
First Report of the Immature Stages of the Leaf-Mining Genus Subclemensia Kozlov, 1987 (Lepidoptera: Incurvariidae), with a Re-Illustration of the Type Species and a Generic Concept Discussion Based on Immature Characters


Abstract
:Simple Summary
Abstract
1. Introduction
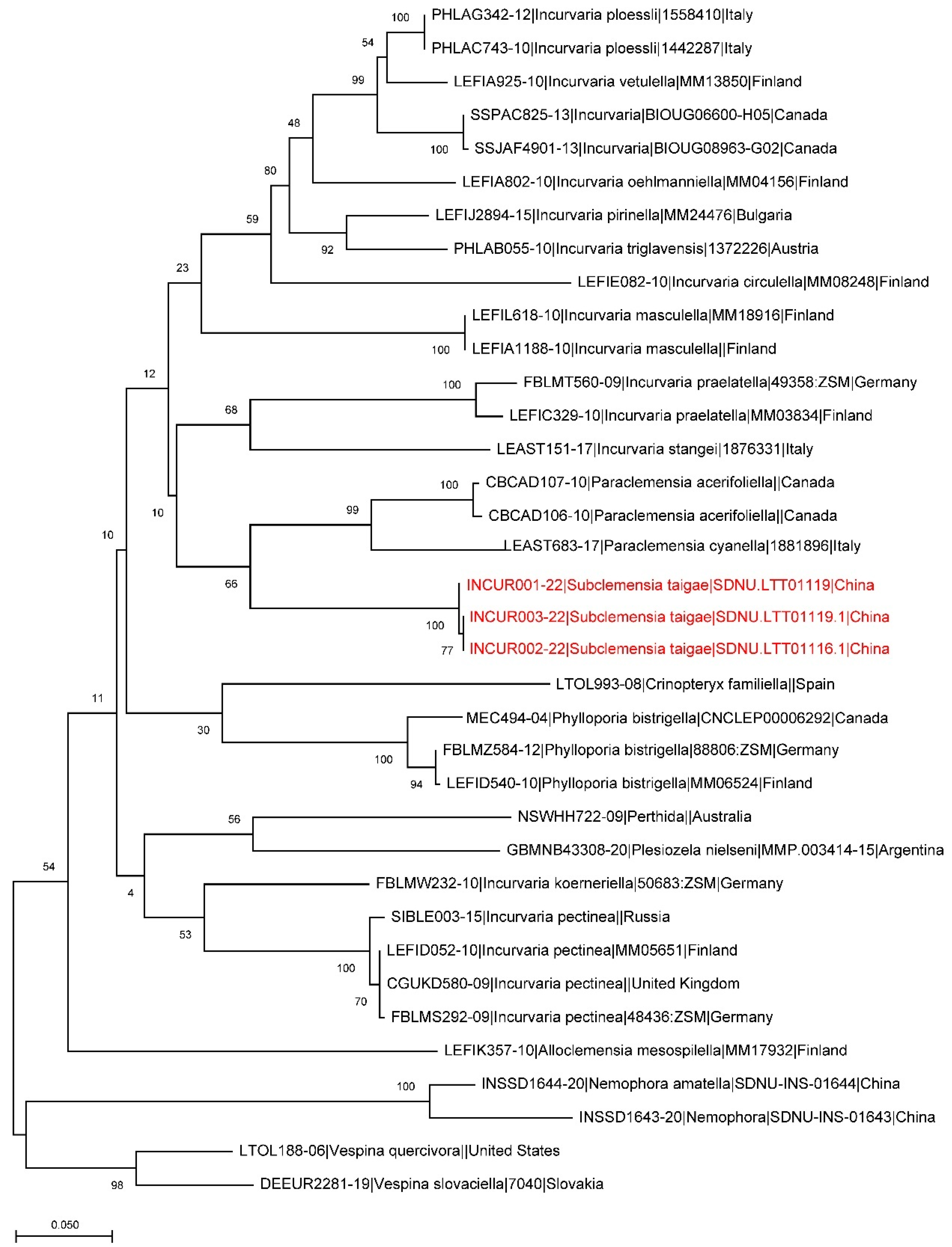
2. Materials and Methods
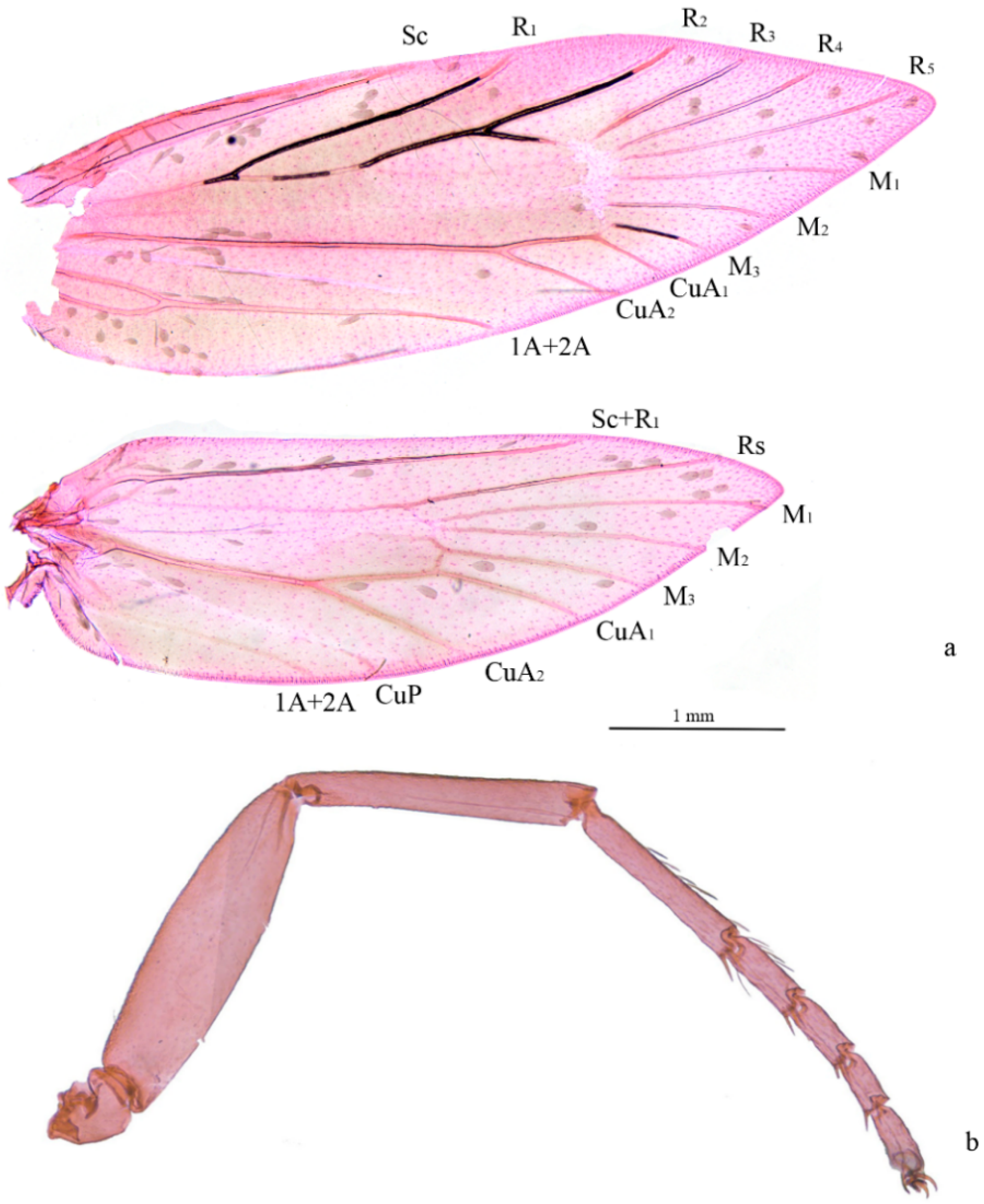
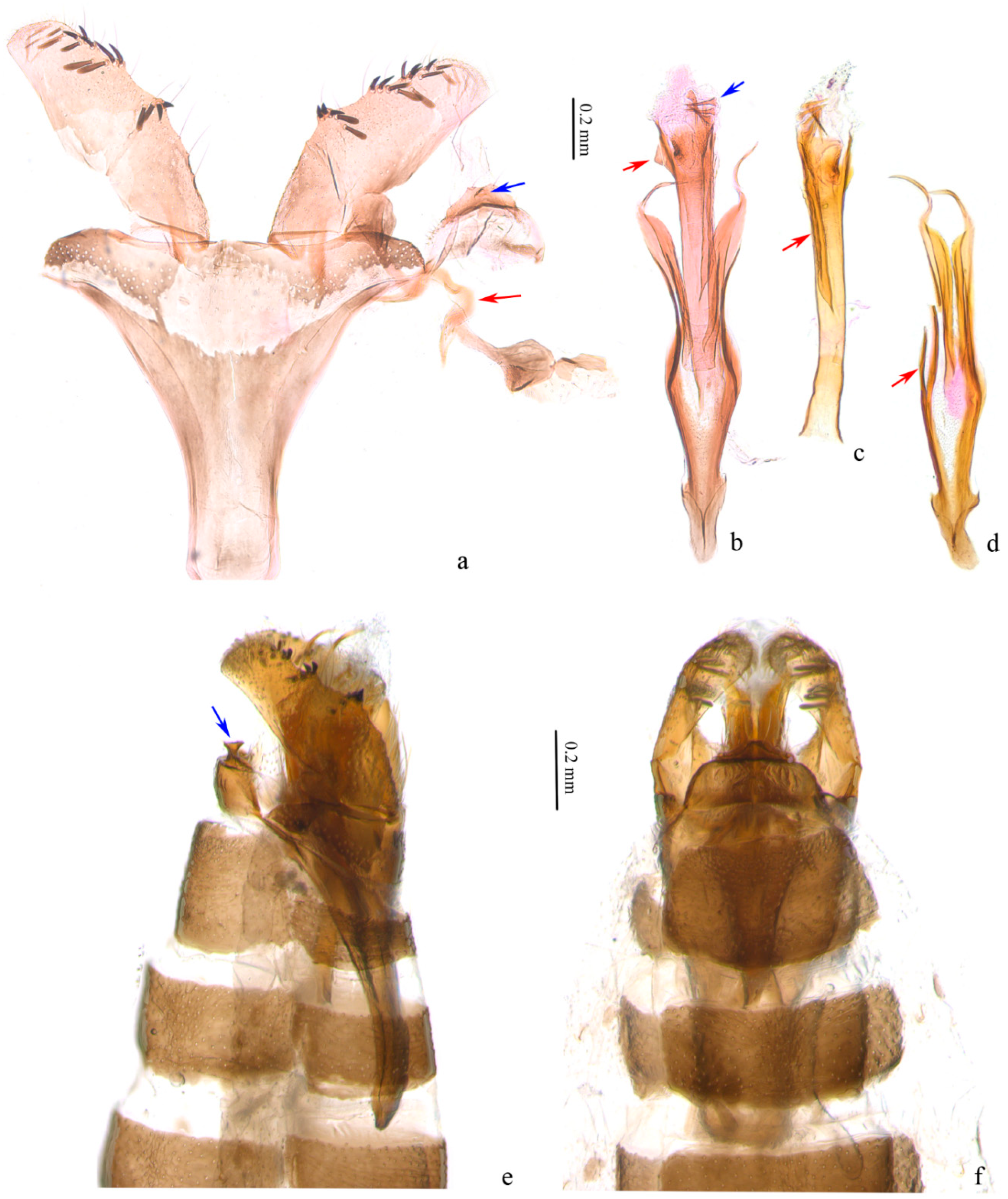
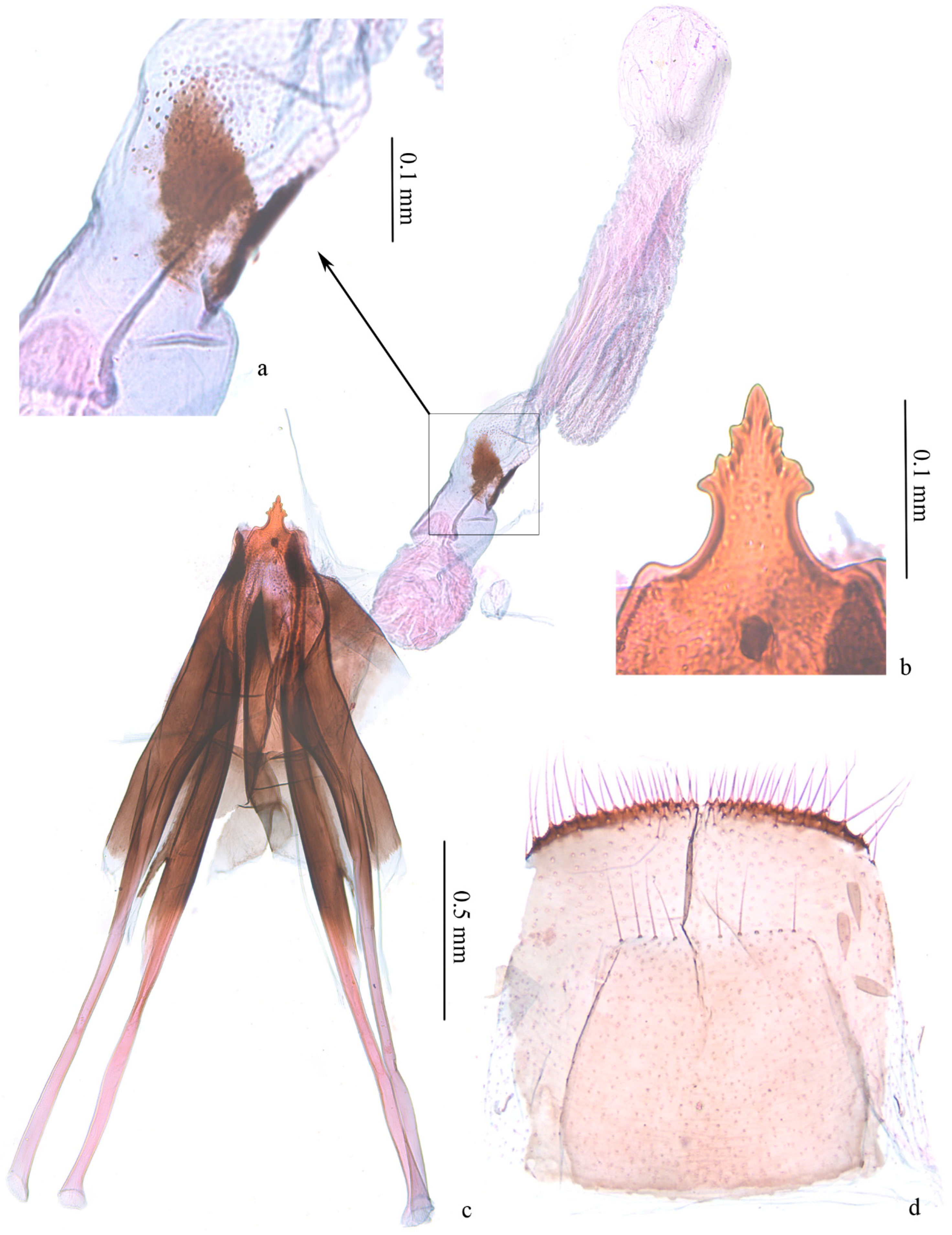
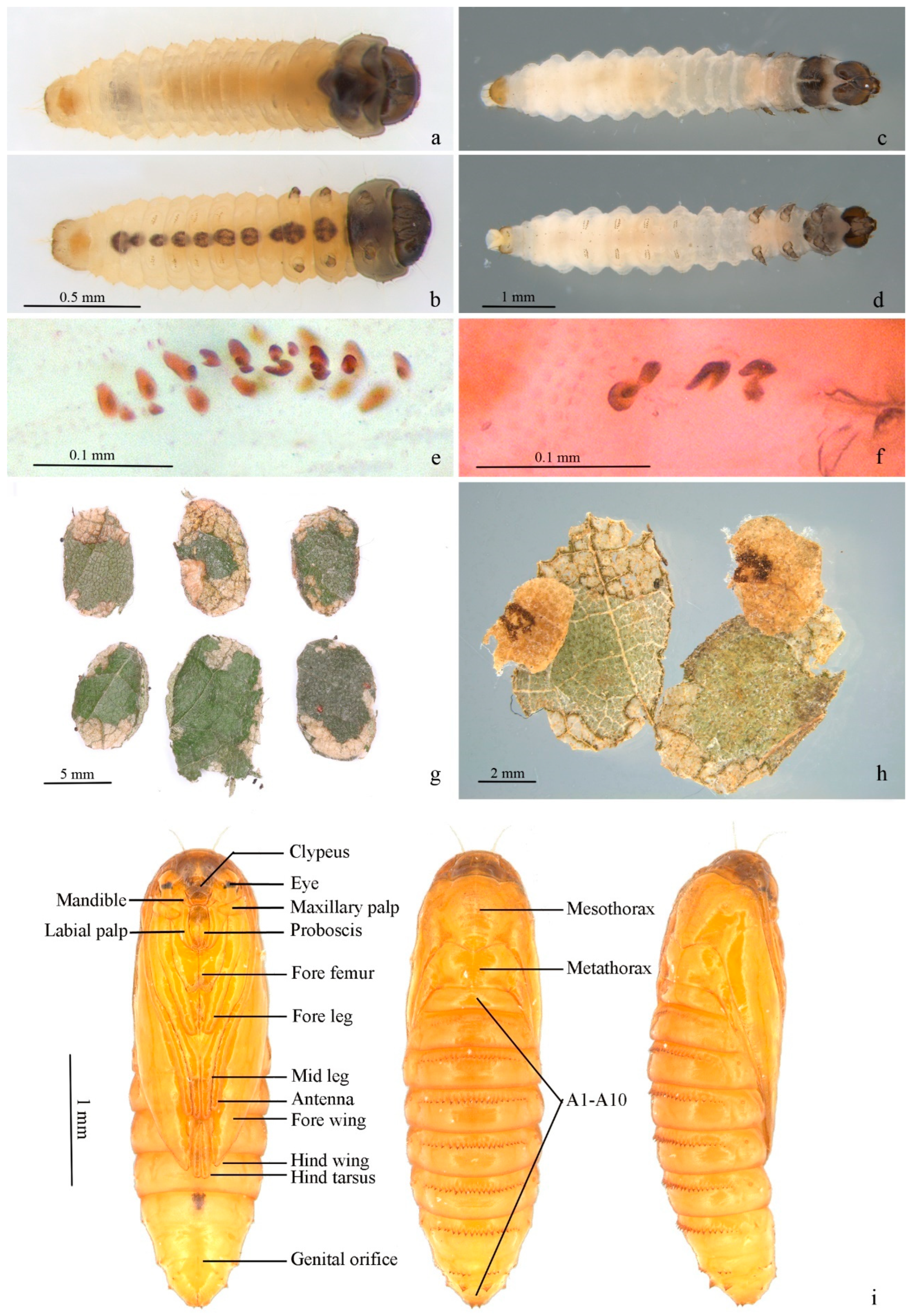
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SDNU | Zoological Collection, Shandong Normal University, Jinan 250014, China |
| A1–A8 | Abdominal segments 1–8 |
| T1–T3 | Thoracic segments 1–3 |
References
