
The Fungus Metarhizium sp. BCC 4849 Is an Effective and Safe Mycoinsecticide for the Management of Spider Mites and Other Insect Pests

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Strains
2.2. Insect-Rearing Culture
2.3. Microscopy of the Infection Process
2.4. Insect Bioassays
2.5. Greenhouse Trials and Application Treatment
2.6. Determination of Conidiation in Different Substrates
2.7. Conidial Powder Formulation and Shelf-Life Assay
2.8. Toxicological Testing in Animal Models
2.9. De Novo Genome Assembly of Metarhizium sp. BCC 4849 and Gene Annotation
2.10. Statistical Analyses
3. Results
3.1. Virulence of Metarhizium sp. Strains against T. truncatus and Four Other Pests
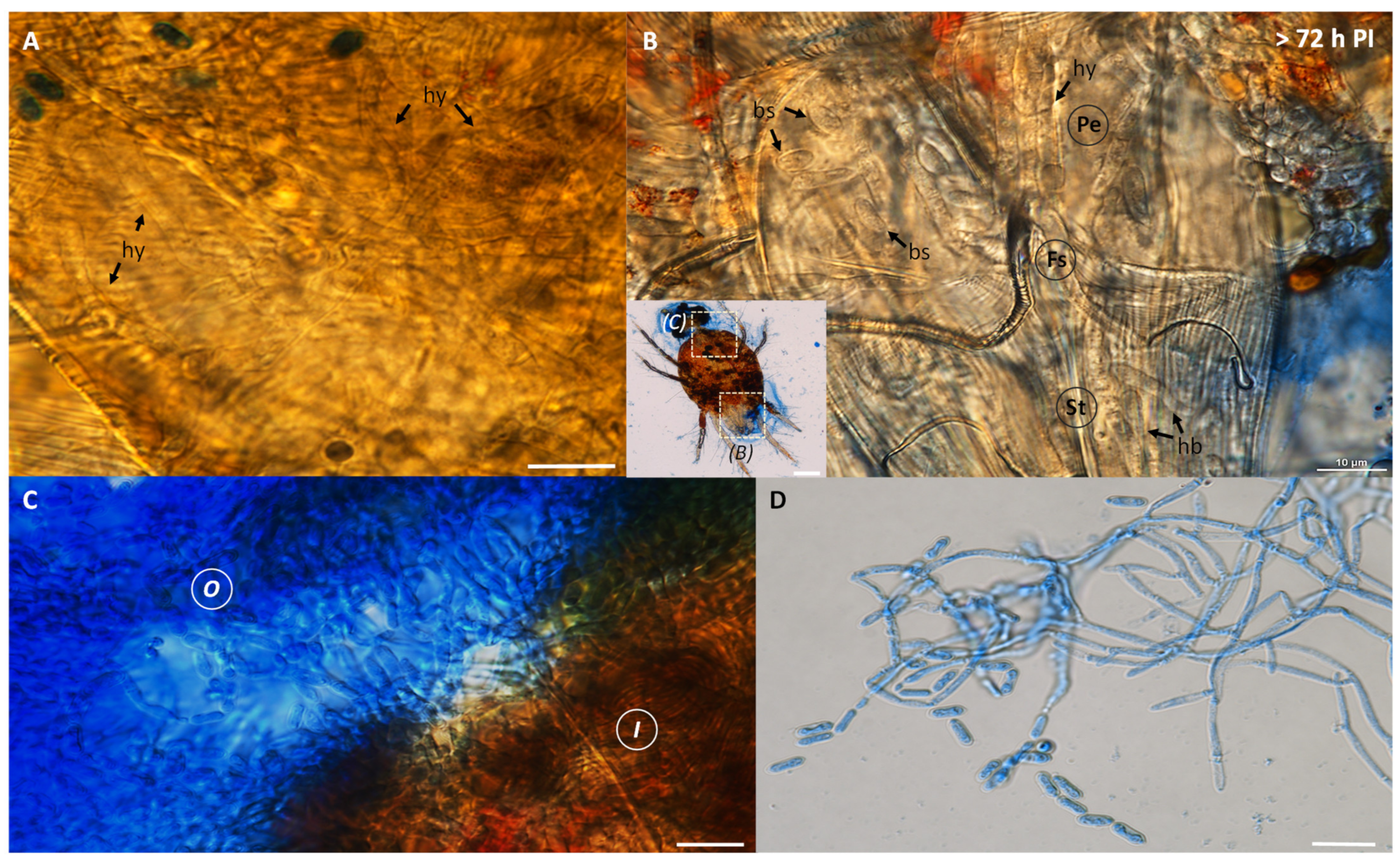
3.2. Metarhizium sp. BCC 4849 Infection of the Cassava Mites
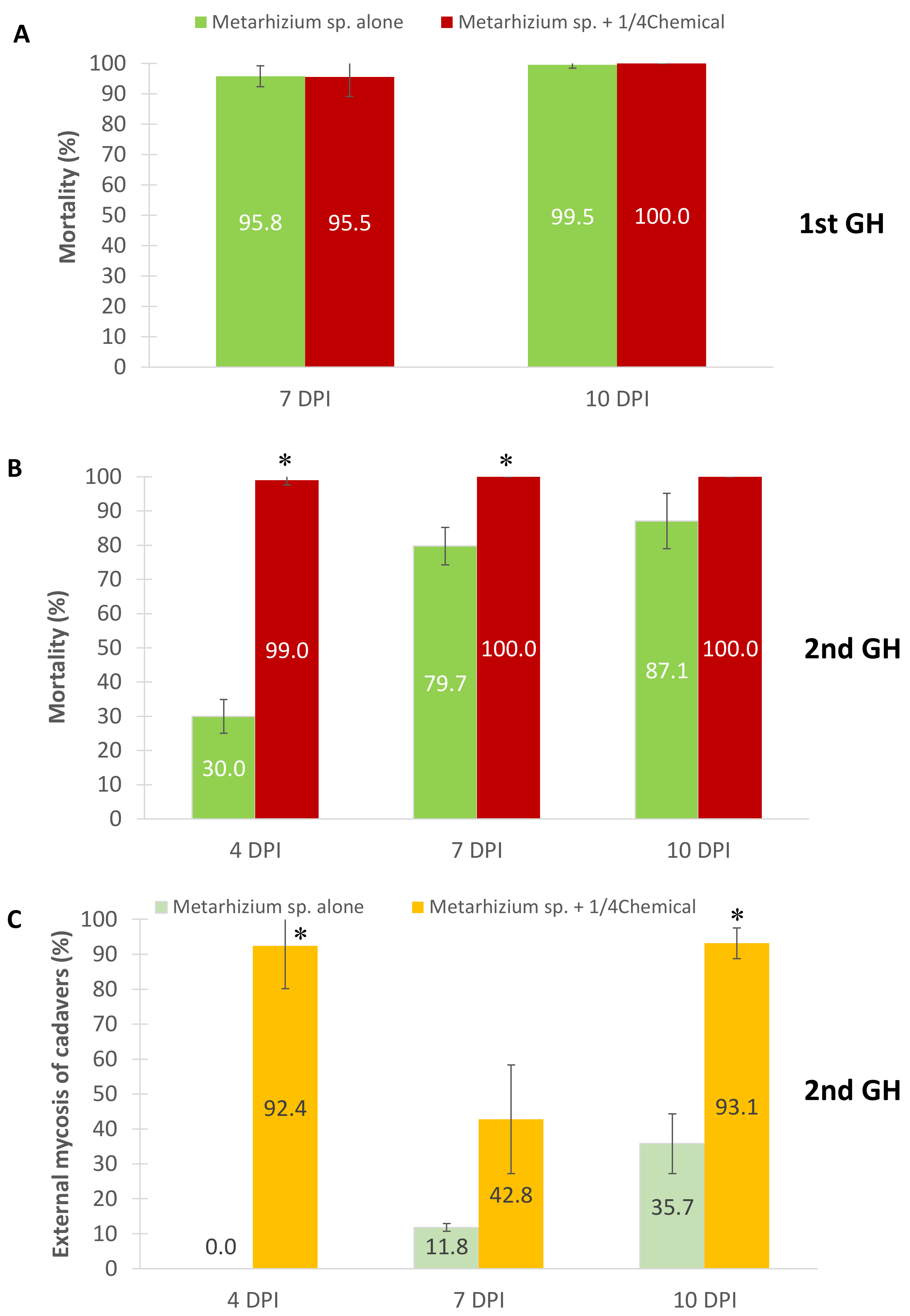
3.3. Efficacy of Metarhizium sp. and Diluted Acaricide Applications on E. africanus Mortality in Greenhouse
3.4. Genome Sequencing and Assembly of Metarhizium sp. BCC 4849
3.5. Effect of Subculture Media and Temperature on Sporulation
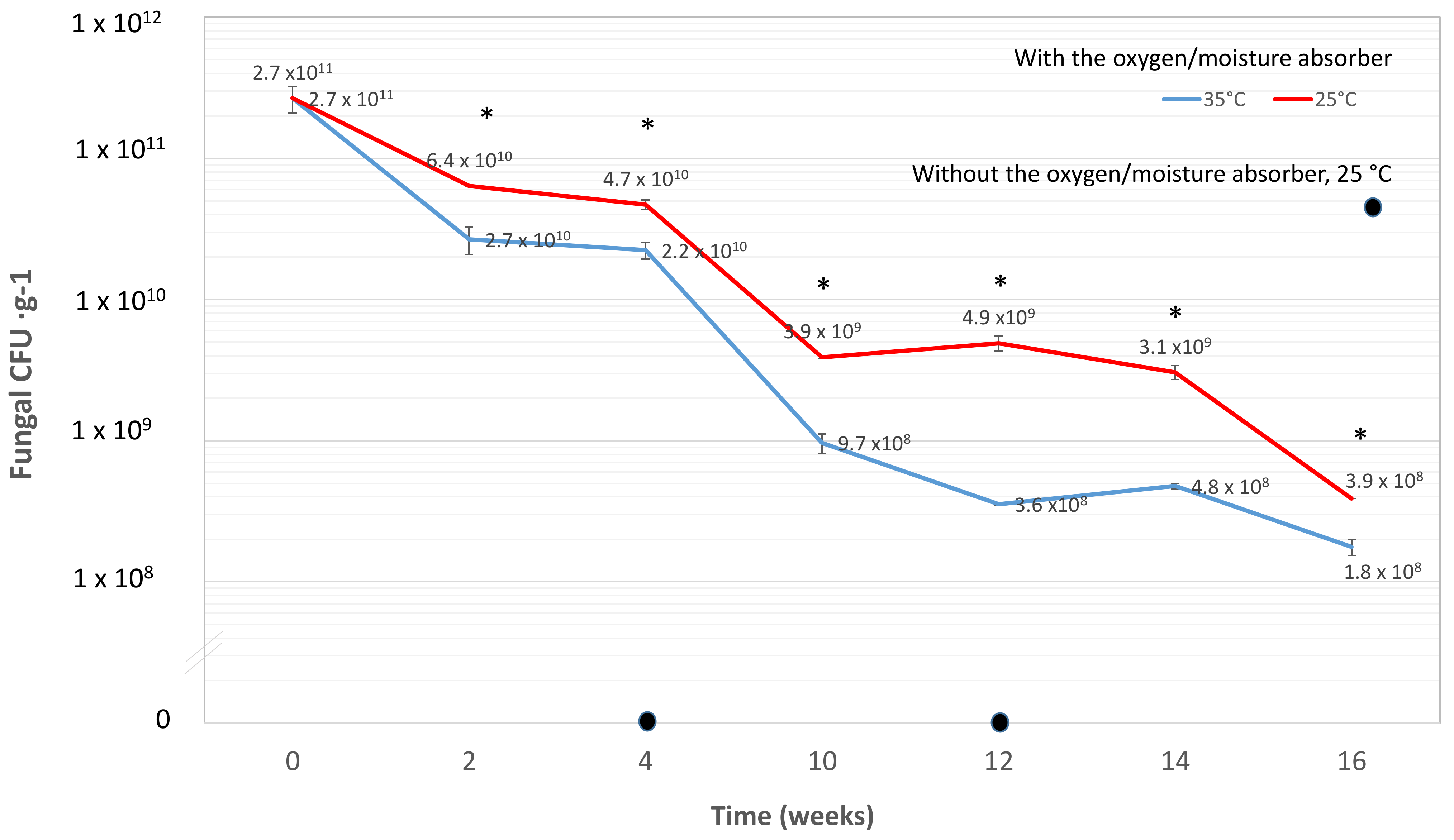
3.6. Effect of Moisture-Oxygen Absorber Sachets for Shelf-Life Extension
3.7. Toxicological Testing in Animal Models
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faria, M.R.D.; Wraight, S.P. Mycoinsecticides and Mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. BioControl 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Ment, D.; Gindin, G.; Rot, A.; Soroker, V.; Glazer, I.; Barel, S.; Samish, M. Novel technique for quantifying adhesion of Metarhizium anisopliae conidia to the tick cuticle. Appl. Environ. Microbiol. 2010, 76, 3521–3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasulo, T.R.; Denmark, H.A. Two spotted spider mite, Tetranychus urticae Koch. (Arachnida: Acari: Tetranychidae). EDIS 2003, 15, 1–5. [Google Scholar] [CrossRef]
