
Diversity and Characterization of Resistance to Pyraclostrobin in Colletotrichum spp. from Strawberry

1
College of Advanced Agricultural Sciences, Zhejiang A&F University, Lin’an, Hangzhou 311300, China
2
Research Institute for the Agriculture Science of Hangzhou, Hangzhou 310013, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agronomy 2023, 13(11), 2824; https://doi.org/10.3390/agronomy13112824
Submission received: 14 October 2023
/
Revised: 9 November 2023
/
Accepted: 14 November 2023
/
Published: 16 November 2023
(This article belongs to the Special Issue Plant Anthracnose: Etiology and Current Management Options)
Abstract
:Strawberry crown rot poses a significant menace to strawberry production during the seedling stage, and the main pathogen is Colletotrichum spp. Pyraclostrobin is one of the main fungicides that have been registered to control anthracnose caused by Colletotrichum spp. The diversity of pathogens and the risk of fungicide resistance may change from year to year. In order to explore the diversity of pathogens causing crown rot and evaluate the resistance risk of pathogens to pyraclostrobin in different years, crown rot samples were collected in Jiande, Zhejiang Province in 2019 and 2021, and the pathogens were identified. Based on the morphological identification and phylogenetic analysis based on ACT, CAL, CHS, GAPDH, and ITS, all 55 strains were identified as C. gloeosporioides species complexes, including 23 C. siamense isolates and 2 C. fructicola isolates in 2019, and all isolates were identified as C. siamense in 2021. C. siamense was the dominant pathogen of strawberry crown rot in 2019 and 2021. The resistance frequencies of the isolates collected in 2019 and 2021 to pyraclostrobin were 69.57% and 100%, respectively. In general, compared to that in 2019, the resistance frequencies of the pathogen to pyraclostrobin increased in 2021. In terms of fitness, there was no significant difference between resistant strains and sensitive strains in the mycelium growth rate, sporulation and spore germination rate. In addition, the resistant mutants exhibited positive cross-resistance to kresoxim-methyl and azoxystrobin. A sequential analysis of cytochrome b gene showed that C. siamense resistance to pyraclostrobin is linked to the G143A point mutation. Our study indicated that the risk of resistance a fungicide gradually increases with the increase in use years, and in order to reduce the emergence and spread of resistant populations, we should choose fungicides of different mechanisms of action for rotation to reduce the risk of resistance development.
1. Introduction
Strawberry is widely cultivated for its good economic benefits and high nutritional value [1]. With the increase in strawberry yield, fungal diseases have emerged as the main factor affecting the yield and quality of strawberry [2]. Strawberry crown rot is a major disease in most strawberry-growing areas, which can lead to serious economic losses. The crown is damaged, causing the above-ground part to be unable to absorb water and nutrients and eventual wilt [3]. Previous studies have shown that the primary pathogen of crown rot in Zhejiang Province is Colletotrichum siamense, followed by C. fructicola [4]. The diversity of this pathogen may change due to host immunity and environmental conditions [5]. It is not clear whether or not the dominant pathogens of crown rot have changed in recent years in strawberry-producing areas of Zhejiang Province. Colletotrichum spp. can be identified via their morphological characteristics such as colonies and spores, but these are easily affected by environmental factors [6]. Currently, numerous significant pathogens are identified via multilocus phylogenetic analysis combined with morphological characteristics [7].
To effectively manage Colletotrichum spp., we are required to consider various control measures. In terms of chemical control, quinone outside inhibitors (QoIs) are one of the fungicides often used to control Colletotrichum spp. [8,9]. The mechanism of action of QoI fungicides involves binding to the ubiquinol-oxidizing (Qo) site, disrupting electron transfer on cytochrome b (Cyt b) and inhibiting ATP production [10]. QoIs fungicides have broad-spectrum activity against fungal species. Pyraclostrobin, a prominent QoI fungicide, is utilized for controlling numerous fungal diseases, since with their widespread applications, resistance has emerged among several plant pathogens [11,12]. Resistance to QoI fungicides may occur because of point mutations in the Cyt b gene. It is reported that amino acid mutations at G143A, F129L and G137R confer varying degrees of QoI resistance in C. acutatum [13]. G143A mutations are the most common in resistant strains and confer high levels of resistance. The mutations F129L and G137R are linked to moderate levels of resistance [14,15]. Forcelini BB et al. collected strawberry samples from 1994 to 2014 to assess the resistance of C. acutatum to QoIs. The results showed that with the continuous use of fungicides, the resistance frequency of C. acutatum to QoI fungicides increased year by year [13]. However, studies have also shown that the frequency of resistance gradually decreases with the use of fungicides [16]. Pyraclostrobin is the main QoI fungicide used to control strawberry crown rot. The resistance level of strawberry crown rot pathogens may change with the continuous use of pyraclostrobin.
Resistant mutations may impose fitness costs in the absence of fungicide selection pressure. Bauske et al. assessed the inherent fitness of pathogen mutants by measuring parameters such as the mycelium growth rate, spore production, and spore germination rate. The results indicated no significant difference in mycelium growth rate and spore germination rate between resistant and sensitive strains [17]. Similar results were obtained in Magnaporthe grisea mutants [18]. The fitness of G143A mutants of M. grisea was not affected and their resistance remained steady. Veloukas et al. reported that the biological characteristics of Botrytis cinerea with the G143A mutation were similar [19]. In addition, mutants resistant to pyraclostrobin are generally cross-resistant to fungicides of the same mechanism of action [20,21]. However, little is known about the fitness, competitiveness and cross-resistance of strains of Colletotrichum spp. that are resistant to pyraclostrobin.
The objectives of this study were to (i) define the dominant pathogen of strawberry crown rot in different years, (ii) determine the resistance frequency of C. siamense to pyraclostrobin in different years, and (iii) assess the fitness of resistant and sensitive strains without fungicide selection pressure.
2. Materials and Methods
2.1. Isolation of Colletotrichum spp.
Strawberry crown rot samples with brown vascular bundles and plant wilting were collected from the same strawberry production area in Jiande, Hangzhou Province (27°02′ to 31°11′ N; 118°01′ to 123°10′ E), in 2019 and 2021. The pathogen was isolated from strawberry crown via the tissue separation method [22]. The soil on the surface of the diseased tissues was cleaned with double-distilled water (ddH2O), followed by being immersed for 30 s in 75% alcohol, being soaked for 2 min in 3% sodium hypochlorite, and being subsequently triple-rinsed with ddH2O. The cleaned tissues were cut into 5 × 5 mm pieces using a sterile scalpel and placed onto potato dextrose agar (PDA) medium containing 100 mg/L of streptomycin sulfate. Cultures were incubated for 2–4 days at 28 °C, and then we transferred the mycelia to a fresh PDA plate [23]. The single-spore isolates were conserved on PDA at 4 °C.
