
Pseudozyma aphidis Enhances Cucumber and Tomato Plant Growth and Yield


Department of Plant Pathology and Microbiology, The Robert H. Smith Faculty of Agriculture, Food and Environment, The Hebrew University of Jerusalem, Rehovot 76100, Israel
*
Author to whom correspondence should be addressed.
Agronomy 2022, 12(2), 383; https://doi.org/10.3390/agronomy12020383
Submission received: 9 January 2022
/
Revised: 25 January 2022
/
Accepted: 1 February 2022
/
Published: 3 February 2022
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The need to supply food for a growing world population, while protecting the environment by using sustainable agriculture, presents a major challenge, in terms of decreasing the use of chemical fertilizers and pesticides. The application of Pseudozyma aphidis cells onto crop plants considerably reduced infection by several pathogens, as well as significantly enhancing plant growth and yield. It was demonstrated in the current work that P. aphidis can enhance cucumber and tomato growth, both in vitro and in the greenhouse. P. aphidis was active when applied either via spray treatment or by seed coating. A significant promotion effect of P. aphidis on tomato and cucumber growth, flowering, and tomato yield and ripening, as compared to control plants, was demonstrated. Specifically, a 10–30% increase in tomato and cucumber plant height and leaf number, a 45% increase in average fruit weight, a 120% increase in tomato fruit ripening, and a flowering time advanced by two weeks were demonstrated. In conclusion, P. aphidis can significantly enhance the growth and yield of crop plants and can, thus, be used as bio-fertilizer.
1. Introduction
The need to supply food for a growing world population, while protecting the environment by using sustainable agriculture, presents a great challenge, in terms of decreasing the use of chemical fertilizers and pesticides. Beneficial microorganisms (BMO) represent a potential resource for environmentally friendly and efficient growth-promoting compounds. BMO are reported to affect plants growth and yields by an array of mechanisms. For example, Phyllobacterium spp. have been shown to affect the plant’s nitrogen metabolism [1], Rhizobacterium spp. were shown to be capable of promoting growth by fixation of nitrogen and by inducing a starvation-like transcriptional response [2], while other microorganisms were demonstrated to secrete plant hormones [3,4]. Fungal biocontrol agents correspond to a BMO group that controls plant pathogens but were also reported to have growth promotion capabilities [5,6,7,8,9]. These agents can enhance growth in a direct or an indirect manner, which is largely related to their antagonistic activity towards phytopathogens, but may be also be attributed to their ability to induce resistance to abiotic stresses, such as salinity or drought, or by detoxification and elimination of heavy metals and other contaminating compounds. Piriformospora indica, for example, enhances yield in barley, not only by activating the plant defense response, but also by reprogramming the plant cell for salt-stress tolerance [5]. P. indica has also been suggested to function by interacting with specific plant cell receptors [7], and it was recently reported that this fungus might stimulate the growth response via a specific molecular pathway involving the activation of phosphatidic acid synthesis in Arabidopsis [6]. Other examples are provided by the biocontrol agent Trichoderma harzianum that increases phosphorous and other nutrient solubilization [9,10], and T. virens that enhance biomass and lateral root formation via auxin-dependent mechanisms in Arabidopsis [8].
It is fair to assume that different beneficial species possess unique sets of mechanisms to enhance the growth of their hosts, and which have yet to be discovered and do not necessarily fall within the commonly accepted paradigms of growth promotion mechanisms. Moreover, BMOs usually use more than one mechanism to enhance plant health or growth. Accordingly, we isolated a unique biologically-active strain of Pseudozyma aphidis (designated isolate L12, Rehovot, Israel, 2004) from strawberry leaves. In contrast to the findings by Avis et al. [11], who reported that P. aphidis, isolated from aphid secretions (isolate CBS 517.83), is not associated with colony collapse of powdery mildew, our strain (isolate L12) exhibited antifungal activity on developed plants and was shown to act as a suppressor of disease symptoms induced by various fungal and bacterial pathogens [12,13,14]. Furthermore, published data indicate that P. aphidis L12 may function by directly secreting antifungal compounds or by parasitism and interaction with pathogenic fungi [15]. Additionally, experimental data suggest that P. aphidis L12 may also function indirectly by activating the plant systemic defense response and by colonizing and competing for space and nutrients on the host plant surface [12]. In the current research, we demonstrated the ability of P. aphidis to promote the growth and yield parameters of its host plant. The use of an environmentally friendly fertilizer mediated by BMOs such as P. aphidis will benefit farmers, consumers, and the environment.
2. Materials and Methods
2.1. Microorganism and Culture Conditions
A Pseudozyma aphidis isolate L12 (Rehovot, Israel, 2004) was maintained on potato dextrose agar (PDA; Difco, Bordeaux, France) at 25 °C and transferred weekly to fresh medium. For liquid cultures, P. aphidis was grown in potato dextrose broth (PDB; Difco, Bordeaux, France) for 4–7 days at 25 °C on a rotary shaker at 150 rpm.