- Strickler, J. Growing potential, hemp could be game changer for American farmers. Acres USA 2018, 48, 22–26. [Google Scholar]
- Palaniswami, M.S.P.; Pillai, K.S.; Nair, R.R.; Mohandas, C. A new cassava pest in India. Cassava Newslett. 1995, 9, 6–7. [Google Scholar]
- Bernardo, E.; Esquevia, E. Seasonal abundance of the cassava red spider mite, Tetranychus kanzawai Kishida, and its predators on some cassava accessions. Ann. Trop. Res. 1981, 3, 197–203. [Google Scholar]
- El-Sharabasy, H.M. Laboratory evaluation of the effect of the entomopathogenic fungi, Hirsutella thompsonii and Paecilomyces fumosoroseus against the citrus brown mite Eutetranychus orientalis (Acari: Tetranychidae). Plant Protect. Sci. 2016, 51, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Shternshis, M.; Andreeva, I.; Trandysheva, M. Effect of host plant on control of Tetranychus urticae by Verticillium (Lecanicillium) lecanii. IOBC/wprs Bull. 2005, 28, 37–40. [Google Scholar]
- Wekesa, V.W.; Maniania, N.K.; Knapp, M.; Boga, H.I. Pathogenicity of Beauveria bassiana and Metarhizium anisopliae to the tobacco spider mite Tetranychus evansi. Exp. Appl. Acarol. 2005, 36, 41–50. [Google Scholar] [CrossRef]
- Bugeme, D.M.; Maniania, N.K.; Knapp, M.; Boga, H.I. Effect of temperature on virulence of Beauveria bassiana and Metarhizium anisopliae isolates to Tetranychus evansi. Exp. Appl. Acarol. 2008, 46, 275–285. [Google Scholar] [CrossRef]
- Lord, J.C. From Metchnikoff to Monsanto and beyond: The path of microbial control. J. Invertebr. Pathol. 2005, 89, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Iwanicki, N.S.A.; Pereira, A.A.; Botelho, A.; Rezende, J.M.; Moral, R.A.; Zucchi, M.I.; Delalibera Junior, I. Monitoring of the field application of Metarhizium anisopliae in Brazil revealed high molecular diversity of Metarhizium spp in insects, soil and sugarcane roots. Sci. Rep. 2019, 9, 4443. [Google Scholar] [CrossRef] [Green Version]
