Morphological, Pathogenic and Toxigenic Variability in the Rice Sheath Rot Pathogen Sarocladium Oryzae

 and
and
Abstract
:
1. Introduction
2. Results
2.1. Morphological Characterisation and Growth Rate
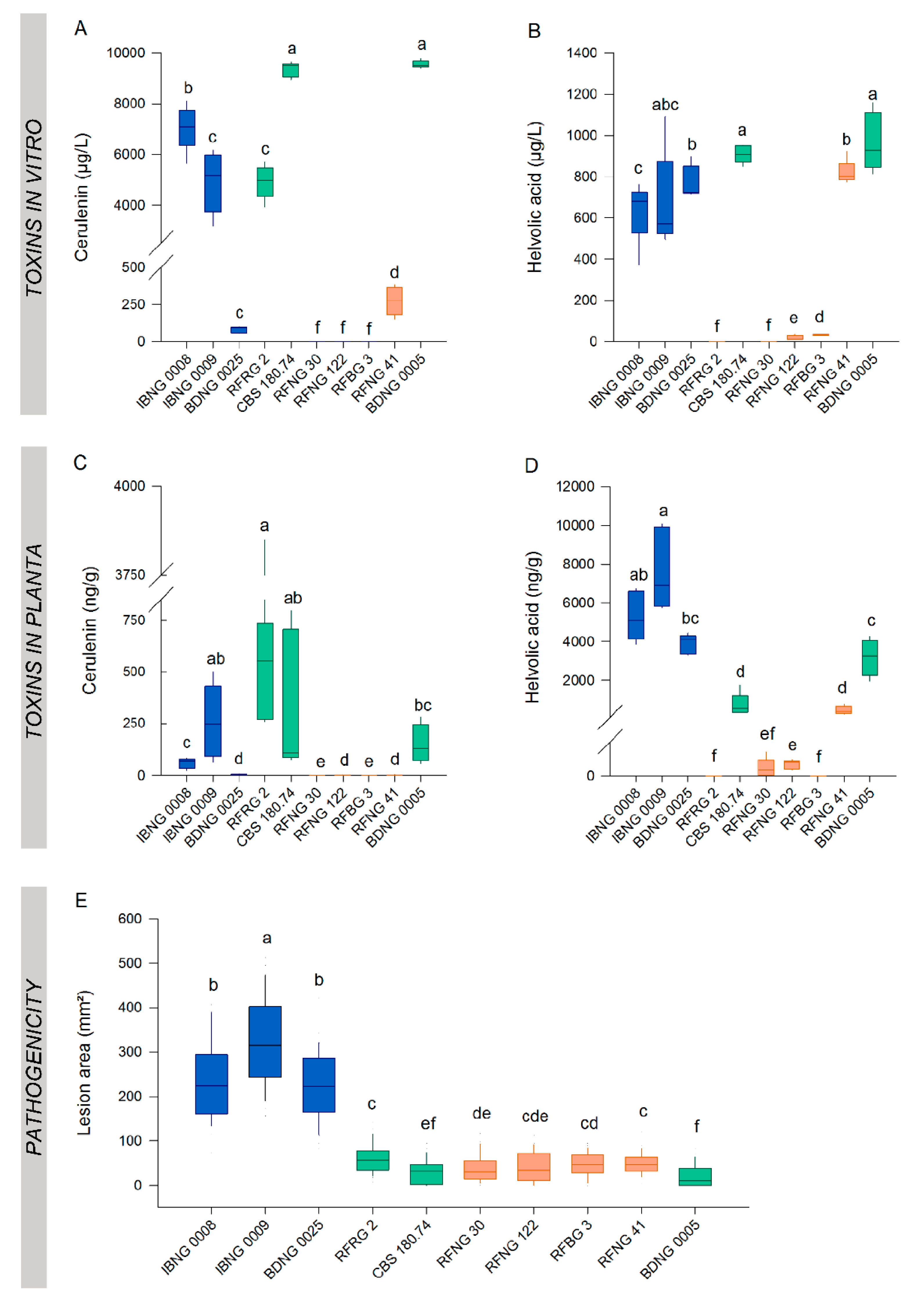
2.2. Pathogenicity
2.3. In Vitro Cerulenin and Helvolic Acid Production
2.4. Cerulenin and Helvolic Acid Production by Different Phenotypes of CBS180.74
2.5. A Comparison of the Toxin Production in Vitro and in Planta
2.6. Disease Severity Correlates with In Planta Helvolic Acid Production
3. Discussion
4. Materials and Methods
4.1. Fungal Isolates
4.2. Morphological Characterization
4.3. Pathogenicity Assays
4.4. Chemical Analysis
4.4.1. Extraction from PDA
4.4.2. Extraction from Liquid Culture
4.4.3. Extraction from Plant Material
4.4.4. Instrumental Analysis
4.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Origin | Phylogenic Group | Reference |
|---|---|---|---|
| CBS 180.74 | India | 1 | [33] |
| CBS 361.75 | Kenya | 1 | [31] |
| CBS 120.817 | Panama | 1 | [31] |
| CBS 485.80 | Australia | 1 | [31] |
| RFRG 2 | Rwanda, Rugeramigozi | 1 | [31] |
| BDNG 0004 | Nigeria, Badaggi | 1 | [31] |
| BDNG 0005 | Nigeria, Badaggi | 1 | [31] |
| BDNG 0007 | Nigeria, Badaggi | 1 | [31] |
| BDNG 0009 | Nigeria, Badaggi | 1 | [31] |
