

Antifungal Activity of Propolis Extracts against Postharvest Pathogen Phlyctema vagabunda
by
 and
and
Iván Urrea
1,
Nolberto Arismendi
2,
Ximena Sepúlveda
1,
Macarena Gerding
1,
Silvana Vero
3 and
Marisol Vargas
1,* 1
Laboratory of Phytopathology, Faculty of Agronomy, Universidad de Concepción, Av. Vicente Méndez 595, Chillán 3780000, Chile
2
Austral Biotech Research Center, Faculty of Science, Universidad Santo Tomás, Av. Ramón Picarte 1130–1160, Valdivia 5090000, Chile
3
Department of Microbiology, Faculty of Chemistry, Universidad de la República, Av. General Flores 2124, Montevideo 11800, Uruguay
*
Author to whom correspondence should be addressed.
Agronomy 2023, 13(1), 104; https://doi.org/10.3390/agronomy13010104
Submission received: 19 October 2022
/
Revised: 29 November 2022
/
Accepted: 30 November 2022
/
Published: 29 December 2022
(This article belongs to the Special Issue Post-harvest Pest and Disease Management)
Abstract
:Phlyctema vagabunda is one of the main postharvest pathogens in late-harvest apples in Chile. The control of this pathogen is mainly through synthetic fungicides or copper-based compounds; however, there are concerns about the residues of these products in food and the environment. Therefore, there is a need for seeking alternative control strategies, in which propolis, a resinous substance collected by bees with antimicrobial properties, could be an environmentally friendly and safe alternative to control for P. vagabunda. The aim of this study was to characterize and determine the pathogenicity of Chilean isolates of P. vagabunda and to evaluate the antifungal activity of propolis against P. vagabunda in vitro and in vivo conditions. The Chilean isolates were identified as P. vagabunda through morphological and molecular characterization. The concentrations of propolis of 0.5% and 0.05% significantly decreased the mycelial growth and germination of conidia by 88.5% and 100%, respectively. The incidence and severity of the disease in apples also decreased by 57.2% and 61.3%, respectively. The use of propolis extract could be a promising alternative for the control of P. vagabunda in apples.
1. Introduction
Bull’s-eye rot of apple fruits has become an important and frequent postharvest disease in apples in Europe, South Africa, and Chile [1,2,3,4]. The disease can be caused by various species, including Phlyctema vagabunda, Neofabraea malicortis, Neofabraea perennans, and Neofabraea kienholzii. In Chile, P. vagabunda was reported for the first time in 2005 [5], and it was described as the only species of the genus Neofabraea [4] until the detection of P. perennans in 2020 [6].
Postharvest diseases represent an important limitation to preserving high-quality commercial fruit for a prolonged storage period [7]. Fruit infection occurs in the field and is favored by long periods of rainfall and temperatures near 20 °C [3], but symptoms show up after 3 to 5 months of cold storage (0 °C), with individual or numerous lesions observed in a single fruit [8]. Disease symptoms are characterized by a circular, slightly sunken lesion with concentric rings that can vary in color from light brown to dark brown, originating from a lenticel in the fruit epidermis [9]. According to the literature, the incidence of bull’s-eye rot may vary from 10 to 60% in late-harvest cultivars such as ‘Cripps Pink’, where they are the most affected varieties, with disease incidences depending on the season and locality [4]. In organic production, incidences in these cultivars can be as high as 80–90%. Furthermore, economic losses can be further increased if the pathogen presents a quarantine condition for export markets [10], as is the case in China [11].
The control strategies for the bull’s-eye rot disease include the removal of fallen fruit and dead tree branches from the orchard floor [12], and the use of fludioxonil and numerous copper applications, but even with the use of fludioxonil, the disease incidence is still around 35% [13]. However, due to growing concerns about chemical residues in the food chain and environmental problems, consumers are calling for a reduction in the use of synthetic fungicides [14]. Furthermore, chemical applications could induce the development of pathogen resistance to the active ingredients [15]. Therefore, there is a need to seek alternative control strategies for bull’s-eye rot disease that are both environmentally friendly and safe for human consumption. Studies on the use of propolis have shown promising results for postharvest disease control [16,17,18]. However, propolis extracts have not been used to control P. vagabunda.
Propolis is a natural resinous product collected by bees from different plant exudates [19]. The bees use propolis as a cementing material to close cracks or spaces that may exist in the hive, as well as having antiseptic properties that maintain hygiene inside the colony [20,21]. The use of propolis has a positive effect on human health, where its antibacterial, antifungal, anti-inflammatory, antiviral, anesthetic, and antioxidant properties stand out [22,23]. Propolis’ positive properties may be associated with its chemical components. In fact, over 850 chemical compounds have been identified in propolis, belonging to flavonoids, terpenes, and phenolics, which vary according to geographical location, plant sources, and bee species [24].
Therefore, the aim of this study was to characterize and determine the pathogenicity of Chilean isolates of P. vagabunda and to evaluate the antifungal activity of propolis methanolic extracts against P. vagabunda in vitro and in vivo.
2. Materials and Methods
2.1. Pathogen Isolation
An apple of the cultivars ‘Cripps Pink’, ‘Fuji’, and ‘Royal Gala’ was collected in autumn 2017 from organic farms in the Cato area, Ñuble Region (36°30′ S–71°48′ W). The samples consisted of a box containing 80 apples (18 kg). Six samples of ‘Cripps Pink’, five of ‘Fuji’, and five of ‘Royal Gala’ were stored at 0 °C for 5 months. Apples with characteristic symptoms of the disease were selected and superficially disinfected with sodium hypochlorite (1.0%). Then symptomatic pieces were cut from the margins of the lesion, which were deposited in Petri dishes with potato dextrose agar medium (PDA) and incubated for 14 days at 20 °C. Pure cultures of P. vagabunda isolates were preserved at −80 °C in 80% (v/v) glycerol until used.
2.2. DNA Extraction from Phlyctema Vagabunda Isolates
The DNA extraction was performed in six isolates of P. vagabunda from cultivar ‘Cripps Pink’, based on the methodology described by Montalva et al. [25], with modifications. Thus, pieces of mycelium were macerated into 1.5 mL microcentrifuge tubes with 200 μL of preheated (65 °C) 4% CTAB extraction buffer (Tris–HCl 100 mM pH 8.0, NaCl 1.4 M, EDTA 20 mM pH 8.0, 4% CTAB w/v, and 0.2% 2-mercaptoethanol (v/v)) using sterile plastic pestles. Another 300 μL of extraction buffer was added, and the macerated samples were then incubated for 30 min at 65 °C. Subsequently, 500 μL of chloroform-octanol (24:1) cooled to −20 °C were added, mixed by inversion, and centrifuged at 4 °C for 10 min at 5000 g. The supernatant phase was extracted, mixed again with 400 μL of chloroform-octanol (24:1), and centrifuged (5 min at 5000× g). After that, 300 μL of the supernatant was placed into a new 1.5 mL tube, and 150 μL ammonium acetate (7.5 M) and 220 μL of cold isopropanol (−20 °C) were added, allowing the precipitation of the nucleic acids at −20 °C for 60 min. After this period, the samples were centrifuged at 13,000 g for 20 min at 4 °C. The precipitated DNA was then washed with 200 μL of ethanol (70%) and centrifuged for 5 min at room temperature. Then, the ethanol was removed, and the DNA pellet was allowed to dry for 40 min at 37 °C. Finally, the DNA pellet was suspended in 50 μL of molecular grade water, quantified using Infinite 200 PRO NanoQuant (Tecan Group Ltd., Männedorf, Switzerland), and stored at −20 °C for later use.
