
Galactomannans for Entrapment of Gliomastix murorum Laccase and Their Use in Reactive Blue 2 Decolorization

 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation, Screening, and Identification of Laccase-Producing Microorganism
2.3. Culture Conditions
2.4. Enzyme Extraction
2.5. Activity Assays
2.6. Preparation of Galactomannan/Laccase Beads
2.7. Reactive Blue 2 Decolorization Assays
2.8. Effect of Temperature and pH on the Enzyme Stability
2.9. Reusability Capacity of Entrapped Laccase
3. Results and Discussion
3.1. Isolation, Screening, and Identification of Laccase-Producing Microorganism
3.2. Laccase Production
3.3. Kinetic Parameters of Free and Entrapped Laccase
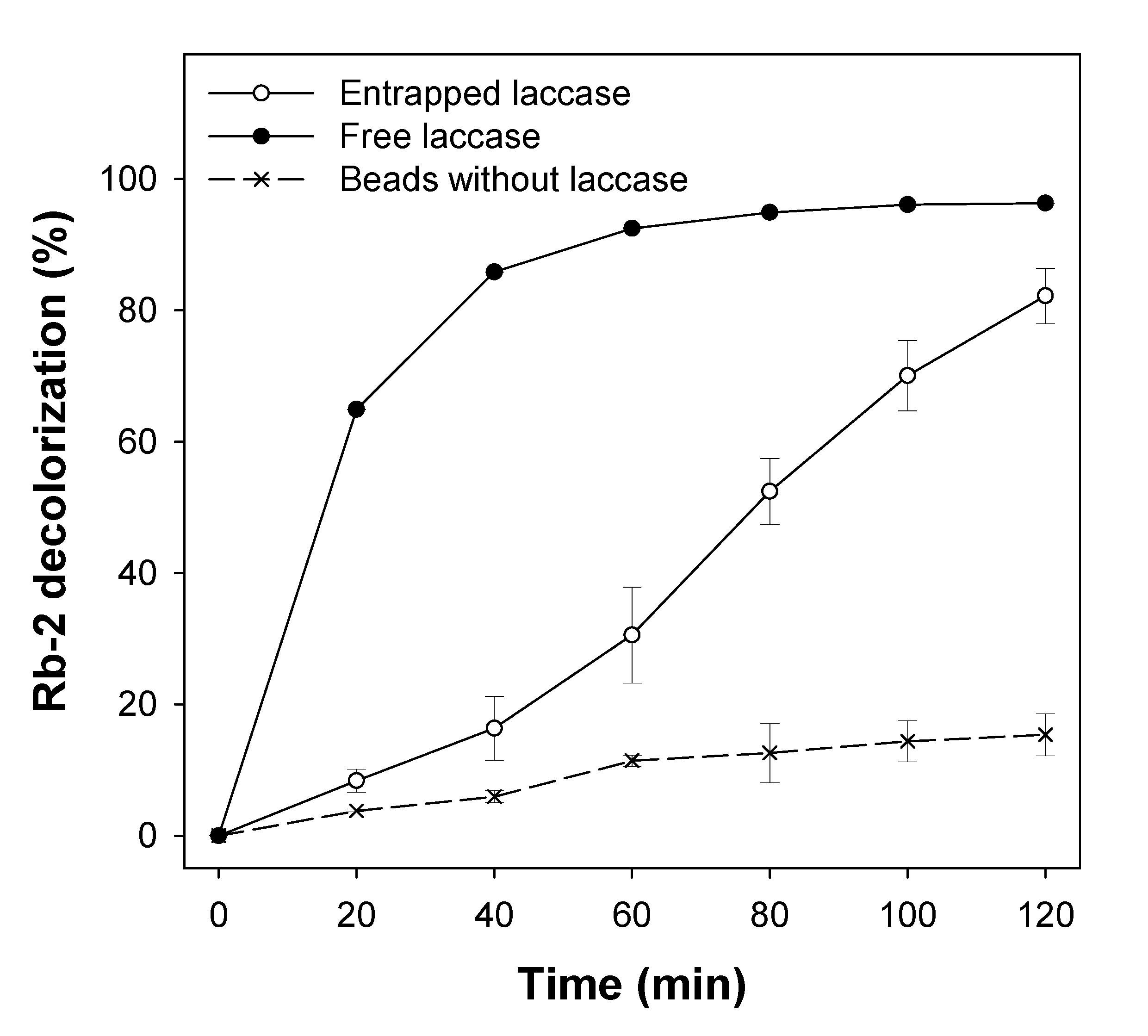
3.4. Evaluation of Laccase in Reactive Blue 2 Decolorization
3.5. Temperature and pH Effects on the Stability of Free and Entrapped Laccase
3.6. Reusability of Entrapped Laccase
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABTS | 2,2′ azino bis (3 ethylbenzthiazoline-6-sulfonic acid) |
| BLAST | Basic Local Alignment Search Tool |
| DNS | dinitrosalisylic acid |
| Rb-2 | Reactive Blue 2 |
References
- Ranjbar, D.; Raeiszadeh, M.; Lewis, L.; MacLachlan, M.J.; Hatzikiriakos, S.G. Adsorptive removal of Congo red by surfactant modified cellulose nanocrystals: A kinetic, equilibrium, and mechanistic investigation. Cellulose 2020, 27, 3211–3232. [Google Scholar] [CrossRef]
- Britos, C.N.; Gianolini, J.E.; Portillo, H.; Trelles, J.A. Biodegradation of industrial dyes by a solvent, metal and surfactant-stable extracellular bacterial laccase. Biocatal. Agric. Biotechnol. 2018, 14, 221–227. [Google Scholar] [CrossRef]
- Dayi, B.; Kyzy, A.D.; Abduloglu, Y.; Cikrikci, K.; Akdogan, H.A. Investigation of the ability of immobilized cells to different carriers in removal of selected dye and characterization of environmentally friendly laccase of Morchella esculenta. Dye. Pigment. 2018, 151, 15–21. [Google Scholar] [CrossRef]
- Lellis, B.; Fávaro-Polonio, C.Z.; Pamphile, J.A.; Polonio, J.C. Effects of textile dyes on health and the environment and bioremediation potential of living organisms. Biotechnol. Res. Innov. 2019, 3, 275–290. [Google Scholar] [CrossRef]
- Papić, S.; Koprivanac, N.; Lončarić Božić, A.; Meteš, A. Removal of some reactive dyes from synthetic wastewater by combined Al(III) coagulation/carbon adsorption process. Dye. Pigment. 2004, 62, 291–298. [Google Scholar] [CrossRef]
- Lin, L.; Zhu, W.; Zhang, C.; Hossain, M.Y.; Oli, Z.B.S.; Pervez, M.N.; Sarker, S.; Hoque, M.I.U.; Cai, Y.; Naddeo, V. Combination of wet fixation and drying treatments to improve dye fixation onto spray-dyed cotton fabric. Sci. Rep. 2021, 11, 15403. [Google Scholar] [CrossRef]