- Aw, K.M.S.; Hue, S.M. Mode of infection of Metarhizium spp. fungus and their potential as biological control agents. J. Fungi 2017, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Leao, M.P.; Tiago, P.V.; Andreote, F.D.; de Araujo, W.L.; de Oliveira, N.T. Differential expression of the pr1A gene in Metarhizium anisopliae and Metarhizium acridum across different culture conditions and during pathogenesis. Genet. Mol. Biol. 2015, 38, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: New insights into host-pathogen interactions. Adv. Genet. 2016, 94, 307–364. [Google Scholar] [CrossRef]
- Gabarty, A.; Salem, H.M.; Fouda, M.A.; Abas, A.A.; Ibrahim, A.A. Pathogencity induced by the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in Agrotis ipsilon (Hufn.). J. Radiat. Res. Appl. Sci. 2019, 7, 95–100. [Google Scholar] [CrossRef]
- St Leger, R.J.; Wang, J.B. Metarhizium: Jack of all trades, master of many. Open Biol. 2020, 10, 200307. [Google Scholar] [CrossRef]
- Houndété, T.A.; Kétoh, G.K.; Hema, O.S.; Brévault, T.; Glitho, I.A.; Martin, T. Insecticide resistance in field populations of Bemisia tabaci (Hemiptera: Aleyrodidae) in West Africa. Pest Manag. Sci. 2010, 66, 1181–1185. [Google Scholar] [CrossRef]
- Song, L.; Li, W.; Guo, H.; Wang, S.; Shen, H. Inheritance of resistance to abamectin+ pyridaben in Tetranychus truncatus Ehara. Chin. J. Entomol. 2019, 56, 150–154. [Google Scholar] [CrossRef]
- Jaramillo, J.; Borgemeister, C.; Ebssa, L.; Gaigl, A.; Tobón, R.; Zimmermann, G. Effect of combined applications of Metarhizium anisopliae (Metsch.) Sorokin (Deuteromycotina: Hyphomycetes) strain CIAT 224 and different dosages of imidacloprid on the subterranean burrower bug Cyrtomenus bergi Froeschner (Hemiptera: Cydnidae). Biol. Control 2005, 34, 12–20. [Google Scholar] [CrossRef]