2.3. Fungal DNA Amplification
The PCR reaction (25 μL) contained 20 ng of DNA, 0.1 mM dNTPs, 2 mM MgCl2, 0.4 µM primers, and one unit of Taq DNA polymerase in 1× Taq PCR buffer. PCR was carried out using the primer described by Cao et al. [26] (F 5′-CTTTCTCCGTTGTCCCATCC-3 and R 5′-GAACATTGCGCATCTGGTCC-3′) that amplify a region of the β-tubulin gene (βtub). The amplification conditions were as follows: an initial denaturation at 94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 58 °C for 30 s, polymerization at 72 °C for 45 s, and an extension step at 72 °C for 5 min. PCR products were separated on a 1.0% (w/v) agarose gel with 0.05 μL mL−1 of SYBR™ Safe DNA Gel Stain (Invitrogen, Carlsbad, CA, USA) in a TBE buffer at 70 V for 60 min. The amplicons were visualized in a UV light transilluminator (TFX-20.M, Vilber Lourmat, Marne-la-Vallée, France).
2.4. Nucleotide Sequencing and Phylogenetic Analyses
To verify the identity of the fungal isolates, PCR amplicons were purified and sequenced by Macrogen Inc. (Seoul, Republic of Korea). The sequences obtained were compared with other β-tubulin genes deposited in the GenBank® database using the Basic Local Alignment Search Tool (BLAST, NCBI) to determine sequence similarities. Additionally, a phylogenetic tree for P. vagabunda isolates reported in Chile and in other parts of the world was constructed with Geneious Prime 2019 software (Biomatters, Auckland, New Zealand) using the neighbor-joining method with 1000 bootstraps. The sequences were submitted to the NCBI GenBank database.
2.5. Effect of Temperature on Mycelial Growth and Conidial Production of Phlyctema Vagabunda
Four Chilean isolates of P. vagabunda (CP5, CP8, CP9, and CP13) from ‘Cripps Pink’ apples were used to determine the effect of temperature on mycelial growth and on the ability to produce conidia, according to the methodology described by Cameldi et al. [1]. In brief, in Petri dishes with tomato agar medium (TA), 6 mm agar discs with the 10-day pathogen mycelium in active growth were placed in the center of the plate and incubated at 0, 5, 10, 15, 20, 25, and 30 °C for 14 days. After that, mycelial growth and morphology were assayed at each incubation temperature.
The presence of conidia was determined by microscopic observation at 10× magnifications (BA310, Motic, Hong Kong, China), and the number of conidia per colony was determined by suspending total conidia in distilled water and quantifying them in a hemocytometer. The longitudinal and transverse dimensions of 50 conidia of each isolate were determined by using the Motic Image Plus 2.0 software.
2.6. Pathogenicity of Phlyctema Vagabunda Isolates In Vivo Condition
The pathogenicity of the isolates CP5, CP8, CP9, and CP13 was validated and determined in ‘Cripps Pink’ apples. Fruits were disinfected superficially by immersion in sodium hypochlorite (1.0% v/v) for 3 min, rinsed four times in distilled water, and allowed to dry in the laminar flow chamber. Then, the equatorial area of the fruit was disinfected with ethanol (70%), and a fruit disk (5 mm) was extracted from this section and replaced by a 10-day mycelial disk of the pathogen. The apples were incubated for 20 days at 20 °C. The disease severity was determined by measuring of rot diameter [27].
2.7. Propolis Extract Source
To evaluate the antifungal activity of propolis against P. vagabunda, four methanolic extracts of propolis obtained previously by Arismendi et al. [28] were used in this study. Two samples, LR15 and LR16, were obtained from apiaries located in the city of Valdivia, Los Ríos Region (39°48′ S–73°17′ W) in the years 2015 and 2016, respectively. The other two samples, RÑ15 and RÑ16, were collected in the city of Chillán, Ñuble Region (36°32′ S–71°59′ W), also in the years 2015 and 2016, respectively. Briefly, propolis samples were kept at 4 °C and protected from light until polyphenols were extracted. The propolis was ground into a powder using a mortar and pestle. Then, 500 mL of methanol (100%) was mixed with 60 g of propolis. The mixtures were kept for 48 h in sealed vessels at room temperature (20 °C) in dark conditions. After this time, the mixture was vacuum filtered to obtain a colored liquid with no waste particles. To separate the methanol solvent from propolis compounds, it was completely evaporated by using a rotary evaporator (R-210, Buchi, Flawil, Switzerland). The extract was stored at 4 °C until its use.
2.8. Effect of Methanolic Propolis Extracts on the Mycelial Growth of Phlyctema Vagabunda Isolates
The effect of the four propolis extracts (RÑ15, RÑ16, LR15, and LR16) on the mycelial growth of P. vagabunda was determined using the methodology described by Yang et al. [18], with modifications. We used isolate CP13 of P. vagabunda since it resulted in the most aggressive isolate. Thus, different concentrations of propolis extract were prepared by dissolving the requisite amounts in 50% ethanol and mixing with organic apple juice agar (AFE® + 20 g agar L−1) to obtain final concentrations of 0.0, 0.01, 0.05, 0.1, 0.15, 0.2, 0.25, 0.30, 0.40, and 0.50%. Agar discs (5 mm) with the 10-day pathogen mycelium in active growth were placed on each plate and incubated at 20 °C for 14 days. The control corresponded to ethanol (50%). Each treatment was performed in triplicate. The diameter of the mycelial growth of P. vagabunda was measured, and the percentage of inhibition of mycelial growth was determined using the formula described by Mohammedi and Atik [29]:
where
Growth inhibition (%) = [(Dc − Dt)/Dc] × 100
Dc = diameter of the colony in the control (mm);
Dt = diameter of the colony in the treatment (mm).
2.9. Effect of Methanolic Propolis Extract on the Conidial Germination in In Vitro Condition
Inhibition of conidia germination of the P. vagabunda isolate was determined using the methodology described above at the following propolis concentrations: 0.00, 0.005, 0.01, 0.05, and 0.1%. In these cases, the propolis (LR16) that showed the greater inhibition of mycelial growth in previous experiments was tested against isolate CP13. Thus, 150 μL of a conidial suspension of the pathogen (1 × 104 conidia mL−1) was spread in Petri dishes with apple juice agar containing different propolis concentrations and incubated at 20 °C for 4 days. The control corresponded to ethanol (50%). Three replicates (plates) were performed per concentration, and a control treatment without propolis was included. The percentage of inhibition of conidial germination was determined with the formula:
where
Inhibition of conidia germination (%) = [(Gc − Gt)/Gc] × 100.
Gc = germination of conidia in the control;
Gt = germination of conidia in the treatment with propolis.
2.10. Effect of Propolis Extract on the Control of Phlyctema Vagabunda in Apple
The antifungal activity of the propolis extract (LR16) in fruits was determined using the methodology described by Daniel et al. [30], with modifications. Organic ‘Cripps Pink’ apples were used, selecting healthy and homogeneous fruits. Fruits were disinfected superficially with 1.0% sodium hypochlorite (v/v), rinsed in distilled water, and allowed to dry in the flow chamber. In the equatorial zone of the fruit, two equidistant wounds (3 × 3 mm) were made, where one corresponded to the treatment with 30 μL of propolis extract (0.5%) dissolved in ethanol (50%) and the other to the control, treated only with ethanol (50%). The ethanol was allowed to evaporate for 60 min, and then, 20 μL of a conidial suspension (5 × 105 conidia mL−1) of P. vagabunda (CP13 isolate) was inoculated into each wound. The fruits (five fruits) were arranged in plastic boxes (one box per replicate, four replicates), and plates with water were used to maintain high humidity (~80% RH). The boxes were incubated at 20 °C for 20 days. The percentage of incidence and severity of the disease were determined according to Vero et al. [27]: The severity was considered as the diameter of rot caused by the disease. The assay was repeated twice.