- Haque, A.N.M.A.; Hannan, M.; Rana, M.M.J.F. Compatibility analysis of reactive dyes by exhaustion-fixation and adsorption isotherm on knitted cotton fabric. Fash. Tex. 2015, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mirzaei, N.; Ghaffari, H.R.; Sharafi, K.; Velayati, A.; Hoseindoost, G.; Rezaei, S.; Mahvi, A.H.; Azari, A.; Dindarloo, K. Modified natural zeolite using ammonium quaternary based material for Acid red 18 removal from aqueous solution. J. Environ. Chem. Eng. 2017, 5, 3151–3160. [Google Scholar] [CrossRef]
- Jabar, J.M.; Odusote, Y.A. Removal of cibacron blue 3G-A (CB) dye from aqueous solution using chemo-physically activated biochar from oil palm empty fruit bunch fiber. Arab. J. Chem. 2020, 13, 5417–5429. [Google Scholar] [CrossRef]
- Pajootan, E.; Arami, M.; Mahmoodi, N.M. Binary system dye removal by electrocoagulation from synthetic and real colored wastewaters. J. Taiwan Inst. Chem. Eng. 2012, 43, 282–290. [Google Scholar] [CrossRef]
- Aravind, P.; Selvaraj, H.; Ferro, S.; Sundaram, M. An integrated (electro- and bio-oxidation) approach for remediation of industrial wastewater containing azo-dyes: Understanding the degradation mechanism and toxicity assessment. J. Hazard. Mater. 2016, 318, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-C.; Hsieh, Y.-H.; Lai, P.-F.; Li, C.-H.; Kao, C.-L. Photodegradation treatment of azo dye wastewater by UV/TiO2 process. Dye. Pigment. 2006, 68, 191–195. [Google Scholar] [CrossRef]
- Katheresan, V.; Kansedo, J.; Lau, S.Y. Efficiency of various recent wastewater dye removal methods: A review. J. Environ. Chem. Eng. 2018, 6, 4676–4697. [Google Scholar] [CrossRef]
- Alvarez, L.H.; Arvizu, I.C.; Garcia-Reyes, R.B.; Martinez, C.M.; Olivo-Alanis, D.; Del Angel, Y.A. Quinone-functionalized activated carbon improves the reduction of congo red coupled to the removal of p-cresol in a UASB reactor. J. Hazard. Mater. 2017, 338, 233–240. [Google Scholar] [CrossRef]
- Bayramoglu, G.; Salih, B.; Akbulut, A.; Arica, M.Y. Biodegradation of Cibacron Blue 3GA by insolubilized laccase and identification of enzymatic byproduct using MALDI-ToF-MS: Toxicity assessment studies by Daphnia magna and Chlorella vulgaris. Ecotoxicol. Environ. Saf. 2019, 170, 453–460. [Google Scholar] [CrossRef]
- Rodríguez, S. Dye removal by immobilised fungi. Biotechnol. Adv. 2009, 27, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Cabana, H.; Ahamed, A.; Leduc, R. Conjugation of laccase from the white rot fungus Trametes versicolor to chitosan and its utilization for the elimination of triclosan. Bioresour. Technol. 2011, 102, 1656–1662. [Google Scholar] [CrossRef] [PubMed]