2.2. Plant Growth and Yield Promotion
An Erlenmeyer flask containing 200 mL of PDB was inoculated with P. aphidis L12, and incubated for 2 days in an orbital shaker at 25 °C and 150 rpm. After incubation, the P. aphidis culture was filtered with sterile Miracloth (Merck, Darmstadt, Germany) and adjusted to a cell density of 108 spores/mL. Two-week-old cucumber (cultivar ‘Beit alpha’) or tomato (cultivar ‘Sharlot’) seedlings were sprayed with a P. aphidis suspension or distilled water (control) once a week. The inoculated plants were incubated in a growth chamber at 25 °C at 90% relative humidity with 16 h of daylight. Two weeks after the first inoculation, cucumber or tomato seedling fresh weight, root and hypocotyl length, as well as number of flowers, open flowers, and leaves, were recorded by double-blind naked eye counting, or using digital electronic scales or a ruler.
Tomato (cultivar ‘Micro-Tom’) seedlings were also grown in the same green house for 10 weeks and growth parameters (number of leaves, plant height, number of flowers, and open flowers) and yield (fruit weight, number of fruits, and ripped fruits) were recorded. Experiments were repeated at least twice.
2.3. Promotion of Root Development
For analysis of the ability to promote root growth, cucumber (cultivar ‘Beit alpha’) or tomato (cultivar ‘Sharlot’) seeds were disinfected in a solution of 3% commercial sodium hypochlorite (NaOCl) for 30 min and rinsed twice with water for 30 min. Then, disinfected seeds were dipped into P. aphidis L12 spore suspension (108 spores/mL) for 1–2 min and dried in the hood, and then incubated on petri dishes containing sterile filter paper imbibed with sterile distilled water for 6–7 days at 25 °C in the dark, with the application of sterile distilled water on alternating days. After incubation, the fresh weight and root length of 10 germinated seeds was recorded. Experiments were performed at least twice.
2.4. Statistical Analysis
A Tukey-Kramer HSD test was used, only after verification that data were normally distributed and sample variances were equal. Significance was accepted at p < 0.05. All experiments described here are representative of at least two independent experiments and presented the same pattern of results.
3. Results
3.1. P. aphidis-Mediated Cucumber Plant Growth-Promoting Effects
Pseudozyma aphidis was tested for its ability to stimulate the growth of cucumber seedlings. Significant differences were detected in terms of plant height (Figure 1a), absolute number of flowers (Figure 1b), and number of blooming flowers (Figure 1c), as well as the number of leaves (Figure 1d) on cucumber plants spray-treated with P. aphidis, as compared with distilled water-sprayed plants. The fresh weight of P. aphidis-treated plants was slightly higher, although not significantly so (Supplementary Figure S1).
