
Predacious Strategies of Nematophagous Fungi as Bio-Control Agents

1
Co-Innovation Center for Sustainable Forestry in Southern China, Department of Forest Protection, College of Forestry, Nanjing Forestry University, Nanjing 210073, China
2
College of Life Science, Nanjing Forestry University, Nanjing 210073, China
*
Author to whom correspondence should be addressed.
Agronomy 2023, 13(11), 2685; https://doi.org/10.3390/agronomy13112685
Submission received: 29 September 2023
/
Revised: 23 October 2023
/
Accepted: 23 October 2023
/
Published: 25 October 2023
(This article belongs to the Special Issue Nematode Diseases and Their Management in Crop Plants)
Abstract
:Plant-parasitic nematodes significantly threaten agriculture and forestry, causing various diseases. They cause annual losses of up to 178 billion dollars worldwide due to their parasitism. Nematophagous fungi (NF) are valuable in controlling or reducing parasitic nematode diseases by killing nematodes through predatory behavior. This article summarizes the strategic approaches adopted by NF to capture, poison, or consume nematodes for food. NF are classified based on their attacking strategies, including nematode trapping, endoparasitism, toxin production, and egg and female parasitism. Moreover, extracellular enzymes such as serine proteases and chitinases also play an important role in the fungal infection of nematodes by disrupting nematode cuticles, which act as essential virulence factors to target the chemical constituents comprising the nematode cuticle and eggshell. Based on the mentioned approaches, it is crucial to consider the mechanisms employed by NF to control nematodes focused on the use of NF as biocontrol agents.
1. Introduction
Nematodes are highly ubiquitous animal groups found throughout the planet. Despite being over 28,000 known species, they are challenging to differentiate. Out of all these species, 16,000 are recognized as parasitic towards animals, insects, or plants [1]. Over 4000 species of plant-parasitic nematodes (PPNs) have been identified [2] which severely threaten agricultural and horticultural crops [3]. These harmful pathogens are hideous crop parasites that cause global annual losses between USD 78 and 125 billion [4,5]. PPNs can actively move in the soil region adjacent to the plant roots, known as the rhizosphere, as well as on the aerial parts of the plant, or within the plant itself. They feed and breed on living organisms, with some nematodes feeding on plant parts such as flowers, leaves, stems, and seeds [6]. However, most of these worms feed on underground plant parts like roots, bulbs, and tubers, causing substantial plant damage ranging from mild harm to destruction [5].
Synthetic nematicides are highly effective at controlling PPNs. However, their widespread impact, toxic environmental effects, and significant government restrictions necessitate exploring alternative measures for PPN control [7]. Pathogenic factors against insects, nematodes, and vertebrates have been documented in bacterial or fungal pathogens [8,9,10]. Biological control agents (BCAs) are promising candidates for managing PPNs due to their environmentally friendly and economically sustainable nature. Many fungi can cause diseases in various nematode groups. About 200 taxonomically diverse species can attack active nematodes [11]. Nematophagous fungi (NF) are natural nematode antagonists and offer promising biocontrol strategies [12]. NF are classified based on the mechanisms by which they invade nematodes: (1) nematode-trapping fungi use morphological hyphae traps, (2) endoparasitic fungi employ spores, (3) egg-parasitic fungi attack nematode eggs or females, and (4) toxin-generating fungi restrain nematodes before invasion [13,14]. NF have long been utilized as biological control agents against PPNs [12,15,16] and animal parasitic nematodes [17].
Extracellular enzymes play a crucial role in the fungal infection of nematodes by disrupting nematode cuticles’ physical and physiological integrity, thereby promoting fungal penetration and colonization [9,10]. It has been shown that extracellular enzymes, such as serine proteases, chitinases, and collagenases act as essential virulence factors that can target the chemical constituents comprising the nematode cuticle and eggshell [18]. Serine proteases are enzymes that hydrolyze peptide bonds catalytically by utilizing a serine residue in the substrate-binding pocket that is specially activated [19,20]. Chitin serves as the structural component of PPNs and their eggshells. Enzymes known as chitinases are found in fungi that parasitize eggs and are utilized to penetrate the eggshells of nematodes. In fact, the first chitinase, Chi43 enzyme, was isolated from two nematode-consuming fungi, Planktothrix rubescens and Pochonia chlamydosporia [21].
Based on previous studies, we have assessed NF as a valuable biocontrol agent and have shown that it has tremendous potential for controlling pathogenic nematodes as an alternative, sustainable, and practical approach.
2. Taxonomic Classification of Nematophagous Fungi
Nematophagous fungi can be used as highly effective biological control agents. They are classified into various fungal taxa groups, including Ascomycota, Oomycota, Pleurotaceae (Basidiomycota), Chytridiomycetes, and Zygomycota [22,23,24]. The taxonomic information for nematophagous fungi is summarized in Figure 1, which includes their division, genus, interaction, and infection structures.
3. Isolation and Characterization of NF
3.1. NF Isolation Using Selective Media
NF have been found in various habitats, such as deciduous leaf litter, coniferous leaf litter, partly revegetated dung, permanent pasture, temporary agricultural grassland, cultivated land, moss cushions, decaying vegetation, and compost peatland coastal vegetation. Coniferous leaf litter has the highest number of these fungi, while peat and dung have the greatest species diversity [26].
The first step in isolating nematophagous fungi is to collect soil or other environmental samples from regions known to harbor nematodes, which are typically obtained from agricultural fields, forests, or gardens. While various techniques can be used for isolating NF, the soil sprinkling and Baermann funnel techniques were the most efficient methods, which can isolate all the predator and endoparasite species with 95% probability [27]. The Baermann funnel technique is more effective than the soil sprinkling method in isolating endoparasitic NF, but it does not capture all endoparasites [28]. Isolated samples are processed using selective media that encourage the growth of NF. Commonly used media include cornmeal agar, potato dextrose agar, and yeast extract agar [29]. The incubation of NF under controlled conditions (23 °C to 26 °C) for several weeks enables them to develop and reproduce [30].
3.2. Morphological and Molecular Characterization
Morphological characterization is a fundamental step in the identification and classification of NF, allowing researchers to distinguish them from non-nematophagous species and understand their functional structures. Fungal colonies are identified based on their physical characteristics, specifically the shape and structure of their spores. These features are critical for distinguishing between different types of NF. In some cases, microscopic examination may be necessary to identify them accurately. The spores possess a diverse range of shapes, sizes, and structures, which may be round, oval, or elongated, and often bear specialized structures or bumps that successfully adhere to nematodes [31].
Similarly, molecular identification is also essential for the accurate identification of NF. In previous studies, a PCR assay using species-specific primers was developed for the rapid and accurate identification of NF Duddingtonia flagrans, which was capable of detecting the fungus in various environmental samples [32]. Similarly, NF Arthrobotrys sinense was studied as a potential biocontrol agent of domestic animal nematode Haemonchus contortus, and five isolates of Arthrobotrys sinense were identified and characterized using molecular techniques [33]. Moreover, a group of nematophagous fungi isolated from soil and animal feces in Mexico were studied to determine their predatory ability against Panagrellus redivivus. The molecular identification of nine isolates was accomplished by amplifying the 18S, 5.8S, and 28S regions with ITS5 and ITS4 oligonucleotides, followed by BLAST sequencing and alignment [34].