- Arregui, L.; Ayala, M.; Gómez-Gil, X.; Gutiérrez-Soto, G.; Hernández-Luna, C.E.; de los Santos, M.H.; Levin, L.; Rojo-Domínguez, A.; Romero-Martínez, D.; Saparrat, M.C.N.; et al. Laccases: Structure, function, and potential application in water bioremediation. Microb. Cell Fact. 2019, 18, 200. [Google Scholar] [CrossRef]
- Blánquez, A.; Rodríguez, J.; Brissos, V.; Mendes, S.; Martins, L.O.; Ball, A.S.; Arias, M.E.; Hernández, M. Decolorization and detoxification of textile dyes using a versatile Streptomyces laccase-natural mediator system. Saudi J. Biol. Sci. 2019, 26, 913–920. [Google Scholar] [CrossRef]
- Mtibaa, R.; Barriuso, J.; de Eugenio, L.; Aranda, E.; Belbahri, L.; Nasri, M.; Martinez, M.J.; Mechichi, T. Purification and characterization of a fungal laccase from the ascomycete Thielavia sp. and its role in the decolorization of a recalcitrant dye. Int. J. Biol. Macromol. 2018, 120 Pt B, 1744–1751. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Ramírez, M.G.; Rivera-Ríos, J.M.; Téllez-Jurado, A.; Maqueda-Gálvez, A.P.; Mercado-Flores, Y.; Arana-Cuenca, A. Screening for thermotolerant ligninolytic fungi with laccase, lipase, and protease activity isolated in Mexico. J. Environ. Manag. 2012, 95, S256–S259. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Bustamante, S.R.; Mondaca-Fernández, I.; Caro-Reyes, R.B.; Gámez-Gutierrez, L.A.; Santos-Villalobos, S.; Meza-Montenegro, M.M.; Balderas-Cortés, J.J. Selección de cepas productoras de enzimas ligninolíticas nativas del Valle del Yaqui. Nova Sci. 2017, 9, 24–36. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Gupta, N. Microbial Laccase: A robust enzyme and its industrial applications. Biologia 2020, 75, 1183–1193. [Google Scholar] [CrossRef]
- Ma, H.F.; Meng, G.; Cui, B.K.; Si, J.; Dai, Y.C. Chitosan crosslinked with genipin as supporting matrix for biodegradation of synthetic dyes: Laccase immobilization and characterization. Chem. Eng. Res. Des. 2018, 132, 664–676. [Google Scholar] [CrossRef]
- Daâssi, D.; Rodríguez-Couto, S.; Nasri, M.; Mechichi, T. Biodegradation of textile dyes by immobilized laccase from Coriolopsis gallica into Ca-alginate beads. Int. Biodeterior. Biodegrad. 2014, 90, 71–78. [Google Scholar] [CrossRef]
