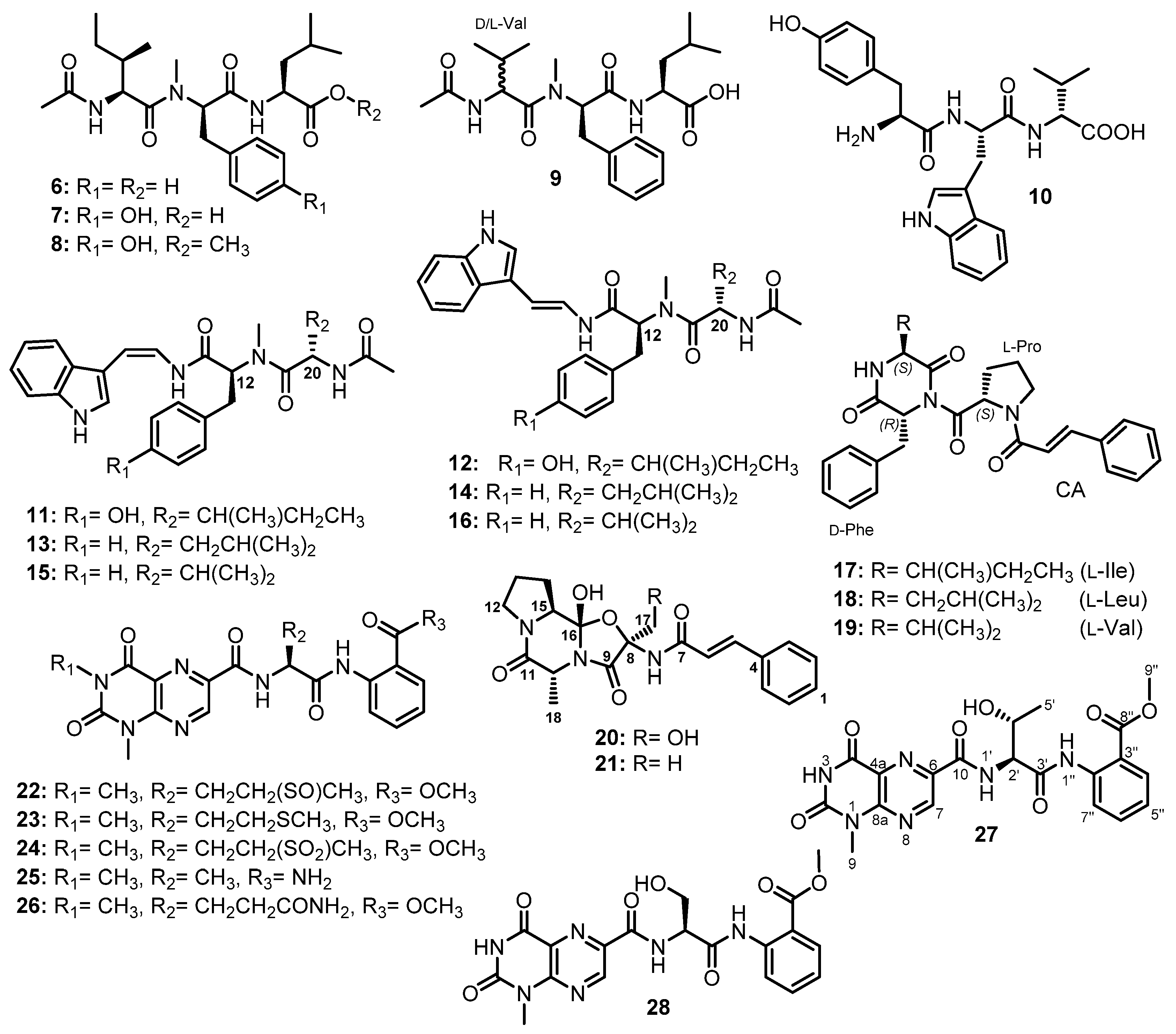{kind=link}
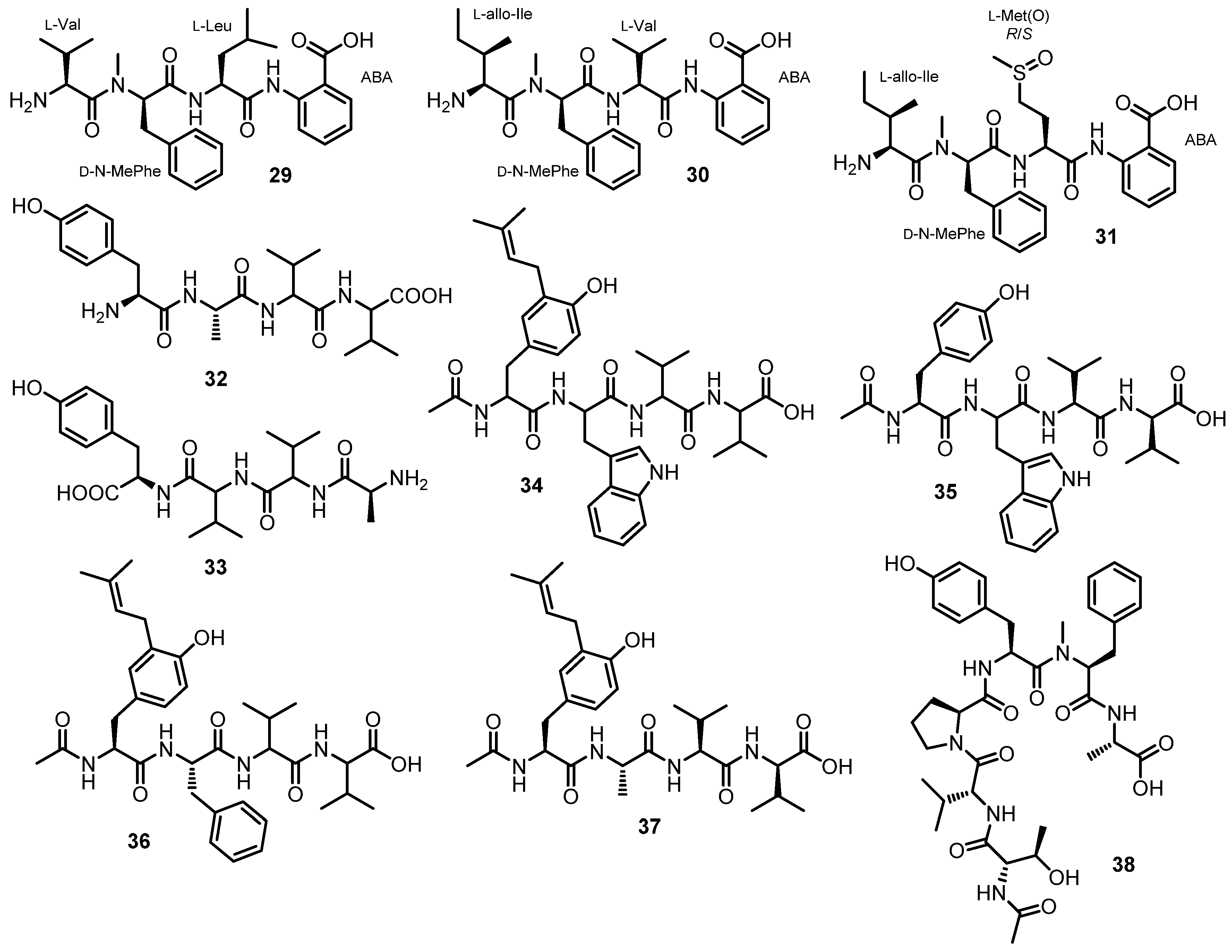{kind=link}
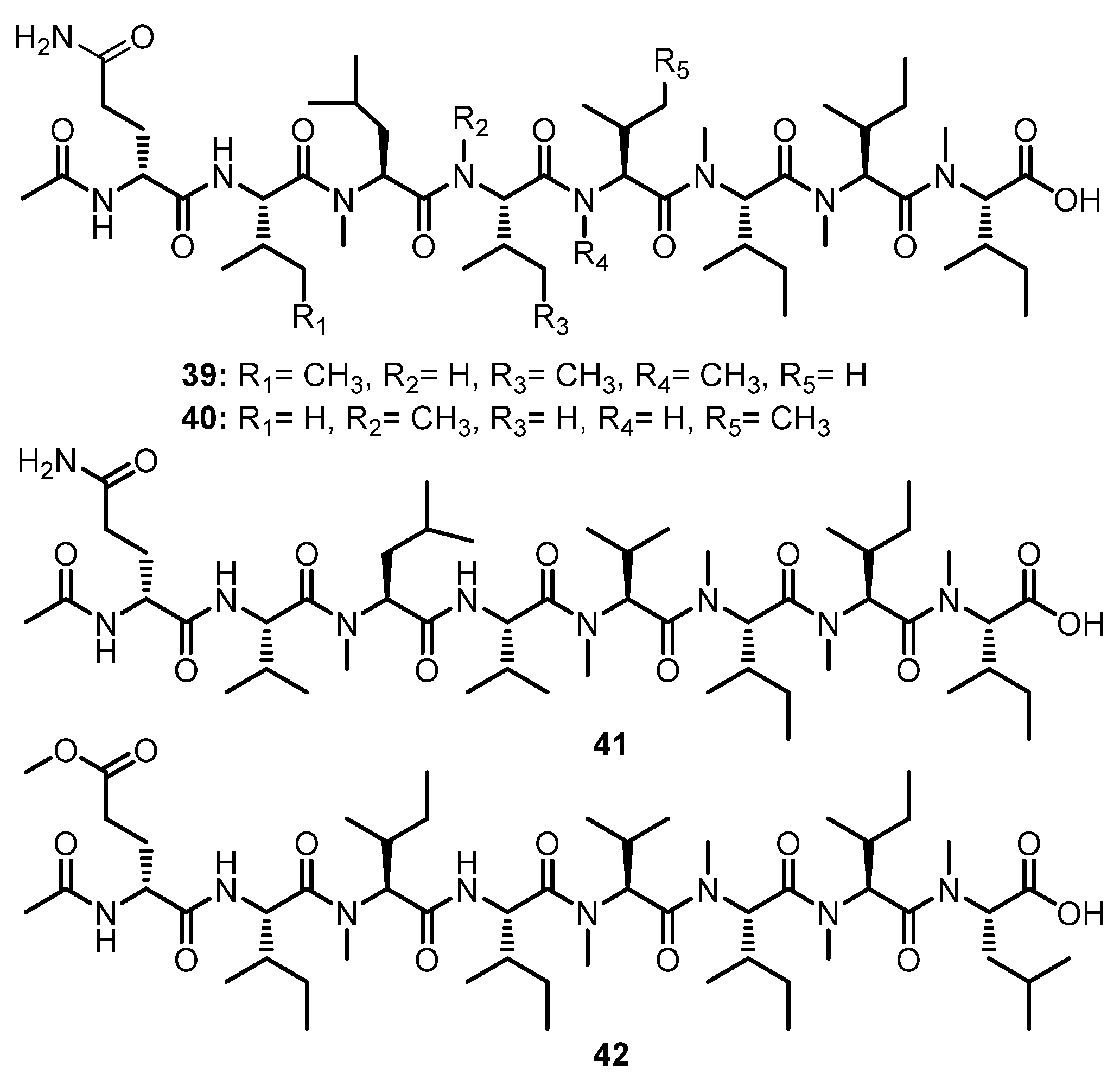{kind=link}
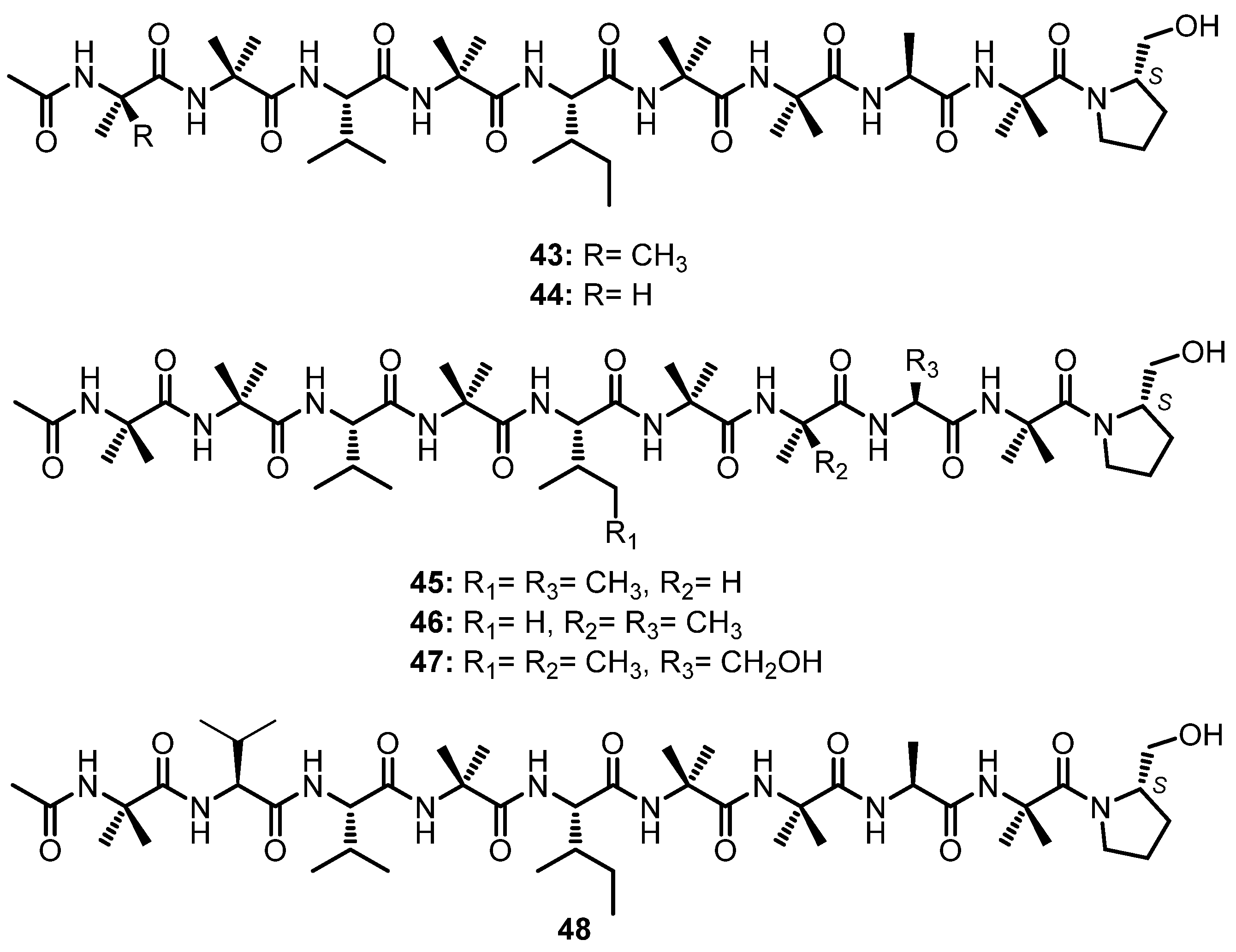{kind=link}
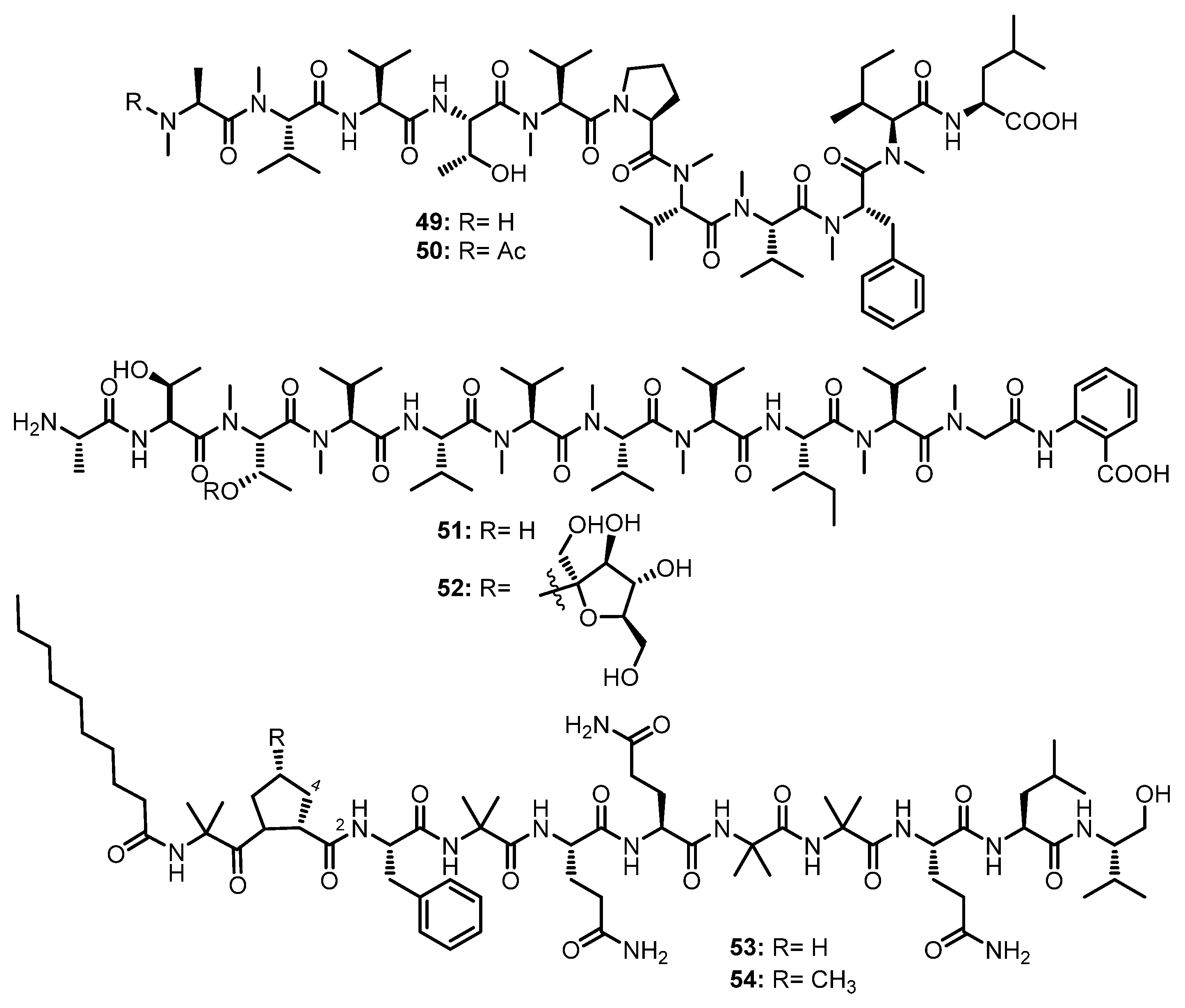{kind=link}