4. Nematophagous Fungi (NF)
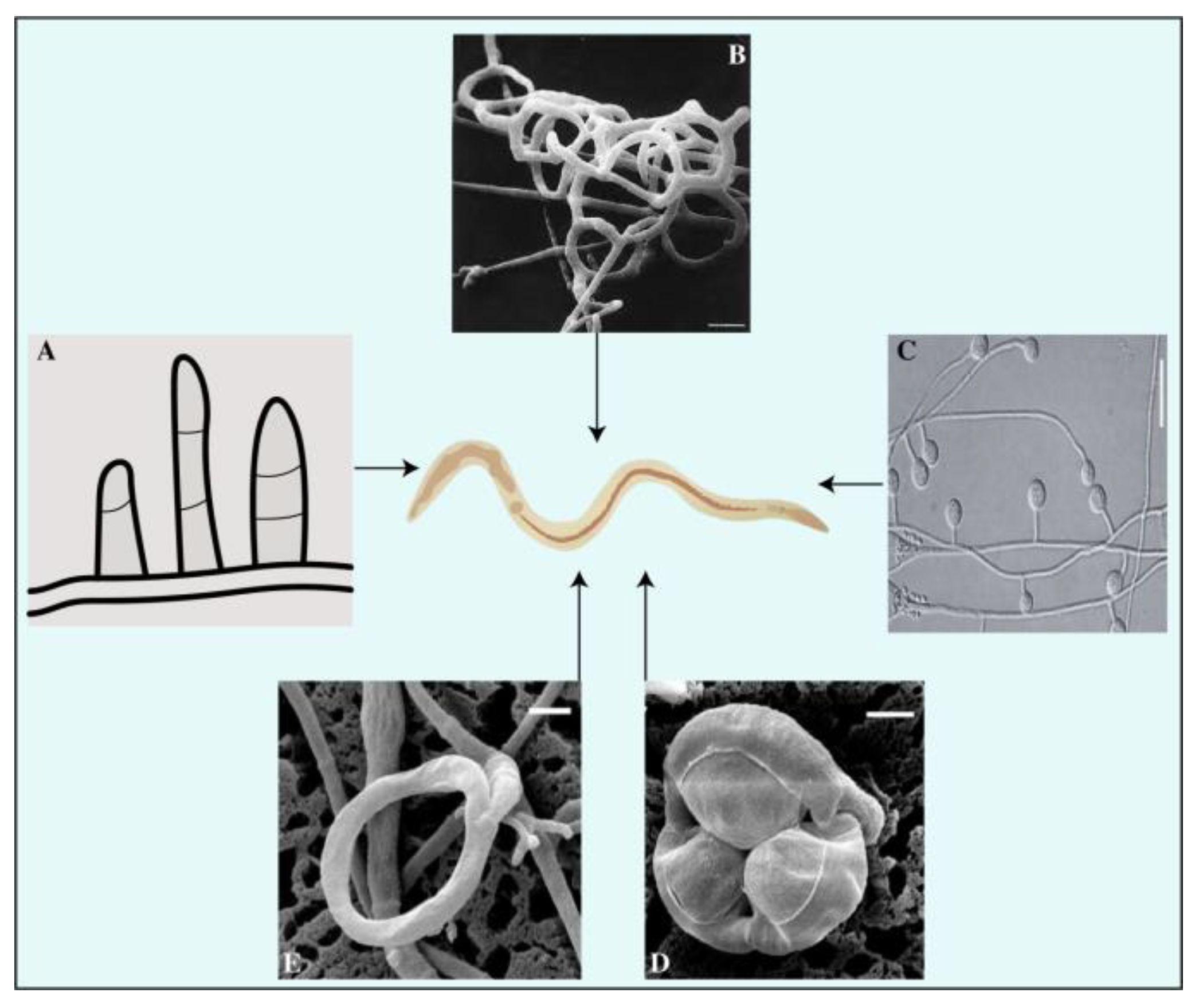
NF are capable of capturing, killing, and digesting nematodes [35]. They reside externally and internally within the host organism, exploiting it for sustenance. These fungi use specific traps to ensnare prey, hyphae tips to parasitize females and eggs, and conidia to adhere while generating toxins to attack nematodes. Based on these strategies, NF are traditionally classified into four groups (Figure 2): (1) The group of fungi that prey on nematodes using specialized traps; (2) some fungi are egg-parasitic and invade nematode eggs or females through their hyphal tips, (3) while others are endoparasitic and use their spores. (4) Additionally, toxin-producing fungi immobilize nematodes before invading them [14,36].
4.1. Predatory Fungi
Predatory fungi employ hyphal structures in nematodes. Studies on nematode-trapping fungi have drawn much attention to their diverse and intricate catching structures [37]. The traps generated by the fungi’s mycelium adversely affect the nematode’s cuticle. The mycelium proliferates within the nematode’s body, resulting in the formation of a penetration peg, whose growth over time causes the hyphae to cover the outer surface of the colonized nematode [38]. The fungus A. oligospora has a unique mechanism for penetrating the cuticle of nematodes via penetrating tubes, and its impact on Meloidogyne javanica in tomato cultivars has been empirically demonstrated [39]. Predatory structures are vital for the life and activity of trapping fungi. Compared to regular hyphae, adhesive traps have a longer lifespan [40,41]. These specialized traps are utilized by over two hundred fungal species (found within the Zygomycota, Basidiomycota, and Ascomycota phyla) to capture free-living nematodes in soil [42]. NF play a vital role in maintaining the population of nematodes through natural methods, such as parasitism, trapping, and poisoning [43,44]. Basidiomycota trapping fungi use spores and adhesive knobs to capture nematodes [45,46,47]. The Orbiliaceae family comprises over 80% of nematode-trapping fungi within the Ascomycota Phylum, where constricting rings, adhesive networks, adhesive branches, adhesive knobs, and non-constricting rings are all used to ensnare nematodes (Figure 2) [48,49]. The study of nematode-trapping fungi in Zygomycota has faced obstacles due to inadequate isolation and culture techniques, despite the growing interest in it [50].
4.1.1. Adhesive Branches
Adhesive branches, also known as adhesive columns, have a simpler morphological structure than other capture organs (Figure 2). These vertical branches consist of one to three cells that merge via anastomosis, forming adhesive hoops or networks with two-dimensional structures resembling crochet or lines. The nematode is easily captured upon contact with the branch due to its complete coverage by a delicate adhesive layer. Due to the proximity of adhesive branches, nematodes frequently become stuck to more sticky hyphae upon contact and struggle to detach themselves. The species commonly found in temperate soils with developed adhesive branches is Dactylella cionopaga [51]. These branches serve as typical trapping mechanisms for Monacrosporium cionopagum and M. gephyrophagum [52]. For example, M. cionopagum produces adhesive branches that trap and immobilize the sugar beet cyst nematode Heterodera schachtii [53]. Likewise, Gamsylella gephyropaga produces adhesive branches to trap nematodes [54,55].
4.1.2. Adhesive Hyphal Network
The adhesive network, widely distributed in fungi, is comprised of an upright lateral branch emerging from a vegetative hypha (Figure 2), extending approximately 20–25 μm from the parent hypha [56], and is characterized by a longer lifespan in comparison to typical hyphae [40]. These adhesive nets are constructed using intricate three-dimensional networks. A. oligospora, with a global distribution, is the most frequently observed species in this specific trapping structure [57]. Adhesive nets form from vegetative hyphae by curving a solitary lateral branch and can combine with parental hyphae. Adhesive nets are regarded as an evolutionary progression from adhesive branches. More lateral hyphae are generated from the parental hyphae, or a loop is formed to generate additional loops once a complex of interconnected loops that extend away from the potential hyphae in all logical directions is established. Nematodes are attracted to the network’s surface, which is coated with a thin layer of adhesives [58].
4.1.3. Adhesive Knobs
Adhesive knobs are specialized cells with a small layer of adhesive covering them (Figure 2). When a nematode becomes ensnared, the contact area between it and the spherical knob is limited, allowing it to resist and free itself. However, upon coming into contact with a flattened, sticky pad, the fungus takes control and traps the nematode. This significantly increases the adhering surface, resulting in a secure binding of the captured nematode, followed by fungal penetration that involves both enzymatic and physical mechanisms. For example, the fungus synthesis of collagenase aids in penetrating the nematode’s cuticle, while the dense sticky pad provides strength and rigidity, allowing the piercing hyphae to move toward the cuticle [51]. Once a spherical infection bulb has formed, assimilative hyphae emerge to consume the internal contents of the nematode [31,59]. Dactylellina arcuata, D. asthenopaga, D. leptosphora, D. copepodii, and D. ellipsospora use adhesive knobs to capture nematodes [60,61,62].