- Campia, P.; Ponzini, E.; Rossi, B.; Farris, S.; Silvetti, T.; Merlini, L.; Brasca, M.; Grandori, R.; Galante, Y.M. Aerogels of enzymatically oxidized galactomannans from leguminous plants: Versatile delivery systems of antimicrobial peptides and enzymes. Carbohydr. Polym. 2017, 158, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Cerqueira, M.A.; Bourbon, A.I.; Pinheiro, A.C.; Martins, J.T.; Souza, B.W.S.; Teixeira, J.A.; Vicente, A.A. Galactomannans use in the development of edible films/coatings for food applications. Trends Food Sci. Technol. 2011, 22, 662–671. [Google Scholar] [CrossRef] [Green Version]
- Zheng, F.; Cui, B.; Wu, X.; Meng, G.; Liu, H.; Si, J. Immobilization of laccase onto chitosan beads to enhance its capability to degrade synthetic dyes. Int. Biodeterior. Biodegrad. 2016, 110, 69–78. [Google Scholar] [CrossRef]
- Yavaşer, R.; Karagözler, A.A. Laccase immobilized polyacrylamide-alginate cryogel: A candidate for treatment of effluents. Process Biochem. 2021, 101, 137–146. [Google Scholar] [CrossRef]
- Kuster, E.; Williams, S.T. Selection of media for isolation of Streptomycetes. Nature 1964, 202, 928–929. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes--application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, G.L. Use of Dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Keesey, J. Biochemica Information: A Revised Biochemical Reference Source; Boehringer Mannheim Biochemicals: Indianapolis, IN, USA, 1987. [Google Scholar]
- Shekher, R.; Sehgal, S.; Kamthania, M.; Kumar, A. Laccase: Microbial sources, production, purification, and potential biotechnological applications. Enzym. Res. 2011, 2011, 217861. [Google Scholar]
- Viswanath, B.; Rajesh, B.; Janardhan, A.; Kumar, A.P.; Narasimha, G. Fungal laccases and their applications in bioremediation. Enzym. Res. 2014, 2014, 163242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myasoedova, N.M.; Renfeld, Z.V.; Podieiablonskaia, E.V.; Samoilova, A.S.; Chernykh, A.M.; Classen, T.; Pietruszka, J.; Kolomytseva, M.P.; Golovleva, L.A. Novel laccase—producing ascomycetes. Microbiology 2017, 86, 503–511. [Google Scholar] [CrossRef]
- Bagewadi, Z.K.; Mulla, S.I.; Ninnekar, H.Z. Purification and immobilization of laccase from Trichoderma harzianum strain HZN10 and its application in dye decolorization. J. Genet. Eng. Biotechnol. 2017, 15, 139–150. [Google Scholar] [CrossRef]
- Lassouane, F.; Ait-Amar, H.; Amrani, S.; Rodriguez-Couto, S. A promising laccase immobilization approach for Bisphenol A removal from aqueous solutions. Bioresour. Technol. 2019, 271, 360–367. [Google Scholar] [CrossRef]
- Othman, A.; Elshafei, A.; Elsayed, M.; Hassan, M. Decolorization of Cibacron Blue 3G-A Dye by Agaricus bisporus CU13 Laccase—Mediator System: A Statistical Study for Optimization via Response Surface Methodology. Annu. Res. Rev. Biol. 2018, 25, 1–13. [Google Scholar] [CrossRef]