- Regier, J.C.; Mitter, C.; Kristensen, N.P.; Davis, D.R.; van Nieukerken, E.J.; Rota, J.; Simonsen, T.J.; Mitter, K.T.; Kawahara, A.Y.; Yen, S.-H.; et al. A molecular phylogeny for the oldest (nonditrysian) lineages of extant Lepidoptera, with implications for classification, comparative morphology and life-history evolution. Syst. Entomol. 2015, 40, 671–704. [Google Scholar] [CrossRef]
- Junnilainen, J. Incurvaria pirinella sp. nov., a new species of the Vetulella species-group (Lepidoptera, Incurvariidae) from Bulgaria, with release of DNA barcodes for European species of Incurvaria. Nota Lepidopterol. 2019, 42, 81–100. [Google Scholar] [CrossRef] [Green Version]
- Hirowatari, T.; Yagi, S.; Ohshima, I.; Huang, G.-H.; Wang, M. Review of the genus Vespina (Lepidoptera, Incurvariidae) with two new species from China and Japan. Zootaxa 2021, 4927, 209–233. [Google Scholar] [CrossRef] [PubMed]
- Karsholt, O.; Kristensen, N.P. Plesiozela, gen. nov. from temperate South America: Apparent sister-group of the previously known Heliozelidae (Lepidoptera: Incurvarioidea: Heliozelidae). Invertebr. Syst. 2003, 17, 39–46. [Google Scholar] [CrossRef]
- Milla, L.; Moussalli, A.; Wilcox, S.A.; Nieukerken, E.J.; Young, D.A.; Halsey, M.; McConville, T.; Jones, T.M.; Kallies, A.; Hilton, D.J. Phylotranscriptomics resolves phylogeny of the Heliozelidae (Adeloidea: Lepidoptera) and suggests a Late Cretaceous origin in Australia. Syst. Entomol. 2020, 45, 128–143. [Google Scholar] [CrossRef]
- Scoble, M.J. Description of Protaephagus capensis. J. Entomol. Soc. S. Afr. 1980, 43, 79–81. [Google Scholar]
- Common, I.F.B. A new genus Perthida for the Western Australian jarrah leaf miner P. glyphopa sp. n. and Tinea phoenicopa Meyrick (Lepidoptera: Incurvariidae). J. Aust. Entomol. Soc. 1969, 8, 128. [Google Scholar] [CrossRef]