Incidence (%) = (number of symptomatic wounds/total number of wounds) × 100.
2.11. Data Analysis
The statistical difference in the disease severity of Chilean isolates of P. vagabunda in ‘Cripps Pink’ apples and the specific effect of propolis extract on conidial germination of P. vagabunda were determined by a one-way ANOVA. Then, post hoc analysis was run with the Tukey HSD test to separate means between treatments. Furthermore, the effect of propolis extracts on mycelial growth was presented as a cluster analysis (standardized data) with the Euclidean metric as a measure of distance. Clusters were subjected to a nonparametric analysis of variance and a comparison of means through the Kruskal–Wallis test. Additionally, the incidence and severity of P. vagabunda in apples and in those treated with propolis were determined by the Student’s t-test. All analyses were run using InfoStat software 2018e (FCA-UNC, Córdoba, Argentina).
3. Results
3.1. Isolation, Characterization and Molecular Identification of the Chilean Phlyctema Vagabunda Isolates
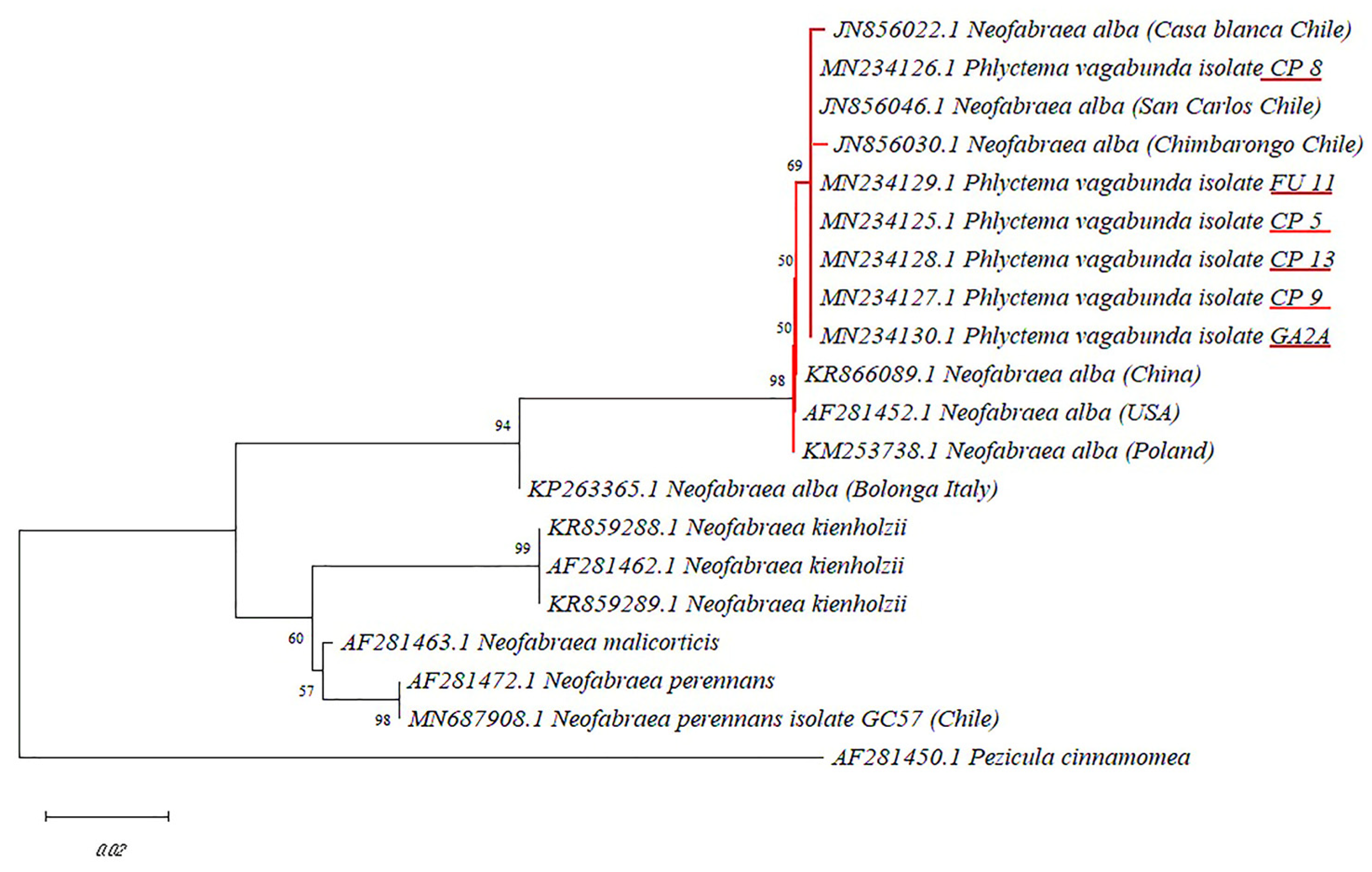
The stored apples ’Cripps Pink’, ‘Fuji’, and ‘Royal Gala’ recorded an incidence of 92%, 80%, and 18%, respectively. In total, 60 isolates of P. vagabunda were obtained from apple fruits that showed bull’s-eye rot symptoms. Moreover, 30, 28, and 2 P. vagabunda isolates were isolated from apples ‘Cripps Pink’, ‘Fuji’, and ‘Royal Gala’, respectively. The isolate colonies showed compact mycelial growth, septate hyphae, and mycelia that were initially white and then turned to a white/pink color, although, some isolates showed colonies of a darker color. Of the thirty ‘Cripps Pink’ isolates, four (CP5, CP8, CP9, and CP13) were selected for identification through sequencing since each of them showed different mycelia colors. Additionally, an isolate (FU11) from ‘Fuji’ and one (GA2A) from ‘Royal Gala’ were also randomly selected for sequencing. A single DNA fragment of 554 bp of the β-tubulin gene was amplified in all the isolates amplified. The DNA sequences showed that all the Chilean isolates of P. vagabunda had 100% homology between them and 98% similarity with P. vagabunda reference sequences obtained from GenBank (Accession KM253738, AF281452, and KR866089). Furthermore, the phylogenetic tree constructed by the neighbor-joining method showed that Chilean P. vagabunda isolates (CP5, FU11, CP13, CP9, CP8, and GA2A) were grouped in the same clade with the reference isolates of P. vagabunda with a significant bootstrap value (98) (Figure 1). Within the P. vagabunda clade, the Chilean isolates formed a subgroup on their own, separated from the P. vagabunda isolate from Bologna, Italy (KP263365) (Figure 1). The sequences of the isolates CP5, FU11, CP13, CP9, CP8, and GA2A were deposited in the GenBank® database under accession numbers MN234125, MN234129, MN234126, MN234128, MN234127, and MN234130, respectively.