| BDNG 0012 | Nigeria, Badaggi | 1 | [31] |
| BDNG 0014 | Nigeria, Badaggi | 1 | [31] |
| BDNG 0022 | Nigeria, Badaggi | 1 | [31] |
| BDNG 0023 | Nigeria, Badaggi | 1 | [31] |
| IBNG 0011 | Nigeria, Ibadan | 1 | [31] |
| IBNG 0012 | Nigeria, Ibadan | 1 | [31] |
| IBNG 0013 | Nigeria, Ibadan | 1 | [31] |
| CBS 399.73 | India | 2 | [33] |
| CBS 101.61 | Mexico | 2 | [31] |
| RFNG 41 | Rwanda, Nyagatare and Rwagitama | 2 | [31] |
| RFNG 33 | Rwanda, Nyagatare and Rwagitama | 2 | [31] |
| RFNG 30 | Rwanda, Nyagatare and Rwagitama | 2 | [31] |
| RFNG 28 | Rwanda, Nyagatare and Rwagitama | 2 | [31] |
| RFNG 122 | Rwanda, Nyagatare and Rwagitama | 2 | [31] |
| RFBG 3 | Rwanda, Bugarama | 2 | [31] |
| RFBG 9 | Rwanda, Bugarama | 2 | [31] |
| CBS 414.81 | Nigeria | 3 | [33] |
| BDNG 0025 | Nigeria, Badaggi | 3 | [31] |
| IBNG 0002 | Nigeria, Ibadan | 3 | [31] |
| IBNG 0008 | Nigeria, Ibadan | 3 | [31] |
| IBNG 0009 | Nigeria, Ibadan | 3 | [31] |
| IBNG 0001 | Nigeria, Ibadan | 3 | [31] |
References
- Mohanty, S. Trends in global rice consumption. Rice Today 2013, 12, 44–45. [Google Scholar]
- Fageria, N. Yield physiology of rice. J. Plant Nutr. 2007, 30, 843–879. [Google Scholar] [CrossRef]
- FAO Rice Market Monitor (RMM); Food and Agriculture Organization United Nation: Rome, Italy, 2018.