- Reda, F.M.; Hassan, N.S.; El-Moghazy, A.-N. Decolorization of synthetic dyes by free and immobilized laccases from newly isolated strain Brevibacterium halotolerans N11 (KY883983). Biocatal. Agric. Biotechnol. 2018, 15, 138–145. [Google Scholar] [CrossRef]
- Sharifi-Bonab, M.; Rad, F.; Mehrabad, J.T. Preparation of laccase-graphene oxide nanosheet/alginate composite: Application for the removal of cetirizine from aqueous solution. J. Environ. Chem. Eng. 2016, 4, 3013–3020. [Google Scholar] [CrossRef]
- Kashefi, S.; Borghei, S.M.; Mahmoodi, N.M. Covalently immobilized laccase onto graphene oxide nanosheets: Preparation, characterization, and biodegradation of azo dyes in colored wastewater. J. Mol. Liq. 2019, 276, 153–162. [Google Scholar] [CrossRef]
- Bilal, M.; Jing, Z.; Zhao, Y.; Iqbal, H.M.N. Immobilization of fungal laccase on glutaraldehyde cross-linked chitosan beads and its bio-catalytic potential to degrade bisphenol A. Biocatal. Agric. Biotechnol. 2019, 19, 101174. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Substrate | Vmax (µmol min−1) * | Km (mM) |
|---|---|---|---|
| Free | |||
| ABTS | 3.05 | 0.28 | |
| Rb-2 | 5.67 | 0.18 | |
| Entrapped | |||
| ABTS | 7.54 | 6.00 | |
| Rb-2 | 1.19 | 1.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero-Soto, I.C.; Martínez-Pérez, R.B.; Rodríguez, J.A.; Camacho-Ruiz, R.M.; Barbachano-Torres, A.; Martín del Campo, M.; Napoles-Armenta, J.; Pliego-Sandoval, J.E.; Concha-Guzmán, M.O.; Angeles Camacho-Ruiz, M. Galactomannans for Entrapment of Gliomastix murorum Laccase and Their Use in Reactive Blue 2 Decolorization. Sustainability 2021, 13, 9019. https://doi.org/10.3390/su13169019
Romero-Soto IC, Martínez-Pérez RB, Rodríguez JA, Camacho-Ruiz RM, Barbachano-Torres A, Martín del Campo M, Napoles-Armenta J, Pliego-Sandoval JE, Concha-Guzmán MO, Angeles Camacho-Ruiz M. Galactomannans for Entrapment of Gliomastix murorum Laccase and Their Use in Reactive Blue 2 Decolorization. Sustainability. 2021; 13(16):9019. https://doi.org/10.3390/su13169019
Chicago/Turabian StyleRomero-Soto, Itzel C., Raúl B. Martínez-Pérez, Jorge A. Rodríguez, Rosa M. Camacho-Ruiz, Alejandra Barbachano-Torres, Martha Martín del Campo, Juan Napoles-Armenta, Jorge E. Pliego-Sandoval, María O. Concha-Guzmán, and María Angeles Camacho-Ruiz. 2021. "Galactomannans for Entrapment of Gliomastix murorum Laccase and Their Use in Reactive Blue 2 Decolorization" Sustainability 13, no. 16: 9019. https://doi.org/10.3390/su13169019