3.2. Effect of Temperature on Mycelial Growth and Conidial Production of Phlyctema Vagabunda
Temperatures of 15 and 20 °C induced the highest mycelial growth for the four Chilean P. vagabunda isolates (CP5, CP8, CP9, and CP13) tested in this assay (Table 1 and Figure 2), where the isolate CP13 outstood with the highest mycelial growth at 20 °C. The mycelial growth of the isolates was significantly lower (nested ANOVA, F = 6.11, p ≤ 0.0001, df = 21.56) at temperatures higher than 20 °C and lower than 15 °C, and at 30 °C no mycelial growth was detected. On the other hand, when Chilean isolates were tested at the same temperature that apple fruits are normally stored (0 °C) and at 5 °C, the fungal mycelial growth was lower, but it never stopped growing (Table 1). The four Chilean isolates showed conidial production only at 0 and 5 °C; no conidia were observed at the other tested temperatures (Table 1).
The conidia produced at 0 and 5 °C by the four isolates (CP5, CP8, CP9, and CP13) were cylindrical, with some being slightly curved. There were no differences in conidia size within the isolates (nested ANOVA F = 0.94, p = 0.49, df = 6.0, 16 media ± 0.5) at both temperatures. Conidia size varied from 9.99 µm × 2.79 µm to 11.11 µm × 3.05 µm (Table 2). There were statistical differences (one-way > ANOVA F = 7.36, p = 0.0005, df = 7.0, 16) in the conidia production per colony, where the isolates CP13 and CP8 reached higher conidia production values (5 °C) than the other Chilean isolates of P. vagabunda (Table 2).
3.3. Pathogenicity of Chilean Phlyctema Vagabunda Isolates on Apple Fruit
All of the Chilean isolates of P. vagabunda showed a 100% disease incidence in apple fruits. However, there were statistical differences (one-way ANOVA F = 3.61, p = 0.016, df = 3, 92) in the disease severity, wherein the Chilean CP13 isolate showed a significantly larger rot diameter than the CP8 isolate, with no significant differences from the CP9 and CP5 isolates (Figure 3 and Figure 4).
3.4. Effect of Propolis Extracts on the Mycelial Growth of Phlyctema Vagabunda
Since CP13 resulted in the most aggressive isolate in apple fruits (Figure 5), propolis methanolic extracts were tested for their effects on the mycelial growth of this isolate. The propolis extracts at different concentrations could be grouped into four clusters (CC = 0.91): Cluster 1, Cluster 2, Cluster 3, and Cluster 4, based on their effect on mycelial growth (Figure 5). In Cluster 1, the propolis extracts LR16: 0.5% and LR15: 0.5% were grouped in the same clade, which showed the greatest inhibition of mycelial growth, with 88.5 and 81.6%, respectively. In the other clusters, the inhibition of mycelial growth was equal to or less than 70%. The groups that showed greater inhibition of mycelial growth (Cluster 1 and Cluster 2) are mainly composed of extracts from the Los Ríos (Figure 5).
3.5. Effect of Propolis Extract on Conidial Germination of Phlyctema Vagabunda in In Vitro Condition
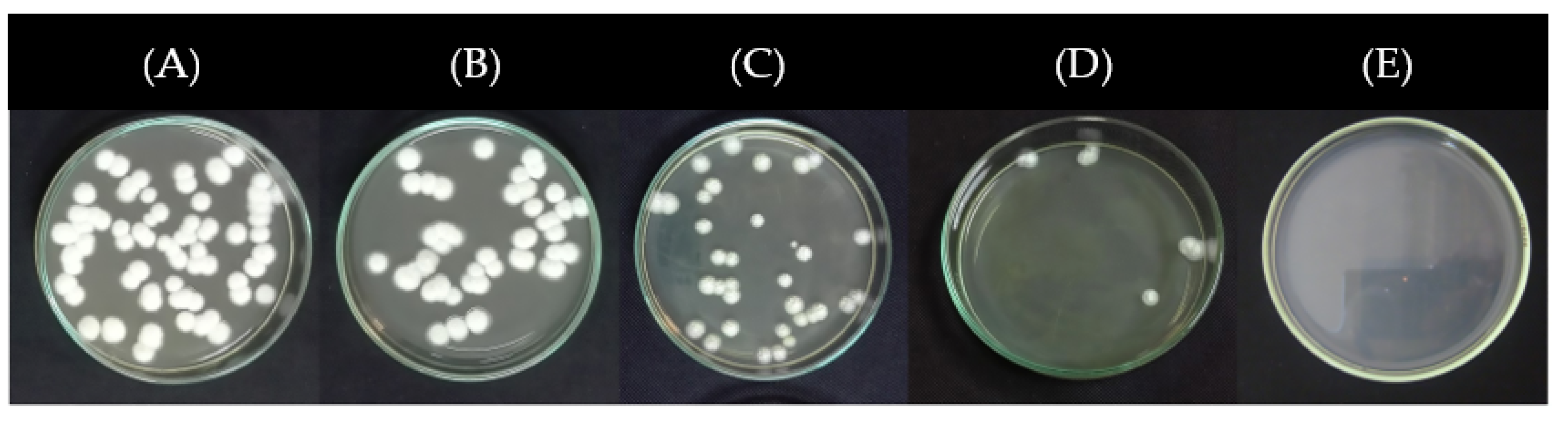
Considering the previous results, the effect of propolis extract LR16 at 0.005, 0.01, 0.05, and 0.1% on the inhibition of conidial germination of CP13 isolate was evaluated. Significant differences (one-way ANOVA F = 110.4, p < 0.001, df = 3, 8) were detected between propolis extract concentrations. Conidia germination inhibition was lower than 5% when exposed to the lowest propolis extract concentrations (0.005 and 0.01%). On the other hand, when the propolis extract concentration increased (0.05 and 0.1%), there was a 100% inhibition of CP13 isolate conidia germination (Figure 6 and Figure 7).
3.6. Effect of Propolis Extract LR16 on the Control of Phlyctema Vagabunda CP13 Isolate in In Vivo Condition
After 20 days of incubation at 20 °C, significant differences (t-test, p < 0.001) were detected in the rot incidence of P. vagabunda (isolate CP13) in apples that were treated with propolis extract (0.5%) and the untreated. In treated apples, the rot incidence decreased by almost 60%, whereas in untreated fruits, the incidence reached 100% (Figure 8A). In addition, the disease severity, expressed as the rot diameter, was significantly different (t-test, p < 0.001) between treated and untreated (control) apples (Figure 8B).
4. Discussion
The Chilean P. vagabunda isolated in this study was characterized by producing colonies with irregular borders and white-rosaceous mycelium when grown on a PDA medium at 20 °C. Similar results were obtained by Neri et al. [31], who indicated that the Chilean P. vagabunda produced colonies with pale white-pink mycelium when grown on a PDA medium at 15 °C. The symptomatology observed in the apple fruits resembles that described in the literature for P. vagabunda, that is, flat to slightly sunken circular brown lesions with concentric rings, with the affected tissues being relatively firm and easily detached from healthy tissue [4,5].