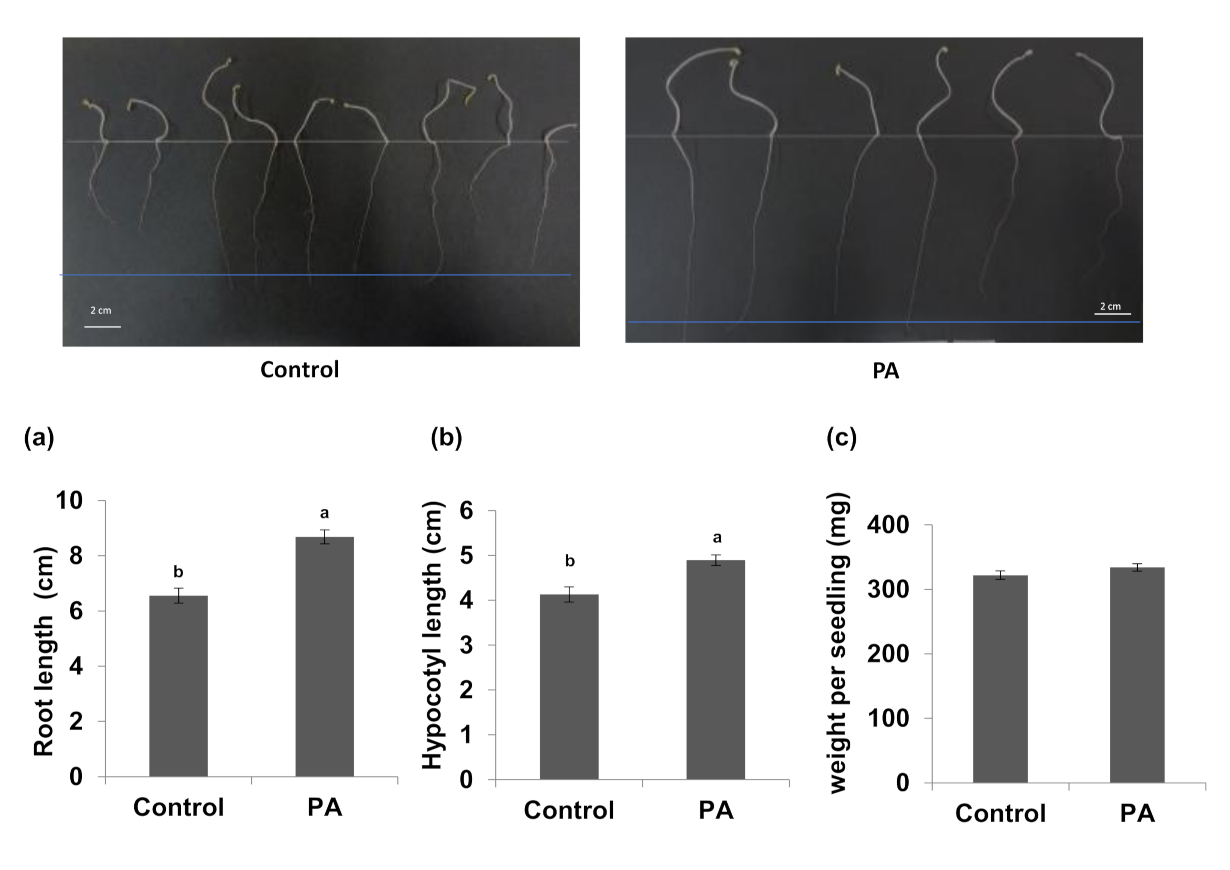
When an in vitro root development assay was carried out on cucumber seedlings after seed treatment with P. aphidis, we observed significant differences in root length (Figure 2a), as well as in aerial plant parts, such as on hypocotyl height (Figure 2b) and seedlings fresh weight (Figure 2c), relative to control plants.
3.2. Growth Enhancement of Tomato Seedlings In Vitro
An in vitro assay of tomato seedling growth enhancement was carried out after seeds were coated with wild type P. aphidis or water, as a control. Incremental increases in average root (Figure 3a) and hypocotyl (Figure 3b) lengths, in addition to average seedling fresh weight (Figure 3c) were observed one week after application of P. aphidis.
3.3. Tomato Plant Growth and Yield Enhancement
Pseudozyma aphidis significantly increased tomato growth and yield in the greenhouse. Micro-Tom tomatoes spray-treated with P. aphidis demonstrated significant increases in leaf numbers (Figure 4a), plant height (Figure 4b), and number of flowers (Figure 4c). Significant effects of P. aphidis treatment on average fruit weight (Figure 5a), number of fruits per plants (Figure 5b), total fruits weight per plant (Figure 5c) and the percentage of ripened fruits per plant (Figure 5d), were also observed.
4. Discussion
In our previous studies, we demonstrated that P. aphidis (isolate L12) relies on complex modes of action to control plant pathogens, including antibiosis, competition, induced resistance, and parasitism [12,13,14,15]. In the present study, it was demonstrated for the first time that epiphytic yeast-like P. aphidis (isolate L12) also affects the growth and yield promotion of tomato and cucumber plants.
Plant growth promotion has typically been investigated using rhizosphere-associated bacteria [16,17]. However, fungi and yeast possess analogous beneficial bioactivities as plant growth-promoting rhizobacteria (PGPR) [8,18,19,20,21,22]. Beneficial fungi are major players in natural and agricultural habitats and ecosystems, where they are involved in organic matter recycling, nutrient availability, and plant health and growth [23,24]. Many reports have demonstrated that arbuscular mycorrhizal fungi (AMF), alone or in combination with plant growth promoting rhizobacteria (PGPRs), such as Bacillus or Pseudomonas, increase nutrient availably for the host plant and promote its development and yield [19,23,25,26].