- Paula, A.R.; Carolino, A.T.; Paula, C.O.; Samuels, R.I. The combination of the entomopathogenic fungus Metarhizium anisopliae with the insecticide imidacloprid increases virulence against the dengue vector Aedes aegypti (Diptera: Culicidae). Parasit Vectors 2011, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Bostein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 9–25. [Google Scholar] [CrossRef] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Xiao, G.; Zheng, P.; Shang, Y.; Su, Y.; Zhang, X.; Liu, X.; Zhan, S.; Leger, R.J.S.; Wang, C. Trajectory and genomic determinants of fungal-pathogen speciation and host adaptation. PNAS 2014, 111, 16796–16801. [Google Scholar] [CrossRef] [Green Version]
- Amnuaykanjanasin, A.; Jirakkakul, J.; Panyasiri, C.; Panyarakkit, P.; Nounurai, P.; Chantasingh, D.; Eurwilaichitr, L.; Cheevadhanarak, S.; Tanticharoen, M. Infection and colonization of tissues of the aphid Myzus persicae and cassava mealybug Phenacoccus manihoti by the fungus Beauveria bassiana. Biol. Control 2012, 58, 379–391. [Google Scholar] [CrossRef]
- St Leger, R.J.; Roberts, D.W.; Staples, R.C. A Model to explain differentiation of appressoria by germlings of Metarhizium anisopliae. J. Invertebr. Pathol. 1991, 57, 299–310. [Google Scholar] [CrossRef]
- Tseng, M.N.; Chung, C.L.; Tzean, S.S. Mechanisms relevant to the enhanced virulence of a dihydroxynaphthalene-melanin metabolically engineered entomopathogen. PLoS ONE 2014, 9, e90473. [Google Scholar] [CrossRef]
- Jeffs, L.B.; Khachatourians, G.G. Estimation of spore hydrophobicity for members of the genera Beauveria, Metarhizium, and Tolypocladium by salt-mediated aggregation and sedimentation. Can. J. Microbiol. 1997, 43, 23–28. [Google Scholar] [CrossRef]
- Udompaisarn, S.; Toopaang, W.; Sae-Ueng, U.; Srisuksam, C.; Wichienchote, N.; Wasuwan, R.; Nahar, N.A.S.; Tanticharoen, M.; Amnuaykanjanasin, A. The polyketide synthase PKS15 has a crucial role in cell wall formation in Beauveria bassiana. Sci. Rep. 2020, 10, 12630. [Google Scholar] [CrossRef] [PubMed]