The highest mycelial growth rate of Chilean isolates of P. vagabunda was obtained at 20 °C, which is similar to other studies [1,32] and different from that obtained by Amaral Carneiro et al., who indicated that the highest colony growth rates were obtained at 15 and 20 °C [33]. The four Chilean isolates analyzed (CP5, CP8, CP9, and CP13) showed conidial production only at 0 and 5 °C (Table 1), and conidia were characterized as morphotype I in vitro condition (Table 2). Similar results were obtained by Neri et al. [31] for the Chilean isolates of P vagabunda. In contrast, Cameldi et al. [1] observed conidia sporulation in temperatures ranging from 0 to 25 °C after 14 days of incubation in vitro conditions and they found that morphotype I (10.7 µm × 2.8 µm) and morphotype II (23.1 µm × 3.3 µm) can be present in TA medium when some isolates were cultured at 15 °C. These differences in conidia morphotypes in vitro of the Chilean isolates in comparison to those from Italy could be due to genetic differences which are evidenced in the phylogenetic tree (Figure 1), where Chilean isolates are genetically separated from the Italian isolate. The molecular characterization of the isolates by partial sequencing of the β-tubulin gene showed that the four analyzed isolates corresponded to P. vagabunda, which agrees with that reported by Soto-Alvear et al. [4]. Within the P. vagabunda clade, the Chilean isolates are grouped on their own and apart from the other isolates, and they are particularly distant from the Italian isolate KP263365. According to De Jong et al. [34], the coding sequences of the β-tubulin gene provide robust phylogenetic information for the genus Neofabraea in comparison to other genomic regions such as ITS or mitochondrial rDNA, clearly demonstrating the genetic differences between closely related species.
The mycelial growth of the fungus was inhibited by 88.5% with propolis from the Los Ríos region (LR16) at a concentration of 0.5%; however, with propolis from the Ñuble region, the inhibition was significantly lower. Similar results were obtained by Ali et al. [35], using a propolis concentration three times higher (1.5%), who managed to reduce the mycelial growth of Colletotrichum gloeosporioides by 87%. Propolis is a source of natural antibiotics, and its extracts have broad-spectrum inhibitory effects on bacteria, fungi, and nematodes [36,37,38].
In this study, the highest antifungal activity was observed in propolis from the Los Ríos region (LR15 and LR16), compared to those from the Ñuble region (RÑ 15 and RÑ 16). This difference in antifungal activity could be derived from the chemical characteristics of propolis, which may be linked to their geographical location and botanical origin [39]. Arismendi et al. [28] determined that the propolis of the Los Ríos region contained high levels of pinocembrin and galangin, compared to those of the Ñuble region (RÑ 15 and RÑ 16). This quantitative difference in antimicrobial compounds present in propolis could be dependent on the collection region, mainly because of the flora from which it is extracted by the bees [36,40,41].
It has been shown that flavonoids such as pinocembrin (flavanone) and galangin (flavonol), have antifungal activity against phytopathogens such as Aspergillus niger, Penicillium notatum, and Fusarium sp. [42,43]. In studies made with propolis from China, strong antifungal activity was demonstrated against citrus blue mold and green mold caused by Penicillium italicum and P. digitatum, respectively, with pinocembrin being identified as one of the main active components [36,44]. The mode of action of pinocembrin is the inhibition of respiration in hyphal cells, which leads to a deficit of energy and damage to cell membranes, accelerating cell death [45].
The use of low concentrations (0.05%) of propolis completely inhibited the germination of P. vagabunda conidia; however, to inhibit mycelial growth, the propolis concentrations had to be increased 10-fold to reduce 88.5% of the mycelial growth of the pathogen. Similar results were obtained with the use of propolis to evaluate the germination of conidia of Botrytis cinerea and Rhizopus stolonifer in strawberries; at a concentration of 0.04%, the germination of conidia was 100% inhibited [18]. The use of propolis at a concentration of 0.5% reduced the incidence of P. vagabunda by 57.1% and the severity by 63.1% in ‘Cripps Pink’ apples. Yang et al. [18] reduced the expression of B. cinerea and R. stolonifer in strawberries by 82% through the use of propolis, yet the control presented only 25% of the natural incidence. Pereira et al. [46] showed that propolis at a concentration of 5% reduced the lesion size caused by P. expansum in apples by 66.5%. Loebler et al. [47] reported that propolis decreased the disease incidence of Stemphylium vesicarium in pears decreased by 25%. In another study where 1.5% chitosan was used in a mixture with propolis (1.5%) for the control of C. gloeosporioides in avocado, no differences were observed when applying the chitosan alone or in a mixture with propolis, but it increased the firmness of the fruits [48]. Propolis, in addition to having antimicrobial activity, could improve some organoleptic characteristics in fruits, parameters not evaluated in this study.
Temperature is an important environmental factor in the growth of fungi both in the field and in cold room conditions [49]. In this sense, it is important to keep in mind that low temperatures (0–5 °C) do not prevent the development of the fungus, which is relevant considering that the fruit is stored in cold rooms (0 °C) for at least three months, where the development of the pathogen continues [1,5]. Therefore, any tool that is developed to control this pathogen must consider temperature as an essential factor in the effectiveness of the control of these potential tools, in the sense that temperature does not limit the potential of control of these technologies. Our study shows that Chilean propolis can strongly affect mycelial growth and conidia germination of P. vagabunda; however, these results are based on temperate climate conditions (20 °C) and not at temperatures as low as 0 °C. Therefore, new assays should be performed under cold storage conditions to validate what was observed in this study.
5. Conclusions
The molecular characterization showed that the six analyzed isolates belonged to the species P. vagabunda. This study also showed that the Chilean propolis extracts had significant in vitro and in vivo antifungal activity against P. vagabunda. Thus, the use of propolis extract could be a promising alternative for the control of bull’s-eye rot disease in apples. However, more studies are required to demonstrate the effect of propolis extract in the control of P. vagabunda in real conditions, especially under cold fruit storage.
Author Contributions
M.V. conceived of this research; I.U., N.A., M.G. and M.V. designed experiments; I.U. and X.S. performed the experiments, the analysis in labs and statistical analysis; I.U., N.A., S.V., M.G. and M.V. wrote the paper and participated in the revisions of it. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Grant CONICYT-PCHA N° 22171680 from the National Research and Development Agency (ANID, Ex-CONICYT), Chile, and CYTED 121RT0110.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of this study, including the data collection, analyses, or interpretation of the data. The funders have an interest in or role in the writing of the manuscript or in the decision to publish the results.