4.1.4. Constricting Rings
Constricting rings are hyphal branches with a circular arrangement typically composed of three cells (Figure 3). These structures are highly sophisticated and actively capture prey. A nematode entering the cavity triggers the rapid expansion of the three surrounding cells, resulting in a threefold increase in their volume. This process effectively seals the orifice and confines the nematode inside the cavity. Subsequently, the hyphae penetrate and assimilate the nematode [63]. Twelve species of hyphomycetes have been discovered to form constricting rings of varying internal diameters ranging from 20 to 40 μm [51]. Constricting rings are distinct from other mechanisms because they encircle nematodes upon contact with the inner edge of the trap, resulting in closure. The nematode is strangled by the expansion of an internally located cell wall of a constricting ring, causing inward swelling through the outer cell wall of these rings. Rapid water intake causes an increase in the volume of the cells forming the ring [64]. The traps of D. brochopaga mutants are considerably larger, almost eight times more than conventional traps. It has been observed that the cells in these traps release fluid droplets, reducing their volume. The humidity of the surrounding environment can be adjusted to facilitate the entry or exit of atmospheric water into or out of these cells. Additionally, a correlation has been established between the ambient humidity level and the frequency of ring closures [65]. The source of the water supply has been investigated, and evidence suggests that it mainly originates primarily from stalk cells or mycelium [66]. Furthermore, once the stalk cell closed, there was no noticeable movement of internal components, suggesting that water from the surrounding environment may have been retained [67]. This is supported by the observation that rings can continue to spread even after being detached from the original stalk on which they first appeared [65,68]. This idea is plausible as live nematodes are typically surrounded by a thin layer of water, which could serve as a sufficient source of moisture for the process of ring closure. Additionally, the ring closure process in D. brochopaga can be chemically induced, in addition to physical methods like touch, elevated temperature, or electrical stimulation [69]. When exposed to solutions containing methanol, ethanol, propanol, or butanol, or to chlorobutanol vapor, this fungus’s traps expanded in 10 to 15 s. In contrast, it is important to note that benzene, ether, and chloroform did not have any detectable effect, suggesting that unidentified variables drive this significant phenomenon [70].
4.1.5. Non-Constricting Rings
Non-constricting rings are a type of three-celled rings that grow on a short supportive stalk originating from prostrate septate hyphae (Figure 3D). The nematodes display passive behavioral responses during the predation process. As noted, the attachment point between the supporting stalk and the ring was weakened. As the nematode attempted to free itself, the ring often detached, indicating that the fungus may have facilitated its escape by allowing it to carry the non-constricting ring tightly wrapped around its body. This seems to be a favored method for achieving widespread dissemination in the soil [71]. For example, fungi, such as Dactylaria candida and D. lysipaga, which produce non-constricting rings, often create adhesive knobs [60,72] and capture nematodes using non-constricting rings [61]. A similar pattern was observed in Dactylellina daliensis via non-constricting rings [73].

4.2. Egg- and Female-Parasitic Fungi
Research on egg- and female-parasitic fungi has been in progress since the 1990s. These fungi employ appressoria (Purpureocillium spp. and Pochonia spp.), zoospores (Nematophthora gynophila), lateral mycelial branches, and penetration pegs to parasitize eggs, females, and other growth stages of the PPNs [78]. An in vitro assessment was conducted to evaluate the parasitism of 10 isolates of P. chlamydosporia on Globodera pallida eggs; the levels of observed pathogenicity ranged from 34% to 49%. The event of impulsive hatching occurs when P. chlamydosporia isolates aggressively parasitize immature eggs as opposed to those containing second-stage juveniles [79]. Additionally, the use of wild-type Beauveria bassiana 08F04 and transformant G10 resulted in a substantial decrease in the cereal cyst nematode (female) population in the roots [80]. In a greenhouse study, it was found that the presence of the arbuscular mycorrhizal fungus (AMF) Glomus etunicatum reduced the population of H. glycines female nematodes by 28.21% in root systems compared to untreated roots. This finding suggests that G. etunicatum may play a role in promoting the ability of host plants to tolerate the presence of the soybean cyst nematode (SCN) [81]. Figure 4 displays fungal species that parasitize the egg and female.
4.3. Endoparasitic Fungi
Endoparasitic fungi are a category of nematophagous fungi which infect nematodes by producing spores. These spores can either be internalized by the nematodes through ingestion, leading to infection, or attach to the nematode epidermis, initiating the infection [14,82,83]. Endoparasites utilize spores, such as conidia and zoospores, for infection, which may attach to the nematode cuticle or be ingested [84]. It has been found that endoparasitic fungi can reduce the number of root-knot nematodes that create galls on tomatoes and alfalfa in greenhouse experiments [14]. These fungi exhibit varying degrees of diversity, with studies indicating differences in their production of conidia per infected nematode. D. coniospora fungi produce a significant amount of conidia, with up to 10,000 per hyphal material, while H. rhossoliensis yields 100–1000 conidia per infected nematode. Conidia germinate immediately, and assimilative hyphae infiltrate and absorb the entire contents of the nematode body, enabling the fungus to penetrate the host’s outer layer [85]. D. coniospora is an aggressive endoparasitic fungus that targets nematodes. The endoparasitic fungus Drechmeria coniospora YMF1.01759 strain exhibited excellent nematode-infecting ability. The study revealed that it hindered nematodes from hatching their eggs, infected them with spores, and produced active metabolites that killed them [86].
4.4. Toxin Production
Some nematophagous fungi produce toxins that kill nematodes and impact plant defense and resistance mechanisms against parasitic nematodes [87,88,89]. Toxin-producing fungi originate from various orders and families. The fungus assaults nematodes via the secretion of inhibitory metabolites without physically interacting and immobilized them [78,90]. After immobilization, the hyphae penetrate the nematode cuticle. Culture filtrates of these fungi contain strong enzymatic (proteolytic and chitinolytic) activities, low-molecular-weight metabolites, and specific non-volatile oil components that cause larval death or inhibit egg hatching [91]. The metabolites secreted by the fungi alter the composition of nematode eggs and prevent embryonic development, rendering them unable to hatch due to their varying shapes and sizes. Similarly, fungi produce toxic chemicals, other than enzymes, that immobilize nematodes and later consume them [92]. Basidiomycetes are the predominant fungi that produce toxins. Recent research on Basidiomycetous fungi (Coprinus comatus and Stropharia rugosoannulata) has revealed that the action mechanisms of these toxins against nematodes are varied and multifaceted [93]. Among Basidiomycetes, numerous Pleurotus species produce toxins with nematotoxic activity [92,94]. For example, P. ostreatus produces trans-2-decenoic acid, a compound obtained from linoleic acid that is detrimental to nematodes, insects, and other fungi [95]. Basidiomycetes are not the only fungi that generate these kinds of toxins; some fungi also produce toxins that are harmful to nematodes, but these are not nematophagous [38]. These compounds exhibit diverse chemical properties, including simple fatty acids or other organic acids such as lactones, pyrones, anthraquinones, benzoquinones, alkaloids, furans, peptaibiotics, and cyclodepsipeptides.
5. Enzymes of NF
Some NF use enzymes to facilitate the infection and digestion of nematodes [38,84]. Nematodes possess physical barriers, consisting of abundant proteins in their composition, that protect them from natural predators [96]. NF penetrate the nematode cuticle and eggshell via enzymatic (protease and chitinase) and mechanical means [78] (Figure 5). The enzymes responsible for breaking down nematode cuticles include alkaline and neutral serine proteases, which facilitate the hydrolysis of the peptide bonds in the cuticle protein [97]. Serine proteases hydrolyze peptide bonds by utilizing a serine residue specifically triggered in the substrate-binding pocket [19,20]. The alkaline serine protease triggers the destruction of cuticles within hours and restrains the nematode P. redivivus [98]. Neutral serine protease produced by Arthrobotrys oligospora causes pathogenicity against nematodes [99]. Arthrobotrys oligospora can control Haemonchus contortus and Caenorhabditis elegans under laboratory conditions [100,101]. The fungus Monacrosporium thaumasium produces a high-level serine protease that has a destructive effect on M. javanica eggs, as demonstrated by [102]. These studies highlight the importance of serine protease as a crucial enzyme in the progression of fungus-initiated infection. The shells of nematode eggs contain high levels of chitin and proteins, which are hydrolyzed by endochitinases and exochitinases, catalyzing the glycosidic bonds between chitin N-acetylglucosamine groups [21]. The nematode-trapping fungus M. thaumasium produces chitinases and exhibits nematicidal activity against the nematode Panagrellus redivivus [103]. Furthermore, chitinases have demonstrated nematicidal activity when used without the presence of fungi [104]. These enzymes are also crucial in the development of infection and shell digestion of nematodes [105].