Yeast are also able to produce a variety of biologically active compounds (e.g., phytohormones, vitamins, amino acids, etc.) that have stimulating effects on the plant and root growth development, helping to increase plant productivity [27,28,29]. In addition, yeast, especially members of the Pseudozyma group, produce different antifungal substances that are involved in its biocontrol capability to reduce plant diseases caused by different fungi [15,30,31], and may also be involved in plant and root growth promotion as a mode of action. Here, we demonstrated that P. aphidis, not only enhances plant protection, but also the growth and yield of different crop plants. It was demonstrated that P. aphidis can significantly enhance plant growth and yield after spraying application onto seedlings grown in a greenhouse. A 13% increase in cucumber plant height, 35% more flowers, 11% more leaves, and 44% more open flowers were measured two weeks after spray application (Figure 1). These increases may be indirectly due to the repressing of pathogens and increasing plant health or by directly improving plant macro- and micro-nutrient uptake or secretion of plant hormones. Tomato seedlings in greenhouse spray-treated with P. aphidis were 30% higher, had 30% more leaves, and flowering time was advanced by two weeks (Figure 4). Transition to flowering in tomato plants is affected by leaf number, with 6–12 leaves usually needed before the vegetative apex is terminated by inflorescence (cultivar-dependent) [32]. Thus, growth promotion and leaf access in P. aphidis-treated plants can explain the advancement in flowering time. When looking at yield parameters, treated tomato plants had an 18% increase in average fruit weight, a 45% increase in fruit weight per plant, a 17% increase in the number of fruits per plant, and a 120% increase in ripened fruit per plant (Figure 5). We assume that the increase in leaf number generated a higher photosynthesis area that eventually resulted in earlier fruiting and higher yields, as also suggested by Xu HL et al. (2001) [33]. Similarly to our results suggesting that P. aphidis can serve dual roles as a biocontrol agent and a plant growth-promoting fungus, other studies on plant growth-promoting fungi, such as arbuscular mycorrhizal fungi or the yeast Torulaspora globosa, have also reported that those growth promoting fungi can suppress plant pathogens [34,35,36,37,38,39,40]. Protecting the host from infection or reducing infection load itself can contribute not only to plant health but also indirectly to better growth and yield parameters.
The current study also demonstrated that seed-coating with P. aphidis enhanced root and shoot elongation upon growth in sterile conditions (Figure 2 and Figure 3). A 37% increase in root length, a 112% increase in hypocotyl length, and a 71% increase in the fresh weight of cucumber seedlings six days after seed treatment (Figure 2) were observed. Seed-coating of tomato seeds reveled a 32% increase in root length and a 18% increase in hypocotyl length one week after treatment (Figure 3). Although P. aphidis is known to be epiphytic, a recent study in our lab demonstrated that it can also be an endophyte (Alster et al., 2022, Microbiol. Spectr., in press). This observation is also supported by the ability of P. aphidis to be active after application by seed-coating. Seed-coating with chemicals is widely used to control against pathogens [41,42,43]. Since the use of chemicals can negatively affect the environment, [44,45] bio-fertilizers and seed-coating with beneficial microorganisms have recently gained attention as promising eco-friendly methods that can be used in both conventional and organic agriculture practices to reduce plant disease and enhance plant growth [26,46,47,48,49,50,51,52,53]
In summary, P. aphidis can promote plant growth, acting directly on plants, as reflected by the augmentation of the various measured plant traits, which were demonstrated in the current report and need to be further characterized, and by indirect mechanisms, such as antimicrobial activity, as demonstrated in our previous work [13,14,15,54]. The growth-promoting ability of P. aphidis demonstrated in the current work on tomato and cucumber plants may also prove to be the case with other crop plants. The use of fungi as bio-fertilizers in agriculture can be beneficial for the environment and for sustainable agriculture.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy12020383/s1, Figure S1: Effects of P. aphidis on cucumber seedling growth promotion.
Author Contributions