2.2. Pathogenicity Assay
Pathogenicity tests were performed to determine whether or not the isolates were pathogenic. Five mycelia pieces (5 mm) were excised from colony’s edge, transferred to a 100 mL potato dextrose (PD) medium and oscillated at 175 r/min and 28 °C to induce conidial production [24]. The concentration of spore suspension was adjusted to 106 conidia/mL. Six-week-old healthy strawberry plants were adopted for inoculation. An amount of 5 mL of the spore suspension was sprayed onto the strawberry crowns. The negative group was inoculated with ddH2O, with each isolate inoculating five plants. The plants were returned to a greenhouse at 28 °C.
2.3. Morphological Characterization
The isolates stored at a 4 °C were activated on a PDA plate incubating for 3 days, and then transferred onto a new PDA plate. After incubating for 7 days, the diameter of the colony was measured, the growth rate was calculated, and colony morphology was observed and recorded [25]; the experiment was conducted in three replications. The mycelium was scraped from the surface of the PDA culture medium using a glass slide and rinses with ddH2O. The mycelium was filtered through three layers of filter paper to obtain a spore suspension. The spore morphology was observed and the spore size was measured under a Scope A1 optical microscope (Nikon Eclipse 80i or Nikon SMZ25, Tokyo, Japan); fifty spores were measured for each isolate. An observation of appressorium was carried out in accordance with Emmett et al. [26], and the concentration of the spore suspension was set at 105 conidia/mL; the 10 μL spore suspension was added to the center of the hydrophobic membrane, and cultured at 28 °C for 14 h. The cover glass was pressed on the hydrophobic membrane, appressorium morphology was observed and the size was measured under the Scope A1 optical microscope (Nikon Eclipse 80i or Nikon SMZ25, Tokyo, Japan). Each isolate was measured, resulting in fifty appressoriums.
2.4. Molecular Identification and Phylogenetic Analysis
Genomic DNA from each isolate was extracted using a fungi genomic DNA rapid extraction kit (B518229-0100; Sangon Biotech, Shanghai, China). The calmodulin (CAL), glyceraldehyde 3-phosphate dehydrogenase (GAPDH), chitin synthase (CHS), actin (ACT) and internal transcribed spacer (ITS) genes were amplified using the primer pairs CL1C/CL2C [27], GDF/GDR [28], CHS-79F/CHS-354R [29], ACT-512F/ACT-783R [30] and ITS-1F/ITS4 [31]. Phylogenetic analysis was carried out following previously described methods [32]. The gene numbers of standard strains utilized in this study are provided in Supplemental Table S1. All sequences were edited using MEGA 7.0 [33]. The clipped sequences were combined in Sequence Martix 1.8. Modeltest 3.7.win, Mrmodeltest2.3 and Win paup4b10-console, as implemented in MrMTgui v1.0, and were used to estimate the best nucleotide substitution model [34]. Bayesian inference (BI) phylogenies were constructed using Mr. Bayes v.3.1.2 [35]. Five simultaneous Markov chains were run for 300,000 generations each. Phylogenetic trees were visualized using Figtree v1.4.4 [36].
2.5. Resistance Detection
The molecular identification results showed that C. siamense was the dominant strain of crown rot, so the resistance level of 53 C. siamense to pyraclostrobin was measured using the differential measurement method. All isolates were cultured on PDA medium, and then transferred onto a new PDA plate. The mycelia plugs were prepared from the edge of the colony and selected and inoculated on the PDA medium plate containing different concentrations of pyraclostrobin. Using 0 μg/mL and 10 μg/mL as the differential concentrations [37], each concentration was inoculated with three PDA plates. After culture for 3 days at 28 °C, sensitive strains were defined as those able to grow at 0 μg/mL and unable to grow at 10 μg/mL, resistant strains were defined as those able to grow at 10 μg/mL. The frequency of resistance was calculated in accordance with the following formula [37]:
Resistance frequency (%) = (number of resistant strains/numbers of total strains) × 100
2.6. Sensitivity Level Determination
Five pyraclostrobin-resistant and five pyraclostrobin-sensitive strains were selected to determine the sensitivity of C. siamense to pyraclostrobin. The tested strains stored in freeze storage tubes were inoculated on the PDA medium and then transferred onto a new PDA plate. Mycelial plugs with a diameter of 5 mm were removed from the edge of the colony and inoculated onto PDA plates containing different concentrations of pyraclostrobin. Sensitive strains were inoculated in PDA medium containing 0.5, 1, 2, 4 and 8 μg/mL of pyraclostrobin, and resistant strains were inoculated in PDA medium containing 2, 4, 8, 16 and 32 μg/mL of pyraclostrobin. The PDA plate without pyraclostrobin was used as the control, and three PDA plates were measured for each concentration. After culture at 28 °C for 7 days, the cross-crossing method was used to measure the colony diameter, which was was measured horizontally and vertically, and calculated the inhibition rate of mycelia [38]. The virulence regression equation, effective inhibitory medium concentration (EC50 value) and correlation coefficient were obtained by regressing the percent relative growth against the log10 values of the fungicide concentrations using SPSS 22.0 software.
2.7. Fitness Determination of Sensitive and Resistant Strains
In terms of mycelial growth, the strains stored at 4 °C were inoculated on a PDA plate, cultured at 28 °C for 3 days and transferred onto a new PDA plate. After incubation for 7 days, the colony diameter of each strains was measured in two approximately perpendicular directions; the mycelia growth rate was calculated. Three plates were measured for each strain.
In terms of sporulation, five mycelia plugs with a diameter of 5 mm were collected from the edge of the colony, transferred onto a 100 mL liquid potato dextrose (PD) medium. and shaken at 175 rpm and 28 °C to induce conidia production [24]. After 5 days, the mycelium was filtered with three layers of filter paper to obtain the spore suspension. Briefly, 10 µL of the spore suspension was pipetted onto a hemocytometer and spore production was counted under an A1 microscope. Three fields were observed for each strain.