- Saito, K.; Dieng, I.; Toure, A.; Somado, E.; Wopereis, M. Rice yield growth analysis for 24 African countries over 1960–2012. Glob. Food Sec. 2015, 5, 62–69. [Google Scholar] [CrossRef]
- Demont, M. Reversing urban bias in African rice markets: A review of 19 national rice development strategies. Glob. Food Sec. 2013, 2, 172–181. [Google Scholar] [CrossRef]
- Balamurugan, P.; Balasubramanian, V. Challenges and opportunities for increasing rice production in sub saharan Africa. J. Innov. Agric. 2017, 4, 1–10. [Google Scholar]
- Sakthivel, N. Sheath rot disease of rice: Current status and control strategies. In Major Fungal Diseases of Rice: Recent Advances; Sreenivasaprasad, S., Johnson, R., Eds.; Kluwer Academic Publishes: Puducherry, India, 2001; pp. 271–283. [Google Scholar]
- Mvuyekure, S.; Sibiya, J.; Derera, J.; Nzungize, J.; Nkima, G. Genetic analysis of mechanisms associated with inheritance of resistance to sheath rot of rice. Plant Breed. 2017, 136, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Mvuyekure, S.; Sibiya, J.; Derera, J.; Nzungize, J.; Nkima, G. Assessment of genetic diversity of rice based on SNP markers for selection of parents for sheath rot (Sarocladium oryzae) resistance breeding. S. Afr. J. Plant Soil 2018, 35, 51–59. [Google Scholar] [CrossRef]
- Bigirimana, V.; Hua, G.; Nyamangyoku, O.; Höfte, M. Rice Sheath Rot: An emerging ubiquitous destructive disease complex. Front. Plant Sci. 2015, 6, 1066. [Google Scholar] [CrossRef] [Green Version]
- Nandakumar, R.; Babu, S.; Viswanathan, R.; Sheela, J.; Raguchander, T.; Samiyappan, R. A new bio-formulation containing plant growth promoting rhizobacterial mixture for the management of sheath blight and enhanced grain yield in rice. Biocontrol 2001, 46, 493–510. [Google Scholar] [CrossRef]
- Ayyadurai, N.; Kirubakaran, S.; Srisha, S.; Sakthivel, N. Biological and molecular variability of Sarocladium oryzae, the sheath rot pathogen of rice (Oryza sativa L.). Curr. Microbiol. 2005, 50, 319–323. [Google Scholar] [CrossRef]
- Tschen, J.; Chen, L.; Hsieh, S.; Wu, T. Isolation and phytotoxic effects of helvolic acid from plant pathogenic fungus Sarocladium oryzae. Bot. Bull. Acad. Sin. 1997, 38, 251–256. [Google Scholar]
- Sakthivel, N.; Amudha, R.; Muthukrishnan, S. Production of phytotoxic metabolites by Sarocladium oryzae. Mycol. Res. 2002, 106, 609–614. [Google Scholar] [CrossRef]
- Hoagland, R. Phytotoxicity of microbial antibiotics helvolic and fusidic acids. Allelopath. J. 2009, 23, 167–174. [Google Scholar]
- Ratnaweera, P.; Williams, D.; de Silva, E.; Wijesundera, R.; Dalisay, D.; Andersen, R. Helvolic acid, an antibacterial nortriterpenoid from a fungal endophyte, Xylaria sp. of orchid Anoectochilus setaceus endemic to Sri Lanka. Mycology 2014, 5, 23–28. [Google Scholar] [CrossRef]
- Hittalmani, S.; Mahesh, H.; Mahadevaiah, C.; Prasannakumar, M. De novo genome assembly and annotation of rice sheath rot fungus Sarocladium oryzae reveals genes involved in Helvolic acid and Cerulenin biosynthesis pathways. BMC Genomics 2016, 17, 271. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, M.; Patra, M.; Albrecht, D.; Chen, D.; Nicolaou, K.; Metzler-Nolte, N.; Bandow, J. Proteomic signature of fatty acid biosynthesis inhibition available for in vivo mechanism-of-action studies. Antimicrob. Agents Chemother. 2011, 55, 2590–2596. [Google Scholar] [CrossRef] [Green Version]