- Nielsen, E.S.; Davis, D.R. A revision of Neotropical Incurvariidae s str, with the description of two new genera and two new species Lepidoptera Incurvarioidea. Steenstrupia 1981, 73, 25–57. [Google Scholar]
- Kozlov, M.V. Moths of subfamilies Incurvariinae and Prodoxinae (Lepidoptera, Adelidae) of the Far East. In Lepidoptera of the Far East of the U.S.S.R.; Institute of Biology and Soil Sciences: Vladivostok, Russia, 1987; pp. 14–24. [Google Scholar]
- Nielsen, E.S. The maple leaf-cutter moth and its allies: A revision of Paraclemensia (Incurvariidae s. str.). Syst. Entomol. 1982, 7, 217–238. [Google Scholar] [CrossRef]
- Ross, D.A. The Maple Leaf Cutter, Paraclemensia acerifoliella (Fitch) (Lepidoptera: Incurvariidae), descriptions of stages. Can. Entomol. 1958, 90, 541–555. [Google Scholar] [CrossRef]
- Pohl, G.R.; Jaeger, C.; Nazari, V.; Schmidt, C.; Richard, D.; Gosche, S. Paraclemensia acerifoliella (Lepidoptera: Incurvariidae) in western Canada: A newly discovered host, an expanded range, and biogeographical considerations. Can. Entomol. 2015, 147, 459–471. [Google Scholar] [CrossRef]
- Lundblad, O. Lilla vinbärsmalen (Incurvaria trimaculella quadrimaculella Höfn.), ett för vårt land nytt skadedjur på vinbär. Flygbl. CentAnst. FörsVäs. JordbrOmr. LandbrEnt. 1927, 121, 1–8. [Google Scholar]
- Lundblad, O. Lilla vinbärsmalen, Incurvaria trimaculella quadrimaculella Höfn., en som skadedjur på vinbär hittills okänd och för vårt land ny malfjäril. Meddn CentAnst. FörsVäs. JordbrOmr. Stockh. Ent. Afd. 1930, 374, 1–25. [Google Scholar]
- Nielsen, E.S. A taxonomic revision of the species of Alloclemensia n. gen. (Lepidoptera: Incurvariidae s. str.). Insect Syst. Evol. 1981, 12, 271–294. [Google Scholar] [CrossRef]
- Li, H. The Gelechiidae of China (I) (Lepidoptera: Gelechioidea); Nankai University Press: Tianjin, China, 2002; p. 504. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- deWaard, J.R.; Ivanova, N.V.; Hajibabaei, M.; Hebert, P.D.N. Assembling DNA Barcodes: Analytical Protocols. Methods Mol. Biol. Environ. Genom. 2008, 2008, 275–293. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K.; Battistuzzi, F.U. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- APG. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- The Plant List. Version 1.1. Published on the Internet. Available online: http://www.theplantlist.org/ (accessed on 8 March 2022).