In terms of the spore germination rate, the spore suspension concentration was adjusted to 105 conidia/mL. Briefly, 20 μL of the spore suspension was taken and applied on water agar medium, cultured at 28 °C. A spore was considered germinated if the germ tube had reached at least half the length of the conidium. When the spore germination rate of the control group reached more than 90%, the data were counted [25]. The number of germinated spores was observed under the Scope A1 optical microscope (Nikon Eclipse 80i or Nikon SMZ25, Tokyo, Japan). We observed 5 fields for each strain, with more than 100 spores in each field.
2.8. Cross-Resistance
Kresoxim-methyl and azoxystrobin was used to assay the cross-resistance to pyraclostrobin [37]. Mycelia plugs with a diameter of 5 mm were taken from the edge of the colony and inoculated on the PDA plates with kresoxim-methyl and azoxystrobin in different concentrations. Sensitive strains were inoculated in PDA medium containing 0.5, 1, 2, 4 and 8 μg/mL of kresoxim-methyl/azoxystrobin, and resistant strains were inoculated in PDA medium containing 2, 4, 8, 16 and 32 μg/mL of kresoxim-methyl/azoxystrobin. After incubation for 7 days at 28 °C, the diameters of the colonies were measured via the cross-crossing method and inhibition rates were calculated to compute the EC50 values. Each concentration was inoculated with three PDA plates.
2.9. Amplification and Sequencing of CsCyt b
Genomic DNA from both resistant and sensitive strains was extracted using the CTAB methods with minor modifications. The mycelium was scraped into 1.5 mL tube, quartz sand and 300 μL of CTAB extract were added, and the mixture was ground for 120 s at 85 Hz. Then, 400 μL CTAB extract was added and the mixture was shaken for 15 s. An amount of 700 μL of trichloromethane was added; the mixture was shaken for 15 s and centrifuged at 12,000 rpm for 10 min. Next, 500 μL of the supernatant was removed and transferred into a new 1.5 mL tube; 500 mL of isopropyl alcohol was added, and the mixture was centrifuged at 12,000 rpm for 10 min. The supernatant was discarded; to it was added 500 mL of 70% ethanol, and then it was centrifuged at 12,000 rpm for 5 min. Finally, the supernatant was discarded, After dryed, 50 μL of ddH2O was added, and the mixture was stored at 4 °C. According to the CsCyt b gene sequence retrieved from NCBI, PCR primers (F: 5′-TCTGCTTTCTTCTTCTTAGTTTA-3′ and R: 5′GGGATAGCACTTATAAGGTTAGT-3′) were designed to amplify the fragment containing the complete coding region of CsCyt b. The PCR reaction system was 50 µL, the upstream and downstream primers were 2 µL each, the template was 1 µL, the 2× mix TaqDNA Mix was 25 µL, and ddH2O was added to 50 µL. The reaction procedure was as follows: 95 °C for 3 min, followed by 35 cycles of 95 °C for 15 s, 56 °C for 15 s, 72 °C for 2 min, and finally 72 °C for 10 min. The PCR products were verified on 1.0% agarose gel at 254 nm (UV) and further sequenced by Tsingke Biotechnology Co., Ltd. (Beijing, China). Using Snapgene v5.2.4, the sequence was aligned and analyzed. The base sequence was compared and translated into amino acids after removing the introns, and the mutation of amino acids was analyzed.
3. Results
3.1. Isolation and Identification of Colletotrichum spp.