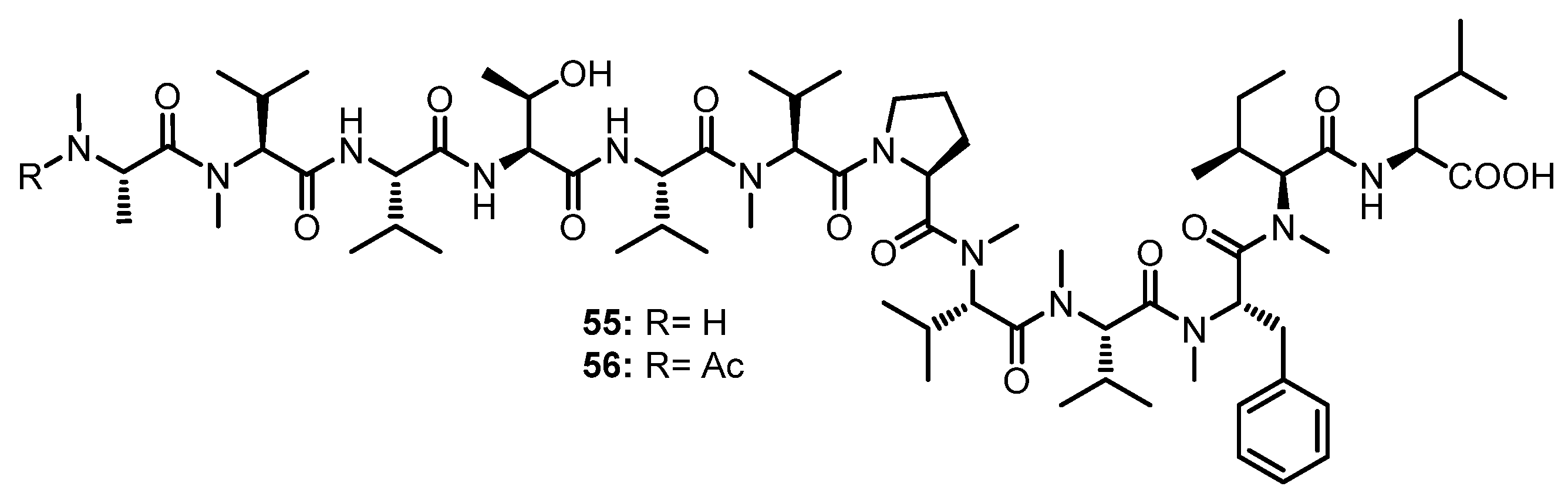{kind=link}
{kind=link}
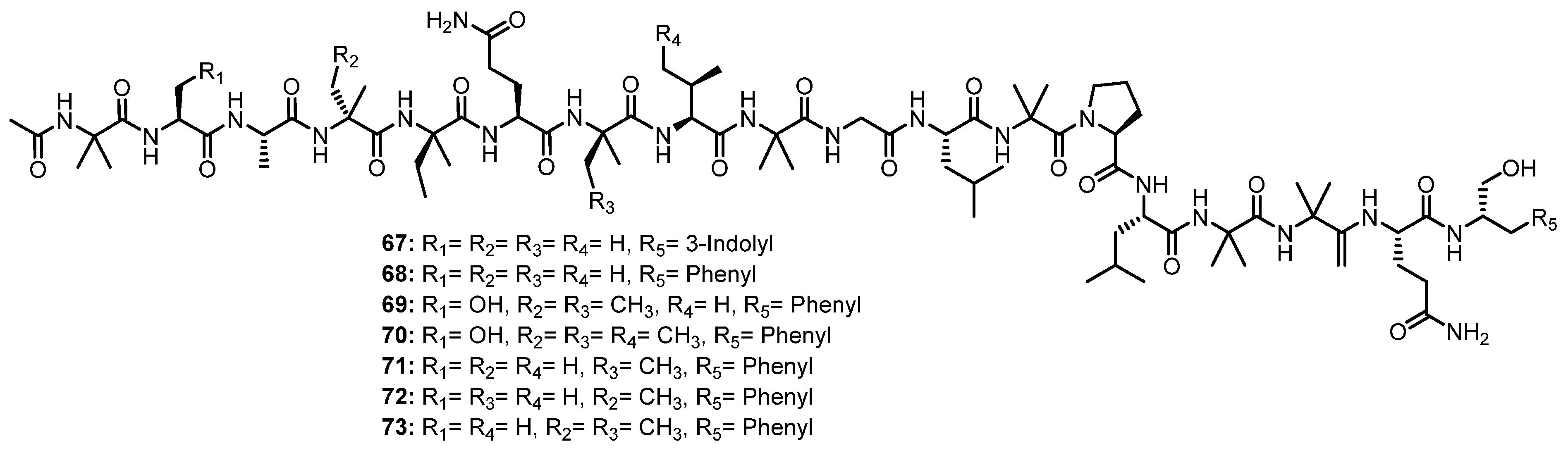{kind=link}
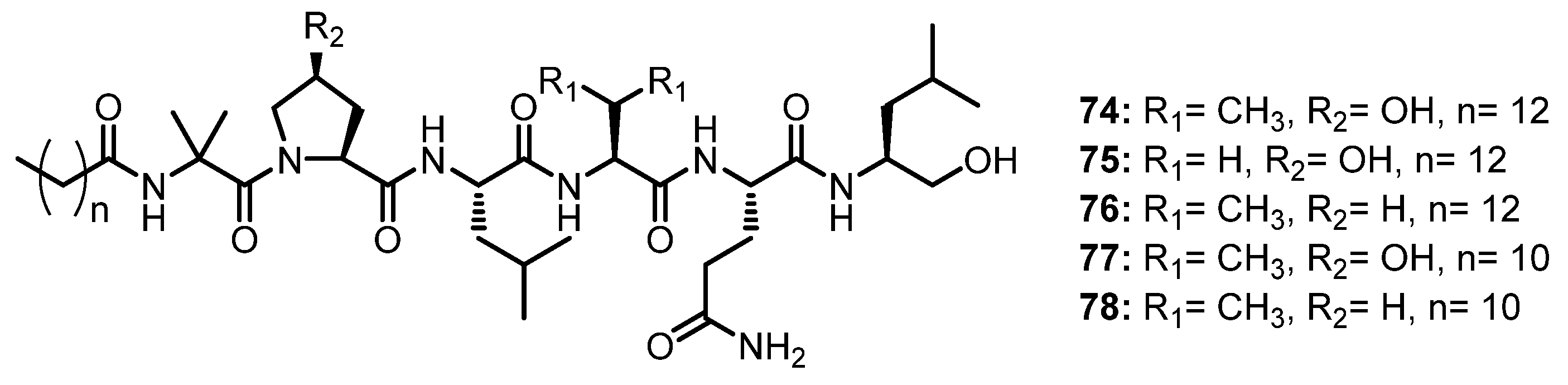{kind=link}
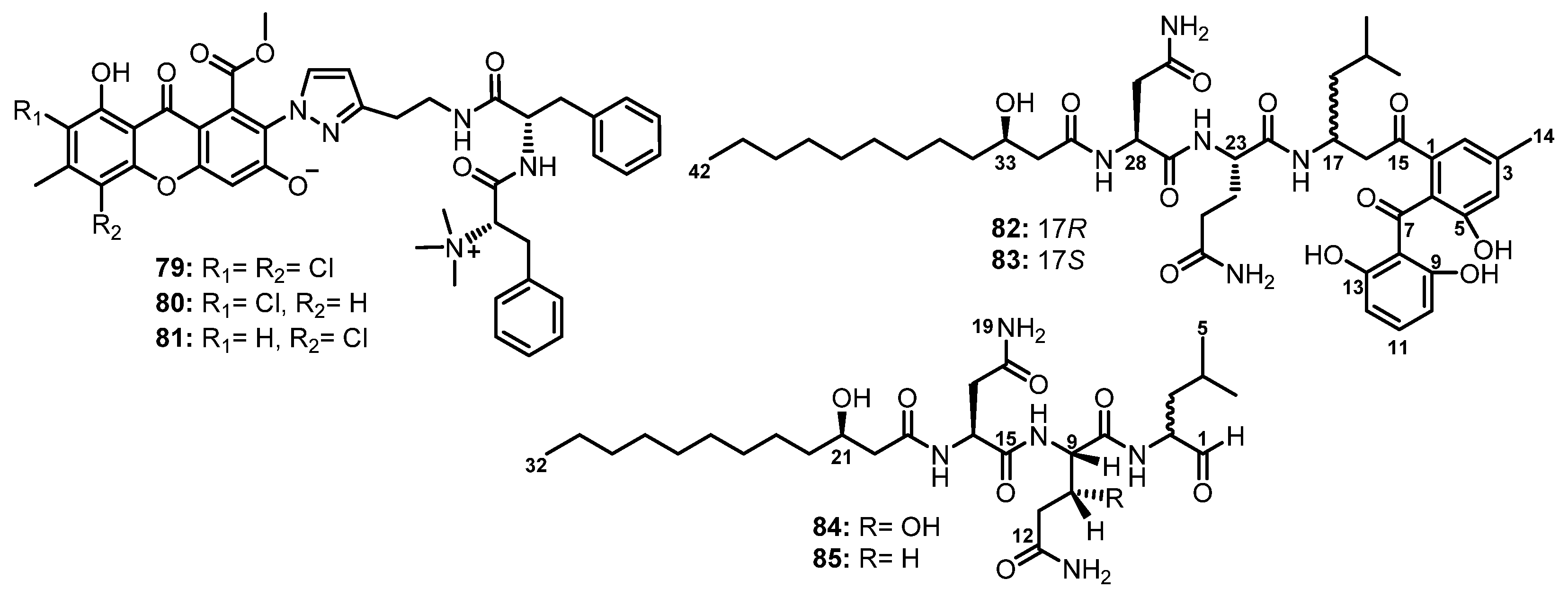{kind=link}
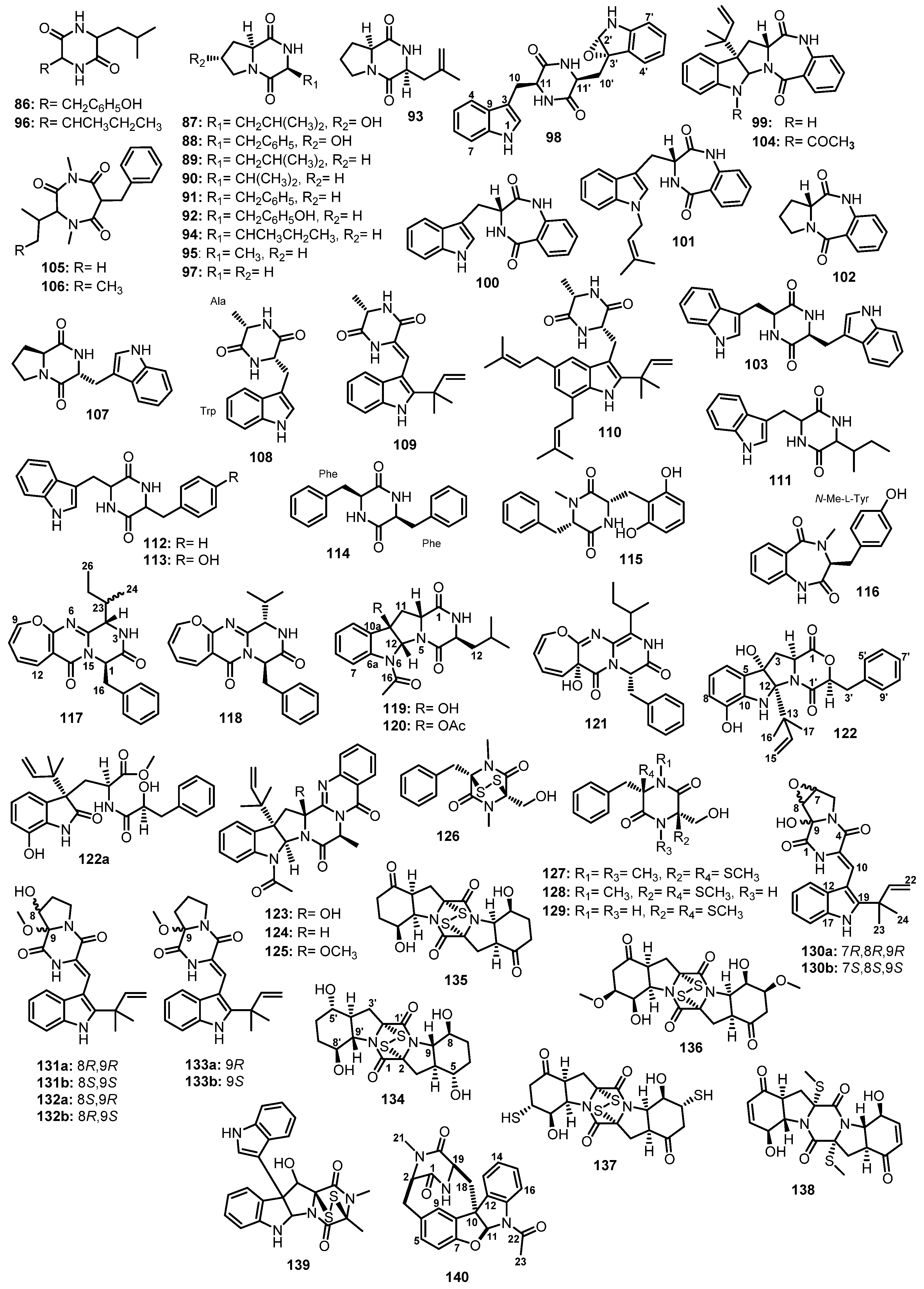{kind=link}