- St Leger, R.J.; Cooper, R.M.; Charnley, A.K. The Effect of melanization of Manduca sexta cuticle on growth and infection by Metarhizium anisopliae. J. Invertebr. Pathol. 1988, 52, 459–470. [Google Scholar] [CrossRef]
- Charnley, A.K.; Leger, S.T. The Role of cuticle-degrading enzymes in fungal pathogenesis in insects. In The Fungal Spore and Disease Initiation in Plants and Animals; Cole, G.T., Hoch, H.C., Eds.; Springer: Boston, MA, USA, 1991; pp. 267–286. [Google Scholar]
- Moino, A., Jr.; Alves, S.B.; Lopes, R.B.; Neves, P.M.O.J.; Pereira, R.M.; Vieira, S.A. External development of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in the subterranean termite Heterotermes tenuis. Sci. Agric. 2002, 59, 267–273. [Google Scholar] [CrossRef]
- Elhakim, E.; Mohamed, O.; Elazouni, I. Virulence and proteolytic activity of entomopathogenic fungi against the two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae). Egypt. J. Biol. Pest Control 2020, 30, 30. [Google Scholar] [CrossRef] [Green Version]
- Pelizza, S.A.; Schalamuk, S.; Simon, M.R.; Stenglein, S.A.; Pacheco-Marino, S.G.; Scorsetti, A.C. Compatibility of chemical insecticides and entomopathogenic fungi for control of soybean defoliating pest, Rachiplusia nu. Rev. Argent. Microbiol. 2018, 50, 189–201. [Google Scholar] [CrossRef]
- Alves, R.T.; Bateman, R.P.; Gunn, J.; Prior, C.; Leather, S.R. Effects of different formulations on viability and medium-term storage of Metarhizium anisopliae conidia. Neotrop. Entomol. 2002, 31, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Moore, D.; Douro-Kpindou, O.K.; Jenkins, N.E.; Lomer, C.J. Effects of moisture content and temperature on storage of Metarhizium flavoviride conidia. Biocontrol Sci. Technol. 2010, 6, 51–62. [Google Scholar] [CrossRef]