In 2019 and 2021, in total, 25 and 30 isolates of Colletotrichum spp. were isolated, respectively. In accordance with Koch’s postulates, all isolates were demonstrated to be pathogenic. According to the morphological and cultural characterizations, all 55 isolates belonged to C. siamense and C. fructicola, including 23 C. siamense isolates and 2 C. fructicola isolates in 2019, while all isolates were identified as C. siamense in 2021. C. siamense colonies are grayish white on the front side and grayish on the back side. C. fructicola colonies have grayish white edges and a celadon center (Figure 1). No significant differences were observed in terms of mycelium growth rate and sporulation between the two Colletotrichum species, and the morphology and size of the spores and appressorium were similar (Table 1). ACT, CAL, CHS, GADPH, and ITS datasets were utilized for constructing a phylogenetic tree. All identified isolates were categorized within the C. gloeosporioides species complex, with 53 isolates belonging to C. siamense and 2 isolates belonging to C. fructicola (Figure 2). Supplementary Table S1 provides the GenBank numbers for all isolates.
3.2. Frequency of Resistance of C. siamense to Pyraclostrobin
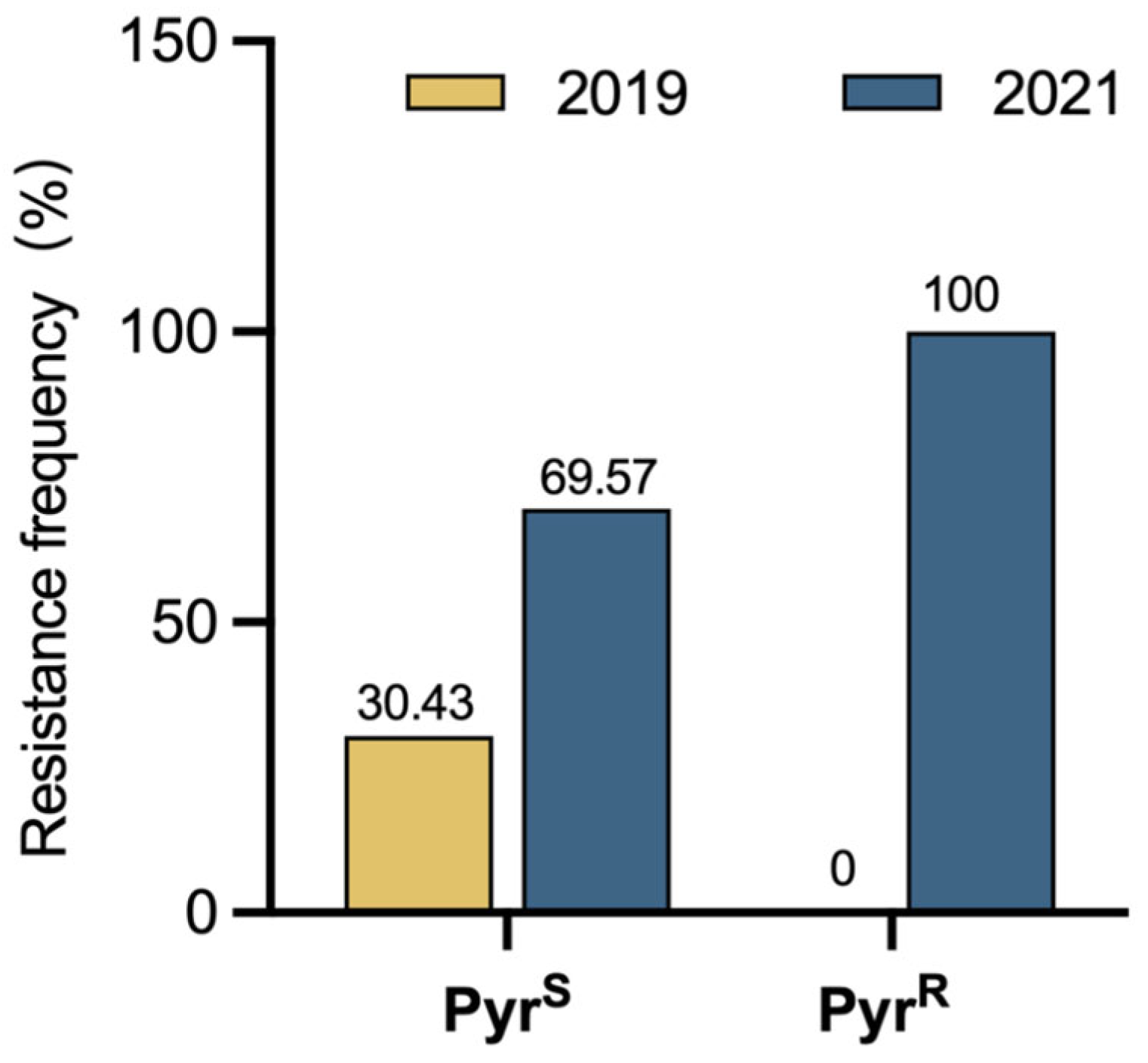
The primary pathogen responsible for strawberry crown rot was C. siamense. Therefore, the resistance level of C. siamense to pyraclostrobin was assessed. Briefly, all 46 strains could grow on the PDA plates containing 10 μg/mL of pyraclostrobin, and were considered resistant strains; the resistance frequency was 86.79%. In 2019, 7 of the 23 C. siamense strains could not grow on the PDA plates containing 10 μg/mL of pyraclostrobin, and the other 16 strains grew on the PDA plates containing 10 μg/mL of pyraclostrobin and were resistant strains; the resistance frequency was 69.57% (Figure 3). In 2021, 30 strains were grown on PDA plates containing 10 μg/mL of pyraclostrobin with a resistance frequency of 100% (Figure 3).
3.3. Sensitivity of C. siamense to Pyraclostrobin
To confirm the sensitivity of strains resistant and sensitive to pyraclostrobin, we calculated their EC50 values of resistant and sensitive strains. The EC50 values for sensitive strains varied from 1.192 to 2.068 μg/mL, and the average EC50 values were 1.586 μg/mL (Table 2). The EC50 values of resistant strains varied from 18.159 to 23.797 μg/mL, with an average EC50 value of 21.158 μg/mL. The mean resistance level was 13.3.
3.4. Fitness Comparison between Resistant and Sensitive Strains
The mycelial growth rate, sporulation quantity and spore germination rate of the resistant and sensitive strains were measured. The results indicated no significant difference in the fitness between the resistant and sensitive strains (Figure 4). The growth rate of the resistant and sensitive strains was (11.16 ± 0.09) mm/d and (11.27 ± 0.06) mm/d, respectively. The sporulation quantities were (1.46 ± 0.11) × 106 conidia/mL and (1.34 ± 0.14) × 106 conidia/mL, respectively. The respective germination rates were (82.15 ± 1.19)% and (85.54 ± 2.31)%, respectively.
3.5. Cross-Resistance of Resistant Mutants
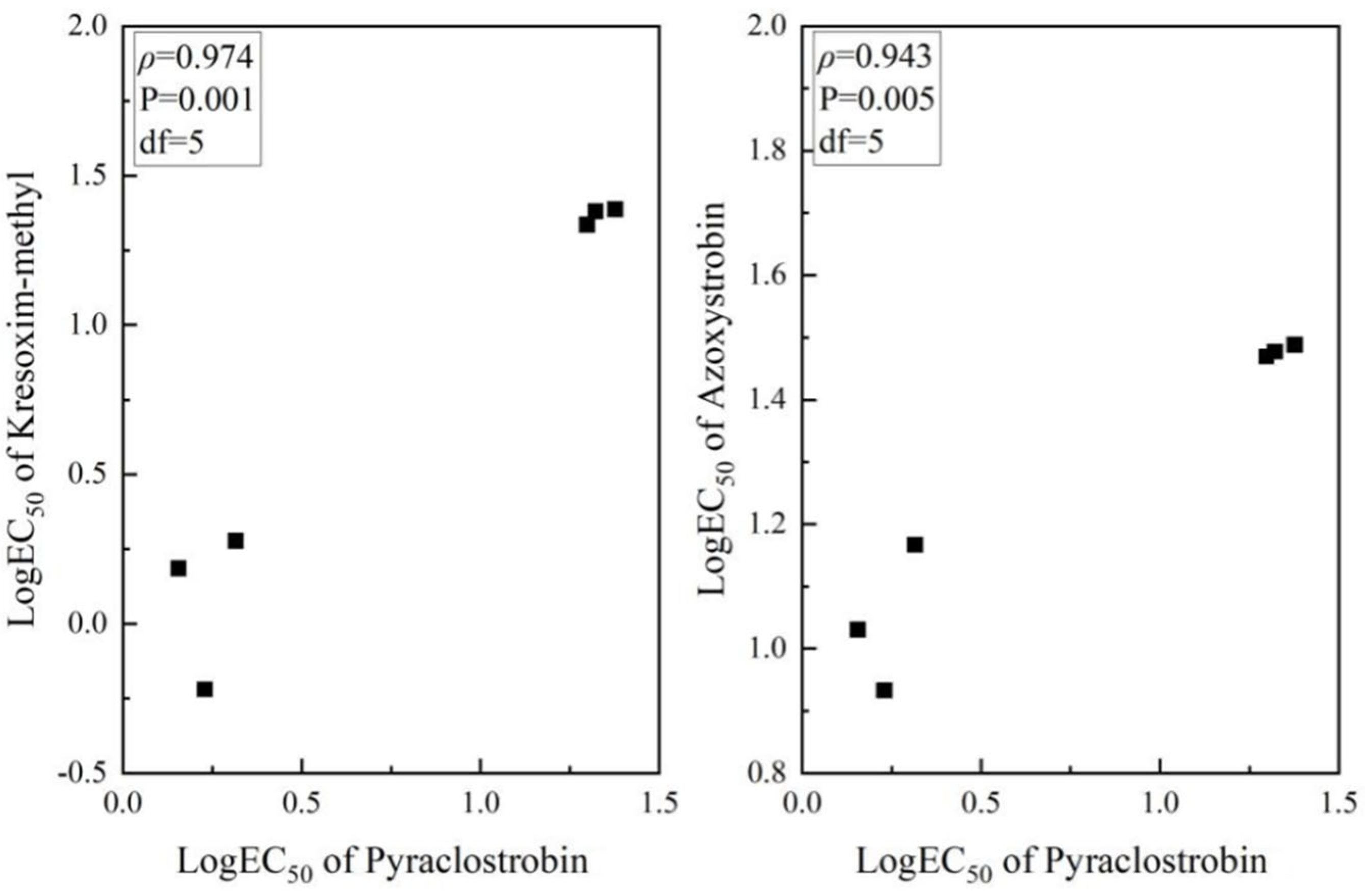
Assessing cross-resistance is crucial for choosing fungicides to control plant pathogens that have developed resistance to specific fungicides. A total of 6 strains were tested for cross-resistance. Consequently, we examined the cross-resistance of pyraclostrobin-resistant strains to kresoxim-methyl and azoxystrobin. The findings demonstrated a significant positive cross-resistance between pyraclostrobin and kresoxim-methyl, azoxystrobin (ρ > 0.8) (Figure 5).