6. Biocontrol Using NF
Biocontrol is considered to be an eco-friendly and sustainable alternative to pesticides [106]. It involves using microbial species, particularly fungus, which has shown significant antagonistic effects against PPNs [107]. Several types of fungi can be harmful to nematodes, but simply being aggressive is not enough to qualify as a dependable biocontrol agent [108]. Two main methods for implementing the use of nematode-killing fungi for nematode biocontrol are adding fungi to the soil or supporting the activity of already-existing fungi via various alterations. Early research on controlling PPNs used fungi, such as Arthrobotrys and Monacrosporium species that trap nematodes. Later, endo-parasitic fungi such as H. rhossoliensis and D. coniospora, and egg-parasitic fungi P. chlamydosporia, were also utilized as biocontrol agents. The efficacy of these agents varies, as reported in [109,110,111]. Due to an advancement in the formulation and soil application of fungal biocontrol agents, as well as a growing comprehension of the biology of these fungi, there is a heightened interest in utilizing nematode-trapping fungi. Additional research is needed to further improve the effectiveness of these biocontrol agents (Table 1).
7. Limitations and Future Directions
One significant limitation of NF as bio-control agents is their often limited host range. Many species are highly specialized to target specific nematode species. This restricts their applicability in situations where multiple nematode species are present. To address this limitation, future research could focus on isolating and characterizing strains with broader host ranges or exploring ways to enhance their adaptability to different nematode species. Techniques such as genetic manipulation and selective breeding may be explored to achieve this diversified goal. The efficacy of NF can be influenced by various environmental conditions as these fungi are susceptible to temperature, humidity, and soil type. Extreme conditions such as extremely dry or hot environments can limit their ability to establish and reproduce effectively. Researchers should investigate strategies to improve the resilience of NF to adverse environmental conditions, possibly through the development of more robust strains or innovative application techniques. NF generally act more slowly than chemical nematicides, which can be a disadvantage when rapid nematode control is required, especially in high-value agricultural settings. In the future, research could focus on enhancing the speed of action of these fungi, possibly by developing more virulent strains or optimizing application methods. The effectiveness of NF can vary from one application to another, where factors such as soil microbial communities and nematode densities can influence their success. These factors should be investigated in more detail to develop strategies that can improve the consistency of nematode control using fungi, which may involve the development of tools for monitoring and predicting fungal performance under different conditions. The registration and approval of NF for commercial use can be challenging and time-consuming due to regulatory requirements and the need for safety assessments. Hence, collaboration between researchers, regulatory agencies, and industry stakeholders is essential to streamline this process and facilitate the adoption of NF as biocontrol agents.
Additionally, developing new and improved formulations should be explored for NF, such as encapsulation or granules, which can ultimately enhance their stability and shelf life, making them more practical for field applications. Research into novel delivery methods, such as sprays or seed coatings, can also help ensure efficient distribution.
Deep investigation should be encouraged to show insights into the impacts of NF on non-target organisms and the environment to ensure their safety and environmental compatibility. Understanding their ecological interactions in greater detail can guide the development of safer and more sustainable bio-control strategies. Exploring biotechnological tools is necessary to enhance the virulence and adaptability of NF to different environmental conditions and nematode species, which can involve identifying and manipulating the genes responsible for crucial predation mechanisms. In addition, integrating NF with other nematode management strategies, such as crop rotation, resistant crop varieties, and chemical nematicides, should be conducted to create more comprehensive and effective nematode control programs where integrated pest management approaches can be developed and tested for specific agricultural systems. Interdisciplinary work and collaboration with regulatory bodies should be prioritized to streamline the approval process for NF-based products, making them more accessible to farmers with industry partners to ultimately scale up production and distribution channels for these bio-control agents. At last, extensive field trials should be conducted to demonstrate the effectiveness of NF in real-world conditions and encourage their adoption by farmers, which requires a proper monitoring system of the long-term effects on soil health and crop productivity to provide practical recommendations for sustainable nematode management.
8. Conclusions
Nematophagous fungi are known for their predatory nature against nematodes. They adopt different approaches for capturing nematodes and gradually consume them. They are divided into groups such as nematode-trapping, eggs- and female-parasitic, endoparasitic, and toxin-producing fungi. Moreover, they also secrete enzymes like serine proteases and chitinases, which immobilize, eventually consume nematodes, and parasitize female eggs. The mentioned knowledge regarding taxonomy, predacious structures, and enzyme involvement of NF is fundamental to be considered while applying them as biocontrol agents against parasitic nematodes in practice.
Author Contributions
The idea was gestated by M.U.R. and B.F.; M.U.R. drafted, and P.C., X.Z. and B.F. helped revise the manuscript. M.U.R. visualized and updated the figures. B.F. supervised the whole study and helped with funding. All authors have read and agreed to the published version of the manuscript.
Funding
This work is supported by the National Natural Science Foundation of China (No. 31970097).
Data Availability Statement
All the data are presented in the main text file.
Acknowledgments
Authors acknowledge the research facilities provided by the Co-Innovation Center for Sustainable Forestry, China.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study, in the collection, analysis, or interpretation of the data, in the writing of the manuscript, or in the decision to publish the results.
References
- Hugot, J.-P.; Baujard, P.; Morand, S. Biodiversity in helminths and nematodes as a field of study: An overview. Nematology 2001, 3, 199–208. [Google Scholar] [CrossRef]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.; Gaur, H.S.; Helder, J.; Jones, M.G.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed]
- Elling, A.A. Major emerging problems with minor meloidogyne species. Phytopathology 2013, 103, 1092–1102. [Google Scholar] [CrossRef] [PubMed]
- Batish, D.R.; Singh, H.P.; Kohli, R.K.; Kaur, S. Eucalyptus essential oil as a natural pesticide. For. Ecol. Manag. 2008, 256, 2166–2174. [Google Scholar] [CrossRef]
- Chitwood, D.J. Research on plant-parasitic nematode biology conducted by the United States Department of Agriculture–Agricultural Research Service. Pest Manag. Sci. Former. Pestic. Sci. 2003, 59, 748–753. [Google Scholar] [CrossRef]