- Bagwan, N.B. Mass multiplication and shelf-life of Metarhizium anisopliae and Beauveria bassiana in solid and liquid formulations. Int. J. Plant Prot. 2011, 4, 34–37. [Google Scholar]
- Mascarin, G.M.; Jackson, M.A.; Behle, R.W.; Kobori, N.N.; Junior, I.D. Improved shelf life of dried Beauveria bassiana blastospores using convective drying and active packaging processes. Appl. Microbiol. Biotechnol. 2016, 100, 8359–8370. [Google Scholar] [CrossRef]
- Faria, M.; Hotchkiss, J.H.; Wraight, S.P. Application of modified atmosphere packaging (gas flushing and active packaging) for extending the shelf life of Beauveria bassiana conidia at high temperatures. Biol. Control. 2012, 61, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Day, J. Practical application of the revolutionary preservation (RP) system for marcasite. In Proceedings of the 14th Triennial Meeting–ICOM Committee for Conservation, The Hague, The Netherlands, 12–16 September 2005. [Google Scholar]
- Zaki, O.; Weekers, F.; Thonart, P.; Tesch, E.; Kuenemann, P.; Jacques, P. Limiting factors of mycopesticide development. Biol. Control 2020, 144, 104220. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Insect Mortality (%) ± s.e.m. |
|---|---|
| BCC 4849 | 82.4 ± 0.1 a |
| BCC 16,000 | 7.9 ± 5.4 c |
| BCC 30,455 | 32.8 ± 18.1 b,c |
| TBRC 4912 | 37.3 ± 16.4 b,c |
| TBRC 4908 | 44.9 ± 20.4 b |
| Control (non-treated) | 11.2 ± 8.6 b,c |
| Target Pest | Inoculation Technique | DPI | Insect Mortality (%) ± s.e.m. |
|---|---|---|---|
| Cassava mite (Tetranychus truncatus) | Topical application | 5 | 82.4 ± 0.1 |
| Bean aphid (Aphis craccivora) | Topical application | 3 | 96.0 ± 5.5 |
| Cassava mealybug (Phenacoccus manihoti) | Topical application | 3 | 92.0 ± 5.7 |
| Sweet potato weevil (Cylas formicarius) | Dipping | 5 | 91.9 ± 7.3 |