3.6. Sequence Analysis of CsCyt b
In order to analyze whether or not the cytb gene of the pyraclostrobin-resistant strain had been mutated, we used primers LT-CytbF3 and LT-CytbR3 to amplify the Cyt b gene fragment of the pyraclostrobin-sensitive and -resistant strains, and a PCR product with a length of 2118 bp was obtained. The amino acid sequence was compared using Snapgene, and the amino acid at the 143rd position of the resistant strain had changed from G (Gly) to Ala (A) (Figure 6).
4. Discussion
Through morphological identification and phylogenetic analysis, the pathogens of strawberry crown rot in 2019 and 2021 were identified as C. gloeosporioides species complexes. Furthermore, 23 C. siamense and 2 C. fructicola species were isolated in 2019, and all 30 isolates were C. siamense in 2021. C. siamense was the advantage pathogen of crown rot in 2019 and 2021, aligning with findings from previous studies. C. siamense was still the dominant pathogen of crown rot in different years [39]. There was no significant difference between C. siamense and C. fructicola in the mycelium growth rate, sporulation quantity, and spore and appressorium size. Previous studies have shown that the appressoria and spores of C. fructicola were larger [39] than those in this study, probably due to the small sample size in this study, which was not representative.
The application of fungicides is a critical measure with which to control anthracnose disease [9]. QoI fungicides are extensively employed to control crown rot induced by Colletotrichum spp., due to their broad spectrum [40]. However, the excessive use of the same fungicide can lead to selection pressure and the emergence of resistance [41]. C. siamense is the predominant pathogen responsible for strawberry crown rot in Zhejiang Province; therefore, we studied the resistance risk of C. siamense to pyraclostrobin in different years. In general, the resistance frequency of C. siamense to pyraclostrobin was 86.79%, of which the resistance frequency in 2019 was 69. 57%. The resistance frequency reached 100% in 2021; the resistance frequency of C. siamense to pyrazinamide increased. It is indicated that C. siamense has developed extensive resistance to pyraclostrobin.
Mutations in the Cyt b gene have been associated with resistance for QoI fungicides. Resistance to QoIs is usually caused by F129L, G137R and G143A substitution in mitochondrial Cyt b genes [13]. The 143rd amino acid mutation of Cyt b causes a high level of resistance to pyraclostrobin [35]. F129L has also been reported in C. cereale, Magnaporthe grisea and other pathogenic fungi [42]. In our study, we amplified and sequenced the Cyt b gene fragments from both sensitive and resistant strains. Only the point mutation of G143A (from glycine to alanine) was detected in resistant strains, suggesting high levels of resistance. F129L and G137R loci mutations were not found in our study. All C. siamense-resistant isolates showed a high level of resistance to pyraclostrobin in the greenhouses.
Resistance to QoI fungicides linked to G143A mutations can affect the structure of the Qo site, and this may reduce the activity of the cytochrome bc1 complex, thereby differentially affecting the adaptability of pathogens [43,44]. No notable distinctions were observed in the growth rate, sporulation rate and spore germination rate between resistant and sensitive strains for C. siamense from strawberry. These results indicated that a high risk of further spread of resistant isolates in the fields. Previous studies had shown adaptive penalties for QoI-resistant isolates in C. gloeosporioides [45], while no significant differences were found between QoI resistance and sensitivity isolates in the C. acutatum species complex [46]. These outcomes highlighted that different Colletotrichum species had different phenotypes for adaptive punishment in resistant isolates. This further indicated that the species of pathogen should be clearly defined in the prevention and control of disease.
It is easy for cross-resistance to occur between fungicides with the same mechanisms of action. Cross-resistance analysis is of great significance for the rational selection of fungicides and the control of plant pathogens resistant to one or more fungicides. We further analyzed the cross-resistance between pyraclostrobin and kresoxim-methyl and azoxystrobin, and found that there was cross-resistance between these three types of QoI fungicides. This is also consistent with previous reports [47]. This study showed that C. siamense developed widespread high-level resistance to pyraclostrobin and that there was no significant difference in biological characteristics between pyraclostrobin-resistant and sensitive strains for C. siamense. It may spreaded widely in the field. In general, C. siamense had a high risk of resistance to pyraclostrobin. Fungicides with different action modes should be replaced to control strawberry crown rot. The results of this study had important guiding significance for the formulation of specific anthracnose control strategies.
5. Conclusions
C. siamense remains the primary pathogen of strawberry crown rot in Zhejiang Province. Strains resitsant to pyraclostrobin have been found in the field and they exhibit high-level resistance. Biological characteristics do not significantly differ between pyraclostrobin-resistant and -sensitive strains. Furthermore, pyraclostrobin exhibits positive cross-resistance along with two other QoI fungicides, indicating that such fungicides should not be further employed for the management of strawberry crown rot.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/agronomy13112824/s1. Table S1: Colletotrichum spp. used in multi-gene analysis in this study.