References
- Cameldi, I.; Neri, F.; Menghini, M.; Pirondi, A.; Nanni, I.M.; Collina, M.; Mari, M. Characterization of Neofabraea vagabunda isolates causing apple bull’s eye rot in Italy (Emilia-Romagna region). Plant Pathol. 2017, 66, 1432–1444. [Google Scholar] [CrossRef]
- Den Breeyen, A.; Rochefort, J.; Russouw, A.; Meitz-Hopkins, J.; Lennox, C.L. Preharvest detection and postharvest incidence of Phlyctema vagabunda on ‘Cripps Pink’ apples in South Africa. Plant Dis. 2020, 104, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Giraud, M.; Coureau, C.; Perrin, J.; Westercamp, P. Apple lenticel rots: State of knowledge on the epidemiology of Neofabraea vagabunda. Acta Hortic. 2021, 1325, 59–66. [Google Scholar] [CrossRef]
- Soto-Alvear, S.; Lolas, M.; Rosales, I.M.; Chávez, E.R.; Latorre, B.A. Characterization of the bull’s eye rot of apple in Chile. Plant Dis. 2013, 97, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriquez, J.L. First report of apple rot caused by Neofabraea alba in Chile. Plant Dis. 2005, 89, 1360. [Google Scholar] [CrossRef] [PubMed]
- González, F.; Salinas, C.; Pinilla, B.; Castillo, A. First report of apple bull’s-eye rot caused by Neofabraea perennans in Chile. Plant Dis. 2020, 104, 1537. [Google Scholar] [CrossRef]
- Aguilar, C.G.; Mazzola, M.; Xiao, C.L. Timing of apple fruit infection by Neofabraea perennans and Neofabraea kienholzii in relation to bull’s-eye rot development in stored apple fruit. Plant Dis. 2017, 101, 800–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spotts, R.A.; Seifert, K.A.; Wallis, K.M.; Sugar, D.; Xiao, C.L.; Serdani, M.; Henriquez, J.L. Description of Cryptosporiopsis kienholzii and species profiles of Neofabraea in major pome fruit growing districts in the Pacific Northwest USA. Mycol. Res. 2009, 113, 1301–1311. [Google Scholar] [CrossRef]
- Vukotić, J.; Stojšin, V.; Nagl, N.; Petreš, M.; Hrustić, J.; Grahovac, M.; Tanović, B. Morphological, molecular, and pathogenic characterization of Neofabraea alba, a postharvest pathogen of apple in Serbia. Agronomy 2022, 12, 2015. [Google Scholar] [CrossRef]
- Lin, H.; Jiang, X.; Yi, J.; Wang, X.; Zuo, R.; Jiang, Z.; Wang, W.; Zhou, E. Molecular identification of Neofabraea species associated with bull’s-eye rot on apple using rolling-circle amplification of partial EF-1α sequence. Can. J. Microbiol. 2018, 64, 57–68. [Google Scholar] [CrossRef]
- Wood, P.N.; Fisher, B.M. The effect of fungicides on spore germination, mycelial growth and lesion development of Phlyctema vagabunda (syn: Neofabraea alba) (bull’s eye rot of apples). N. Z. Plant Prot. 2017, 70, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Wenneker, M.; Thomma, B.P.H.J. Latent postharvest pathogens of pome fruit and their management: From single measures to a systems intervention approach. Eur. J. Plant Pathol. 2020, 156, 663–681. [Google Scholar] [CrossRef] [Green Version]
- Lolas, M.; Díaz, G.; Mendez, R.; Cáceres, M.; Neubauer, L. Evaluation of the efficacy of fungicide fludioxonil in the postharvest control of bull’s eye rot (Neofabraea alba) in Chile. Acta Hortic. 2016, 1144, 461–464. [Google Scholar] [CrossRef]
- Mesías, F.J.; Martín, A.; Hernández, A. Consumers’ growing appetite for natural foods: Perceptions towards the use of natural preservatives in fresh fruit. Food Res. Int. 2021, 150, 110749. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fernández-Ortuño, D.; Grabke, A.; Schnabel, G. Resistance to fludioxonil in Botrytis cinerea isolates from blackberry and strawberry. Phytopathology 2014, 104, 724–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, M.A.; Vallejo, A.M.; Ballester, A.-R.; Zampini, C.; Isla, M.I.; López-Rubio, A.; Fabra, M.J. Antifungal edible coatings containing Argentinian propolis extract and their application in raspberries. Food Hydrocoll. 2020, 107, 105973. [Google Scholar] [CrossRef]
- Pobiega, K.; Kraśniewska, K.; Derewiaka, D.; Gniewosz, M. Comparison of the antimicrobial activity of propolis extracts obtained by means of various extraction methods. J. Food Sci. Technol. 2019, 56, 5386–5395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Zhou, Y.; Ye, J.; Fan, G.; Peng, L.; Pan, S. Effects of poplar buds as an alternative to propolis on postharvest diseases control of strawberry fruits: Poplar buds to control postharvest disease of strawberry. J. Sci. Food Agric. 2016, 96, 2136–2141. [Google Scholar] [CrossRef]
- Soylu, E.M.; Ozdemir, A.E.; Erturk, E.; Sahinler, N.; Soylu, S. Antifungal activity of propolis against postharvest disease agent Penicillium digitatum. Asian J. Chem. 2008, 20, 4823–4830. [Google Scholar]
- Akbay, E.; Özenirler, Ç.; Çelemli, Ö.G.; Durukan, A.B.; Onur, M.A.; Sorkun, K. Effects of propolis on warfarin efficacy. Pol. J. Cardio-Thorac. Surg. 2017, 1, 43–46. [Google Scholar] [CrossRef] [Green Version]
- Kasiotis, K.M.; Anastasiadou, P.; Papadopoulos, A.; Machera, K. Revisiting Greek propolis: Chromatographic analysis and antioxidant activity study. PLoS ONE 2017, 12, e0170077. [Google Scholar] [CrossRef] [PubMed]
- Mutlu Sariguzel, F.; Berk, E.; Koc, A.N.; Sav, H.; Demir, G. Antifungal activity of propolis against yeasts isolated from blood culture: In vitro evaluation: Antifungal activity of propolis. J. Clin. Lab. Anal. 2016, 30, 513–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omar, R.; Igoli, J.O.; Zhang, T.; Gray, A.I.; Ebiloma, G.U.; Clements, C.J.; Fearnley, J.; Edrada Ebel, R.; Paget, T.; de Koning, H.P.; et al. The chemical characterization of nigerian propolis samples and their activity against Trypanosoma brucei. Sci. Rep. 2017, 7, 923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šturm, L.; Ulrih, N.P. Advances in the propolis chemical composition between 2013 and 2018: A review. eFood 2020, 1, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Montalva, C.; Arismendi, N.; Barta, M.; Rojas, E. Molecular differentiation of recently described Neozygites osornensis (Neozygitales: Neozygitaceae) from two morphologically similar species. J. Invertebr. Pathol. 2014, 115, 92–94. [Google Scholar] [CrossRef]
- Cao, D.; Li, X.; Cao, J.; Wang, W. PCR detection of the three neofabraea pathogenic species responsible for apple bull’s eye rot. Adv. Microbiol. 2013, 3, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Vero, S.; Mondino, P.; Burgueño, J.; Soubes, M.; Wisniewski, M. Characterization of biocontrol activity of two yeast strains from Uruguay against blue mold of apple. Postharvest Biol. Technol. 2002, 26, 91–98. [Google Scholar] [CrossRef]
- Arismendi, N.; Vargas, M.; López, M.D.; Barría, Y.; Zapata, N. Promising antimicrobial activity against the honey bee parasite Nosema ceranae by methanolic extracts from Chilean native plants and propolis. J. Apic. Res. 2018, 57, 522–535. [Google Scholar] [CrossRef]
- Mohammedi, Z.; Atik, F. Fungitoxic effect of natural extracts on mycelial growth, spore germination and aflatoxin B1 production of Aspergillus flavus. Aust. J. Crop Sci. 2013, 7, 293–298. [Google Scholar]
- Daniel, C.K.; Lennox, C.L.; Vries, F.A. In vivo application of garlic extracts in combination with clove oil to prevent postharvest decay caused by Botrytis cinerea, Penicillium expansum and Neofabraea alba on apples. Postharvest Biol. Technol. 2015, 99, 88–92. [Google Scholar] [CrossRef]