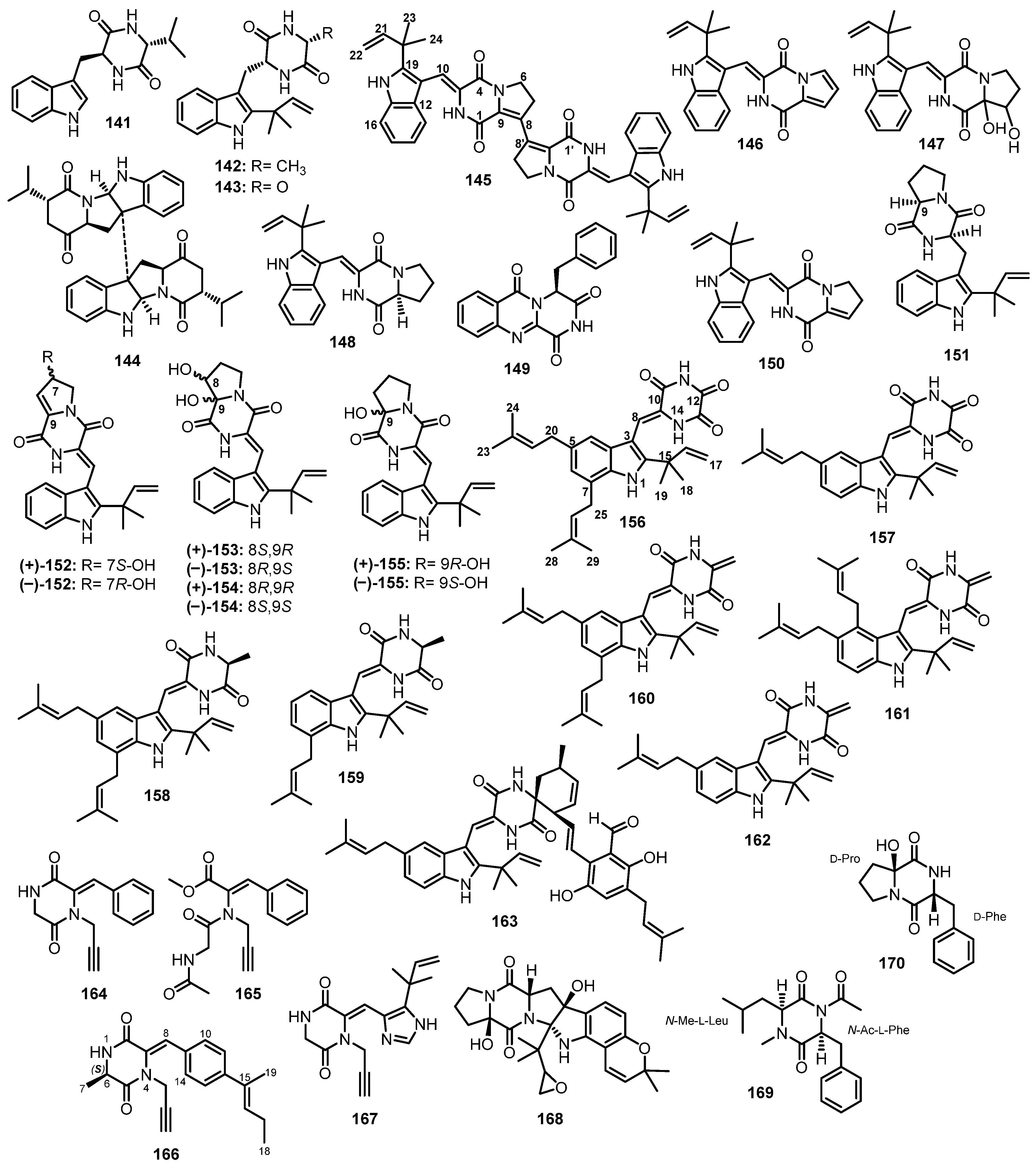{kind=link}
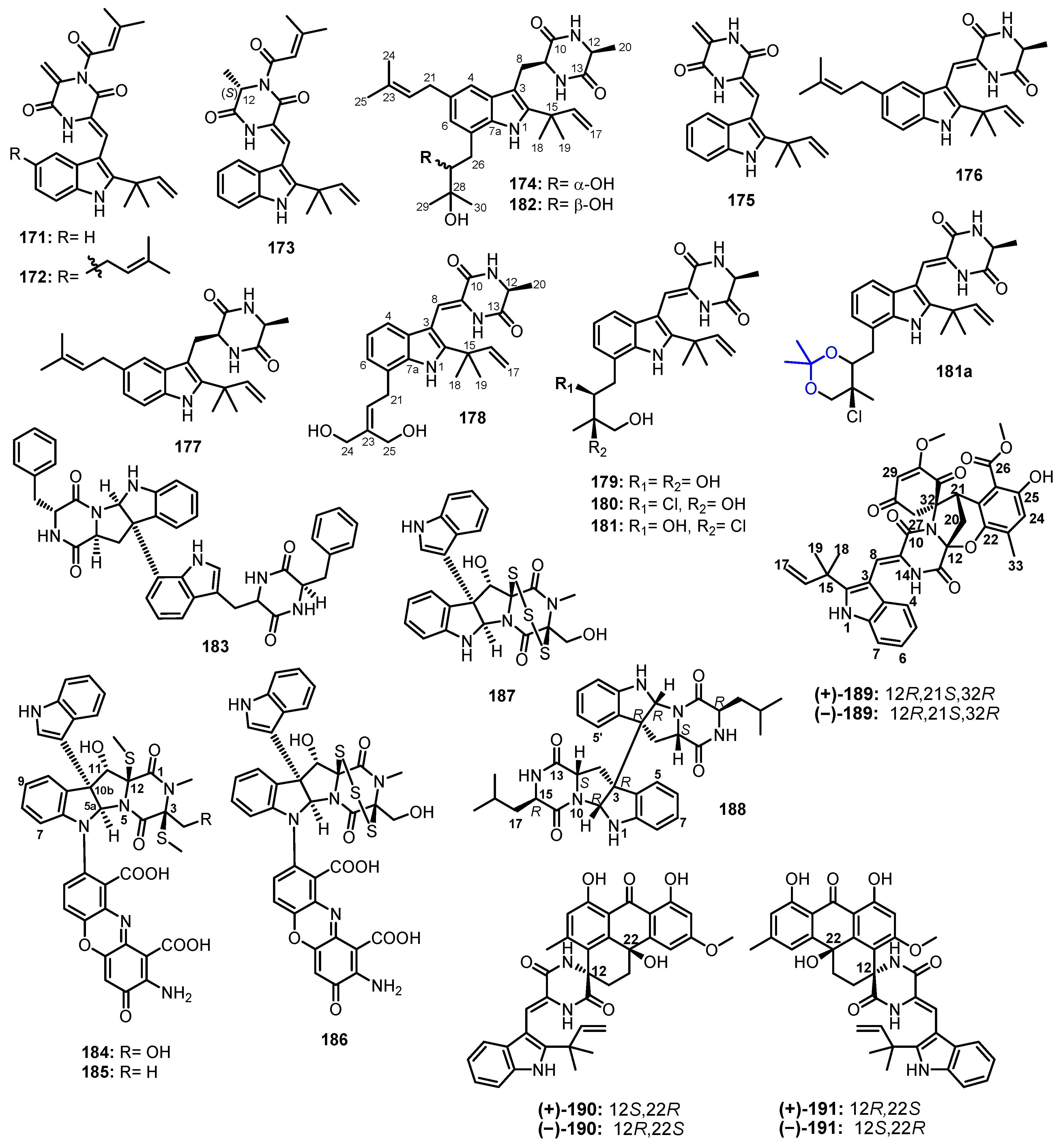{kind=link}
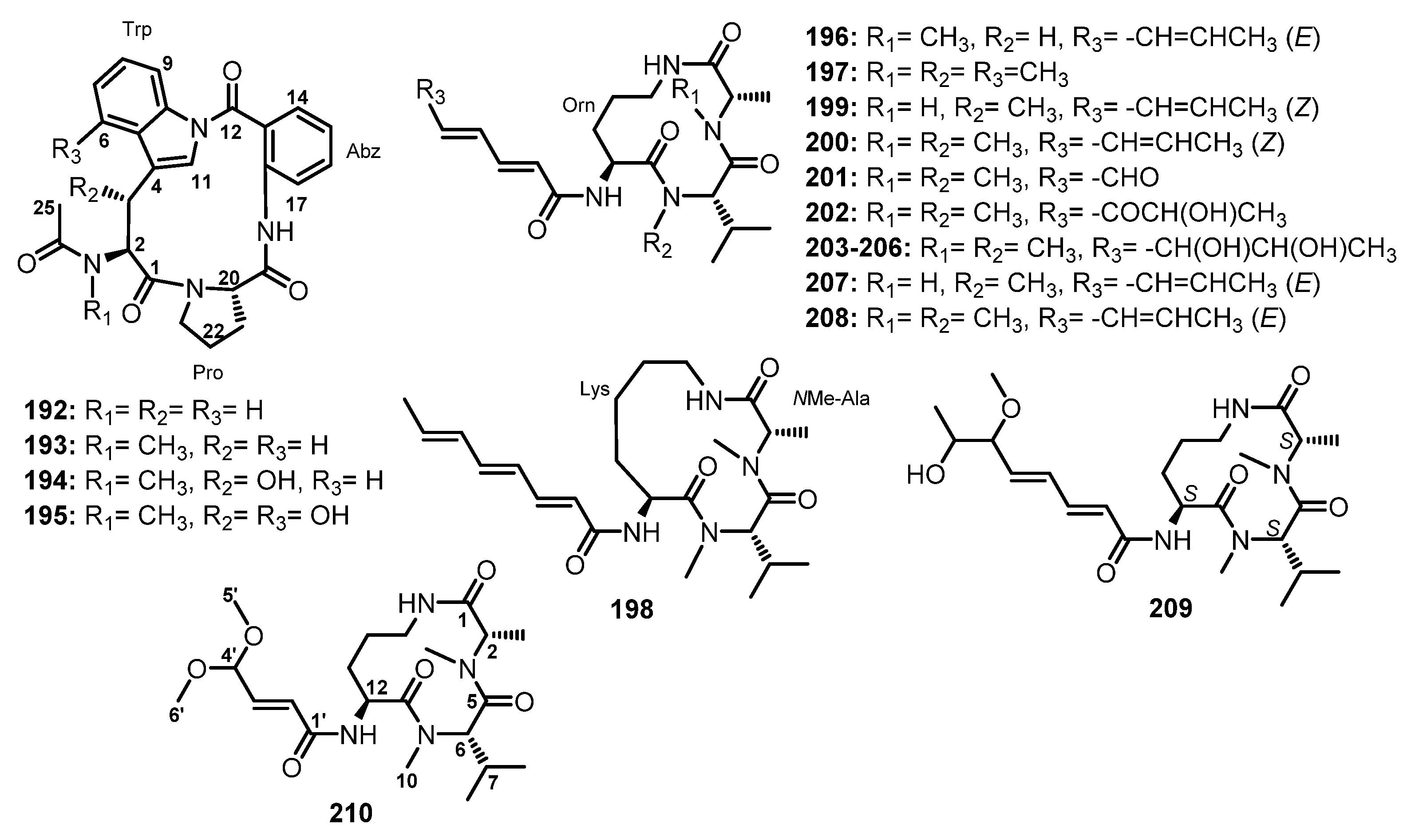{kind=link}
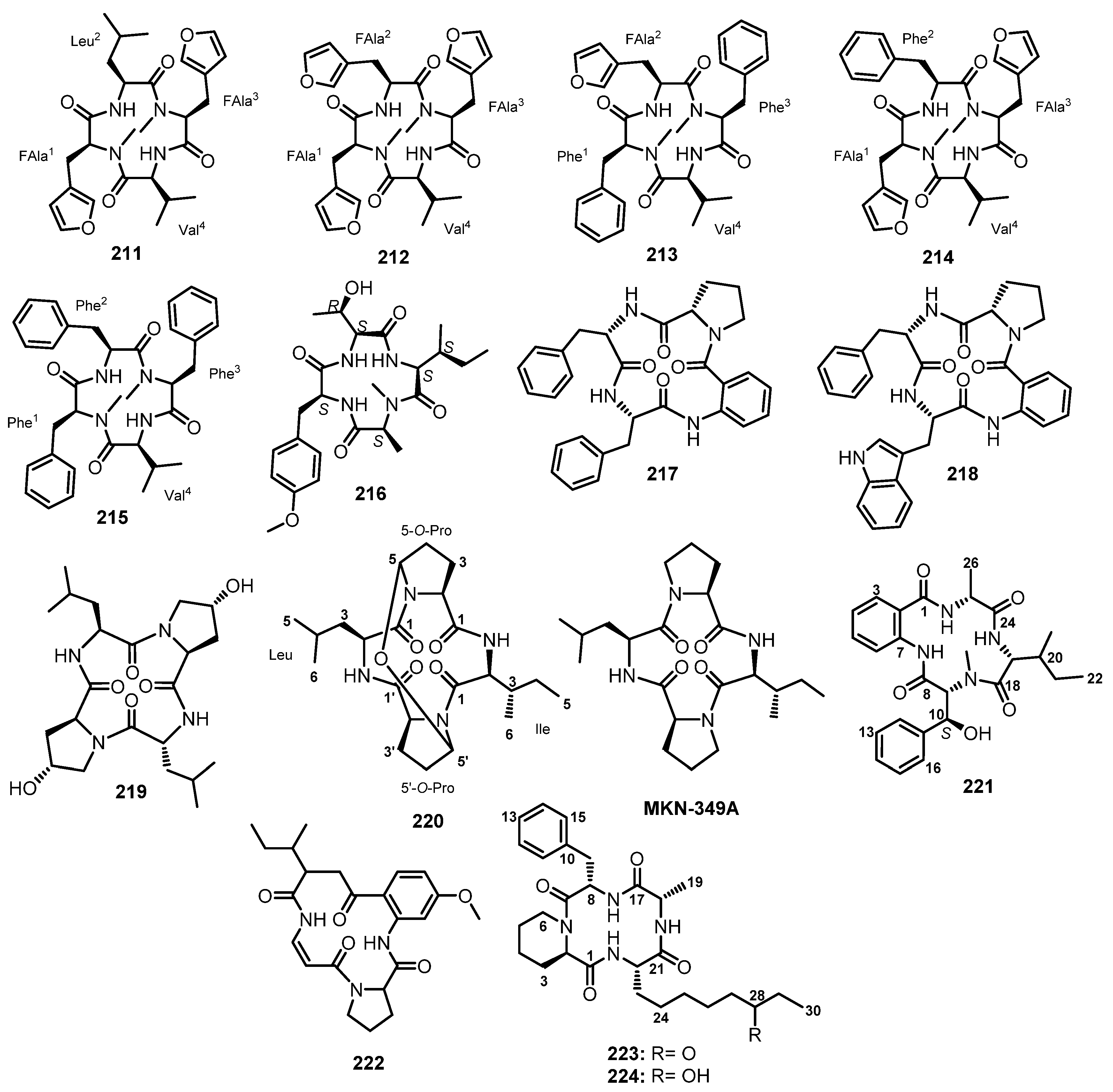{kind=link}
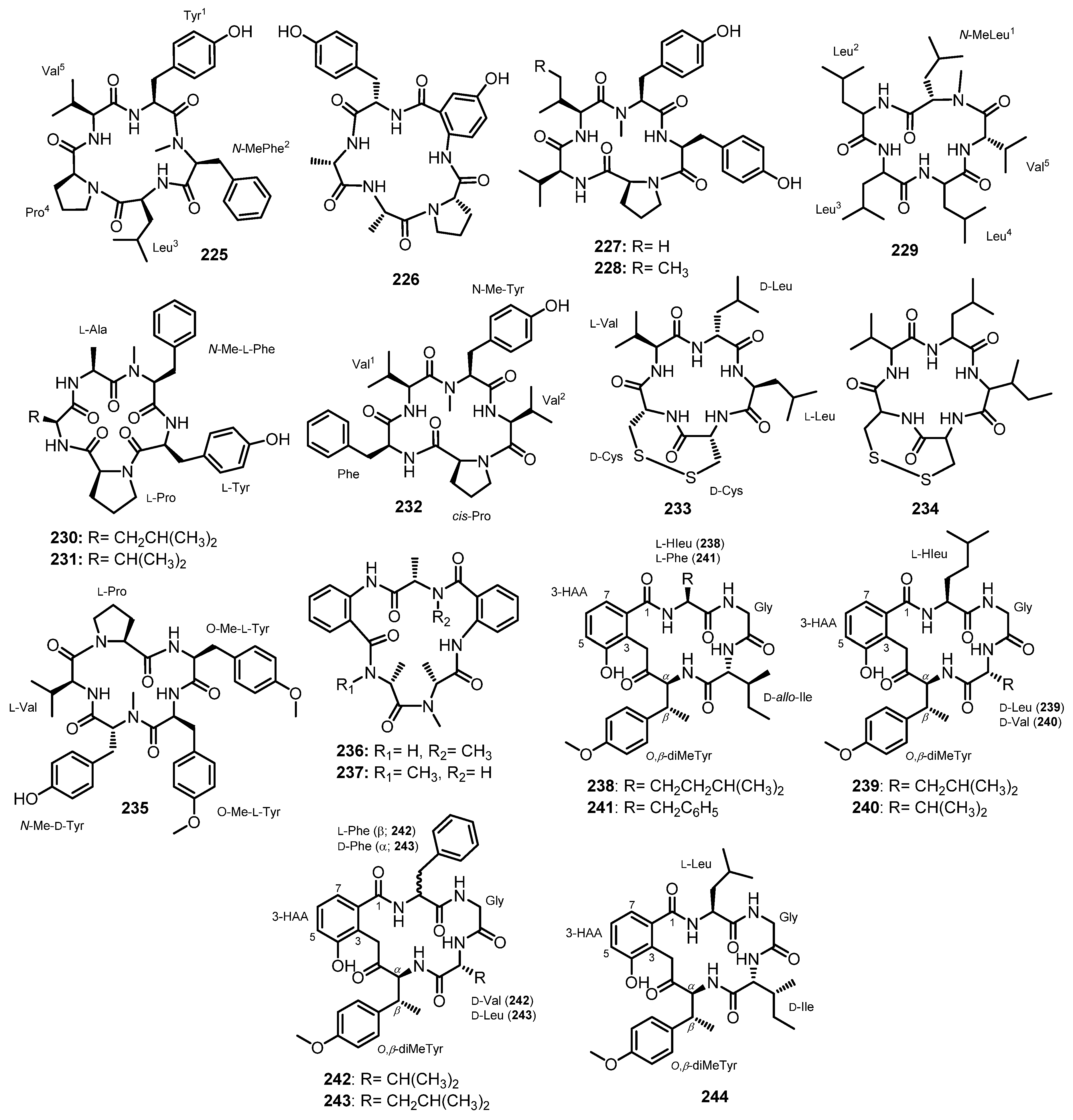{kind=link}