Author Contributions
Conceptualization, H.Y. and C.Z.; methodology, Y.L., H.Y. and C.Z.; software, S.H. and W.X.; validation, H.Y. and C.Z.; formal analysis, Y.L., W.X. and S.H.; investigation, S.Z. and S.H.; writing—original draft preparation, S.Z. and S.H.; writing—review and editing, H.Y. and C.Z.; visualization, S.Z. and S.H.; supervision, H.Y. and C.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Agriculture and Social Development Research Project of Hangzhou (202203A07), and the Joint-Extension Project of important Agriculture Technology in Zhejiang Province (2021XTTGSC02-4).
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barbey, C.R.; Lee, S.; Verma, S.; Bird, K.A.; Yocca, A.E.; Edger, P.P.; Knapp, S.J.; Whitaker, V.M.; Folta, K.M. Disease resistance genetics and genomics in octoploid strawberry. G3 Genes Genom. Genet. 2019, 9, 3315–3332. [Google Scholar] [CrossRef]
- Farzaneh, M.; Kiani, H.; Sharifi, R.; Reisi, M.; Hadian, J. Chemical composition and antifungal effects of three species of Satureja (S. hortensis, S. spicigera, and S. khuzistanica) essential oils on the main pathogens of strawberry fruit. Postharvest Biol. Technol. 2015, 109, 145–151. [Google Scholar] [CrossRef]
- Paynter, M.; Gomez, A.; Ko, L.; Herringtonet, M.E. Research into crown rot and wilt diseases of strawberries in Queensland. Acta Hortic. 2016, 1117, 163–170. [Google Scholar] [CrossRef]
- Chen, X.Y.; Dai, D.J.; Zha, S.F.; Shen, Y.; Wang, H.D.; Zhang, C.Q. Genetic diversity of Colletotrichum spp. causing strawberry anthracnose in Zhejiang, China. Plant Dis. 2020, 104, 1351–1357. [Google Scholar] [CrossRef] [PubMed]
- Cobey, S. Pathogen evolution and the immunological niche. Ann. N. Y. Acad. Sci. 2014, 1320, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum–current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef]
- Liu, F.; Ma, Z.Y.; Hou, L.W.; Diao, Y.Z.; Wu, W.P.; Damm, U.; Song, S.; Cai, L. Updating species diversity of Colletotrichum, with a phylogenomic overview. Stud. Mycol. 2022, 101, 1–56. [Google Scholar] [CrossRef]
- Haack, S.E.; Ivors, K.L.; Holmes, G.J.; Förster, H.; Adaskaveg, J.E. Natamycin, a new bio-fungicide for managing crown rot of strawberry caused by QoI-Resistant Colletotrichum acutatum. Plant Dis. 2018, 102, 1687–1695. [Google Scholar] [CrossRef]
- Han, Y.C.; Zeng, X.G.; Xiang, F.Y.; Zhang, Q.H.; Guo, C.; Chen, F.Y.; Gu, Y.C. Carbendazim sensitivity in populations of Colletotrichum gloeosporioides complex infecting strawberry and yams in Hubei Province of China. J. Integr. Agric. 2018, 17, 1391–1400. [Google Scholar] [CrossRef]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; ParrDobrzanski, B. The strobilurin fungicides. Pest Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef]
- Fernández-Ortuño, D.; Torés, J.A.; De Vicente, A.; Pérez-García, A. Mechanisms of resistance to QoI fungicides in phytopathogenic fungi. Int. Microbiol. 2008, 11, 1–9. [Google Scholar] [PubMed]
- Wei, L.; Zheng, H.; Zhang, P.; Chen, W.; Zheng, J.; Chen, C.; Cao, A. Molecular and biochemical characterization of Colletotrichum gloeosporioides isolates resistant to azoxystrobin from grape in China. Plant Pathol. 2021, 70, 1300–1309. [Google Scholar] [CrossRef]
- Forcelini, B.B.; Seijo, T.E.; Amiri, A.; Peres, N.A. Resistance in strawberry isolates of Colletotrichum acutatum from Florida to quinone outside inhibitor fungicides. Plant Dis. 2016, 100, 2050–2056. [Google Scholar] [CrossRef]
- Gisi, U.; Sierotzki, H.; Cook, A.; McCaffery, A. Mechanisms influencing the evolution of resistance to Qo inhibitor fungicides. Pest Manag. Sci. 2002, 58, 859–867. [Google Scholar] [CrossRef]
- Sierotzki, H.; Frey, R.; Wullschleger, J.; Palmero, S.; Karlin, S.; Godwin, J.; Gisi, U. Cytochrome b gene sequence and structure of Pyrenophora teres and P. tritici-repentis and implications for QoI resistance. Pest Manag. Sci. 2007, 63, 225–233. [Google Scholar] [CrossRef]
- Runno-Paurson, E.; Nassar, H.; Tähtjärv, T.; Eremeev, V.; Hansen, M.; Niinemets, Ü. High temporal variability in late blight pathogen diversity, virulence, and fungicide resistance in potato breeding fields: Results from a long-term monitoring study. Plants 2022, 11, 2426. [Google Scholar] [CrossRef]
- Bauske, M.J.; Gudmestad, N.C. Parasitic fitness of fungicide-resistant and -sensitive isolates of Alternaria solani. Plant Dis. 2018, 102, 666–673. [Google Scholar] [CrossRef]
- Avila-Adame, C.; Köller, W. Characterization of spontaneous mutants of Magnaporthe grisea expressing stable resistance to the Qo-inhibiting fungicide azoxystrobin. Curr. Genet. 2003, 42, 332–338. [Google Scholar] [CrossRef]
- Veloukas, T.; Kalogeropoulou, P.; Markoglou, A.N.; Karaoglanidis, G.S. Fitness and competitive ability of Botrytis cinerea field isolates with dual resistance to SDHI and QoI fungicides, associated with several sdhB and the cytb G143A mutations. Phytopathology 2014, 104, 347–356. [Google Scholar] [CrossRef]
- Oiki, S.; Yaguchi, T.; Urayama, S.; Hagiwara, D. Wide distribution resistance to the fungicides fludioxonil and iprodione in Penicillium species. PLoS ONE 2022, 17, e0262521. [Google Scholar] [CrossRef]
- Liu, S.M.; Hai, F.; Jiang, J. Sensitivity to fludioxonil of Botrytis cinerea isolates from tomato in Henan province of China and characterizations of fludioxonil-resistant mutants. J. Phytopathol. 2017, 165, 98–104. [Google Scholar] [CrossRef]