- Neri, F.; Crucitti, D.; Negrini, F.; Pacifico, D.; Ceredi, G.; Carimi, F.; Lolas, M.A.; Collina, M.; Baraldi, E. New insight into morphological and genetic diversity of Phlyctema vagabunda and Neofabraea kienholzii causing bull’s eye rot on apple and pear. Plant Pathol. 2022, 1–22. [Google Scholar] [CrossRef]
- Hortova, B.; Novotny, D.; Erban, T. Physiological characteristics and pathogenicity of eight Neofabraea isolates from apples in Czechia. Eur. J. Hortic. Sci. 2014, 79, 327–334. [Google Scholar]
- Amaral Carneiro, G.; Walcher, M.; Storti, A.; Baric, S. Phylogenetic diversity and phenotypic characterization of Phlyctema vagabunda (syn. Neofabraea alba ) and Neofabraea kienholzii causing postharvest bull’s eye rot of apple in Northern Italy. Plant Dis. 2022, 106, 451–463. [Google Scholar] [CrossRef] [PubMed]
- de Jong, S.N.; Lévesque, C.A.; Verkley, G.J.M.; Abeln, E.C.A.; Rahe, J.E.; Braun, P.G. Phylogenetic relationships among Neofabraea species causing tree cankers and bull’s-eye rot of apple based on DNA sequencing of ITS nuclear rDNA, mitochondrial rDNA, and the β-tubulin gene. Mycol. Res. 2001, 105, 658–669. [Google Scholar] [CrossRef]
- Ali, A.; Cheong, C.; Noosheen, Z. Composite effect of propolis and gum arabic to control postharvest anthracnose and maintain quality of papaya during storage. Int. J. Agric. Biol. 2014, 1, 1117–1122. [Google Scholar]
- Yang, S.Z.; Peng, L.T.; Su, X.J.; Chen, F.; Cheng, Y.J.; Fan, G.; Pan, S.Y. Bioassay-guided isolation and identification of antifungal components from propolis against Penicillium italicum. Food Chem. 2011, 127, 210–215. [Google Scholar] [CrossRef]
- Petruzzi, L.; Rosaria Corbo, M.; Campaniello, D.; Speranza, B.; Sinigaglia, M.; Bevilacqua, A. Antifungal and antibacterial effect of propolis: A comparative hit for food-borne pseudomonas, enterobacteriaceae and fungi. Foods 2020, 9, 559. [Google Scholar] [CrossRef]
- Paula, L.A.L.; Cândido, A.C.B.B.; Santos, M.F.C.; Caffrey, C.R.; Bastos, J.K.; Ambrósio, S.R.; Magalhães, L.G. Antiparasitic properties of propolis extracts and their compounds. Chem. Biodivers. 2021, 18, e2100310. [Google Scholar] [CrossRef]
- Barrientos, L.; Herrera, C.L.; Montenegro, G.; Ortega, X.; Veloz, J.; Alvear, M.; Cuevas, A.; Saavedra, N.; Salazar, L.A. Chemical and botanical characterization of Chilean propolis and biological activity on cariogenic bacteria Streptococcus mutans and Streptococcus sobrinus. Braz. J. Microbiol. 2013, 44, 577–585. [Google Scholar] [CrossRef] [Green Version]
- Wagh, V.D. Propolis: A wonder bees product and its pharmacological potentials. Adv. Pharmacol. Sci. 2013, 2013, 308249. [Google Scholar] [CrossRef] [Green Version]
- Bueno-Silva, B.; Marsola, A.; Ikegaki, M.; Alencar, S.M.; Rosalen, P.L. The effect of seasons on Brazilian red propolis and its botanical source: Chemical composition and antibacterial activity. Nat. Prod. Res. 2017, 31, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, E.N.; Sampietro, D.A.; Soberon, J.R.; Sgariglia, M.A.; Vattuone, M.A. Propolis from the northwest of Argentina as a source of antifungal principles. J. Appl. Microbiol. 2006, 101, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Probst, I.; Sforcin, J.; Vlm, R.; Fernandes, A.; Fernandes Júnior, A. Antimicrobial activity of propolis and essential oils and synergism between these natural products. J. Venom. Anim. Toxins Trop. Dis. 2011, 17, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Peng, L.; Cheng, Y.; Chen, F.; Pan, S. Control of citrus green and blue molds by Chinese propolis. Food Sci. Biotechnol. 2010, 19, 1303–1308. [Google Scholar] [CrossRef]
- Peng, L.; Yang, S.; Cheng, Y.J.; Chen, F.; Pan, S.; Fan, G. Antifungal activity and action mode of pinocembrin from propolis against Penicillium italicum. Food Sci. Biotechnol. 2012, 21, 1533–1539. [Google Scholar] [CrossRef]
- Pereira, L.; Cunha, A.; Almeida-Aguiar, C. Portuguese propolis from Caramulo as a biocontrol agent of the apple blue mold. Food Control 2022, 139, 109071. [Google Scholar] [CrossRef]
- Loebler, M.; Sánchez, C.; Muchagato Maurício, E.; Diogo, E.; Santos, M.; Vasilenko, P.; Cruz, A.S.; Mendes, B.; Gonçalves, M.; Duarte, M.P. Potential application of propolis extracts to control the growth of Stemphylium vesicarium in “Rocha” pear. Appl. Sci. 2020, 10, 1990. [Google Scholar] [CrossRef] [Green Version]
- Marino, A.K.; Junior, J.S.P.; Magalhães, K.M.; Mattiuz, B.-H. Chitosan-propolis combination inhibits anthracnose in “Hass” avocados. Emir. J. Food Agric. 2018, 681. [Google Scholar] [CrossRef]
- Hoa, H.T.; Wang, C.-L. The effects of temperature and nutritional conditions on mycelium growth of two oyster mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus). Mycobiology 2015, 43, 14–23. [Google Scholar] [CrossRef]
Figure 1.
Phylogenic tree showing the relationships between Neofabraea spp. isolates published in GenBank and the isolates obtained in this study (CP5, CP8, CP9, CP13, FU11, and GA2A), inferred by the neighbor-joining method based on the partial sequencing of the β-tubulin gene. Pezicula cinnamomea was used as an outgroup.
Figure 1.
Phylogenic tree showing the relationships between Neofabraea spp. isolates published in GenBank and the isolates obtained in this study (CP5, CP8, CP9, CP13, FU11, and GA2A), inferred by the neighbor-joining method based on the partial sequencing of the β-tubulin gene. Pezicula cinnamomea was used as an outgroup.

Figure 2.
Effect of temperature on the growth and conidial production of Phlyctema vagabunda on tomato agar medium, 14 days post inoculation.
Figure 2.
Effect of temperature on the growth and conidial production of Phlyctema vagabunda on tomato agar medium, 14 days post inoculation.

Figure 3.
Disease severity (rot diameter) of Chilean CP13, CP9, CP5, and CP8 isolates of Phlyctema vagabunda in ‘Cripps Pink’ apples incubated at 20 °C for 20 days. Letters on bar indicate statistical differences between isolates after Tukey test (p < 0.05).
Figure 3.
Disease severity (rot diameter) of Chilean CP13, CP9, CP5, and CP8 isolates of Phlyctema vagabunda in ‘Cripps Pink’ apples incubated at 20 °C for 20 days. Letters on bar indicate statistical differences between isolates after Tukey test (p < 0.05).

Figure 4.
Disease severity of Chilean CP13 isolate of Phlyctema vagabunda in ‘Cripps Pink’ apples inoculated with sterile distilled water (A), mycelium disc (B), and conidia suspension (C). The fruit was incubated at 20 °C for 20 days.
Figure 4.
Disease severity of Chilean CP13 isolate of Phlyctema vagabunda in ‘Cripps Pink’ apples inoculated with sterile distilled water (A), mycelium disc (B), and conidia suspension (C). The fruit was incubated at 20 °C for 20 days.

Figure 5.
Phenogram of the cluster analysis of the inhibition of the mycelial growth of Phlyctema vagabunda (CP13 isolate) by the four propolis extracts (RÑ15, RÑ16, RL15, and RL16) at different concentrations (0.01, 0.05, 0.1, 0.2, 0.25, 0.3, and 0.5%). Average—average linkage and Euclidean distance. Means with the different letter is showing a significant difference (p < 0.05) between clusters.