- Gnanamanickam, S.; Mew, T. Interactions between Sarocladium oryzae and stem attacking fungal pathogens of rice. Plant. Soil 1991, 138, 213–219. [Google Scholar] [CrossRef]
- Côrtes, M.; Silva-Lobo, V.; Filippi, M.; Lima, D.; Prabhu, A. Potential for using crude extract of Sarocladium oryzae for suppression of rice blast. Trop. Plant. Pathol. 2014, 39, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Brakhage, A. Regulation of fungal secondary metabolism. Nat. Rev. Microbiol. 2013, 11, 21–32. [Google Scholar] [CrossRef]
- Fox, E.; Howlett, B. Secondary metabolism: Regulation and role in fungal biology. Curr. Opin. Microbiol. 2008, 11, 481–487. [Google Scholar] [CrossRef]
- Ryan, M.; Bridge, P.; Smith, D.; Jeffries, P. Phenotypic degeneration occurs during sector formation in Metarhizium anisopliae. J. Appl. Microbiol. 2002, 93, 163–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamaruzzaman, M.; He, G.; Wu, M.; Zhang, J.; Yang, L.; Chen, W.; Li, G. A novel partitivirus in the hypovirulent isolate qt5-19 of the plant pathogenic fungus Botrytis cinerea. Viruses 2019, 11, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, V.; Mascarin, G.M.; da Silva Lopes, M.; Alves, M.C.D.F.; Rezende, J.M.; Gatti, M.S.V.; Dunlap, C.A.; Delalibera Júnior, Í. Identification of double-stranded RNA viruses in Brazilian strains of Metarhizium anisopliae and their effects on fungal biology and virulence. Plant. Gene 2017, 11, 49–58. [Google Scholar] [CrossRef]
- Shah, F.; Butt, T. Influence of nutrition on the production and physiology of sectors produced by the insect pathogenic fungus Metarhizium anisopliae. FEMS Microbiol. Lett. 2005, 250, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawe, A.L.; Nuss, D.L. Hypoviruses and Chestnut Blight: Exploiting viruses to understand and modulate fungal pathogenesis. Annu. Rev. Genet. 2001, 35, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lee, J.; Yun, S.; So, K.; Ko, Y.; Kim, Y.; Park, S.; Kim, D. A mutant of the Bck1 homolog from Cryphonectria parasitica resulted in sectorization with an impaired pathogenicity. Mol. Plant Microbe Interact. 2016, 29, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Saravanakumar, D.; Lavanya, N.; Muthumeena, K.; Raguchander, T.; Samiyappan, R. Fluorescent pseudomonad mixtures mediate disease resistance in rice plants against sheath rot (Sarocladium oryzae) disease. Biocontrol 2009, 54, 273–286. [Google Scholar] [CrossRef]
- Chowdhury, T.; Mian, M.; Mia, M.; Rafii, M.; Latif, M. Agro-ecological variations of sheath rot disease of rice caused by Sarocladium oryzae and DNA fingerprinting of the pathogen’s population structure. Genet. Mol. Res. 2015, 14, 18140–18152. [Google Scholar] [CrossRef]
- Afolabi, O.O.; Peeters, K.J.; Onwughalu, J.; Oyetunji, O.; Oludare, A.; Ogunbayo, A.; Oni, F.; Höfte, M. Characterization of Sarocladium oryzae and pathogenic Fusarium species associated with sheath rot disease of rice in Mali and Nigeria. (unpublished; manuscript in preparation).
- Bigirimana, V.; Hua, G.K.H.; Bertier, L.; Peeters, K.J.; Nyamangyoku, O.I.; Höfte, M. Sarocladium oryzae and Fusarium spp. are associated with rice sheath rot in Rwanda. (unpublished; manuscript in preparation).