Conceptualization, M.L.; methodology, I.S. and C.E.C.; validation, M.L., C.E.C. and I.S.; formal analysis, M.L.; C.E.C. and I.S.; investigation, C.E.C. and I.S.; writing—original draft preparation, M.L.; writing—review and editing, M.L.; supervision, M.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Israel Science Foundation, grant number 1375/14.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mantelin, S.; Desbrosses, G.; Larcher, M.; Tranbarger, T.J.; Cleyet-Marel, J.-C.; Touraine, B. Nitrate-dependent control of root architecture and N nutrition are altered by a plant growth-promoting Phyllobacterium sp. Planta 2006, 223, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Schwachtje, J.; Karojet, S.; Thormählen, I.; Bernholz, C.; Kunz, S.; Brouwer, S.; Schwochow, M.; Köhl, K.; Van Dongen, J.T. A Naturally associated Rhizobacterium of Arabidopsis thaliana induces a starvation-like transcriptional response while promoting growth. PLoS ONE 2011, 6, e29382. [Google Scholar] [CrossRef] [PubMed]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenblueth, M.; Martinez-Romero, E. Bacterial endophytes and their interactions with hosts. Mol. Plant-Microbe Interact. 2006, 19, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Waller, F.; Achatz, B.; Baltruschat, H.; Fodor, J.; Becker, K.; Fischer, M.; Heier, T.; Hückelhoven, R.; Neumann, C.; von Wettstein, D.; et al. The endophytic fungus Piriformospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield. Proc. Natl. Acad. Sci. USA 2005, 102, 13386–13391. [Google Scholar] [CrossRef] [Green Version]
- Camehl, I.; Drzewiecki, C.; Vadassery, J.; Shahollari, B.; Sherameti, I.; Forzani, C.; Munnik, T.; Hirt, H.; Oelmüller, R. The OXI1 kinase pathway mediates Piriformospora indica-induced growth promotion in Arabidopsis. PLoS Pathog. 2011, 7, e1002051. [Google Scholar] [CrossRef]
- Vadassery, J.; Ranf, S.; Drzewiecki, C.; Mithofer, A.; Mazars, C.; Scheel, D.; Lee, J.; Oelmuller, R. A cell wall extract from the endophytic fungus Piriformospora indica promotes growth of Arabidopsis seedlings and induces intracellular calcium elevation in roots. Plant J. Cell Mol. Biol. 2009, 59, 193–206. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Cortés-Penagos, C.; López-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [Green Version]
- Yedidia, I.; Srivastva, A.K.; Kapulnik, Y.; Chet, I. Effect of Trichoderma harzianum on microelement concentrations and increased growth of cucumber plants. Plant Soil 2001, 235, 235–242. [Google Scholar] [CrossRef]
- Li, R.-X.; Cai, F.; Pang, G.; Shen, Q.-R.; Li, R.; Chen, W. Solubilisation of phosphate and micronutrients by Trichoderma harzianum and its relationship with the promotion of tomato plant growth. PLoS ONE 2015, 10, e0130081. [Google Scholar] [CrossRef] [Green Version]
- Avis, T.J.; Caron, S.J.; Boekhout, T.; Hamelin, R.C.; Bélanger, R.R. Molecular and physiological analysis of the powdery mildew antagonist Pseudozyma flocculosa and related fungi. Phytopathology 2001, 91, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Buxdorf, K.; Rahat, I.; Levy, M. Pseudozyma aphidis induces ethylene-independent resistance in plants. Plant Signal. Behav. 2013, 8, e26273. [Google Scholar] [CrossRef] [PubMed]
- Barda, O.; Shalev, O.; Alster, S.; Buxdorf, K.; Gafni, A.; Levy, M. Pseudozyma aphidis induces salicylic-acid-independent resistance to Clavibacter michiganensis in tomato plants. Plant Dis. 2015, 99, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Buxdorf, K.; Rahat, I.; Gafni, A.; Levy, M. The epiphytic fungus Pseudozyma aphidis induces jasmonic acid- and salicylic acid/nonexpressor of PR1-independent local and systemic resistance. Plant Physiol. 2013, 161, 2014–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gafni, A.; Calderón, E.C.; Harris, R.; Buxdorf, K.; Dafa-Berger, A.; Zeilinger-Reichert, E.; Levy, M. Biological control of the cucurbit powdery mildew pathogen Podosphaera xanthii by means of the epiphytic fungus Pseudozyma aphidis and parasitism as a mode of action. Front. Plant Sci. 2015, 6, 132. [Google Scholar] [CrossRef] [Green Version]
- Compant, S.; Van Der Heijden, M.G.; Sessitsch, A. Climate change effects on beneficial plant-microorganism interactions. FEMS Microbiol. Ecol. 2010, 73, 197–214. [Google Scholar] [CrossRef]
- Pérez-Montaño, F.; Alias-Villegas, C.; Bellogín, R.A.; del Cerro, P.; Espuny, M.R.; Jiménez-Guerrero, I.; López-Baena, F.J.; Ollero, F.; Cubo, T. Plant growth promotion in cereal and leguminous agricultural important plants: From microorganism capacities to crop production. Microbiol. Res. 2014, 169, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, T.; Banik, A.; Mukhopadhyay, S.K. Plant growth-promoting traits of a thermophilic strain of the Klebsiella group with its effect on rice plant growth. Curr. Microbiol. 2020, 77, 2613–2622. [Google Scholar] [CrossRef]