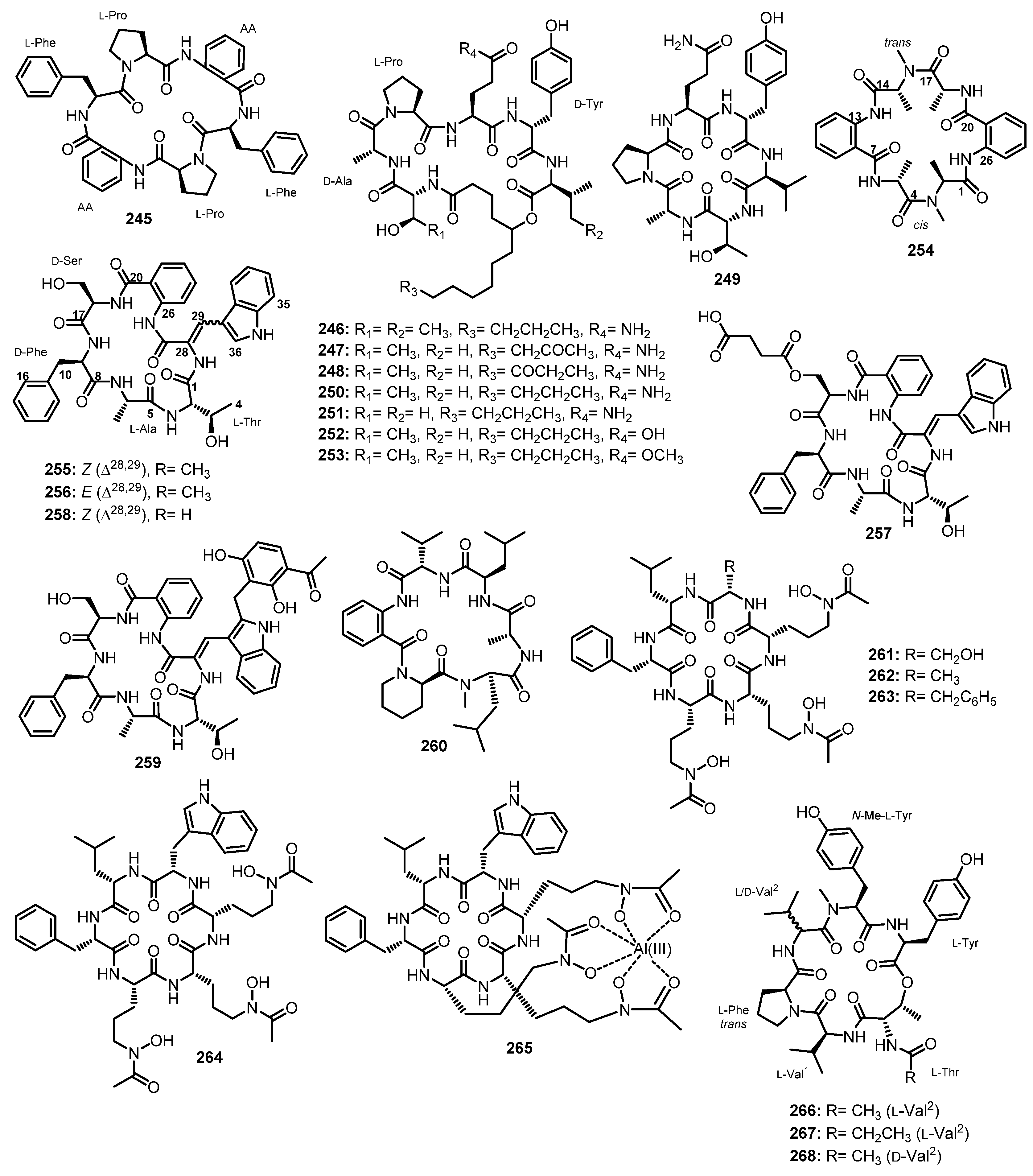{kind=link}
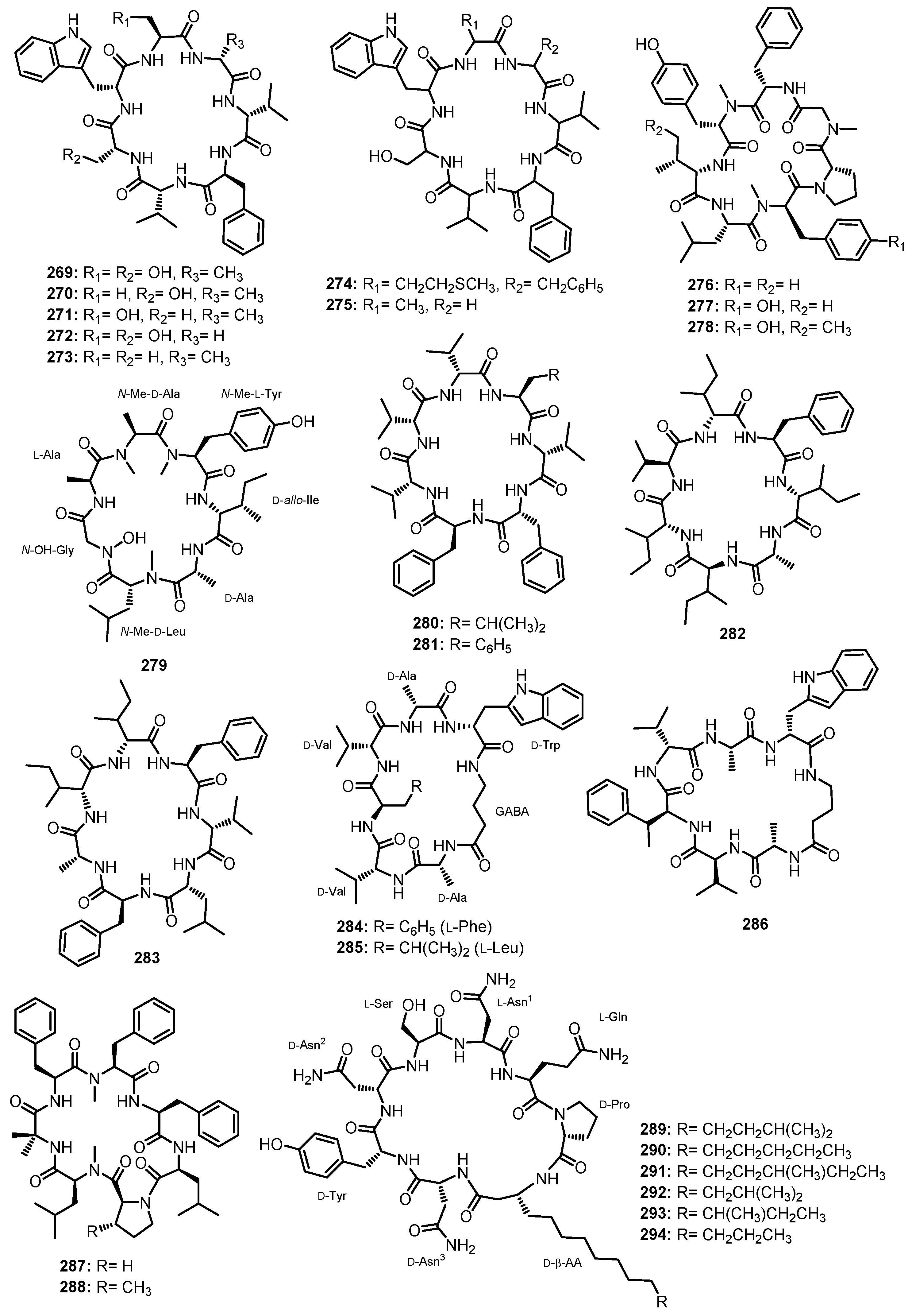{kind=link}
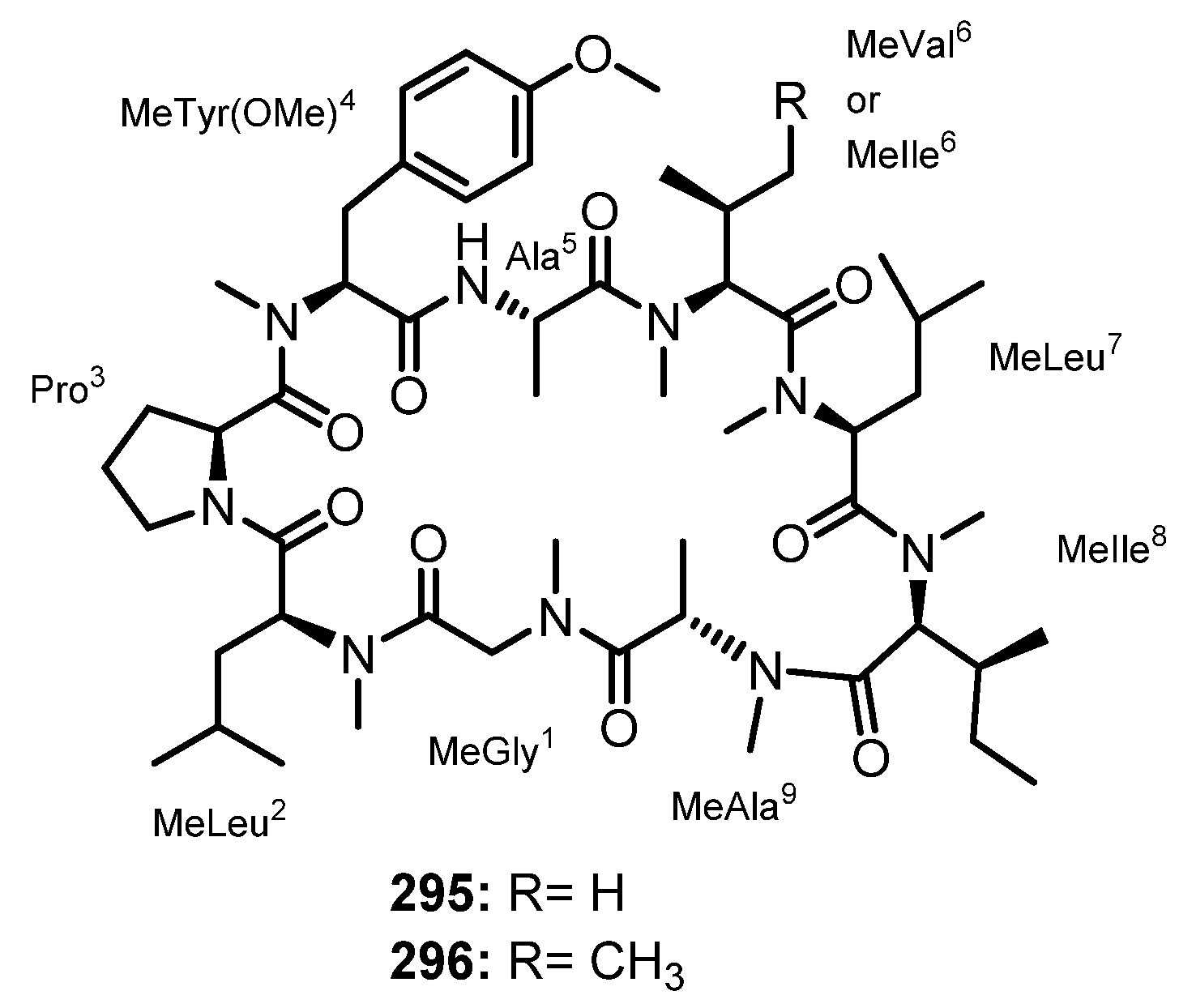{kind=link}
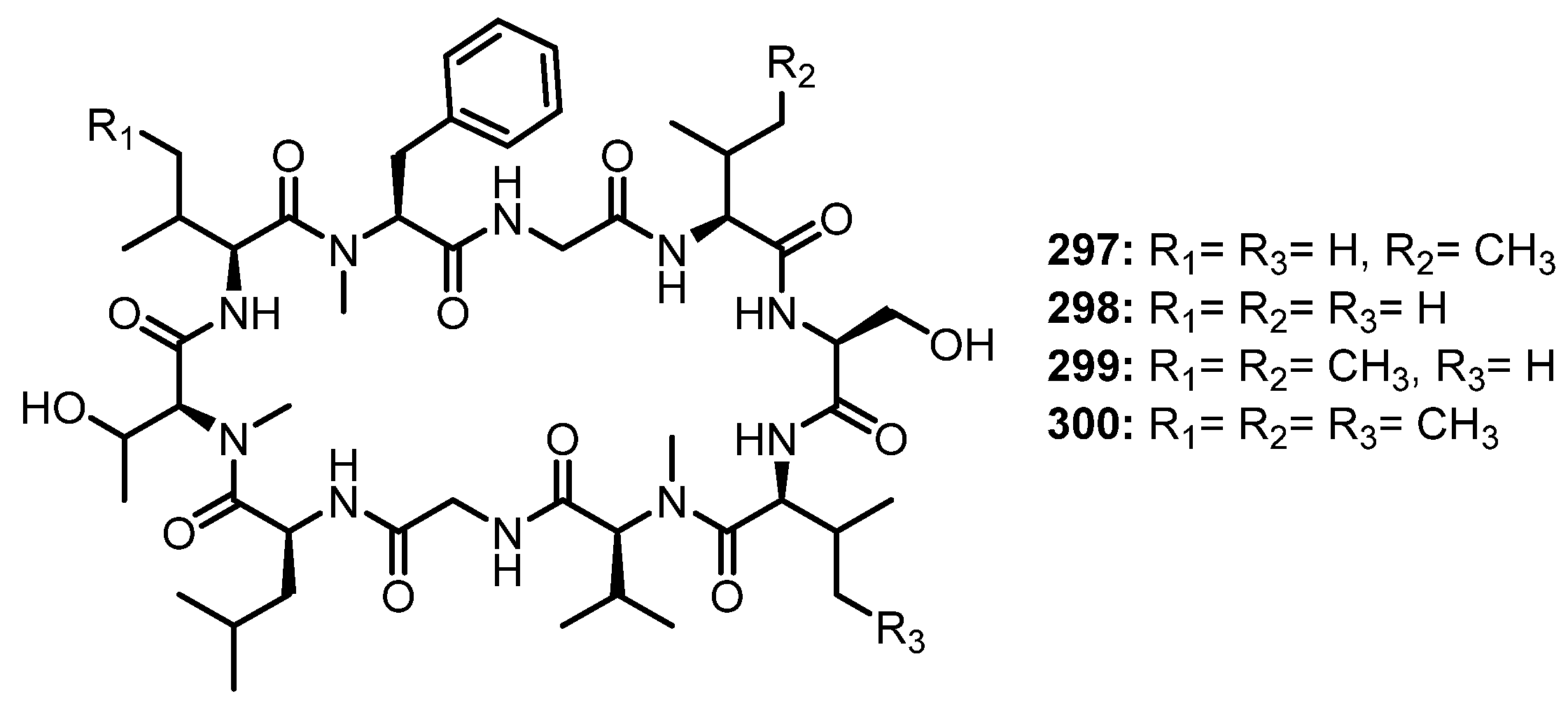{kind=link}
{kind=link}
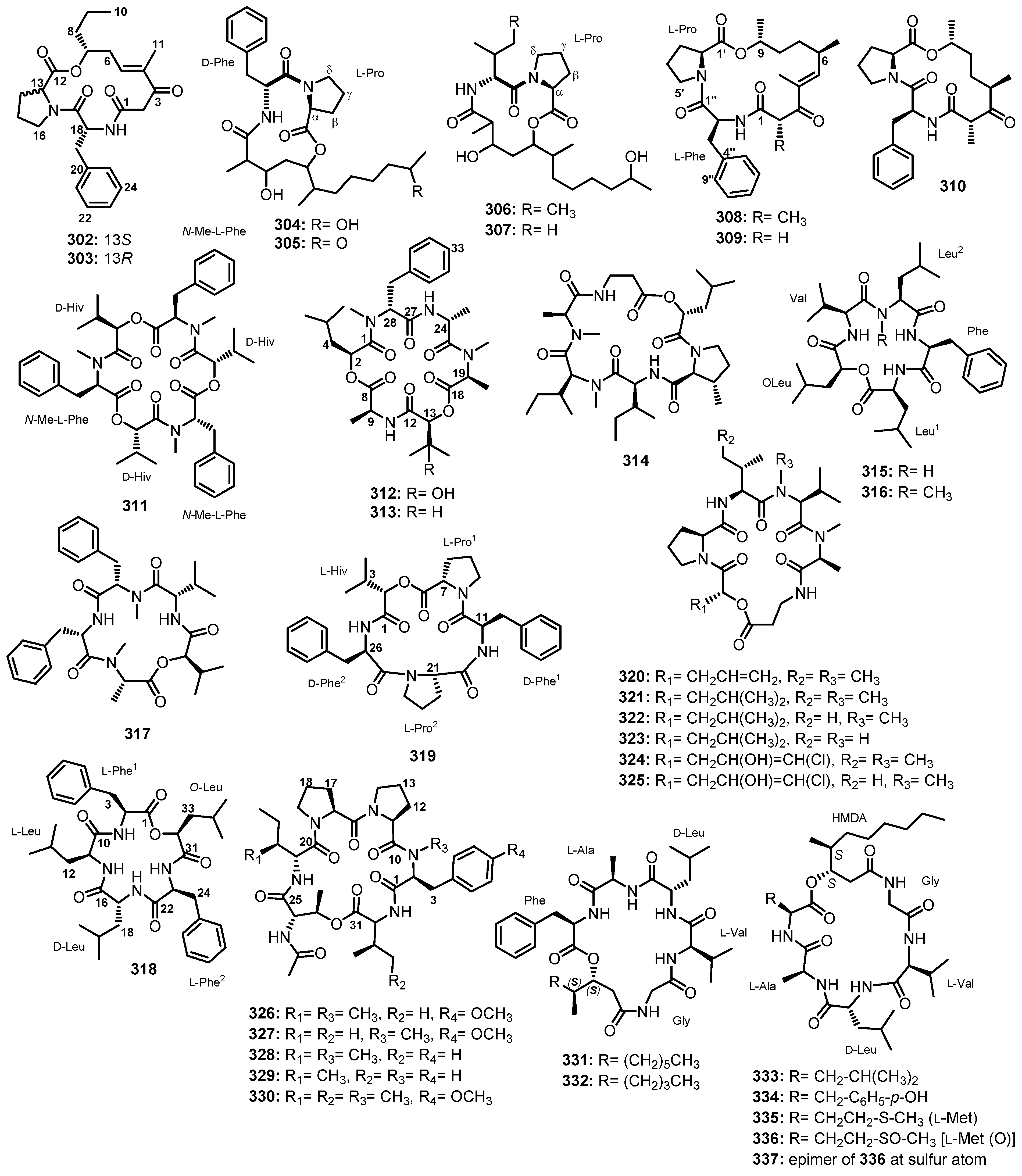{kind=link}