- Zimmerman, E. Insects of Hawaii; Microlepidoptera Part 1; University of Hawai’i Press: Honolulu, HI, USA, 1978; Volume 9, p. 881. [Google Scholar]
- Denis, M.; Schiffermüller, I. Ankündung Eines Systematischen Werkes von den Schmetterlingen der Wienergegend; Augustin Bernardi: Wien, Austria, 1775; p. 323. [Google Scholar]
- Kuprijanov, A.V. Towards a natural system of the Incurvarioidea. Part 3: Excurvaria gen. nov. for Incurvaria praelatella (Denis & Schiffermüller, 1775) (Lepidoptera, Incurvariidae s. str.). Atalanta 1994, 25, 391–398. [Google Scholar]
- Haworth, A.H. Lepidoptera Britannica, Sistens Digestionem Novam Insectorum Lepidopterorum Quae in Magne Britannia Reperiuntur, Larvarum Pabulo, Temporeque Pascendi, Expansione Alarum; Mensibusque Volandi; Synonymis Atque Locis Observationibusque Variis; R. Taylor: London, UK, 1828; Volume 4, pp. 513–609. [Google Scholar]
- Okamoto, H.; Hirowatari, T. Distributional records and biological notes on Japanese species of the family Incurvariidae (Lepidoptera). Trans. Lepidopterol. Soc. Jpn. 2004, 55, 173–195. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | BOLD Process ID | Collecting Date | Country | Depository |
|---|---|---|---|---|
| Alloclemensia mesospilella | LEFIK357-10 | 2 June 2009 | Finland | University of Oulu, Zoological Museum |
| Crinopteryx familiella | LTOL993-08 | 3 January 2001 | Spain | University of Maryland |
| Incurvaria circulella | LEFIE082-10 | - | Finland | University of Oulu, Zoological Museum |
| Incurvaria koerneriella | FBLMW232-10 | 30 April 2005 | Germany | Research Collection of Peter Lichtmannecker |
| Incurvaria masculella | LEFIL618-10 | - | Finland | University of Oulu, Zoological Museum |
| Incurvaria masculella | LEFIA1188-10 | 19 May 2007 | Finland | University of Oulu |
| Incurvaria oehlmanniella | LEFIA802-10 | - | Finland | University of Oulu, Zoological Museum |
| Incurvaria pectinea | CGUKD580-09 | 9 February 2007 | United Kingdom | Natural History Museum, London |
| Incurvaria pectinea | FBLMS292-09 | 15 April 2009 | Germany | Research Collection of Theo Gruenewald |
| Incurvaria pectinea | SIBLE003-15 | 18 June 2010 | Russia | Institut National de la Recherche Agronomique, Zoologie Forestiere |
| Incurvaria pectinea | LEFID052-10 | 16 May 2007 | Finland | University of Oulu, Zoological Museum |
| Incurvaria pirinella | LEFIJ2894-15 | 3 May 2013 | Bulgaria | Research Collection of Jari Junnilainen |
| Incurvaria ploessli | PHLAG342-12 | 21 July 1991 | Italy | Tiroler Landesmuseen |
| Incurvaria ploessli | PHLAC743-10 | 21 July 1991 | Italy | Tiroler Landesmuseen |
| Incurvaria praelatella | FBLMT560-09 | 24 June 2004 | Germany | Research Collection of Alfred Haslberger |
| Incurvaria praelatella | LEFIC329-10 | 19 June 2006 | Finland | University of Oulu, Zoological Museum |
| Incurvaria sp. | SSPAC825-13 | 14 July 2012 | Canada | Centre for Biodiversity Genomics |
| Incurvaria sp. | SSJAF4901-13 | 21 July 2012 | Canada | Centre for Biodiversity Genomics |
| Incurvaria stangei | LEAST151-17 | 23 June 2017 | Italy | Tiroler Landesmuseen |
| Incurvaria triglavensis | PHLAB055-10 | 1 July 2009 | Austria | Tiroler Landesmuseen |
| Incurvaria vetulella | LEFIA925-10 | - | Finland | University of Oulu, Zoological Museum |
| Nemophora amatella | INSSD1644-20 | 12 May 2019 | China | Shandong Normal University |
| Nemophora sp. | INSSD1643-20 | 1 July 2017 | China | Shandong Normal University |
| Paraclemensia acerifoliella | CBCAD107-10 | 9 June 2009 | Canada | Centre for Biodiversity Genomics |