- Mukherjee, A.; Verma, J.P.; Gaurav, A.K.; Chouhan, G.K.; Patel, J.S.; Hesham, A.E.-L. Yeast a potential bio-agent: Future for plant growth and postharvest disease management for sustainable agriculture. Appl. Microbiol. Biotechnol. 2020, 104, 1497–1510. [Google Scholar] [CrossRef]
- Ramadan, A.; Mohamad, H.; Saad, A. Effect of soil amendment with yeasts as bio-fertilizers on the growth and productivity of sugar beet. Afr. J. Agric. Res. 2013, 8, 46–56. [Google Scholar] [CrossRef]
- Ignatova, L.V.; Brazhnikova, Y.V.; Berzhanova, R.Z.; Mukasheva, T.D. Plant growth-promoting and antifungal activity of yeasts from dark chestnut soil. Microbiol. Res. 2015, 175, 78–83. [Google Scholar] [CrossRef]
- Ignatova, L.; Brazhnikova, Y.; Berzhanova, R.; Mukasheva, T. The effect of application of micromycetes on plant growth, as well as soybean and barley yields. Acta Biochim. Pol. 2015, 62, 669–675. [Google Scholar] [CrossRef] [PubMed]
- de la Hoz, J.P.; Rivero, J.; Azcón-Aguilar, C.; Urrestarazu, M.; Pozo, M.J. Mycorrhiza-induced resistance against foliar pathogens is uncoupled of nutritional effects under different light intensities. J. Fungi 2021, 7, 402. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Zapata, J.A.; Marrufo-Zapata, D.; Guadarrama, P.; Carrillo-Sánchez, L.; Hernández-Cuevas, L.; Caamal-Maldonado, A. Impact of weed control on arbuscular mycorrhizal fungi in a tropical agroecosystem: A long-term experiment. Mycorrhiza 2012, 22, 653–661. [Google Scholar] [CrossRef]
- Vafadar, F.; Amooaghaie, R.; Otroshy, M. Effects of plant-growth-promoting rhizobacteria and arbuscular mycorrhizal fungus on plant growth, stevioside, NPK, and chlorophyll content of Stevia rebaudiana. J. Plant 2014, 9, 128–136. [Google Scholar] [CrossRef]
- Yadav, A.N.; Kour, D.; Kaur, T.; Devi, R.; Yadav, N. Agriculturally Important Fungi for Crop Productivity: Current Research and Future Challenges. In Agriculturally Important Fungi for Sustainable Agriculture: Volume 1: Perspective for Diversity and Crop Productivity; Yadav, A.N., Mishra, S., Kour, D., Yadav, N., Kumar, A., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 275–286. [Google Scholar]
- El-Tarabily, K.A.; Nassar, A.H.; Hardy, G.E.; Sivasithamparam, K. Plant growth promotion and biological control of Pythium aphanidermatum, a pathogen of cucumber, by endophytic actinomycetes. J. Appl. Microbiol. 2009, 106, 13–26. [Google Scholar] [CrossRef]
- Xin, G.; Glawe, D.; Doty, S.L. Characterization of three endophytic, indole-3-acetic acid-producing yeasts occurring in Populus trees. Mycol. Res. 2009, 113, 973–980. [Google Scholar] [CrossRef]
- Botha, A. The importance and ecology of yeasts in soil. Soil Biol. Biochem. 2011, 43, 1–8. [Google Scholar] [CrossRef]
- Hammami, W.; Castro, C.Q.; Rémus-Borel, W.; Labbé, C.; Bélanger, R.R. Ecological basis of the interaction between Pseudozyma flocculosa and powdery mildew fungi. Appl. Environ. Microbiol. 2011, 77, 926–933. [Google Scholar] [CrossRef] [Green Version]
- Teichmann, B.; Labbé, C.; Lefebvre, F.; Bölker, M.; Linne, U.; Bélanger, R.R. Identification of a biosynthesis gene cluster for flocculosin a cellobiose lipid produced by the biocontrol agent Pseudozyma flocculosa. Mol. Microbiol. 2011, 79, 1483–1495. [Google Scholar] [CrossRef]
- Sawhney, V.K.; Greyson, R.I. On the initiation of the inflorescence and floral organs in tomato (Lycopersicon esculentum). Can. J. Bot. 1972, 50, 1493–1495. [Google Scholar] [CrossRef]
- Xu, H.-L.; Wang, R.; Mridha, M.A.U. Effects of Organic Fertilizers and a Microbial Inoculant on Leaf Photosynthesis and Fruit Yield and Quality of Tomato Plants. J. Crop Prod. 2001, 3, 173–182. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic press: Cambridge, MA, USA, 2010. [Google Scholar]
- Dehne, H.W. Interaction between Vesicular-Arbuscular Mycorrhizal Fungi and Plant Pathogens; The American Phytopathological Society: Saint Paul, MN, USA, 1982. [Google Scholar]
- Azcón-Aguilar, C.; Barea, J. Arbuscular mycorrhizas and biological control of soil-borne plant pathogens—An overview of the mechanisms involved. Mycorrhiza 1997, 6, 457–464. [Google Scholar] [CrossRef]
- Hyakumachi, M. Plant-growth-promoting fungi from turfgrass rhizosphere with potential for disease suppression. Soil Microorg. 1994, 44, 53–68. [Google Scholar] [CrossRef]
- Meera, M.; Shivanna, M.; Kageyama, K.; Hyakumachi, M. Plant growth promoting fungi from zoysiagrass rhizosphere as potential inducers of systemic resistance in cucumbers. Phytopathology 1994, 84, 1399–1406. [Google Scholar] [CrossRef]