- Palomares-Rius, J.E.; Escobar, C.; Cabrera, J.; Vovlas, A.; Castillo, P. Anatomical alterations in plant tissues induced by plant-parasitic nematodes. Front. Plant Sci. 2017, 8, 1987. [Google Scholar] [CrossRef]
- Chitwood, D.J. Nematicides; Wiley Online Library: Hoboken, NJ, USA, 2003. [Google Scholar]
- Joshi, L.; St Leger, R.J.; Bidochka, M.J. Cloning of a cuticle-degrading protease from the entomopathogenic fungus, Beauveria bassiana. FEMS Microbiol. Lett. 1995, 125, 211–217. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, N.; Zhang, K. Extracellular enzymes serving as virulence factors in nematophagous fungi involved in infection of the host. Res. Microbiol. 2004, 155, 811–816. [Google Scholar] [CrossRef]
- Yang, J.; Tian, B.; Liang, L.; Zhang, K.Q. Extracellular enzymes and the pathogenesis of nematophagous fungi. Appl. Microbiol. Biotechnol. 2007, 75, 21–31. [Google Scholar] [CrossRef]
- KARAKAŞ, M. Nematode-destroying fungi: Infection structures, interaction mechanisms and biocontrol. Commun. Fac. Sci. Univ. Ank. Ser. C Biol. 2020, 29, 176–201. [Google Scholar]
- Degenkolb, T.; Vilcinskas, A. Metabolites from nematophagous fungi and nematicidal natural products from fungi as an alternative for biological control. Part I: Metabolites from nematophagous ascomycetes. Appl. Microbiol. Biotechnol. 2016, 100, 3799–3812. [Google Scholar] [CrossRef] [PubMed]
- Kendrick, B. The Fifth Kingdom, 3rd ed.; Mycologue Publications: Sidney, BC, Canada, 2001. [Google Scholar]
- Liu, X.; Xiang, M.; Che, Y. The living strategy of nematophagous fungi. Mycoscience 2009, 50, 20–25. [Google Scholar] [CrossRef]
- Duponnois, R.; Chotte, J.; Sall, S.; Cadet, P. The effects of organic amendments on the interactions between a nematophagous fungus Arthrobotrys oligospora and the root-knot nematode Meloidogyne mayaguensis parasitizing tomato plants. Biol. Fertil. Soils 2001, 34, 1–6. [Google Scholar] [CrossRef]
- Singh, K.; Jaiswal, R.; Kumar, N.; Kumar, D. Nematophagous fungi associated with root galls of rice caused by Meloidogyne graminicola and its control by Arthrobotrys dactyloides and Dactylaria brochopaga. J. Phytopathol. 2007, 155, 193–197. [Google Scholar] [CrossRef]
- Carvalho, R.; Araújo, J.; Braga, F.; Ferreira, S.; Araujo, J.; Silva, A.; Frassy, L.; Alves, C. Biological control of Ancylostomosis in dogs using the nematode-trapping fungus Monacrosporium thaumasium in southeastern Brazil. Vet. Parasitol. 2009, 165, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liang, L.; Li, J.; Zhang, K.Q. Nematicidal enzymes from microorganisms and their applications. Appl. Microbiol. Biotechnol. 2013, 97, 7081–7095. [Google Scholar] [CrossRef]
- Schultz, R.; Liebman, M. Structure-function relationship in protein families. In Textbook of Biochemistry with Clinical Correlations; Wiley-Liss: New York, NY, USA, 1997; pp. 1–116. [Google Scholar]
- Siezen, R.J.; Leunissen, J.A. Subtilases: The superfamily of subtilisin-like serine proteases. Protein Sci. 1997, 6, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Tikhonov, V.E.; Lopez-Llorca, L.V.; Salinas, J.; Jansson, H.-B. Purification and characterization of chitinases from the nematophagous fungi Verticillium chlamydosporium and V. suchlasporium. Fungal Genet. Biol. 2002, 35, 67–78. [Google Scholar] [CrossRef]
- Gams, W.; Zare, R. A taxonomic review of the clavicipitaceous anamorphs parasitizing nematodes and other microinvertebrates. Clavicipitalean Fungi 2003, 19, 17–74. [Google Scholar]
- Wijayawardene, N.; Hyde, K.; Al-Ani, L.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.; Zhao, R.; Aptroot, A.; Leontyev, D.; Saxena, R. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Yadav, B.; Singh, U.B.; Malviya, D.; Vishwakarma, S.K.; Ilyas, T.; Shafi, Z.; Shahid, M.; Singh, H.V. Nematophagous Fungi: Biology, Ecology and Potential Application. In Detection, Diagnosis and Management of Soil-Borne Phytopathogens; Springer: Berlin/Heidelberg, Germany, 2023; pp. 309–328. [Google Scholar]
- Dackman, C.; Nordbring-Hertz, B. Conidial traps—A new survival structure of the nematode-trapping fungus Arthrobotrys oligospora. Mycol. Res. 1992, 96, 194–198. [Google Scholar] [CrossRef]
- Gray, N. Ecology of nematophagous fungi: Methods of collection, isolation and maintenance of predatory and endoparasitic fungi. Mycopathologia 1984, 86, 143–153. [Google Scholar] [CrossRef]
- Bailey, F.; Gray, N. The comparison of isolation techniques for nematophagous fungi from soil. Ann. Appl. Biol. 1989, 114, 125–132. [Google Scholar] [CrossRef]
- Gray, N.F. Ecology of nematophagous fungi: Comparison of the soil sprinkling method with the Baermann funnel technique in the isolation of endoparasites. Soil Biol. Biochem. 1984, 16, 81–83. [Google Scholar] [CrossRef]
- Doolotkeldieva, T.; Bobushova, S.; Muratbekova, A.; Schuster, C.; Leclerque, A. Isolation, Identification, and Characterization of the Nematophagous Fungus Arthrobotrys oligospora from Kyrgyzstan. Acta Parasitol. 2021, 66, 1349–1365. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Balán, F.; Landeros-Jaime, F.; González-Garduño, R.; Cazapal-Monteiro, C.; Arias-Vázquez, M.S.; Aguilar-Tipacamú, G.; Esquivel-Naranjo, E.U.; Mosqueda, J. High Predatory Capacity of a Novel Arthrobotrys oligospora Variety on the Ovine Gastrointestinal Nematode Haemonchus contortus (Rhabditomorpha: Trichostrongylidae). Pathogens 2021, 10, 815. [Google Scholar] [CrossRef] [PubMed]
- Barron, G.L. The Nematode-Destroying Fungi; Canadian Biological Publications Ltd.: Guelph, ON, Canada, 1977. [Google Scholar]
- Kelly, P.; Good, B.; Fitzpatrick, R.; Hanrahan, J.P.; Waal, T.D.T.D. Development and application of a PCR diagnostic assay for the accurate and rapid identification of the nematophagous fungus Duddingtonia flagrans. Mycol. Res. 2008, 112, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.-J.; Li, E.-L.; Jing, C.-X.; Ma, L.; Cai, K.-Z. Isolation, identification and characterization of the nematophagous fungus Arthrobotrys (Monacrosporium) sinense from China. Acta Parasitol. 2018, 63, 325–332. [Google Scholar] [CrossRef]
- Sánchez-Martínez, E.; Aguilar-Marcelino, L.; Hernández-Romano, J.; Castaeda-Ramírez, G.S.; Mendoza-De-Gives, P. Taxonomic and biological characterization and predatory activity of four nematophagous fungi isolates of Arthrobotrys species from Tapachula, Chiapas, Mexico. Arch. Agron. Soil Sci. 2023, 69, 327–343. [Google Scholar] [CrossRef]
- Nordbring-Hertz, B.; Jansson, H.-B.; Tunlid, A. Nematophagous Fungi. In eLS; Wiley Online Library: Hoboken, NJ, USA, 2011. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.; Askary, T.H. Fungal and bacterial nematicides in integrated nematode management strategies. Egypt. J. Biol. Pest Control 2018, 28, 74. [Google Scholar] [CrossRef]
- Corda, A.K.J. Pracht-Flora Europaeischer, Schimmelbildungen; Bei G. Fleischer: Leipzig, Germany, 1839. [Google Scholar]
- de Freitas Soares, F.E.; Sufiate, B.L.; de Queiroz, J.H. Nematophagous fungi: Far beyond the endoparasite, predator and ovicidal groups. Agric. Nat. Resour. 2018, 52, 1–8. [Google Scholar]
- Mostafanezhad, H.; Sahebani, N.; Nourinejhad Zarghani, S. Induction of resistance in tomato against root-knot nematode Meloidogyne javanica with salicylic acid. J. Crop Prot. 2014, 3, 499–508. [Google Scholar]