| Paraclemensia acerifoliella | CBCAD106-10 | 9 June 2009 | Canada | Centre for Biodiversity Genomics |
| Subclemensia taigae | INCUR003-22 | 14 August 2021 | China | Shandong Normal University |
| Subclemensia taigae | INCUR002-22 | 14 August 2021 | China | Shandong Normal University |
| Subclemensia taigae | INCUR001-22 | 9 September 2021 | China | Shandong Normal University |
| Paraclemensia cyanella | LEAST683-17 | 20 May 2004 | Italy | Tiroler Landesmuseen |
| Perthida sp. | NSWHH722-09 | 28 December 2008 | Australia | Centre for Biodiversity Genomics |
| Phylloporia bistrigella | MEC494-04 | 19 June 2004 | Canada | Canadian National Collection of Insects, Arachnids and Nematodes |
| Phylloporia bistrigella | FBLMZ584-12 | 16 July 2010 | Germany | Research Collection of Peter Lichtmannecker |
| Phylloporia bistrigella | LEFID540-10 | 3 July 2007 | Finland | University of Oulu, Zoological Museum |
| Plesiozela nielseni | GBMNB43308-20 | - | Argentina | Mined from GenBank, NCBI |
| Vespina quercivora | LTOL188-06 | 4 December 1992 | United States | University of Maryland |
| Vespina slovaciella | DEEUR2281-19 | 18 April 2015 | Slovakia | Research Collection of Zdenko Tokar |
| Leaf Mine before Hole of Cut-Off | Portable Case of Mature Larva | Mature Larva | Pupa | |
|---|---|---|---|---|
| S. taigae | A very short and irregular corridor, or a patch | Outer layer composed of two pieces of leaf tissue | No sclerotized zones on tergite of T2 and T3 | 1. Hind leg reaching middle of A6; 2. A single row of spines on A2–8. |
| A. mesospilella | A slender line, same in width (Lepiforum) | Outer layer composed of several pieces of leaf tissue [8] | Paired small sclerotization on lateral side of tergite of T2 and T3 (Lepiforum) | 1. Hind leg reaching middle of A6 (Lepiforum); 2. Not examined. |
| A. unifasciata | Not available | Outer layer composed of one piece of leaf tissue [8] | Paired small sclerotization on lateral side of tergite of T2 [8] | 1. Hind leg reaching posterior margin of A8; 2. A single row of spines on A3–8 [8]. |
| P. acerifoliella | A broad corridor or a patch [12] | Outer layer composed of one piece of leaf tissue (Pohl et al. 2015: Figure 13) [12], but can also be of two pieces | Large sclerotization on T2 and T3 | 1. Hind leg beyond anterior margin of A8; 2. A single row of spines on A2–8 [11]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, T.; Geng, X.; Tang, Y.; Li, B.; Zhang, H.; Teng, K. First Report of the Immature Stages of the Leaf-Mining Genus Subclemensia Kozlov, 1987 (Lepidoptera: Incurvariidae), with a Re-Illustration of the Type Species and a Generic Concept Discussion Based on Immature Characters. Insects 2022, 13, 440. https://doi.org/10.3390/insects13050440
Liu T, Geng X, Tang Y, Li B, Zhang H, Teng K. First Report of the Immature Stages of the Leaf-Mining Genus Subclemensia Kozlov, 1987 (Lepidoptera: Incurvariidae), with a Re-Illustration of the Type Species and a Generic Concept Discussion Based on Immature Characters. Insects. 2022; 13(5):440. https://doi.org/10.3390/insects13050440
Chicago/Turabian StyleLiu, Tengteng, Xiaoping Geng, Ye Tang, Baozhu Li, Haixin Zhang, and Kaijian Teng. 2022. "First Report of the Immature Stages of the Leaf-Mining Genus Subclemensia Kozlov, 1987 (Lepidoptera: Incurvariidae), with a Re-Illustration of the Type Species and a Generic Concept Discussion Based on Immature Characters" Insects 13, no. 5: 440. https://doi.org/10.3390/insects13050440