Figure 5.
Phenogram of the cluster analysis of the inhibition of the mycelial growth of Phlyctema vagabunda (CP13 isolate) by the four propolis extracts (RÑ15, RÑ16, RL15, and RL16) at different concentrations (0.01, 0.05, 0.1, 0.2, 0.25, 0.3, and 0.5%). Average—average linkage and Euclidean distance. Means with the different letter is showing a significant difference (p < 0.05) between clusters.

Figure 6.
Effect of propolis extracts LR16 at 0% (A), 0.005% (B), 0.01% (C), 0.05% (D), and 0.1% (E) on the germination of Phlyctema vagabunda (CP13 isolate) conidia. The cultures were incubated at 20 °C for 4 days.
Figure 6.
Effect of propolis extracts LR16 at 0% (A), 0.005% (B), 0.01% (C), 0.05% (D), and 0.1% (E) on the germination of Phlyctema vagabunda (CP13 isolate) conidia. The cultures were incubated at 20 °C for 4 days.

Figure 7.
Effect of propolis extracts (LR16) at different concentrations on the germination of Phlyctema vagabunda (CP13 isolate) conidia. Lowercase letters indicate statistical differences between propolis concentrations after Tukey test (p < 0.05).
Figure 7.
Effect of propolis extracts (LR16) at different concentrations on the germination of Phlyctema vagabunda (CP13 isolate) conidia. Lowercase letters indicate statistical differences between propolis concentrations after Tukey test (p < 0.05).

Figure 8.
Incidence (A) and severity (B) of Phlyctema vagabunda (CP13 isolate) in apples treated with 0.5% propolis LR16. Letters indicate statistical differences (t-test, p < 0.05) between treatments after 20 days of incubation at 20 °C.
Figure 8.
Incidence (A) and severity (B) of Phlyctema vagabunda (CP13 isolate) in apples treated with 0.5% propolis LR16. Letters indicate statistical differences (t-test, p < 0.05) between treatments after 20 days of incubation at 20 °C.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effect of temperature on mycelial growth and conidial production of Phlyctema vagabunda. The isolates were cultured on tomato agar at 0, 5, 10, 15, 20, 25, and 30 °C for 14 days.
Table 1.
Effect of temperature on mycelial growth and conidial production of Phlyctema vagabunda. The isolates were cultured on tomato agar at 0, 5, 10, 15, 20, 25, and 30 °C for 14 days.
| Temperature (°C) | Isolate | Mean (SE) | Conidial Production |
|---|---|---|---|
| 20 | CP 13 | 20.50 (0.5) a | − |
| 15 | CP 5 | 19.92 (0.36) ab | − |
| 20 | CP 8 | 19.67 (0.66) ab | − |
| 15 | CP 9 | 19.50 (1.00) ab | − |
| 15 | CP 13 | 19.50 (0.00) ab | − |
| 15 | CP 8 | 19.17 (0.44) ab | − |
| 20 | CP 9 | 19.17 (0.22) ab | − |
| 20 | CP 5 | 17.75 (0.8) b | − |
| 10 | CP 9 | 15.00 (0.00) c | − |
| 25 | CP 5 | 14.50 (1.00) c | − |
| 10 | CP 8 | 14.33 (0.08) cd | − |
| 10 | CP 5 | 13.58 (0.22) cd | − |
| 10 | CP 13 | 12.92 (0.30) cd | − |
| 25 | CP 9 | 11.67 (0.16) de | − |
| 25 | CP 13 | 9.83 (0.88) ef | − |
| 25 | CP 8 | 8.17 (0.22) fg | − |
| 5 | CP 5 | 6.33 (0.66) gh | + |
| 5 | CP 8 | 5.50 (0.00) ghi | + |
| 5 | CP 9 | 5.50 (0.28) ghi | + |
| 5 | CP 13 | 5.17 (0.92) hi | + |
| 0 | CP 13 | 3.83 (0.16) hi | + |
| 0 | CP 8 | 3.50 (0.50) i | + |
| 0 | CP 9 | 3.25 (0.38) i | + |
| 0 | CP 5 | 3.00 (0.00) i | + |
| 30 | CP 9 | 0.00 (0.00) j | − |
| 30 | CP 13 | 0.00 (0.00) j | − |
| 30 | CP 8 | 0.00 (0.00) j | − |
| 30 | CP 5 | 0.00 (0.00) j | − |
Data represent the average of the isolates. Different letters indicate growth differences for an isolate at different temperatures based on Tukey HSD test (p < 0.05). Symbol + mean conidia production and symbol − mean no conidial.
Table 2.
Population and morphological characterization of conidia produced by the isolates CP 5, CP 8, CP 9, and CP 13 at 0 and 5 °C after 14 days of incubation in tomato agar.
Table 2.
Population and morphological characterization of conidia produced by the isolates CP 5, CP 8, CP 9, and CP 13 at 0 and 5 °C after 14 days of incubation in tomato agar.
| Isolate | Growth Temperature (°C) | Conidia N° Colonies−1 | Length (µm) | Width (µm) |
|---|---|---|---|---|
| CP 5 | 0 | 1.44 × 105 ± 2.6 × 104 ab | 10.42 ± 1.9 a | 2.74 ± 0.2 a |
| CP 8 | 0 | 1.78 × 105 ± 1.0 × 104 a | 10.42 ± 1.4 a | 2.97 ± 0.1 a |
| CP 9 | 0 | 9.68 × 104 ± 2.2 × 103 b | 9.99 ± 1.9 a | 2.79 ± 0.1 a |
| CP 13 | 0 | 1.03 × 105 ± 1.4 × 103 b | 10.78 ± 2.5 a | 3.06 ± 0.1 a |
| CP 5 | 5 | 1.05 × 105 ± 1.3 × 104 b | 10,94 ± 1.6 a | 2.87 ± 0.2 a |
| CP 8 | 5 | 1.34 × 105 ± 2.5 × 104 ab | 10,10 ± 2.4 a | 2.83 ± 0.2 a |
| CP 9 | 5 | 9.91 × 104 ± 3.3 × 103 b | 10.72 ± 0.9 a | 2.87 ± 0.2 a |
| CP 13 | 5 | 2.02 × 105 ± 8.6 × 102 a | 11.11 ± 2.5 a | 3.05 ± 0.3 a |
Different letters in the same column indicate statistical differences according to Tukey HSD test (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Urrea, I.; Arismendi, N.; Sepúlveda, X.; Gerding, M.; Vero, S.; Vargas, M. Antifungal Activity of Propolis Extracts against Postharvest Pathogen Phlyctema vagabunda. Agronomy 2023, 13, 104. https://doi.org/10.3390/agronomy13010104
AMA Style
Urrea I, Arismendi N, Sepúlveda X, Gerding M, Vero S, Vargas M. Antifungal Activity of Propolis Extracts against Postharvest Pathogen Phlyctema vagabunda. Agronomy. 2023; 13(1):104. https://doi.org/10.3390/agronomy13010104
Chicago/Turabian StyleUrrea, Iván, Nolberto Arismendi, Ximena Sepúlveda, Macarena Gerding, Silvana Vero, and Marisol Vargas. 2023. "Antifungal Activity of Propolis Extracts against Postharvest Pathogen Phlyctema vagabunda" Agronomy 13, no. 1: 104. https://doi.org/10.3390/agronomy13010104
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.