- Shivanna, M.; Meera, M.; Hyakumachi, M. Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat. Crop Prot. 1996, 15, 497–504. [Google Scholar] [CrossRef]
- Nutaratat, P.; Srisuk, N.; Arunrattiyakorn, P.; Limtong, S. Plant growth-promoting traits of epiphytic and endophytic yeasts isolated from rice and sugar cane leaves in Thailand. Fungal Biol. 2014, 118, 683–694. [Google Scholar] [CrossRef]
- Ehsanfar, S.; Modarres-Sanavy, S. Crop protection by seed coating. Commun. Agric. Appl. Biol. Sci. 2005, 70, 225–229. [Google Scholar]
- Elzein, A.; Heller, A.; Ndambi, B.; De Mol, M.; Kroschel, J.; Cadisch, G. Cytological investigations on colonization of sorghum roots by the mycoherbicide Fusarium oxysporum f. sp. strigae and its implications for Striga control using a seed treatment delivery system. Biol. Control 2010, 53, 249–257. [Google Scholar] [CrossRef]
- Ziani, K.; Ursúa, B.; Maté, J.I. Application of bioactive coatings based on chitosan for artichoke seed protection. Crop Prot. 2010, 29, 853–859. [Google Scholar] [CrossRef]
- Rundlöf, M.; Andersson, G.K.S.; Bommarco, R.; Fries, I.; Hederström, V.; Herbertsson, L.; Jonsson, O.; Klatt, B.K.; Pedersen, T.R.; Yourstone, J.; et al. Seed coating with a neonicotinoid insecticide negatively affects wild bees. Nature 2015, 521, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.G.; Atwood, L.W.; Morris, M.B.; Mortensen, D.A.; Koide, R.T. Evidence for indirect effects of pesticide seed treatments on weed seed banks in maize and soybean. Agric. Ecosyst. Environ. 2016, 216, 269–273. [Google Scholar] [CrossRef]
- Rocha, I.; Ma, Y.; Souza-Alonso, P.; Vosátka, M.; Freitas, H.; Oliveira, R.S. Seed Coating: A Tool for Delivering Beneficial Microbes to Agricultural Crops. Front. Plant Sci. 2019, 10, 1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shweta, B.; Maheshwari, D.K.; Dubey, R.C.; Arora, D.S.; Bajpai, V.K.; Kang, S.C. Beneficial effects of fluorescent pseudomonads on seed germination, growth promotion, and suppression of charcoal rot in groundnut (Arachis hypogea L.). J. Microbiol. Biotechnol. 2008, 18, 1578–1583. [Google Scholar]
- Kaewkham, T.; Hynes, R.K.; Siri, B. The effect of accelerated seed ageing on cucumber germination following seed treatment with fungicides and microbial biocontrol agents for managing gummy stem blight by Didymella bryoniae. Biocontrol Sci. Technol. 2016, 26, 1048–1061. [Google Scholar] [CrossRef]
- Kasem, W.; Ghareeb, A.; Marwa, E. Seed morphology and seed coat sculpturing of 32 taxa of family Brassicaceae. J. Am. Sci. 2011, 7, 166–178. [Google Scholar]
- Zohora, U.S.; Ano, T.; Rahman, M.S. Biocontrol of Rhizoctonia solani K1 by iturin A producer Bacillus subtilis RB14 seed treatment in tomato plants. Adv. Microbiol. 2016, 6, 424–431. [Google Scholar] [CrossRef] [Green Version]
- Kipngeno, P.; Losenge, T.; Maina, N.; Kahangi, E.; Juma, P. Efficacy of Bacillus subtilis and Trichoderma asperellum against Pythium aphanidermatum in tomatoes. Biol. Control 2015, 90, 92–95. [Google Scholar] [CrossRef]
- Bennett, A.J.; Mead, A.; Whipps, J.M. Performance of carrot and onion seed primed with beneficial microorganisms in glasshouse and field trials. Biol. Control 2009, 51, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y. Seed coating with beneficial microorganisms for precision agriculture. Biotechnol. Adv. 2019, 37, 107423. [Google Scholar] [CrossRef]
- Calderón, C.E.; Rotem, N.; Harris, R.; Vela-Corcía, D.; Levy, M. Pseudozyma aphidis activates reactive oxygen species production, programmed cell death and morphological alterations in the necrotrophic fungus Botrytis cinerea. Mol. Plant Pathol. 2018, 20, 562–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Promoting effects of Pseudozyma aphidis on cucumber seedling growth. Plant height (a), absolute number of flowers (b), number of blooming flowers (c), and number of leaves (d) per plant were measured two weeks after application of P. aphidis L12 spore suspensions (PA, 108 spores/mL) or water (Control). The different letters in each panel denote statistically significant differences (n = 10 per treatment, p < 0.05, Tukey–Kramer HSD test).
Figure 1.
Promoting effects of Pseudozyma aphidis on cucumber seedling growth. Plant height (a), absolute number of flowers (b), number of blooming flowers (c), and number of leaves (d) per plant were measured two weeks after application of P. aphidis L12 spore suspensions (PA, 108 spores/mL) or water (Control). The different letters in each panel denote statistically significant differences (n = 10 per treatment, p < 0.05, Tukey–Kramer HSD test).

Figure 2.