- Giraldo, A.; Gené, J.; Sutton, D.; Madrid, H.; de Hoog, G.; Cano, J.; Crous, P.; Guarro, J. Phylogeny of Sarocladium (Hypocreales). Persoonia 2015, 34, 10–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, M.; Amudha, R.; Jayachandran, S.; Sakthivel, N. Detection and quantification of phytotoxic metabolites of Sarocladium oryzae in sheath rot-infected grains of rice. Lett. Appl. Microbiol. 2002, 34, 398–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Butt, T.; St Leger, R. Colony sectorization of Metarbizium anisopliae is a sign of ageing. Microbiology 2005, 151, 3223–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal-plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Pischetsrieder, M.; St Leger, R.J.; Wang, C. Associated links among mtDNA glycation, oxidative stress and colony sectorization in Metarhizium anisopliae. Fungal Genet. Biol. 2008, 45, 1300–1306. [Google Scholar] [CrossRef]
- Heller, J.; Tudzynski, P. Reactive oxygen species in phytopathogenic fungi: Signaling, development, and disease. Annu. Rev. Phytopathol. 2011, 49, 369–390. [Google Scholar] [CrossRef]
- García-Pedrajas, M.D.; Cañizares, M.C.; Sarmiento-Villamil, J.L.; Jacquat, A.G.; Dambolena, J.S. Mycoviruses in biological control: From basic research to field implementation. Phytopathology 2019, 109, 1828–1839. [Google Scholar] [CrossRef]
- Brusini, J.; Robin, C. Mycovirus transmission revisited by in situ pairings of vegetatively incompatible isolates of Cryphonectria parasitica. J. Virol. Methods 2013, 187, 435–442. [Google Scholar] [CrossRef]
- Bigirimana, V.P. Characterization of Sheath Rot Pathogens from Major Rice-Growing Areas in Rwanda; Ghent University: Gent, Belgium, 2016. [Google Scholar]
- Sakthivel, N.; Gnanamanickam, S. Evaluation of Pseudomonas fluorescens for suppression of Sheath Rot disease and for enhancement of grain yields in rice (Oryza sativa L.). Appl. Environ. Microbiol. 1987, 53, 2056–2059. [Google Scholar] [CrossRef] [Green Version]
- Perneel, M.; Heyrman, J.; Adiobo, A.; De Maeyer, K.; Raaijmakers, J.; De Vos, P.; Höfte, M. Characterization of CMR5c and CMR12a, novel fluorescent Pseudomonas strains from the cocoyam rhizosphere with biocontrol activity. J. Appl. Microbiol. 2007, 103, 1007–1020. [Google Scholar] [CrossRef] [Green Version]
- Omura, S. The antibiotic cerulenin, a novel tool for biochemistry as an inhibitor of fatty acid synthesis. Bacteriol. Rev. 1976, 40, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Haeck, A.; Van Langenhove, H.; Harinck, L.; Kyndt, T.; Gheysen, G.; Höfte, M.; Demeestere, K. Trace analysis of multi-class phytohormones in Oryza sativa using different scan modes in high-resolution Orbitrap mass spectrometry: Method validation, concentration levels, and screening in multiple accessions. Anal. Bioanal. Chem. 2018, 410, 4527–4539. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peeters, K.J.; Haeck, A.; Harinck, L.; Afolabi, O.O.; Demeestere, K.; Audenaert, K.; Höfte, M. Morphological, Pathogenic and Toxigenic Variability in the Rice Sheath Rot Pathogen Sarocladium Oryzae. Toxins 2020, 12, 109. https://doi.org/10.3390/toxins12020109
Peeters KJ, Haeck A, Harinck L, Afolabi OO, Demeestere K, Audenaert K, Höfte M. Morphological, Pathogenic and Toxigenic Variability in the Rice Sheath Rot Pathogen Sarocladium Oryzae. Toxins. 2020; 12(2):109. https://doi.org/10.3390/toxins12020109
Chicago/Turabian StylePeeters, Kaat José, Ashley Haeck, Lies Harinck, Oluwatoyin Oluwakemi Afolabi, Kristof Demeestere, Kris Audenaert, and Monica Höfte. 2020. "Morphological, Pathogenic and Toxigenic Variability in the Rice Sheath Rot Pathogen Sarocladium Oryzae" Toxins 12, no. 2: 109. https://doi.org/10.3390/toxins12020109