- Veenhuis, M.; Nordbring-Hertz, B.; Harder, W. An electron-microscopical analysis of capture and initial stages of penetration of nematodes byArthrobotrys oligospora. Antonie Van Leeuwenhoek 1985, 51, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Bedekovic, T.; Brand, A.C. Microfabrication and its use in investigating fungal biology. Mol. Microbiol. 2022, 117, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, K.; Liu, X. Taxonomy of Nematophagous Fungi; China Science Publishing & Media Ltd: Beijing, China, 2000. [Google Scholar]
- Jaffee, B.; Tedford, E.; Muldoon, A. Tests for density-dependent parasitism of nematodes by nematode-trapping and endoparasitic fungi. Biol. Control 1993, 3, 329–336. [Google Scholar] [CrossRef]
- Linford, M.; Yap, F.; OLIVEIRA, J.M. Reduction of soil populations of the root-knot nematode during decomposition of organic matter. Soil Sci. 1938, 45, 127–142. [Google Scholar] [CrossRef]
- Poloczek, E.; Webster, J. Conidial traps in Nematoctonus (nematophagous Basidiomycetes). Nova Hedwig. 1994, 59, 201–205. [Google Scholar]
- Dürschner-Pelz, U. Traps of Nematoctonus leiosporus—An unusual feature of an endoparasitic nematophagous fungus. Trans. Br. Mycol. Soc. 1987, 88, 129–130. [Google Scholar] [CrossRef]
- Thorn, R.G.; Moncalvo, J.-M.; Reddy, C.; Vilgalys, R. Phylogenetic analyses and the distribution of nematophagy support a monophyletic Pleurotaceae within the polyphyletic pleurotoid-lentinoid fungi. Mycologia 2000, 92, 241–252. [Google Scholar] [CrossRef]
- Hyde, K.D.; Swe, A.; Zhang, K.-Q. Nematode-trapping fungi. In Nematode-Trapping Fungi; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–12. [Google Scholar]
- Swe, A.; Li, J.; Zhang, K.; Pointing, S.; Jeewon, R.; Hyde, K. Nematode-trapping fungi. Curr. Res. Environ. Appl. Mycol. 2011, 1, 1–26. [Google Scholar]
- Saikawa, M. Ultrastructural studies on zygomycotan fungi in the Zoopagaceae and Cochlonemataceae. Mycoscience 2011, 52, 83–90. [Google Scholar] [CrossRef]
- Poinar, G.O.; Jansson, H.-B. Diseases of Nematodes; CRC Press: Boca Raton, FL, USA, 1988; Volume 1. [Google Scholar]
- Saxena, G. Biological control of root-knot and cyst nematodes using nematophagous fungi. In Root Biology; Springer: Berlin/Heidelberg, Germany, 2018; pp. 221–237. [Google Scholar]
- Andersson, K.-M.; Kumar, D.; Bentzer, J.; Friman, E.; Ahrén, D.; Tunlid, A. Interspecific and host-related gene expression patterns in nematode-trapping fungi. Bmc Genom. 2014, 15, 968. [Google Scholar] [CrossRef] [PubMed]
- Jaffee, B.; Strong, D. Strong bottom-up and weak top-down effects in soil: Nematode-parasitized insects and nematode-trapping fungi. Soil Biol. Biochem. 2005, 37, 1011–1021. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, K.-Q.; Hyde, K. The ecology of nematophagous fungi in natural environments. In Nematode-Trapping Fungi; Springer: Berlin/Heidelberg, Germany, 2014; pp. 211–229. [Google Scholar]
- Nordbring-Hertz, B.; Friman, E.; Veenhuis, M. Hyphal fusion during initial stages of trap formation in Arthrobotrys oligospora. Antonie Van Leeuwenhoek 1989, 55, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.-M.; Zhang, K.-Q. Arthrobotrys oligospora: A model organism for understanding the interaction between fungi and nematodes. Mycology 2011, 2, 59–78. [Google Scholar] [CrossRef]
- Pala Martinelli, P.R.; dos Santos, J.M. Scanning electron microscopy of nematophagous fungi associated Tylenchulus semipenetrans and Pratylenchus jaehni. Biosci. J. 2010, 26, 809–816. [Google Scholar]
- Gray, N. Fungi attacking vermiform nematodes. Dis. Nematodes 1988, 2, 3–38. [Google Scholar]
- Li, Y.; Hyde, K.D.; Jeewon, R.; Cai, L.; Vijaykrishna, D.; Zhang, K. Phylogenetics and evolution of nematode-trapping fungi (Orbiliales) estimated from nuclear and protein coding genes. Mycologia 2005, 97, 1034–1046. [Google Scholar] [CrossRef]
- Hai-Yan, W.; Dong-Geun, K.; Xun-Bo, Z. First report of an unrecorded nematode-trapping fungus species Dactylellina candidum in Korea. Afr. J. Microbiol. Res. 2012, 6, 203–205. [Google Scholar] [CrossRef]
- Xie, H.; Aminuzzaman, F.; Xu, L.; Lai, Y.; Li, F.; Liu, X. Trap induction and trapping in eight nematode-trapping fungi (Orbiliaceae) as affected by juvenile stage of Caenorhabditis elegans. Mycopathologia 2010, 169, 467–473. [Google Scholar] [CrossRef]
- Liu, K.; Tian, J.; Xiang, M.; Liu, X. How carnivorous fungi use three-celled constricting rings to trap nematodes. Protein Cell 2012, 3, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Heintz, C.; Pramer, D. Ultrastructure of nematode-trapping fungi. J. Bacteriol. 1972, 110, 1163–1170. [Google Scholar] [CrossRef] [PubMed]
- Insell, J.; Zachariah, K. The mechanism of the ring trap of the predacious hyphomycete Dactylella brochopaga Drechsler. Protoplasma 1978, 95, 175–191. [Google Scholar] [CrossRef]
- Rudek, W.T. The constriction of the trapping rings in Dactylaria brochopaga. Mycopathologia 1975, 55, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Barron, G. Nematophagous fungi: A new Harposporium producing aerial arthroconidia. Can. J. Bot. 1979, 57, 886–889. [Google Scholar] [CrossRef]
- Barron, G. Host range studies for Haptoglossa and a new species, Haptoglossa intermedia. Can. J. Bot. 1989, 67, 1645–1648. [Google Scholar] [CrossRef]
- Higgins, M.; Pramer, D. Fungal morphogenesis: Ring formation and closure by Arthrobotrys dactyloides. Science 1967, 155, 345–346. [Google Scholar] [CrossRef] [PubMed]
- Zachariah, K. Chemical induction of trap closure in Dactylella brochopaga. Protoplasma 1989, 148, 87–93. [Google Scholar] [CrossRef]
- Askary, T.H.; Martinelli, P.R.P. Biocontrol Agents of Phytonematodes; CABI: Egham, UK, 2015. [Google Scholar]
- Drechsler, C. Some hyphomycetes that prey on free-living terricolous nematodes. Mycologia 1937, 29, 447–552. [Google Scholar] [CrossRef]
- Su, H.; Hao, Y.e.; Yang, X.; Yu, Z.; Deng, J.; Mo, M. A new species of Dactylellina producing adhesive knobs and non-constricting rings to capture nematodes. Mycotaxon 2008, 105, 313. [Google Scholar]
- Jiang, X.; Xiang, M.; Liu, X. Nematode-Trapping Fungi. Microbiol. Spectr. 2017, 5, 5-1. [Google Scholar] [CrossRef] [PubMed]
- Nordbring-Hertz, B. Morphogenesis in the nematode-trapping fungus Arthrobotrys oligospora—An extensive plasticity of infection structures. Mycologist 2004, 18, 125–133. [Google Scholar] [CrossRef]
- Li, J.; Zou, C.; Xu, J.; Ji, X.; Niu, X.; Yang, J.; Huang, X.; Zhang, K.-Q. Molecular mechanisms of nematode-nematophagous microbe interactions: Basis for biological control of plant-parasitic nematodes. Annu. Rev. Phytopathol. 2015, 53, 67–95. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhang, W.; Lai, Y.; Xiang, M.; Wang, X.; Zhang, X.; Liu, X. Drechslerella stenobrocha genome illustrates the mechanism of constricting rings and the origin of nematode predation in fungi. BMC Genom. 2014, 15, 114. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Llorca, L.; Maciá-Vicente, J.; Jansson, H.-B. Mode of action and interactions of nematophagous fungi. In Integrated Management and Biocontrol of Vegetable and Grain Crops Nematodes; Springer: Berlin/Heidelberg, Germany, 2008; pp. 51–76. [Google Scholar]
- Dos Santos, M.C.V.; Horta, J.; Moura, L.; Pires, D.V.; Conceicao, I.; Abrantes, I.; Costa, S.R. An integrative approach for the selection of Pochonia chlamydosporia isolates for biocontrol of potato cyst and root knot nematodes. Phytopathol. Mediterr. 2019, 58, 187–199. [Google Scholar]
- Zhang, J.; Fu, B.; Lin, Q.; Riley, I.T.; Ding, S.; Chen, L.; Cui, J.; Yang, L.; Li, H. Colonization of Beauveria bassiana 08F04 in root-zone soil and its biocontrol of cereal cyst nematode (Heterodera filipjevi). PLoS ONE 2020, 15, e0232770. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, T.; Antoniolli, Z.I.; Sordi, E.; Carvalho, I.R.; Bortoluzzi, E.C. Use of the Glomus etunicatum as biocontrol agent of the soybean cyst nematode. Res. Soc. Dev. 2021, 10, e7310615132. [Google Scholar] [CrossRef]