| Oriental fruit fly (Bactrocera dorsalis) | Mixing with dry conidia | 6 | 98.9 ± 1.9 |
| Study Type | Study Guideline | Animal | Dose Range/Level | GHS 3 Toxic Classification |
|---|---|---|---|---|
| Acute oral toxicity | OECD GLP 1 423 | Rats | LD50 > 2000 mg/kg | Class 5 (Unclassified) |
| Acute dermal toxicity | OECD GLP 402 | Rats | LD50 > 2000 mg/kg | Class 5 (Unclassified) |
| Acute dermal irritation/Corrosion | OECD GLP 404 | Rabbits | - | Class 5 (Unclassified) |
| Acute eye irritation/Corrosion | OECD GLP 405 | Rabbits | - | Category 2B (mild irritant) |
| Acute pulmonary toxicity/Pathogenecity | U.S. EPA OCSPP 2 885.3150 | Rats | No mortality at 1.05 × 108 CFU |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wasuwan, R.; Phosrithong, N.; Promdonkoy, B.; Sangsrakru, D.; Sonthirod, C.; Tangphatsornruang, S.; Likhitrattanapisal, S.; Ingsriswang, S.; Srisuksam, C.; Klamchao, K.; et al. The Fungus Metarhizium sp. BCC 4849 Is an Effective and Safe Mycoinsecticide for the Management of Spider Mites and Other Insect Pests. Insects 2022, 13, 42. https://doi.org/10.3390/insects13010042
Wasuwan R, Phosrithong N, Promdonkoy B, Sangsrakru D, Sonthirod C, Tangphatsornruang S, Likhitrattanapisal S, Ingsriswang S, Srisuksam C, Klamchao K, et al. The Fungus Metarhizium sp. BCC 4849 Is an Effective and Safe Mycoinsecticide for the Management of Spider Mites and Other Insect Pests. Insects. 2022; 13(1):42. https://doi.org/10.3390/insects13010042
Chicago/Turabian StyleWasuwan, Rudsamee, Natnapha Phosrithong, Boonhiang Promdonkoy, Duangjai Sangsrakru, Chutima Sonthirod, Sithichoke Tangphatsornruang, Somsak Likhitrattanapisal, Supawadee Ingsriswang, Chettida Srisuksam, Kewarin Klamchao, and et al. 2022. "The Fungus Metarhizium sp. BCC 4849 Is an Effective and Safe Mycoinsecticide for the Management of Spider Mites and Other Insect Pests" Insects 13, no. 1: 42. https://doi.org/10.3390/insects13010042