Effects of Pseudozyma aphidis on in vitro cucumber seedlings root development. Root length (a), hypocotyl height (b), and seedling fresh weight (c) were measured 6 days after cucumber seeds were dipped into P. aphidis L12 spore suspensions (PA, 108 spores/mL) or water (Control). The different letters in each panel denote statistically significant differences (n = 10 per treatment, p < 0.05, Tukey-Kramer HSD test). Representative pictures of roots development after different treatments are presented above.
Figure 2.
Effects of Pseudozyma aphidis on in vitro cucumber seedlings root development. Root length (a), hypocotyl height (b), and seedling fresh weight (c) were measured 6 days after cucumber seeds were dipped into P. aphidis L12 spore suspensions (PA, 108 spores/mL) or water (Control). The different letters in each panel denote statistically significant differences (n = 10 per treatment, p < 0.05, Tukey-Kramer HSD test). Representative pictures of roots development after different treatments are presented above.

Figure 3.
Effects of Pseudozyma aphidis on tomato seedling growth. ‘Sherlot’ tomato seeds were coated with P. aphidis L12 (PA, 108 spores/mL) or water (Control) and grown on sterile paper at 25 °C. (a) Average root length ± SE, (b) average hypocotyl length ± SE, and (c) average seedling fresh weight ± SE were measured one week after application. The different letters in each panel denote statistically significant differences (n = 24 per treatment, p < 0.05, Tukey–Kramer HSD test). Representative pictures of roots development after different treatments are presented above.
Figure 3.
Effects of Pseudozyma aphidis on tomato seedling growth. ‘Sherlot’ tomato seeds were coated with P. aphidis L12 (PA, 108 spores/mL) or water (Control) and grown on sterile paper at 25 °C. (a) Average root length ± SE, (b) average hypocotyl length ± SE, and (c) average seedling fresh weight ± SE were measured one week after application. The different letters in each panel denote statistically significant differences (n = 24 per treatment, p < 0.05, Tukey–Kramer HSD test). Representative pictures of roots development after different treatments are presented above.

Figure 4.
Effect of Pseudozyma aphidis on growth parameters of Micro-Tom tomato plants. (a) Average ± SE number of leaves (b) average ± SE plant height and number of flowers (c). Plants were sprayed with P. aphidis L12 (PA, 108 spores/mL) or water (Control). The different letters in each panel denote statistically significant differences according to a Tukey–Kramer HSD test (n = 12 per treatment; p < 0.05). Representative pictures of plants after different treatments are presented above.
Figure 4.
Effect of Pseudozyma aphidis on growth parameters of Micro-Tom tomato plants. (a) Average ± SE number of leaves (b) average ± SE plant height and number of flowers (c). Plants were sprayed with P. aphidis L12 (PA, 108 spores/mL) or water (Control). The different letters in each panel denote statistically significant differences according to a Tukey–Kramer HSD test (n = 12 per treatment; p < 0.05). Representative pictures of plants after different treatments are presented above.

Figure 5.
Effect of Pseudozyma aphidis on Micro-Tom tomato plant yield parameters. (a) Average fruit weight ± SE, (b) number of fruits collected ± SE per plant 10 weeks after treatment with P. aphidis L12 (PA, 108 spores/mL) or water as a control, (c) total fruits weight per plant and (d) percentage of ripened fruit per plant ± SE. The different letters in each panel denote statistically significant differences according to Tukey–Kramer HSD test (n = 10—12 per treatment, p < 0.05). Representative pictures of the collected fruits from plants after different treatments are presented above.
Figure 5.
Effect of Pseudozyma aphidis on Micro-Tom tomato plant yield parameters. (a) Average fruit weight ± SE, (b) number of fruits collected ± SE per plant 10 weeks after treatment with P. aphidis L12 (PA, 108 spores/mL) or water as a control, (c) total fruits weight per plant and (d) percentage of ripened fruit per plant ± SE. The different letters in each panel denote statistically significant differences according to Tukey–Kramer HSD test (n = 10—12 per treatment, p < 0.05). Representative pictures of the collected fruits from plants after different treatments are presented above.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shoam, I.; Calderon, C.E.; Levy, M. Pseudozyma aphidis Enhances Cucumber and Tomato Plant Growth and Yield. Agronomy 2022, 12, 383. https://doi.org/10.3390/agronomy12020383
AMA Style
Shoam I, Calderon CE, Levy M. Pseudozyma aphidis Enhances Cucumber and Tomato Plant Growth and Yield. Agronomy. 2022; 12(2):383. https://doi.org/10.3390/agronomy12020383
Chicago/Turabian StyleShoam, Idan, Claudia E. Calderon, and Maggie Levy. 2022. "Pseudozyma aphidis Enhances Cucumber and Tomato Plant Growth and Yield" Agronomy 12, no. 2: 383. https://doi.org/10.3390/agronomy12020383
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.