- Dijksterhuis, J.; Harder, W.; Wyss, U.; Veenhuis, M. Colonization and digestion of nematodes by the endoparasitic nematophagous fungus Drechmeria coniospora. Mycol. Res. 1991, 95, 873–878. [Google Scholar] [CrossRef]
- Tunlid, A.; Jansson, H.-B.; Nordbring-Hertz, B. Fungal attachment to nematodes. Mycol. Res. 1992, 96, 401–412. [Google Scholar] [CrossRef]
- Braga, F.R.; de Araújo, J.V. Nematophagous fungi for biological control of gastrointestinal nematodes in domestic animals. Appl. Microbiol. Biotechnol. 2014, 98, 71–82. [Google Scholar] [CrossRef]
- Nordbring-Hertz, B.; Jansson, H.; Tunlid, A. Encyclopedia of Life Sciences; Wiley: Chichester, UK, 2006. [Google Scholar]
- Wan, J.; Dai, Z.; Zhang, K.; Li, G.; Zhao, P. Pathogenicity and metabolites of endoparasitic nematophagous fungus Drechmeria coniospora YMF1.01759 against nematodes. Microorganisms 2021, 9, 1735. [Google Scholar] [CrossRef] [PubMed]
- Sarker, M.S.; Mohiuddin, K.; Al-Ani, L.K.T.; Hassan, M.N.; Akter, R.; Hossain, M.S.; Khand, M. Effect of bio-nematicide and bau-biofungicide against root-knot (Meloidogyne spp.) of soybean. Malays. J. Sustain. Agric. 2020, 4, 44. [Google Scholar] [CrossRef]
- Comans-Pérez, R.J.; Sánchez, J.E.; Al-Ani, L.K.T.; González-Cortázar, M.; Castañeda-Ramírez, G.S.; Mendoza-de Gives, P.; Sánchez-García, A.D.; Millán-Orozco, J.; Aguilar-Marcelino, L. Biological control of sheep nematode Haemonchus contortus using edible mushrooms. Biol. Control 2021, 152, 104420. [Google Scholar] [CrossRef]
- Girardi, N.S.; Sosa, A.L.; Etcheverry, M.G.; Passone, M.A. In vitro characterization bioassays of the nematophagous fungus Purpureocillium lilacinum: Evaluation on growth, extracellular enzymes, mycotoxins and survival in the surrounding agroecosystem of tomato. Fungal Biol. 2022, 126, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, S. Fungal secondary metabolites in biological control of crop pests. In Handbook of Industrial Mycology; CRC Press: Boca Raton, FL, USA, 2004; pp. 742–767. [Google Scholar]
- Westphal, A.; Becker, J. Components of soil suppressiveness against Heterodera schachtii. Soil Biol. Biochem. 2001, 33, 9–16. [Google Scholar] [CrossRef]
- Satou, T.; Kaneko, K.; Li, W.; Koike, K. The toxin produced by Pleurotus ostreatus reduces the head size of nematodes. Biol. Pharm. Bull. 2008, 31, 574–576. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Li, X.; Li, G.; Pan, Y.; Zhang, K. Acanthocytes of Stropharia rugosoannulata function as a nematode-attacking device. Appl. Environ. Microbiol. 2006, 72, 2982–2987. [Google Scholar] [CrossRef] [PubMed]
- Nordbring-Hertz, B.; Neumeister, H.; Sjollema, K.; Veenhuis, M. A conidial trap-forming mutant of Arthrobotrys oligospora. Mycol. Res. 1995, 99, 1395–1398. [Google Scholar] [CrossRef]
- Kwok, O.; Plattner, R.; Weisleder, D.; Wicklow, D. A nematicidal toxin from Pleurotus ostreatus NRRL 3526. J. Chem. Ecol. 1992, 18, 127–136. [Google Scholar] [CrossRef]
- Ekino, T.; Yoshiga, T.; Takeuchi-Kaneko, Y.; Kanzaki, N. Transmission electron microscopic observation of body cuticle structures of phoretic and parasitic stages of Parasitaphelenchinae nematodes. PLoS ONE 2017, 12, e0179465. [Google Scholar] [CrossRef]
- Liang, L.; Meng, Z.; Ye, F.; Yang, J.; Liu, S.; Sun, Y.; Guo, Y.; Mi, Q.; Huang, X.; Zou, C.; et al. The crystal structures of two cuticle-degrading proteases from nematophagous fungi and their contribution to infection against nematodes. FASEB J. 2010, 24, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Huang, X.; Tian, B.; Sun, H.; Duan, J.; Wu, W.; Zhang, K. Characterization of an extracellular serine protease gene from the nematophagous fungus Lecanicillium psalliotae. Biotechnol. Lett. 2005, 27, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Minglian, Z.; Minghe, M.; Keqin, Z. Characterization of a neutral serine protease and its full-length cDNA from the nematode-trapping fungus Arthrobotrys oligospora. Mycologia 2004, 96, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Junwei, W.; Qingling, M.; Jun, Q.; Weisheng, W.; Shuangqing, C.; Jianxun, L.; Chunguang, Z.; Chuangfu, C. The recombinant serine protease XAoz1 of Arthrobotrys oligospora exhibits potent nematicidal activity against Caenorhabditis elegans and Haemonchus contortus. FEMS Microbiol. Lett. 2013, 344, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, X.; Bai, N.; Yang, X.; Zhang, K.-Q.; Yang, J. Transcriptomic Analysis Reveals That Rho GTPases Regulate Trap Development and Lifestyle Transition of the Nematode-Trapping Fungus Arthrobotrys oligospora. Microbiol. Spectr. 2022, 10, e01759-21. [Google Scholar] [CrossRef] [PubMed]
- de Souza Gouveia, A.; de Freitas Soares, F.E.; Morgan, T.; Sufiate, B.L.; Tavares, G.P.; Braga, F.R.; Monteiro, T.S.A.; Geniêr, H.L.A.; de Freitas, L.G.; de Queiroz, J.H. Enhanced production of Monacrosporium thaumasium protease and destruction action on root-knot nematode Meloidogyne javanica eggs. Rhizosphere 2017, 3, 13–15. [Google Scholar] [CrossRef]
- de Freitas Soares, F.E.; de Queiróz, J.H.; Braga, F.R.; de Oliveira Tavela, A.; Araújo, J.M.; da Fonseca, L.A.; de Souza Gouveia, A.; de Araújo, J.V. Action of the nematophagous fungus Pochonia chlamydosporia on Dioctophyma renale eggs. Biocontrol Sci. Technol. 2014, 24, 399–406. [Google Scholar] [CrossRef]
- Soares, F.E.; Braga, F.R.; Araújo, J.V.; dos Santos Lima, W.; Mozer, L.R.; Queiróz, J.H. In vitro activity of a serine protease from Monacrosporium thaumasium fungus against first-stage larvae of Angiostrongylus vasorum. Parasitol. Res. 2012, 110, 2423–2427. [Google Scholar] [CrossRef]
- Khan, A.; Williams, K.L.; Nevalainen, H.K. Effects of Paecilomyces lilacinus protease and chitinase on the eggshell structures and hatching of Meloidogyne javanica juveniles. Biol. Control 2004, 31, 346–352. [Google Scholar] [CrossRef]
- Rodrigues, L.; Fortini, R.; CR Neves, M. Impacts of the use of biological pest control on the technical efficiency of the Brazilian agricultural sector. Int. J. Environ. Sci. Technol. 2023, 20, 1–16. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.; Kannangara, S.D.; Promputtha, I. Fungi vs. fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef] [PubMed]
- Moosavi, M.R.; Zare, R. Fungi as biological control agents of plant-parasitic nematodes. In Plant Defence: Biological Control; Springer: Berlin/Heidelberg, Germany, 2020; pp. 333–384. [Google Scholar]
- Nordbring-Hertz, B. Nematophagous fungi: Strategies for nematode exploitation and for survival. Microbiol. Sci. 1988, 5, 108–116. [Google Scholar] [PubMed]
- Silva, S.D.; Carneiro, R.M.; Faria, M.; Souza, D.A.; Monnerat, R.G.; Lopes, R.B. Evaluation of Pochonia chlamydosporia and Purpureocillium lilacinum for suppression of Meloidogyne enterolobii on tomato and banana. J. Nematol. 2017, 49, 77. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.-J.; Ariyawansa, H.A.; Becker, J.O.; Yang, J.-i. The evaluation of egg-parasitic fungi Paraboeremia taiwanensis and Samsoniella sp. for the biological control of Meloidogyne enterolobii on Chinese cabbage. Microorganisms 2020, 8, 828. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Vasconcellos, R.; Ramos, M.; Guimarães, M.; Yatsuda, A.; Vieira-Bressan, M. In vitro interaction of Brazilian strains of the nematode-trapping fungi Arthrobotrys spp. on Panagrellus sp. and Cooperia punctata. Memórias Do Inst. Oswaldo Cruz 2001, 96, 861–864. [Google Scholar] [CrossRef] [PubMed]