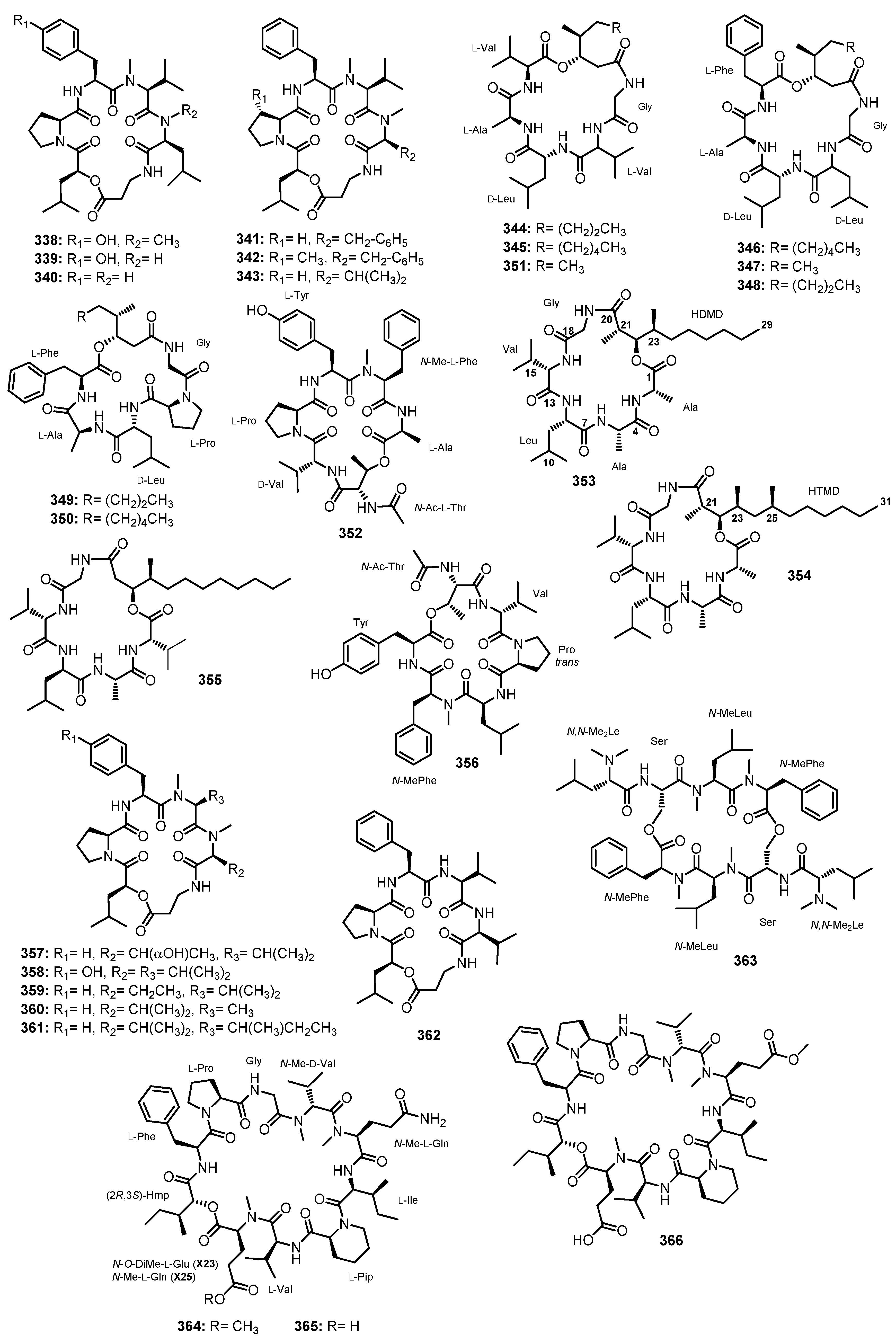{kind=link}
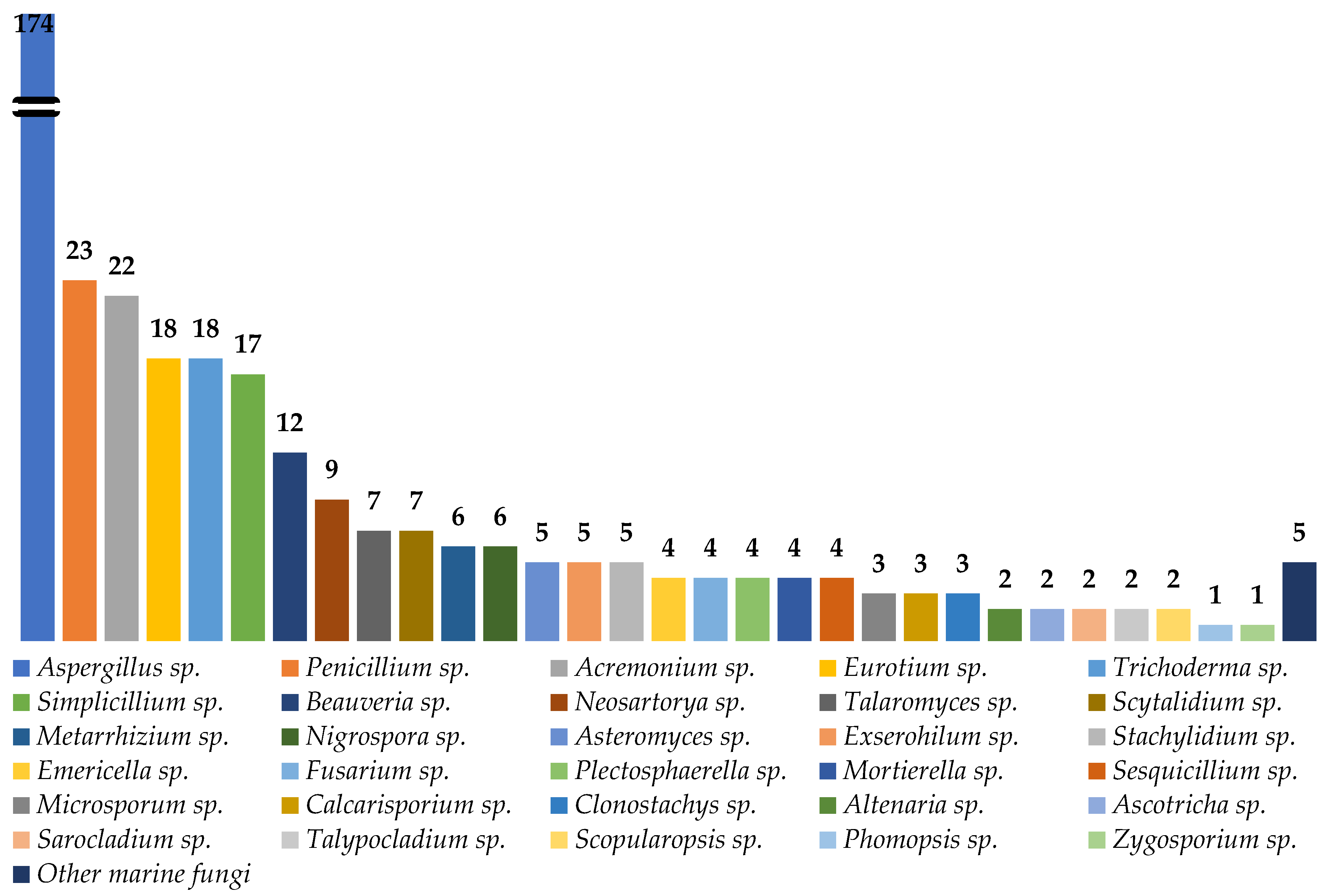{kind=link}
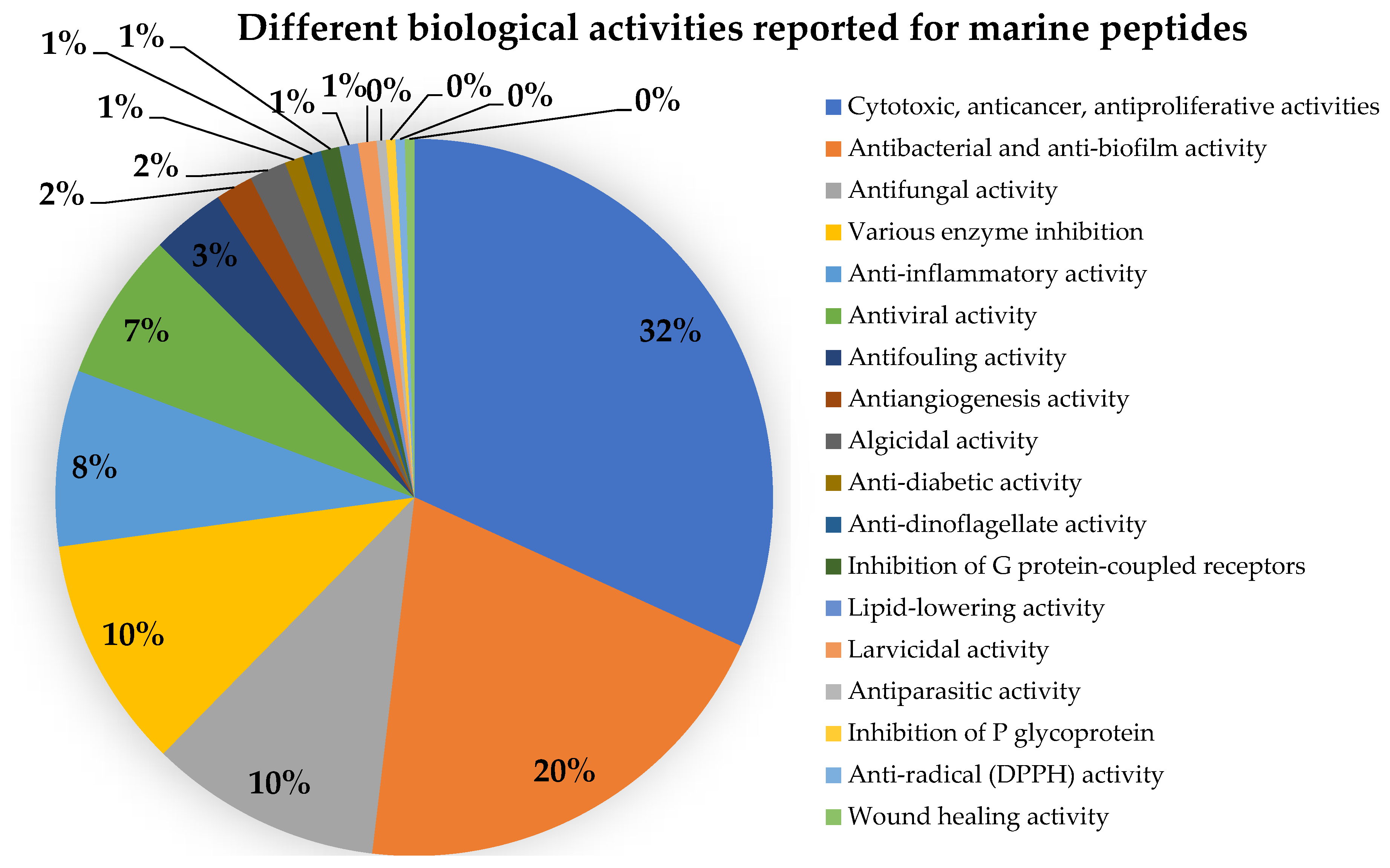{kind=link}
Table 1.
Peptides reported from marine-derived fungi from January 1991 to June 2023.