- Phoulivong, S.; Cai, L.; Chen, H.; Mckenzie, E.H.C.; Abdelsalam, K.; Chukeatirote, E. Colletotrichum gloeosporioides is not a common pathogen on tropical fruits. Fungal Divers. 2020, 44, 33–43. [Google Scholar] [CrossRef]
- Noman, E.; Al-Gheethi, A.A.; Rahman, N.K.; Talip, B.; Mohamed, R.; Kadir, O.A. Single spore isolation as a simple and efficient technique to obtain fungal pure culture. Earth Environ. Sci. 2018, 140, 10255. [Google Scholar] [CrossRef]
- Emmett, R.W.; Parbery, D.G. Appressoria. Annu. Rev. Phytopathol. 1975, 13, 147–165. [Google Scholar] [CrossRef]
- O’Donnell, K.; Nirenberg, H.I.; Aoki, T.; Cigelnik, E. A multigene phylogeny of the Gibberella fujikuroi species complex: Detection of additional phylogenetically distinct species. Mycoscience 2000, 41, 61–78. [Google Scholar] [CrossRef]
- Templeton, M.D.; Rikkerink, E.H.; Solon, S.L.; Crowhurst, R.N. Cloning and molecular characterization of the glyceraldehyde-3-phosphate dehydrogenase-encoding gene and cDNA from the plant pathogenic fungus Glomerella cingulata. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef]
- Damm, U.; Woudenberg, J.H.C.; Cannon, P.F.; Crous, P.W. Colletotrichum species with curved conidia from herbaceous hosts. Fungal Divers. 2009, 39, 45. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA 5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Vaidya, G.; Lohman, D.J.; Meier, R. Sequence matrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Page, R.D. Tree view: An application to display phylogenetic trees on personal computers. Bioinformatics 1996, 12, 357–358. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Hu, X.R.; Zhang, C.Q. Molecular detection of QoI resistance in Colletotrichum gloeosporioides causing strawberry anthracnose based on loop-mediated isothermal amplification assay. Plant Dis. 2019, 103, 1319–1325. [Google Scholar] [CrossRef] [PubMed]
- Usman, H.M.; Tan, Q.; Karim, M.M.; Adnan, M.; Yin, W.X.; Zhu, F.X.; Luo, C.X. Sensitivity of Colletotrichum fructicola and Colletotrichum siamense of peach in China to multiple classes of fungicides and characterization of pyraclostrobin-resistant isolates. Plant Dis. 2021, 105, 3459–3465. [Google Scholar] [CrossRef]
- Gale, L.R.; Bryant, J.D.; Calvo, S.; Giese, H.; Katan, T.; O’Donnell, K. Chromosome complement of the fungal plant pathogen Fusarium graminearum based on genetic and physical mapping and cytological observations. Genetics 2005, 171, 985–1001. [Google Scholar] [CrossRef]
- Cai, L.; Hyde, K.D.; Taylor, P.W.J.; Weir, B.; Waller, J.; Abang, M.M.; Zhang, J.Z.; Yang, Y.L.; Phoulivong, S.; Liu, Z.Y.; et al. A polyphasic approach for studying Colletotrichum. Fungal Divers. 2009, 39, 183–204. [Google Scholar]
- Hu, S.D.; Wu, J.Y.; Yang, X.Q.; Yu, H.; Zhang, C.Q. Involvement of CYP51A and CYP51B in growth, reproduction, pathogenicity, and sensitivity to fungicides in Colletotrichum siamense. Agronomy 2023, 13, 239. [Google Scholar] [CrossRef]
- Piccirillo, G.; Carrieri, R.; Polizzi, G.; Azzaro, A.; Lahoz, E.; Fernández-Ortuño, D.; Vitale, A. In vitro and in vivo activity of QoI fungicides against Colletotrichum gloeosporioides causing fruit anthracnose in Citrus sinensis. Sci. Hortic. 2018, 236, 90–95. [Google Scholar] [CrossRef]
- Russell, P.E. 1995. Fungicide resistance: Occurrence and management. J. Agric. Sci. 1995, 124, 317–323. [Google Scholar] [CrossRef]
- Young, J.R.; Tomaso-Peterson, M.; Tredway, L.P.; De La Cerda, K. Occurrence and molecular identification of azoxystrobin-resistant Colletotrichum cereale isolates from golf course putting greens in the southern united states. Plant Dis. 2010, 94, 751–757. [Google Scholar] [CrossRef]
- Fisher, N.; Brown, A.C.; Sexton, G.; Cook, A.; Windass, J.; Meunier, B. Modeling the Qo site of crop pathogens in Saccharomyces cerevisiae cytochrome b. Eur. J. Biochem. 2004, 271, 2264–2271. [Google Scholar] [CrossRef] [PubMed]
- Fisher, N.; Meunier, B. Molecular basis of resistance to cytochrome bc1 inhibitors. FEMS Yeast Res. 2008, 8, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Diao, Y.; Wang, W.; Hao, J.; Imran, M.; Duan, H.; Liu, X.L. Assessing the risk for resistance and elucidating the genetics of Colletotrichum truncatum that is only sensitive to some DMI fungicides. Front. Microbiol. 2017, 8, 1779. [Google Scholar] [CrossRef]
- Forcelini, B.B.; Rebello, C.S.; Wang, N.Y.; Peres, N.A. Fitness, competitive ability, and mutation stability of isolates of Colletotrichum acutatum from strawberry resistant to QoI fungicides. Phytopathology 2018, 108, 462–468. [Google Scholar] [CrossRef]
- Duan, Y.B.; Xin, W.J.; Lu, F.; Li, T.; Li, M.X.; Wu, J.; Wang, J.X.; Zhou, M.G. Benzimidazole-and QoI-resistance in Corynespora cassiicola populations from greenhouse-cultivated cucumber: An emerging problem in China. Pestic. Biochem. Phys. 2019, 153, 95–105. [Google Scholar] [CrossRef]
Figure 1.
Colony morphology of of Colletotrichum siamense and C. fructicola on PDA (a1,a2,b1,b2), conidia (a3,b3), and appressoria (a4,b4); scale bar = 20 µm in (a3,b3,a4,b4).
Figure 1.
Colony morphology of of Colletotrichum siamense and C. fructicola on PDA (a1,a2,b1,b2), conidia (a3,b3), and appressoria (a4,b4); scale bar = 20 µm in (a3,b3,a4,b4).

Figure 2.