- Balbino, H.M.; de Souza Gouveia, A.; Monteiro, T.S.A.; Morgan, T.; de Freitas, L.G. Overview of the nematophagous fungus Duddingtonia flagrans. Biocontrol Sci. Technol. 2022, 32, 911–929. [Google Scholar] [CrossRef]
- Chen, S.; Liu, X. Control of the soybean cyst nematode by the fungi Hirsutella rhossiliensis and Hirsutella minnesotensis in greenhouse studies. Biol. Control 2005, 32, 208–219. [Google Scholar] [CrossRef]
- Khan, A.; Williams, K.L.; Nevalainen, H.K. Infection of plant-parasitic nematodes by Paecilomyces lilacinus and Monacrosporium lysipagum. BioControl 2006, 51, 659–678. [Google Scholar] [CrossRef]
- Manzanilla-Lopez, R.H.; Esteves, I.; Finetti-Sialer, M.M.; Hirsch, P.R.; Ward, E.; Devonshire, J.; Hidalgo-Díaz, L. Pochonia chlamydosporia: Advances and challenges to improve its performance as a biological control agent of sedentary endo-parasitic nematodes. J. Nematol. 2013, 45, 1. [Google Scholar]
- Bourne, J.M.; Kerry, B.; De Leij, F. The importance of the host plant on the interaction between root-knot nematodes Meloidogyne spp. and the nematophagous fungus, Verticillium chlamydosporium Goddard. Biocontrol Sci. Technol. 1996, 6, 539–548. [Google Scholar] [CrossRef]
- Bourne, J.; Kerry, B. Effect of the host plant on the efficacy of Verticillium chlamydosporium as a biological control agent of root-knot nematodes at different nematode densities and fungal application rates. Soil Biol. Biochem. 1998, 31, 75–84. [Google Scholar] [CrossRef]
- Pires, D.; Vicente, C.S.L.; Inácio, M.L.; Mota, M. The Potential of Esteya spp. for the Biocontrol of the Pinewood Nematode, Bursaphelenchus xylophilus. Microorganisms 2022, 10, 168. [Google Scholar] [CrossRef]
- Kerry, B. Biocontrol: Fungal parasites of female cyst nematodes. J. Nematol. 1980, 12, 253–259. [Google Scholar] [PubMed]
- Kerry, B.R.; Crump, D.H. Two fungi parasitic on females of cystnematodes (Heterodera spp.). Trans. Br. Mycol. Soc. 1980, 74, 119–125. [Google Scholar] [CrossRef]
- Barron, G.L.; Thorn, R.G. Destruction of nematodes by species of Pleurotus. Can. J. Bot. 1987, 65, 774–778. [Google Scholar] [CrossRef]
- Marlin, M.A.; Wolf, A.; Alomran, M.M.; Carta, L.K.; Newcombe, G. Nematophagous Pleurotus Species Consume Some Nematode Species but Are Themselves Consumed by Others. Forests 2019, 10, 404. [Google Scholar] [CrossRef]
- Khan, M.; Tanaka, K. Purpureocillium lilacinum for plant growth promotion and biocontrol against root-knot nematodes infecting eggplant. PLoS ONE 2023, 18, e0283550. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Gupta, A.; Dalela, M.; Sharma, S.; Sayyed, R.Z.; Enshasy, H.A.E.; Elsayed, E.A. Linking Organic Metabolites as Produced by Purpureocillium Lilacinum 6029 Cultured on Karanja Deoiled Cake Medium for the Sustainable Management of Root-Knot Nematodes. Sustainability 2020, 12, 8276. [Google Scholar] [CrossRef]
- Sharon, E.; Bar-Eyal, M.; Chet, I.; Herrera-Estrella, A.; Kleifeld, O.; Spiegel, Y. Biological control of the root-knot nematode Meloidogyne javanica by Trichoderma harzianum. Phytopathology 2001, 91, 687–693. [Google Scholar] [CrossRef]
- Sahebani, N.; Hadavi, N. Biological control of the root-knot nematode Meloidogyne javanica by Trichoderma harzianum. Soil Biol. Biochem. 2008, 40, 2016–2020. [Google Scholar] [CrossRef]
- Heidari, F.; Olia, M. Biological control of root-knot nematode, Meloidogyne javanica, using vermicompost and fungus Trichoderma harzianum on tomato. Iran. J. Plant Pathol. 2016, 52, 109–124. [Google Scholar]
- Lei, H.-M.; Wang, J.-T.; Hu, Q.-Y.; Li, C.-Q.; Mo, M.-H.; Zhang, K.-Q.; Li, G.-H.; Zhao, P.-J. 2-Furoic acid associated with the infection of nematodes by Dactylellina haptotyla and its biocontrol potential on plant root-knot nematodes. Microbiol. Spectr. 2023, 11, e01896-01823. [Google Scholar] [CrossRef]
- Shinya, R.; Aiuchi, D.; Kushida, A.; Tani, M.; Kuramochi, K.; Koike, M. Effects of fungal culture filtrates of Verticillium lecanii (Lecanicillium spp.) hybrid strains on Heterodera glycines eggs and juveniles. J. Invertebr. Pathol. 2008, 97, 291–297. [Google Scholar] [CrossRef]
- Hussain, M.; Zouhar, M.; Ryšánek, P. Suppression of Meloidogyne incognita by the entomopathogenic fungus Lecanicillium muscarium. Plant Dis. 2018, 102, 977–982. [Google Scholar] [CrossRef]
Figure 1.
Representative types of nematophagous fungi from different taxonomical groups and their infection structures [25].
Figure 1.
Representative types of nematophagous fungi from different taxonomical groups and their infection structures [25].

Figure 2.
Mode of action of nematophagous fungi.

Figure 4.
The nematode egg- and female-parasitic fungi and their infection modes.

Figure 5.
General representation of nematophagous fungi’s enzymes against cuticle, females, and eggs.
Figure 5.
General representation of nematophagous fungi’s enzymes against cuticle, females, and eggs.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Nematophagous fungi and their application as biocontrol agents.
| Nematophagous Fungi | Application as Biocontrol Agent | References |
|---|---|---|
| Arthrobotrys spp. | Prey free-living nematode (Panagrellus spp.) in vitro | [112] |
| Duddingtonia flagrans | Use for management of parasitic nematodes in domestic animals and plants | [113] |
| Hirsutella rhossiliensis | Effectively control soybean cyst nematode, Heterodera glycines | [114] |
| Paecilomyces lilacinus | Infects eggs, juveniles, and females of Meloidogyne javanica and also immature cysts of H. avenae and Radopholus similis | [115] |
| P. chlamydosporia | Control plant (semi-) endo-parasitic nematodes of the genera Globodera, Heterodera, Meloidogyne, Nacobbus, and Rotylenchulus | [116] |
| Verticillium chlamydosporium | Biological control agent for root-knot nematodes (Meloidogyne spp.) | [117,118] |
| Esteya spp. | Control pinewood nematode (PWN), Bursaphelenchus xylophilus causal agent of pine wilt disease (PWD) | [119] |
| Nematophthora gynophila | Reduces populations of the cereal cyst nematode, Heterodera spp. | [120,121] |
| Pleurotus spp. | Control nematodes such as, Poikilolaimus oxycercus, C. elegans, Pristionchus aerivorus, P. redivivus, and Acrobeloides amurensis | [122,123] |
| Purpureocillium lilacinum | Effective biocontrol agent against the root-knot nematode M. incognita | [124,125] |
| Trichodermaharzianum | Potentially control root-knot nematode M. javanica | [126,127,128] |
| Dactylellina haptotyla | Traps M. incognita by adhesive knobs and non-constricting loops in environment | [31,129] |
| Verticillium lecanii | Culture filtrates show toxicity against different stages of H. glycines | [130] |
| Lecanicillium muscarium | Effective against the root knot nematode M. incognita. | [131] |
| Lagenidiaceous | Attacks females of cyst-nematodes (Heterodera spp.) | [121] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rahman, M.U.; Chen, P.; Zhang, X.; Fan, B. Predacious Strategies of Nematophagous Fungi as Bio-Control Agents. Agronomy 2023, 13, 2685. https://doi.org/10.3390/agronomy13112685
AMA Style
Rahman MU, Chen P, Zhang X, Fan B. Predacious Strategies of Nematophagous Fungi as Bio-Control Agents. Agronomy. 2023; 13(11):2685. https://doi.org/10.3390/agronomy13112685
Chicago/Turabian StyleRahman, Mati Ur, Peng Chen, Xiuyu Zhang, and Ben Fan. 2023. "Predacious Strategies of Nematophagous Fungi as Bio-Control Agents" Agronomy 13, no. 11: 2685. https://doi.org/10.3390/agronomy13112685
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.