| Compound | Fungal Species/Strain No. | Source of Marine-Derived Fungi | Bioactivity | Ref. |
|---|---|---|---|---|
| Linear dipeptides | ||||
| Simplicilliumtide G (1) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifouling and cytotoxic activities. | [38] |
| Simplicilliumtide H (2) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifouling and cytotoxic activities. | [38] |
| Coniosulfide E (3) | Aspergillus unguis IV17-109 | Deep-sea shrimp. | - | [39] |
| Penicamide A (4) | Penicillium sp. SCSIO 41512 | Marine. | - | [40] |
| Penicamide B (5) | Penicillium sp. SCSIO 41512 | Marine. | - | [40] |
| Linear tripeptides | ||||
| Simplicilliumtide C (6) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifouling activity. | [38] |
| Simplicilliumtide D (7) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifouling activity. | [38] |
| Simplicilliumtide E (8) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifouling and cytotoxic activities. | [38] |
| Simplicilliumtide F (9) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifouling activity. | [38] |
| Aspergillipeptide E (10) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata. | Antiviral activity. | [41] |
| Aspergillamides C (11) | Aspergillus terreus SCSIO41008 | Marine sponge Callyspongia sp. | - | [42] |
| Aspergillamide D (12) | Aspergillus terreus SCSIO41008 | Marine sponge Callyspongia sp. | - | [42] |
| Aspergillamides A (13) | Aspergillus terreus SCSIO41008 | Marine sponge Callyspongia sp. | - | [42] |
| Aspergillamides B (14) | Aspergillus terreus SCSIO41008 | Marine sponge Callyspongia sp. | - | [42] |
| cis-L-phenylalaninamide (15) | Aspergillus terreus SCSIO41008 | Marine sponge Callyspongia sp. | - | [42] |
| trans-L-phenylalaninamide (16) | Aspergillus terreus SCSIO41008 | Marine sponge Callyspongia sp. | - | [42] |
| Asterripeptide A (17) | Aspergillus terreus LM.5.2 | Marine mangrove Kandelia candel. | Cytotoxic activity. | [43] |
| Asterripeptide B (18) | Aspergillus terreus LM.5.2 | Marine mangrove Kandelia candel. | Cytotoxic and inhibition of SrtA activities. | [43] |
| Asterripeptide C (19) | Aspergillus terreus LM.5.2 | Marine mangrove Kandelia candel. | Cytotoxic and inhibition of SrtA activities. | [43] |
| Talaropeptin A (20) | Talaromyces purpureogenus CX11 | Marine. | Antifungal activity. | [44] |
| Talaropeptin B (21) | Talaromyces purpureogenus CX11 | Marine. | Antifungal activity. | [44] |
| Penilumamide (22) | Penicillium sp. (strain CNL-338) | Marine red alga Laurencia sp. | - | [45] |
| Aspergillus sp. XS-20090B15 | Marine gorgonian Muricella abnormaliz. | - | [46] | |
| Aspergillus sp. (33241) | Marine mangrove Bruguiera sexangula var. rhynchopetala. | - | [47] | |
| Penilumamide B (23) | Aspergillus sp. XS-20090B15 | Marine gorgonian Muricella abnormaliz. | - | [46] |
| Penilumamide C (24) | Aspergillus sp. XS-20090B15 | Marine gorgonian Muricella abnormaliz. | - | [46] |
| Penilumamide D (25) | Aspergillus sp. XS-20090B15 | Marine gorgonian Muricella abnormaliz. | - | [46] |
| Aspergilumamide A (26) | Aspergillus sp. (33241) | Marine mangrove Bruguiera sexangula var. rhynchopetala. | - | [47] |
| Terrelumamide A (27) | Aspergillus terreus FA009 | Marine sediment. | Anti-diabetic and anticancer activities. | [48] |
| Terrelumamide B (28) | Aspergillus terreus FA009 | Marine sediment. | Anti-diabetic and anticancer activities. | [48] |
| Linear tetrapeptides | ||||
| Simplicilliumtide A (29) | Simplicillium obclavatum EIODSF 020 | Marine sediment. | Antifouling and cytotoxic activities. | [38] |
| Simplicilliumtide B (30) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifouling activity. | [38] |
| Simplicilliumtide I (31) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | - | [49] |
| Aspergillipeptide F (32) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata. | - | [41] |
| Aspergillipeptide G (33) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata. | - | [41] |
| Aspergillipeptide H (34) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata Nutting. | - | [50] |
| Aspergillipeptide I (35) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata Nutting. | - | [50] |
| Aspergillipeptide J (36) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata Nutting. | - | [50] |
| Aspergillipeptide K (37) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata Nutting | - | [50] |
| Linear hexapeptides | ||||
| FJ120DPB (38) | Aspergillus ochraceopetaliformis. | Marine sediment. | Inhibition of enzyme sortase A (SrtA) activity. | [51] |
| Linear octapeptides | ||||
| RHM1 (39) | Acremonium sp. (UCSC coll. no. 021172 cKZ) | Marine sponge Teichaxinella sp. (coll. no. 02172). | Antibacterial and Cytotoxic activities. | [52] |
| RHM2 (40) | Acremonium sp. (UCSC coll. no. 021172 cKZ) | Marine sponge Teichaxinella sp. (coll. no. 02172). | Cytotoxic activity. | [52] |
| RHM3 (41) | Acremonium sp. (UCSC coll. no. 021172 cKZ) | Marine sponge Teichaxinella sp. (coll. no. 02172). | - | [53] |
| RHM4 (42) | Acremonium sp. (UCSC coll. no. 021172 cKZ) | Marine sponge Teichaxinella sp. (coll. no. 02172). | - | [53] |
| Linear nonapeptides | ||||
| Aspereline A (43) | Trichoderma asperellum | Marine sediment. | - | [54] |
| Aspereline B (44) | Trichoderma asperellum | Marine sediment. | - | [54] |
| Aspereline C (45) | Trichoderma asperellum | Marine sediment. | - | [54] |
| Aspereline D (46) | Trichoderma asperellum | Marine sediment. | - | [54] |
| Aspereline E (47) | Trichoderma asperellum | Marine sediment. | - | [54] |
| Aspereline F (48) | Trichoderma asperellum | Marine sediment. | - | [54] |
| Linear undecapeptides | ||||
| Talaropeptide A (49) | Talaromyces sp. (CMB-TU011) | An unidentified marine tunicate. | Antibacterial activity. | [55] |
| Talaropeptide C (50) | Talaromyces sp. (CMB-TU011) | An unidentified marine tunicate. | - | [55] |
| Dictyonamide A (51) | The fungus strain K063. | Marine red alga Ceratodictyon spongiosum. | Inhibition of cyclin-dependent kinase 4 activity. | [56] |
| Dictyonamide B (52) | The fungus strain K063. | Marine red alga Ceratodictyon spongiosum. | - | [57] |
| Tolypocaibol A (53) | Tolypocladium sp. | Marine microalga Spongomorpha arcta. | Antibacterial activity. | [57] |
| Tolypocaibol B (54) | Tolypocladium sp. | Marine microalga Spongomorpha arcta. | Antibacterial activity. | [57] |
| Linear dodecapeptides | ||||
| Talaropeptide B (55) | Talaromyces sp. (CMB-TU011) | An unidentified marine tunicate. | Antibacterial activity. | [55] |
| Talaropeptide D (56) | Talaromyces sp. (CMB-TU011) | An unidentified marine tunicate. | - | [55] |
| Linear pentadecapeptides | ||||
| Efrapeptin G (57) | Acremonium sp. (UCSC coll. no. 021172 cKZ) | Marine sponge Teichaxinella sp. (coll. no. 02172). | Antibacterial and Cytotoxic activities. | [52,53] |
| Efrapeptins Eα (58) | Acremonium sp. (UCSC coll. no. 021172 cKZ) | Marine sponge Teichaxinella sp. (coll. no. 02172). | Cytotoxic activity. | [53] |
| Efrapeptin H (59) | Acremonium sp. (UCSC coll. no. 021172 cKZ) | Marine sponge Teichaxinella sp. (coll. no. 02172). | - | [53] |
| Efrapeptin E (60) | Acremonium sp. (UCSC coll. no. 021172 cKZ) | Marine sponge Teichaxinella sp. (coll. no. 02172). | - | [53] |
| Efrapeptin F (61) | Acremonium sp. (UCSC coll. no. 021172 cKZ) | Marine sponge Teichaxinella sp. (coll. no. 02172). | Cytotoxic activity. | [53] |
| Pentadecaibin I (62) | Trichoderma sp. MMS1255 | Marine sediment. | Antibacterial and cytotoxic activities. | [58] |
| Pentadecaibin II (63) | Trichoderma sp. MMS1255 | Marine sediment. | Cytotoxic activity. | [58] |
| Pentadecaibin III (64) | Trichoderma sp. MMS1255 | Marine sediment. | Antibacterial and cytotoxic activities. | [58] |
| Pentadecaibin IV (65) | Trichoderma sp. MMS1255 | Marine sediment. | Cytotoxic activity. | [58] |
| Pentadecaibin V (66) | Trichoderma sp. MMS1255 | Marine sediment. | Antibacterial and cytotoxic activities. | [58] |
| Linear octadecapeptides | ||||
| Trichorzins A (67) | Trichoderma sp. GXIMD 01001 | Marine sponge Haliclona sp. | Antibacterial and cytotoxic activities. | [59] |
| Trichorzin B (68) | Trichoderma sp. GXIMD 01001 | Marine sponge Haliclona sp. | Antibacterial and cytotoxic activities. | [59] |
| Trichorzin C (69) | Trichoderma sp. GXIMD 01001 | Marine sponge Haliclona sp. | Antibacterial and cytotoxic activities. | [59] |
| Trichorzin D (70) | Trichoderma sp. GXIMD 01001 | Marine sponge Haliclona sp. | Antibacterial and cytotoxic activities. | [59] |
| Trichorzin E (71) | Trichoderma sp. GXIMD 01001 | Marine sponge Haliclona sp. | Antibacterial and cytotoxic activities. | [59] |
| Trichorzin F (72) | Trichoderma sp. GXIMD 01001 | Marine sponge Haliclona sp. | Antibacterial and cytotoxic activities. | [59] |
| Trichorzin G (73) | Trichoderma sp. GXIMD 01001 | Marine sponge Haliclona sp. | Antibacterial and cytotoxic activities. | [59] |
| Lipophilic linear hexapeptides | ||||
| Halovir A (74) | Scytalidium sp. | Marine. | Antiviral activity. | [60] |
| Halovir B (75) | Scytalidium sp. | Marine. | Antiviral activity. | [60] |
| Halovir C (76) | Scytalidium sp. | Marine. | Antiviral activity. | [60] |
| Halovir D (77) | Scytalidium sp. | Marine. | Antiviral activity. | [60] |
| Halovir E (78) | Scytalidium sp. | Marine. | Antiviral activity. | [60] |
| Other linear peptides | ||||
| Flavipeside A (79) | Aspergillus flavipes 164013 | Marine sponge Dysidea sp. | Inhibition of pancreatic lipase. | [61] |
| Flavipeside B (80) | Aspergillus flavipes 164013 | Marine sponge Dysidea sp. | Inhibition of pancreatic lipase. | [61] |
| Flavipeside C (81) | Aspergillus flavipes 164013 | Marine sponge Dysidea sp. | Inhibition of pancreatic lipase. | [61] |
| Asperphenin A (82) | Aspergillus sp. | Marine-submerged decaying wood. | Antiproliferative activity. | [62] |
| Asperphenin B (83) | Aspergillus sp. | Marine-submerged decaying wood. | Antiproliferative activity. | [62] |
| Fellutamide A (84) | Penicillium fellutanum Biourge | Marine fish Apogon endekataenta Bleeker. | Cytotoxic activity. | [63] |
| Fellutamide B (85) | Penicillium fellutanum Biourge | Marine fish Apogon endekataenta Bleeker. | Cytotoxic activity. | [63] |
| Cyclic dipeptides | ||||
| cis-cyclo(Leucyl-Tyrosyl) (86) | Penicillium sp. F37 | Marine sponge Axinella corrugate | Anti-biofilm formation. | [64] |
| cyclo-(trans-4-hydroxy-L-Pro-L-Leu) (87) | Aspergillus niger BRF-074 | Marine sediment. | - | [65] |
| cyclo-(trans-4-hydroxy-L-Pro-L-Phe) (88) | Aspergillus niger BRF-074 | Marine sediment. | - | [65] |
| cyclo-(L-Pro-L-Leu) (89) | Aspergillus niger BRF-074 | Marine sediment. | - | [65] |
| Fusarium sp. RWS56-10 | Marine hydrothermal vent sediment. | - | [66] | |
| cyclo-(L-Pro-L-Val) (90) | Aspergillus niger BRF-074 | Marine sediment. | - | [65] |
| cyclo-(Phe-Pro) (91) | Penicillium sp. WF-06 | Marine sediment. | - | [86] |
| Aspergillus terreus SCSIO41008 | Marine sponge Callyspongia sp. | - | [42] | |
| Aspergillus niger BRF-074 | Marine sediment. | - | [65] | |
| cyclo-(L-Pro-L-Tyr) (92) | Aspergillus ochraceopetaliformis DSW-2 | Sea water. | Weak cytotoxic activity. | [78] |
| Aspergillus niger BRF-074 | Marine sediment. | - | [65] | |
| Penicillium chrysogenum DXY-1 | Marine sediment. | Antibacterial activity and anti-biofilm formation | [67] | |
| Penicimutide (93) | Penicillium purpurogenum G59 | Marine sediment. | Cytotoxic activity. | [68] |
| cyclo-(L-Ile-L-Pro) (94) | Penicillium purpurogenum G59 | Marine sediment. | - | [68] |
| cyclo-(Pro-Ala) (95) | Ascotricha sp. ZJ-M-5 | Marine. | - | [69] |
| cyclo-(Ile-Leu) (96) | Ascotricha sp. ZJ-M-5 | Marine. | - | [69] |
| cyclo-(Gly-Pro) (97) | Penicillium sp. WF-06 | Marine sediment. | - | [86] |
| Fellutanine A analogue (98) | Neosartorya glabra KUFA 0702 | Marine sponge Mycale sp. | - | [70] |
| Aszonalenin (99) | Neosartorya glabra KUFA 0702 | Marine sponge Mycale sp. | - | [70] |
| Neosartorya takakii KUFC 7898 | Marine alga Amphiroa sp. | - | [72] | |
| (3R)-3-(1H-Indol-3-ylmethyl)-3,4-dihydro-1H-1,4-benzodiazepine-2,5-dione (100) | Neosartorya glabra KUFA 0702 | Marine sponge Mycale sp. | - | [70] |
| Takakiamide (101) | Neosartorya glabra KUFA 0702 | Marine sponge Mycale sp. | - | [70] |
| Neosartorya takakii KUFC 7898 | Marine alga Amphiroa sp. | - | [72] | |
| (11aR)-2,3-Dihydro-1H-pyrrolo{2,1-c}{1,4}benzodiazepine-5,11(10H,11aH)-dione (102) | Neosartorya glabra KUFA 0702 | Marine sponge Mycale sp. | - | [70] |
| Fellutanine A (103) | Neosartorya glabra KUFA 0702 | Marine sponge Mycale sp. | - | [70] |
| Acyl aszonalenin (104) | Aspergillus carneus (MST-MF156) | Estuarine sediment. | - | [71] |
| Neosartorya takakii KUFC 7898 | Marine alga Amphiroa sp. | - | [72] | |
| Terretrione B (105) | Aspergillus terreus SCSIO41008 | Marine sponge Callyspongia sp. | - | [42] |
| Terretrione C (106) | Aspergillus terreus SCSIO41008 | Marine sponge Callyspongia sp. | - | [42] |
| Brevianamide F (107) | Aspergillus terreus SCSIO41008 | Marine sponge Callyspongia sp. | - | [42] |
| cyclo-(L-Trp-L-Ala) (108) | Eurotium chevalieri MUT2316 | Marine sponge Grantia compressa. | Antibacterial and algicidal activities. | [73] |
| Penicillium sp. WF-06 | Marine sediment. | - | [86] | |
| Neoechinulin A (109) | Eurotium chevalieri MUT2316 | Marine sponge Grantia compressa. | Algicidal activity. | [73] |
| Microsporum sp. | Marine red algae. | Cytotoxic activity and induction of apoptosis. | [74] | |
| Echinulin (110) | Eurotium chevalieri MUT2316 | Marine sponge Grantia compressa. | Antibacterial and algicidal activities. | [73] |
| Eurotium repens | Marine sponge Suberites domuncula | - | [75] | |
| Eurotium chevalieri KUFA 0006 | Mangrove plant Rhizophora mucronata Poir. | - | [88] | |
| cyclo-(L-Trp-L-Ile) (111) | Aspergillus niger EN-13 | Marine brown alga Colpomenia sinuosa. | - | [76] |
| cyclo-(L-Trp-L-Phe) (112) | Aspergillus niger EN-13 | Marine brown alga Colpomenia sinuosa. | - | [76] |
| cyclo-(L-Trp-L-Tyr) (113) | Aspergillus niger EN-13 | Marine brown alga Colpomenia sinuosa. | - | [76] |
| (3S,6S)-3,6-dibenzylpiperazine-2,5-dione (114) | Aspergillus candidus KUFA0062 | Marine sponge Epipolasis sp. | - | [77] |
| Mactanamide (115) | Aspergillus ochraceopetaliformis DSW-2 | Sea water. | Weak cytotoxic activity. | [78] |
| 14-Hydroxy-cyclopeptine (116) | Aspergillus sp. SCSIOW2 | Marine deep-sea sediment. | Inhibition of nitric oxide production. | [79] |
| Protuboxepin A (117) | Aspergillus sp. SF-5044 | Marine intertidal sediment. | Cytotoxic activity. | [80] |
| Protuboxepin B (118) | Aspergillus sp. SF-5044 | Marine intertidal sediment. | - | [80] |
| Protubonine A (119) | Aspergillus sp. SF-5044 | Marine intertidal sediment. | - | [80] |
| Protubonine B (120) | Aspergillus sp. SF-5044 | Marine intertidal sediment. | - | [80] |
| Oxepinamide E (121) | Aspergillus sp. (BM-05 and BM-05ML) | Marine brown alga Sargassum sp. | [81] | |
| Shornephine A (122) | Aspergillus sp. (CMB-M081F) | Marine sediment. | Inhibition of P-glycoprotein | [82] |
| 15b-β-Hydroxy-5-N-acetyladreemin (123) | Aspergillus sp. (CMB-M081F) | Marine sediment. | - | [82] |
| 5-N-Acetyladreemin (124) | Aspergillus sp. (CMB-M081F) | Marine sediment. | - | [82] |
| 15b-β-Methoxy-5-N-acetyladreemin (125) | Aspergillus sp. (CMB-M081F) | Marine sediment. | - | [82] |
| Hyalodendrin (126) | Asteromyces cruciatus 763 | An unidentified decaying green alga. | - | [83] |