The Colletotrichum gloeosporioides species complex was determined as the identity of the phylogenetic tree using Bayesian inference. The sequencing analysis included the ACT, CAL, CHS, GAPDH, and ITS genes DNA sequence. The scale bar shows 0.08 expected changes per site.
Figure 2.
The Colletotrichum gloeosporioides species complex was determined as the identity of the phylogenetic tree using Bayesian inference. The sequencing analysis included the ACT, CAL, CHS, GAPDH, and ITS genes DNA sequence. The scale bar shows 0.08 expected changes per site.

Figure 3.
Frequency of resistance to pyraclostrobin in Colletotrichum siamensis in 2019 and 2021. PyrR represents pyraclostrobin-resistant strain, and PyrS represents pyraclostrobin-sensitive strain.
Figure 3.
Frequency of resistance to pyraclostrobin in Colletotrichum siamensis in 2019 and 2021. PyrR represents pyraclostrobin-resistant strain, and PyrS represents pyraclostrobin-sensitive strain.

Figure 4.
Comparison of mycelium growth rate (a), sporulation quantity (b) and spore germination rate (c) between pyraclostrobin-resistant and -sensitive strains of Colletotrichum siamense. PyrR represents the pyraclostrobin-resistant strain, PyrS represents the pyraclostrobin-sensitive strain, and “ns” means no significant difference.
Figure 4.
Comparison of mycelium growth rate (a), sporulation quantity (b) and spore germination rate (c) between pyraclostrobin-resistant and -sensitive strains of Colletotrichum siamense. PyrR represents the pyraclostrobin-resistant strain, PyrS represents the pyraclostrobin-sensitive strain, and “ns” means no significant difference.

Figure 5.
Cross−resistance between pyraclostrobin, and kresoxim−methyl and azoxystrobin. p < 0.05 indicates there is a significant difference. ρ > 0.8 indicates that there is cross-resistance. The black squares represent different strains.
Figure 5.
Cross−resistance between pyraclostrobin, and kresoxim−methyl and azoxystrobin. p < 0.05 indicates there is a significant difference. ρ > 0.8 indicates that there is cross-resistance. The black squares represent different strains.

Figure 6.
Comparison of Cyt b gene sequences between pyraclostrobin-resistant and -sensitive strains of Colletotrichum siamense. JD-HX-A-12-1, ZS-ZJ-111-1, JD-ZS-11, JD-ZJ-14-2, and JD-HY-20 are pyraclostrobin-sensitive strains; H121, H211, H63, H116, and H85 are pyraclostrobin-resistant strains.
Figure 6.
Comparison of Cyt b gene sequences between pyraclostrobin-resistant and -sensitive strains of Colletotrichum siamense. JD-HX-A-12-1, ZS-ZJ-111-1, JD-ZS-11, JD-ZJ-14-2, and JD-HY-20 are pyraclostrobin-sensitive strains; H121, H211, H63, H116, and H85 are pyraclostrobin-resistant strains.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Size of spore, appresoria, hyphae growth rates, and sporulation of Colletotrichum siamense and C. fructicola.
Table 1.
Size of spore, appresoria, hyphae growth rates, and sporulation of Colletotrichum siamense and C. fructicola.
| Species | Strain Number | Conidia y | Appressoria y | Growth Rate (mm/Day) y | Sporulation (×106) y | ||
|---|---|---|---|---|---|---|---|
| Length (μm) | Width (μm) | Length (μm) | Width (μm) | ||||
| C. siamense | JD-HY-20 | 15.26 ± 0.80 a | 5.55 ± 0.22 a | 6.58 ± 0.33 a | 5.92 ± 0.57 a | 10.91 ± 0.11 a | 15.25 ± 2.80 a |
| H35 | 15.60 ± 0.36 a | 5.83 ± 0.19 a | 6.79 ± 0.15 a | 6.13 ± 0.36 a | 11.51 ± 0.65 a | 15.6 ± 3.36 a | |
| C. fructicola | JD-ZJ-5 | 16.83 ± 0.14 a | 5.86 ± 0.14 a | 6.19 ± 0.15 a | 6.59 ± 0.32 a | 12.58 ± 0.22 a | 16.06 ± 4.14 a |
| JD-ZJ-9 | 16.64 ± 0.23 a | 6.74 ± 0.15 a | 6.70 ± 0.42 a | 6.37 ± 0.23 a | 12.67 ± 0.12 a | 16.83 ± 3.23 a | |
y Data are the mean ± standard error. The same letters represent the data being not statistically different (p > 0.05) according to the least significant different (LSD) test.
Table 2.
The EC50 values of strains resistant and sensitive to pyraclostrobin.
| Isolate | Origin | EC50 (μg/mL) |
|---|---|---|
| JD-HX-A-12-1 (PyrS) | Crown of strawberry | 1.544 |
| ZS-ZJ-111-1 (PyrS) | Crown of strawberry | 1.695 |
| JD-ZJ-11 (PyrS) | Crown of strawberry | 1.192 |
| JD-ZJ-14-2 (PyrS) | Crown of strawberry | 1.431 |
| JD-HY-20 (PyrS) | Crown of strawberry | 2.068 |
| H121 (PyrR) | Crown of strawberry | 20.974 |
| H211 (PyrR) | Crown of strawberry | 23.797 |
| H63 (PyrR) | Crown of strawberry | 18.159 |
| H116 (PyrR) | Crown of strawberry | 21.740 |
| H85 (PyrR) | Crown of strawberry | 21.120 |
Note: PyrR represents pyraclostrobin-resistant strain, and PyrS represents pyraclostrobin-sensitive strain.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hu, S.; Zhang, S.; Xiao, W.; Liu, Y.; Yu, H.; Zhang, C. Diversity and Characterization of Resistance to Pyraclostrobin in Colletotrichum spp. from Strawberry. Agronomy 2023, 13, 2824. https://doi.org/10.3390/agronomy13112824
AMA Style
Hu S, Zhang S, Xiao W, Liu Y, Yu H, Zhang C. Diversity and Characterization of Resistance to Pyraclostrobin in Colletotrichum spp. from Strawberry. Agronomy. 2023; 13(11):2824. https://doi.org/10.3390/agronomy13112824
Chicago/Turabian StyleHu, Shuodan, Shuhan Zhang, Wenfei Xiao, Yahui Liu, Hong Yu, and Chuanqing Zhang. 2023. "Diversity and Characterization of Resistance to Pyraclostrobin in Colletotrichum spp. from Strawberry" Agronomy 13, no. 11: 2824. https://doi.org/10.3390/agronomy13112824
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.