| Gliovictin (127) | Asteromyces cruciatus 763 | An unidentified decaying green alga. | - | [83] |
| 1N-norgliovictin (128) | Asteromyces cruciatus 763 | An unidentified decaying green alga. | - | [83] |
| Bis-N-norgliovictin (129) | Asteromyces cruciatus 763 | An unidentified decaying green alga. | Antibacterial and antifungal activities. | [83] |
| (±)-7,8-epoxy-brevianamide Q ((±)-130) | Aspergillus versicolor MF180151 | Marine sediment. | - | [84] |
| (±)-8-hydroxy-brevianamide R ((±)-131) | Aspergillus versicolor MF180151 | Marine sediment. | - | [84] |
| (±)-8-epihydroxy-brevianamide R ((±)-132) | Aspergillus versicolor MF180151 | Marine sediment. | - | [84] |
| (±)-Brevianamide R ((±)-133) | Aspergillus versicolor MF180151 | Marine sediment. | - | [84] |
| Aspergillus versicolor HBU-7 | Sea mud sample. | - | [90] | |
| Rostratin A (134) | Exserohilum rostratum (Drechsler) CNK-630 | Marine cyanobacterial mat. | Cytotoxic activity. | [85] |
| Rostratin B (135) | Exserohilum rostratum (Drechsler) CNK-630 | Marine cyanobacterial mat. | Cytotoxic activity. | [85] |
| Rostratin C (136) | Exserohilum rostratum (Drechsler) CNK-630 | Marine cyanobacterial mat. | Cytotoxic activity. | [85] |
| Rostratin D (137) | Exserohilum rostratum (Drechsler) CNK-630 | Marine cyanobacterial mat. | Cytotoxic activity. | [85] |
| Exserohilone (138) | Exserohilum rostratum (Drechsler) CNK-630 | Marine cyanobacterial mat. | - | [85] |
| Gliocladine C (139) | Penicillium sp. WF-06 | Marine Sediment. | Cytotoxic activity. | [86] |
| Azonazine (140) | Aspergillus insulicola | Marine sediment. | Anti-angiogenesis activity. | [87] |
| (11R,14S)-3-(1H-Indol-3ylmethyl)-6-isopropyl-2,5-piperazinedione (141) | Eurotium chevalieri KUFA 0006 | Mangrove plant Rhizophora mucronata Poir. | - | [88] |
| Preechinulin (142) | Eurotium chevalieri KUFA 0006 | Mangrove plant Rhizophora mucronata Poir. | - | [88] |
| Neoechinulin E (143) | Eurotium chevalieri KUFA 0006 | Mangrove plant Rhizophora mucronata Poir. | - | [88] |
| Eurocristatine (144) | Eurotium chevalieri KUFA 0006 | Mangrove plant Rhizophora mucronata Poir. | - | [88] |
| Brevianamide S (145) | Aspergillus versicolor (MF030) | Marine sediment. | Antibacterial activity. | [89] |
| Brevianamide T (146) | Aspergillus versicolor (MF030) | Marine sediment. | - | [89] |
| Brevianamide U (147) | Aspergillus versicolor (MF030) | Marine sediment. | - | [89] |
| Brevianamide V (148) | Aspergillus versicolor (MF030) | Marine sediment. | - | [89] |
| Aspergillus versicolor HBU-7 | Sea mud sample. | Cytotoxic activity. | [90] | |
| Brevianamide N (149) | Aspergillus versicolor (MF030) | Marine sediment. | - | [89] |
| Brevianamide K (150) | Aspergillus versicolor (MF030) | Marine sediment. | - | [89] |
| Aspergillus versicolor HBU-7 | Sea mud sample. | - | [90] | |
| Deoxy brevianamide E (151) | Aspergillus versicolor (MF030) | Marine sediment. | - | [89] |
| Aspergillus versicolor HBU-7 | Sea mud sample. | - | [90] | |
| (±)-Brevianamide Z ((±)-152) | Aspergillus versicolor HBU-7 | Sea mud sample. | - | [90] |
| (±)-Brevianamide Z1 ((±)-153) | Aspergillus versicolor HBU-7 | Sea mud sample. | - | [90] |
| (±)-Brevianamide X ((±)-154) | Aspergillus versicolor HBU-7 | Sea mud sample. | - | [90] |
| (±)-Brevianamide Q ((±)-155) | Aspergillus versicolor HBU-7 | Sea mud sample. | - | [90] |
| 12-Demethyl-12-oxo-eurotechinulin B (156) | Eurotium rubrum G2 | Marine semi-mangrove plant Hibiscus tiliaceus LINN. | Cytotoxic activity. | [91] |
| Variecolorin J (157) | Eurotium rubrum G2 | Marine semi-mangrove plant Hibiscus tiliaceus LINN | - | [91] |
| Eurotechinulin B (158) | Eurotium rubrum G2 | Marine semi-mangrove plant Hibiscus tiliaceus LINN | - | [91] |
| Variecolorin G (159) | Eurotium rubrum G2 | Marine semi-mangrove plant Hibiscus tiliaceus LINN | Cytotoxic activity. | [91] |
| Alkaloid E-7 (160) | Eurotium rubrum G2 | Marine semi-mangrove plant Hibiscus tiliaceus LINN | Cytotoxic activity. | [91] |
| Aspergillus sp. FS445 | Marine deep sediment. | Anti-inflammatory activity. | [95] | |
| Cryptoechinuline G (161) | Eurotium rubrum G2 | Marine semi-mangrove plant Hibiscus tiliaceus LINN | - | [91] |
| Aspergillus sp. FS445 | Marine deep sediment. | Anti-inflammatory activity. | [95] | |
| Isoechinulin B (162) | Eurotium rubrum G2 | Marine semi-mangrove plant Hibiscus tiliaceus LINN | - | [91] |
| Aspergillus sp. FS445 | Marine deep sediment. | Anti-inflammatory activity. | [95] | |
| 7-Isopentenylcryptoechinuline D (163) | Eurotium rubrum G2 | Marine semi-mangrove plant Hibiscus tiliaceus LINN | - | [91] |
| Sclerotioloid A (164) | Aspergillus sclerotiorum ST0501 | Inner tissue of marine sponge. | Anti-inflammatory activity. | [92] |
| Sclerotioloid B (165) | Aspergillus sclerotiorum ST0501 | Inner tissue of marine sponge. | Anti-inflammatory activity. | [92] |
| Sclerotioloid C (166) | Aspergillus sclerotiorum ST0501 | Inner tissue of marine sponge. | Weak anti-inflammatory activity. | [92] |
| Gartryprostatin C (167) | Aspergillus sclerotiorum ST0501 | Inner tissue of marine sponge. | - | [92] |
| Speramide C (168) | Aspergillus sclerotiorum ST0501 | Inner tissue of marine sponge. | - | [92] |
| Asperopiperazine A (169) | Aspergillus sp. DY001 | Red Sea tunicate Didemnum sp. | Antibacterial, antifungal, and cytotoxic activities. | [93] |
| Asperopiperazine B (170) | Aspergillus sp. DY001 | Red Sea tunicate Didemnum sp. | Antibacterial, antifungal, and cytotoxic activities. | [93] |
| Asperchinulin A (171) | Aspergillus sp. FS445 | Marine deep sediment. | - | [95] |
| Asperchinulin B (172) | Aspergillus sp. FS445 | Marine deep sediment. | Anti-inflammatory activity. | [95] |
| Asperchinulin C (173) | Aspergillus sp. FS445 | Marine deep sediment. | Anti-inflammatory activity. | [95] |
| Asperchinulin D (174) | Aspergillus sp. FS445 | Marine deep sediment. | - | [95] |
| Neoechinulin B (175) | Aspergillus sp. FS445 | Marine deep sediment. | Anti-inflammatory activity. | [95] |
| Antibacterial activity. | [96] | |||
| Isoechinulin A (176) | Aspergillus sp. FS445 | Marine deep sediment. | - | [95] |
| Tardioxopiperazine A (177) | Aspergillus sp. FS445 | Marine deep sediment. | - | [95] |
| 24,25-dihydroxyvariecolorin G (178) | Aspergillus chevalieri CS-122 | Marine deep-sea cold seep. | Antibacterial activity. | [96] |
| 25-hydroxyrubrumazine B (179) | Aspergillus chevalieri CS-122 | Marine deep-sea cold seep. | Antibacterial activity. | [96] |
| 22-chloro-25-hydroxyrubrumazine B (180) | Aspergillus chevalieri CS-122 | Marine deep-sea cold seep. | Antibacterial activity. | [96] |
| 25-hydroxyvariecolorin F (181) | Aspergillus chevalieri CS-122 | Marine deep-sea cold seep. | Antibacterial activity. | [96] |
| 27-epi-aspechinulin D (182) | Aspergillus chevalieri CS-122 | Marine deep-sea cold seep. | Antibacterial activity. | [96] |
| Asperazine (183) | Aspergillus niger | Caribbean sponge Hyrtiosproteus sp. | - | [97] |
| Plectosphaeroic acid A (184) | Plectosphaerella cucumerina | Marine sediment. | Inhibition of indoleamine 2,3-dioxygenase. | [98] |
| Plectosphaeroic acid B (185) | Plectosphaerella cucumerina | Marine sediment. | Inhibition of indoleamine 2,3-dioxygenase. | [98] |
| Plectosphaeroic acid C (186) | Plectosphaerella cucumerina | Marine sediment. | Inhibition of indoleamine 2,3-dioxygenase. | [98] |
| T988 A (187) | Plectosphaerella cucumerina | Marine sediment. | - | [98] |
| Diketopiperazine dimer (188) | Aspergillus violaceofuscus | Marine Sponge Reniochalina sp. | Anti-inflammatory activity. | [99] |
| Variecolortin A (189) | Eurotium sp. SCSIO F452 | Marine sediment. | DPPH• radical scavenging activity. | [100] |
| Variecolortin B (190) | Eurotium sp. SCSIO F452 | Marine sediment. | Cytotoxic activity. | [100] |
| Variecolortin C (191) | Eurotium sp. SCSIO F452 | Marine sediment. | Cytotoxic activity. | [100] |
| Cyclic tripeptides | ||||
| Psychrophilin E (192) | Aspergillus sp. (BN-05 & BM-05ML) | Marine brown alga Sargassum sp. | Cytotoxic activity. | [81] |
| Aspergillus versicolor ZLN-60 | Marine mud. | - | [101] | |
| Psychrophilin F (193) | Aspergillus versicolor ZLN-60 | Marine mud. | - | [101] |
| Psychrophilin G (194) | Aspergillus versicolor ZLN-60 | Marine mud. | Lipid-lowering activity. | [101] |
| Psychrophilin H (195) | Aspergillus versicolor ZLN-60 | Marine mud. | - | [101] |
| Sclerotiotide A (196) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | Antifungal activity. | [102] |
| Sclerotiotide B (197) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | Antifungal activity. | [102] |
| Aspergillus ochraceopetaliformis DSW-2 | Sea water. | Weak cytotoxic activity. | [78] | |
| Sclerotiotide C (198) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | - | [102] |
| Sclerotiotide D (199) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | - | [102] |
| Sclerotiotide E (200) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | - | [102] |
| Sclerotiotide F (201) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | Antifungal activity. | [102] |
| Aspergillus ochraceopetaliformis DSW-2 | Sea water. | Weak cytotoxic activity. | [78] | |
| Sclerotiotide G (202) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | - | [102] |
| Sclerotiotide H (203) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | - | [102] |
| Sclerotiotide I (204) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | Antifungal activity. | [102] |
| Sclerotiotide J (205) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | - | [102] |
| Sclerotiotide K (206) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | - | [102] |
| JBIR-15 (207) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | Antifungal activity. | [102] |
| Aspochracin (208) | Aspergillus sclerotiorum PT06-1 | Marine salt sediment. | - | [102] |
| Sclerotiotide L (209) | Aspergillus violaceofuscus | Marine sponge Reniochalina sp. | Weak anti-inflammatory activity. | [99] |
| Sclerotiotide M (210) | Aspergillus ochraceopetaliformis DSW-2 | Sea water. | Weak cytotoxic activity. | [78] |
| Cyclic tetrapeptides | ||||
| Endolide A (211) | Stachylidium sp. | Marine sponge Callyspongia sp. sf. C. flammea. | Inhibition of G-protein-coupled receptors. | [103] |
| Endolide B (212) | Stachylidium sp. | Marine sponge Callyspongia sp. sf. C. flammea. | Inhibition of G-protein-coupled receptors. | [103] |
| Endolide C (213) | Stachylidium sp. 293 K04 | Marine sponge Callyspongia sp. sf. C. flammea. | - | [104] |
| Endolide D (214) | Stachylidium sp. 293 K04 | Marine sponge Callyspongia sp. sf. C. flammea. | - | [104] |
| Hirsutide (215) | Stachylidium sp. 293 K04 | Marine sponge Callyspongia sp. sf. C. flammea. | - | [104] |
| Violaceomide A (216) | Aspergillus violaceofuscus | Marine Sponge Reniochalina sp. | Anti-inflammatory activity. | [99] |
| Sartoryblabramide A (217) | Neosartorya glabra KUFA 0702 | Marine sponge Mycale sp. | - | [70] |
| Sartoryblabramide B (218) | Neosartorya glabra KUFA 0702 | Marine sponge Mycale sp. | - | [70] |
| cyclo-(L-leucyl-trans-4-hydroxy-L-prolyl-D-leucyl-trans-4-hydroxy-L-proline) (219) | Co-culture of Phomopsis sp. K38 & Altenaria sp. E33 | Mangrove. | Antifungal activity. | [105] |
| 5,5ʹ-epoxy-MKN-349A (220) | Penicilluim sp. GD6 | Marine mangrove Bruguiera gymnorrhiza. | - | [107] |
| Asperterrestide A (221) | Aspergillus terreus SCSGAF0162 | Marine gorgonian Echinogorgia aurantiaca. | Antiviral and cytotoxic activities. | [108] |
| Compound (222) | Aspergillus flavipes | Marine isopod Ligia oceanica. | - | [109] |
| Microsporin A (223) | Microsporum cf. gypseum | Marine bryozoan Bugula sp. | Cytotoxic and histone deacethylase inhibitory activities. | [110] |
| Microsporin B (224) | Microsporum cf. gypseum | Marine bryozoan Bugula sp. | Cytotoxic activity. | [110] |
| Cyclic penapeptides | ||||
| Caletasin (225) | Aspergillus sp. MEXU27854 | Marine intertidal sand. | - | [111] |
| Asperpeptide A (226) | Aspergillus sp. XS-20090B15 | Marine gorgonian Muricella abnormaliz. | Antibacterial activity. | [46] |
| Cotteslosin A (227) | Aspergillus versicolor (MST-MF495) | Low-tide region. | Cytotoxic activity. | [112] |
| Cotteslosin B (228) | Aspergillus versicolor (MST-MF495) | Low-tide region. | - | [112] |
| Lajollamide A (229) | Asteromyces cruciatus 763 | An unidentified decaying green alga. | Antibacterial activity. | [83] |
| JG002CPA (230) | Aspergillus allahabadii | Marine sediment. | Inhibition of enzyme sortase A (SrtA) activity. | [51] |
| JG002CPA (231) | Aspergillus allahabadii | Marine sediment. | Inhibition of sortase A (SrtA) and isocitrate lyase (ICL) activities. | [51] |
| Asperflomide (232) | Aspergillus flocculosus 16D-1 | Marine sponge Phakellia fusca. | Inhibition of tankyrase1/2 activity. | [113] |
| Malformin C (233) | Aspergillus sp. SCSIOW2 | Marine sediment. | Algicidal activity. | [114] |
| Aspergillus niger BRF-074 | Marine sediment. | - | [65] | |
| Aspergillus niger | Marine sponge Hyrtiosproteus sp. | - | [97] | |
| Malformin A1 (234) | Aspergillus niger BRF-074 | Marine sediment. | - | [65] |
| Aspergillipeptide D (235) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata. | Antiviral activity. | [41] |
| Versicotide A (236) | Aspergillus versicolor ZLN-60 | Marine mud. | - | [115] |
| Versicotide B (237) | Aspergillus versicolor ZLN-60 | Marine mud. | - | [115] |
| Pseudoviridinutan A (238) | Aspergillus pseudoviridinutans TW58-5 | Marine hydrothermal vent sediment. | Anti-inflammatory activity. | [116] |
| Pseudoviridinutan B (239) | Aspergillus pseudoviridinutans TW58-5 | Marine hydrothermal vent sediment. | Anti-inflammatory activity. | [116] |
| Pseudoviridinutan C (240) | Aspergillus pseudoviridinutans TW58-5 | Marine hydrothermal vent sediment. | Anti-inflammatory activity. | [116] |
| Pseudoviridinutan D (241) | Aspergillus pseudoviridinutans TW58-5 | Marine hydrothermal vent sediment. | Anti-inflammatory activity. | [116] |
| Pseudoviridinutan E (242) | Aspergillus pseudoviridinutans TW58-5 | Marine hydrothermal vent sediment. | Anti-inflammatory activity. | [116] |
| Pseudoviridinutan F (243) | Aspergillus pseudoviridinutans TW58-5 | Marine hydrothermal vent sediment. | Anti-inflammatory activity. | [116] |
| Pseudoviridinutan G (244) | Aspergillus pseudoviridinutans TW58-5 | Marine hydrothermal vent sediment. | Anti-inflammatory activity. | [116] |
| Cyclic hexapeptides | ||||
| Aspersymmetide A (245) | Aspergillus versicolor (TA01-14) | Marine gorgonian coral Carijoa sp. (GX-WZ-2010001). | Cytotoxic activity. | [117] |
| Simplicilliumtide J (246) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifungal and antiviral activities. | [49] |
| Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifungal activity. | [118] | |
| Simplicilliumtide K (247) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | - | [49] |
| Simplicilliumtide L (248) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | - | [49] |
| Simplicilliumtide M (249) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | - | [49] |
| Verlamelin A (250) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifungal and antiviral activities. | [49] |
| Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifungal activity. | [118] | |
| Verlamelin B (251) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifungal and antiviral activities. | [49] |
| Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifungal activity. | [118] | |
| Simplicilliumtide N (252) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifungal activity. | [118] |
| Simplicilliumtide O (253) | Simplicillium obclavatum EIODSF 020 | Marine deep-sea sediment. | Antifungal activity. | [118] |
| Versicotide C (254) | Aspergillus versicolor ZLN-60 | Marine mud. | - | [101] |
| Sclerotide A (255) | Aspergillus sclerotiorum PT06-1 | Putain sea salt filed. | Antifungal activity. | [119] |
| Aspergillus sclerotiorum SCSIO41031 | Marine soft coral. | - | [120] | |
| Sclerotide B (256) | Aspergillus sclerotiorum PT06-1 | Putain sea salt filed. | Antifungal, antibacterial, and cytotoxic activities. | [119] |
| Sclerotide C (257) | Aspergillus sclerotiorum SCSIO41031 | Marine soft coral. | - | [120] |
| Sclerotide D (258) | Aspergillus sclerotiorum SCSIO41031 | Marine soft coral. | - | [120] |
| Sclerotide E (259) | Aspergillus sclerotiorum SCSIO41031 | Marine soft coral. | - | [120] |
| Similanamide (260) | Aspergillus similanensis KUFA0013 | Marine sponge Rhabdermia sp. | Cytotoxic activity. | [121] |
| Acremonpeptide A (261) | Acremonium persicinum SCSIO115. | Marine sediment. | Antiviral activity. | [122] |
| Acremonpeptide B (262) | Acremonium persicinum SCSIO115. | Marine sediment. | Antiviral activity. | [122] |
| Acremonpeptide C (263) | Acremonium persicinum SCSIO115. | Marine sediment. | Antiviral activity. | [122] |
| Acremonpeptide D 264) | Acremonium persicinum SCSIO115. | Marine sediment. | Antiviral activity. | [122] |
| Al(III)-acremonpeptide D (265) | Acremonium persicinum SCSIO115. | Marine sediment. | Antiviral activity. | [122] |
| Petrosamide A (266) | Aspergillus sp. 151304 | Marine sponge Petrosia sp. | Inhibition of pancreatic lipase activity. | [123] |
| Petrosamide A (267) | Aspergillus sp. 151304 | Marine sponge Petrosia sp. | Inhibition of pancreatic lipase activity. | [123] |
| Petrosamide A (268) | Aspergillus sp. 151304 | Marine sponge Petrosia sp. | Inhibition of pancreatic lipase activity. | [123] |
| Cyclic heptapeptides | ||||
| Asperversiamide A (269) | Aspergillus versicolor (CHNSCLM-0063) | Marine coral. | Antibacterial activity. | [124] |
| Asperversiamide B (270) | Aspergillus versicolor (CHNSCLM-0063) | Marine coral. | Antibacterial and anti-Mycobacterium tuberculosis activities. | [124] |
| Asperversiamide C (271) | Aspergillus versicolor (CHNSCLM-0063) | Marine coral. | Antibacterial activity. | [124] |
| Asperheptatide A (272) | Aspergillus versicolor (CHNSCLM-0063) | Marine gorgonian coral Rumphella aggregata. | Anti-Mycobacterium tuberculosis activity. | [125] |
| Asperheptatide B (273) | Aspergillus versicolor (CHNSCLM-0063) | Marine gorgonian coral Rumphella aggregata. | Anti-Mycobacterium tuberculosis activity. | [125] |
| Asperheptatide C (274) | Aspergillus versicolor (CHNSCLM-0063) | Marine gorgonian coral Rumphella aggregata. | - | [125] |
| Asperheptatide D (275) | Aspergillus versicolor (CHNSCLM-0063) | Marine gorgonian coral Rumphella aggregata. | - | [125] |
| Cordyheptapeptide C (276) | Acremonium persicinum SCSIO115 | Marine sediment. | Cytotoxic activity. | [126] |
| Cordyheptapeptide D (277) | Acremonium persicinum SCSIO115 | Marine sediment. | Cytotoxic activity. | [126] |
| Cordyheptapeptide E (278) | Acremonium persicinum SCSIO115 | Marine sediment. | Cytotoxic activity. | [126] |
| Talarolide A (279) | Talaromyces sp. (CMB-TU011) | Marine tunicate. | - | [127] |
| Mortiamide A (280) | Mortierella sp. RKAG 110 | Marine sediment. | - | [128] |
| Mortiamide B (281) | Mortierella sp. RKAG 110 | Marine sediment. | - | [128] |
| Mortiamide C (282) | Mortierella sp. RKAG 110 | Marine sediment. | - | [128] |
| Mortiamide D (283) | Mortierella sp. RKAG 110 | Marine sediment. | - | [128] |
| Unguisin A (284) | Emericella unguis | Stomolopus meliagris (medusa) | Antibacterial activity. | [129] |
| Aspergillus unguis 6-20-6 GDMCC 60337 | Seaweed. | Antibacterial activity. | [130] | |
| Aspergillus unguis IV17-109 | Deep-sea shrimp. | - | [39] | |
| Unguisin B (285) | Emericella unguis | Stomolopus meliagris (medusa). | Antibacterial activity. | [129] |
| Unguisin E (286) | Aspergillus sp. AF119 | Soil of Xiamen beach. | - | [131] |
| Scytalidamide A (287) | Scytalidium sp. CNC-310 | Marine green alga Halimedia sp. | Cytotoxic activity. | [132] |
| Scytalidamide B (288) | Scytalidium sp. CNC-310 | Marine green alga Halimedia sp. | Cytotoxic activity. | [132] |
| Maribasin C (289) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata Nutting. | Antifungal activity. | [50] |
| Maribasin D (290) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata Nutting. | Antifungal activity. | [50] |
| Maribasin E (291) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata Nutting. | Antifungal activity. | [50] |
| Maribasin A (292) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata Nutting. | Antifungal activity. | [50] |
| Maribasin B (293) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata Nutting. | Antifungal activity. | [50] |
| Marihysin A (294) | Aspergillus sp. SCSIO 41501 | Marine gorgonian Melitodes squamata Nutting | - | [50] |
| Cyclic nonapeptides | ||||
| Clonostachysin A (295) | Clonostachys rogersoniana (HJK9) | Marine sponge Halicondria japonica. | Anti-dinoflagellate activity. | [133] |
| Clonostachysin B (296) | Clonostachys rogersoniana (HJK9) | Marine sponge Halicondria japonica. | Anti-dinoflagellate activity. | [133] |
| Cyclic decapeptides | ||||
| Auyuittuqamide A (297) | Sesquicillium microsporum RKAG 186 | Marine sediment (intertidal zone). | - | [134] |
| Auyuittuqamide B (298) | Sesquicillium microsporum RKAG 186 | Marine sediment (intertidal zone). | - | [134] |
| Auyuittuqamide C (299) | Sesquicillium microsporum RKAG 186 | Marine sediment (intertidal zone). | - | [134] |
| Auyuittuqamide D (300) | Sesquicillium microsporum RKAG 186 | Marine sediment (intertidal zone). | - | [134] |
| Linear depsipeptides | ||||
| Waspergillamide A (301) | Aspergillus sp. CMB-W031 | Marine mud dauber wasp Sceliphron sp. | - | [135] |
| Cyclic depsipeptides | ||||
| Saroclide A (302) | Sarocladium kiliense HDN11-112 | Mangrove plant Aricennia marina | Lipid-lowering activity. | [136] |
| Saroclide B (303) | Sarocladium kiliense HDN11-112 | Mangrove plant Aricennia marina | - | [136] |
| Acremolide A (304) | Acremonium sp. (MST-MF588a) | Estuarine sediment. | - | [137] |
| Acremolide B (305) | Acremonium sp. (MST-MF588a) | Estuarine sediment. | - | [137] |
| Acremolide C (306) | Acremonium sp. (MST-MF588a) | Estuarine sediment. | - | [137] |
| Acremolide D (307) | Acremonium sp. (MST-MF588a) | Estuarine sediment. | - | [137] |
| Calcaripeptide A (308) | Calcarisporium sp. strain KF525 | Sea. | - | [138] |
| Calcaripeptide B (309) | Calcarisporium sp. strain KF525 | Sea. | - | [138] |
| Calcaripeptide C (310) | Calcarisporium sp. strain KF525 | Sea. | - | [138] |
| Beauvericin (311) | Aspergillus terreus (no. GX7-3B) | Marine mangrove Bruguiera gymnoihiza (Linn.) Savigny. | Anti-acetylcholinesterase and cytotoxic activity. | [140] |
| Fusarium sp. (No.DZ27) | Marine mangrove. | Cytotoxic activity and induction of apoptosis. | [141] | |
| Guangomide A (312) | Strain No. 001314c | A yellow fan-shaped marine sponge (Coll. No. 00314). | Antibacterial activity. | [142] |
| Guangomide B (313) | Strain No. 001314c | A yellow fan-shaped marine sponge (Coll. No. 00314). | Antibacterial activity. | [142] |
| Homodestcardin (314) | Strain No. 001314c | A yellow fan-shaped marine sponge (Coll. No. 00314). | - | [142] |
| Sansalvamide A (315) | Fusarium sp. | Marine seagrass Halodule wrightii | Cytotoxic activity. | [145,146] |
| N-methylsansalvamide (316) | Fusarium sp. CNL-619 | Marine mangrove green alga Avrainvillea sp. | Cytotoxic activity. | [146] |
| Acremonamide (317) | Acremonium sp. CNQ-049 | Marine sediment. | Wound healing activity. | [147] |
| Zygosporamide (318) | Zygosporium masonii (CNK458) | Marine cyanobacterium. | Cytotoxic activity. | [148] |
| Alternaramide (319) | Altenaria sp. SF-5016 | Marine shoreline sediment. | Antibacterial and inhibition of protein tyrosine phosphatase 1B activity. | [149] |
| Destruxin A (320) | Metarrhizium sp. (strain number 001103) | Marine sponge Pseudoceratina purpurea (coll. no. 00103). | Cytotoxic activity. | [53] |
| Destruxin B (321) | Metarrhizium sp. (strain number 001103) | Marine sponge Pseudoceratina purpurea (coll. no. 00103). | - | [53] |
| Destruxin B2 (322) | Metarrhizium sp. (strain number 001103) | Marine sponge Pseudoceratina purpurea (coll. no. 00103). | Cytotoxic activity. | [53] |
| Desmethyldestruxin B (323) | Metarrhizium sp. (strain number 001103) | Marine sponge Pseudoceratina purpurea (coll. no. 00103). | Cytotoxic activity. | [53] |
| Destruxin E chlorohydrin (324) | Metarrhizium sp. (strain number 001103) | Marine sponge Pseudoceratina purpurea (coll. no. 00103). | Cytotoxic activity. | [53] |
| Destruxin E2 chlorohydrin (325) | Metarrhizium sp. (strain number 001103) | Marine sponge Pseudoceratina purpurea (coll. no. 00103). | Cytotoxic activity. | [53] |
| Aspergillicin A (326) | Aspergillus carneus (MST-MF156) | Marine estuarine sediment. | - | [71] |
| Aspergillicin B (327) | Aspergillus carneus (MST-MF156) | Marine estuarine sediment. | - | [71] |
| Aspergillicin C (328) | Aspergillus carneus (MST-MF156) | Marine estuarine sediment. | - | [71] |
| Aspergillicin D (329) | Aspergillus carneus (MST-MF156) | Marine estuarine sediment. | - | [71] |
| Aspergillicin E (330) | Aspergillus carneus (MST-MF156) | Marine estuarine sediment. | - | [71] |
| Scopularide A (331) | Scopulariopsis brevicaulis NCPF 2177 | Marine sponge Tethya aurantium. | Cytotoxic activity. | [151] |
| Aspergillus flavus | Marine soft coral Sarcophyton ehrenbergi. | Larvicidal and inhibition of pancreatic lipase activity. | [152] | |
| Nigrospora oryzae PF18 | Marine sponge Phakellia fusca | - | [153] | |
| Penicillium chrysogenum (CHSCLM-0003) | Marine gorgonian coral Carijoa sp. (GX-WZ-2010001). | Cytotoxic activity. | [159] | |
| Scopularide B (332) | Scopulariopsis brevicaulis NCPF 2177 | Marine sponge Tethya aurantium. | Cytotoxic activity. | [151] |
| Aspergillus flavus | Marine soft coral Sarcophyton ehrenbergi. | Larvicidal and inhibition of pancreatic lipase activity. | [152] | |
| Penicillium chrysogenum (CHSCLM-0003) | Marine gorgonian coral Carijoa sp. (GX-WZ-2010001). | Cytotoxic activity. | [159] | |
| Oryzamide A (333) | Nigrospora oryzae PF18 | Marine sponge Phakellia fusca. | - | [153] |
| Oryzamide B (334) | Nigrospora oryzae PF18 | Marine sponge Phakellia fusca. | - | [153] |
| Oryzamide C (335) | Nigrospora oryzae PF18 | Marine sponge Phakellia fusca. | - | [153] |
| Oryzamide D (336) | Nigrospora oryzae PF18 | Marine sponge Phakellia fusca. | - | [153] |
| Oryzamide E (337) | Nigrospora oryzae PF18 | Marine sponge Phakellia fusca. | - | [153] |
| Isaridin G (338) | Beauveria feline EN-135 | An unidentified marine bryozoan. | Antibacterial activity. | [157] |
| Desmethylisaridin G (339) | Beauveria feline EN-135 | An unidentified marine bryozoan. | Antibacterial activity. | [157] |
| Desmethylisaridin C1 (340) | Beauveria feline EN-135 | An unidentified marine bryozoan. | Antibacterial activity. | [157] |
| Isaridin A (341) | Beauveria feline EN-135 | An unidentified marine bryozoan. | - | [157] |
| Isaridin B (342) | Beauveria feline EN-135 | An unidentified marine bryozoan. | - | [157] |
| Isaridin E (343) | Beauveria feline EN-135 | An unidentified marine bryozoan. | Antibacterial activity. | [157] |
| Beauveria feline SYSU-MS7908 | Marine ascidian Styela plicata. | - | [161] | |
| Chrysogeamide A (344) | Penicillium chrysogenum (CHSCLM-0003) | Marine gorgonian coral Carijoa sp. (GX-WZ-2010001). | Anti-angiogenesis activity. | [159] |
| Chrysogeamide B (345) | Penicillium chrysogenum (CHSCLM-0003) | Marine gorgonian coral Carijoa sp. (GX-WZ-2010001). | Anti-angiogenesis activity. | [159] |
| Chrysogeamide C (346) | Penicillium chrysogenum (CHSCLM-0003) | Marine gorgonian coral Carijoa sp. (GX-WZ-2010001). | - | [159] |
| Chrysogeamide D (347) | Penicillium chrysogenum (CHSCLM-0003) | Marine gorgonian coral Carijoa sp. (GX-WZ-2010001). | - | [159] |
| Chrysogeamide E (348) | Penicillium chrysogenum (CHSCLM-0003) | Marine gorgonian coral Carijoa sp. (GX-WZ-2010001). | - | [159] |
| Chrysogeamide F (349) | Penicillium chrysogenum (CHSCLM-0003) | Marine gorgonian coral Carijoa sp. (GX-WZ-2010001). | - | [159] |
| Chrysogeamide G (350) | Penicillium chrysogenum (CHSCLM-0003) | Marine gorgonian coral Carijoa sp. (GX-WZ-2010001). | - | [159] |
| Nodupetide (351) | Penicillium chrysogenum (CHSCLM-0003) | Marine gorgonian coral Carijoa sp. (GX-WZ-2010001). | Antibacterial and anti-angiogenesis activities. | [159] |
| FJ120DPA (352) | Aspergillus ochraceopetaliformis. | Marine sediment. | Inhibition of sortase A (SrtA) activity. | [51] |
| Emericellamide A (353) | Emericella sp. (strain CNL-878) | Marine green alga Halimeda sp. | Antibacterial and cytotoxic activities. | [160] |
| Emericellamide B (354) | Emericella sp. (strain CNL-878) | Marine green alga Halimeda sp. | Antibacterial and cytotoxic activities. | [160] |
| Scopularide I (355) | Aspergillus sclerotiorum SCSIO41031 | Marine soft coral. | Anti-acetylcholinesterase and cytotoxic activities. | [120] |
| Asperflosamide (356) | Aspergillus flocculosus 16D-1 | Marine sponge Phakellia fusca. | Inhibition of tankyrase1/2 activity. | [113] |
| Isaridin I (357) | Beauveria feline SYSU-MS7908 | Marine ascidian Styela plicata. | - | [161] |
| Isaridin J (358) | Beauveria feline SYSU-MS7908 | Marine ascidian Styela plicata. | Antifungal activity. | [161] |
| Isaridin K (359) | Beauveria feline SYSU-MS7908 | Marine ascidian Styela plicata. | - | [161] |
| Isaridin L (360) | Beauveria feline SYSU-MS7908 | Marine ascidian Styela plicata. | Antifungal activity. | [161] |
| Isaridin M (361) | Beauveria feline SYSU-MS7908 | Marine ascidian Styela plicata. | - | [161] |
| Isaridin N (362) | Beauveria feline SYSU-MS7908 | Marine ascidian Styela plicata. | - | [161] |
| IB-01212 (363) | Clonostachys sp. ESNA-A009 | An unidentified marine sponge. | Cytotoxic activity. | [162] |
| - | - | Anti-parasite activity. | [166] | |
| Phaeosphamides A (364) | Phaeosphaeriopsis sp. Esf-30 | Rhizosphere sediment of a mangrove plant Bruguiera gymnorhiza. | Cytotoxic activity. | [164] |
| Phaeosphamides B (365) | Phaeosphaeriopsis sp. Esf-30 | Rhizosphere sediment of a mangrove plant Bruguiera gymnorhiza. | - | [164] |
| Sch 217048 (366) | Phaeosphaeriopsis sp. Esf-30 | Rhizosphere sediment of a mangrove plant Bruguiera gymnorhiza. | - | [164] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hafez Ghoran, S.; Taktaz, F.; Sousa, E.; Fernandes, C.; Kijjoa, A. Peptides from Marine-Derived Fungi: Chemistry and Biological Activities. Mar. Drugs 2023, 21, 510. https://doi.org/10.3390/md21100510
AMA Style
Hafez Ghoran S, Taktaz F, Sousa E, Fernandes C, Kijjoa A. Peptides from Marine-Derived Fungi: Chemistry and Biological Activities. Marine Drugs. 2023; 21(10):510. https://doi.org/10.3390/md21100510
Chicago/Turabian StyleHafez Ghoran, Salar, Fatemeh Taktaz, Emília Sousa, Carla Fernandes, and Anake Kijjoa. 2023. "Peptides from Marine-Derived Fungi: Chemistry and Biological Activities" Marine Drugs 21, no. 10: 510. https://doi.org/10.3390/md21100510
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.