
Revisiting the Raractocetus Fossils from Mesozoic and Cenozoic Amber Deposits (Coleoptera: Lymexylidae)
by
 , , , , and
, , , , and
Yan-Da Li
1,2 ,
,
David Peris
3,4,
Shûhei Yamamoto
5,
Yun Hsiao
6,7 ,
,
Alfred F. Newton
8 and
Chen-Yang Cai
1,2,*
1
State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Centre for Excellence in Life and Palaeoenvironment, Chinese Academy of Sciences, Nanjing 210008, China
2
School of Earth Sciences, University of Bristol, Life Sciences Building, Tyndall Avenue, Bristol BS8 1TQ, UK
3
Departament de Dinàmica de la Terra i de l’Oceà, Facultat de Ciències de la Terra, Universitat de Barcelona, 08028 Barcelona, Spain
4
Institut de Recerca de la Biodiversitat (IRBio), Universitat de Barcelona, 08028 Barcelona, Spain
5
Hokkaido University Museum, Hokkaido University, Kita 10, Nishi 8, Kita-ku, Sapporo 060-0810, Japan
6
Australian National Insect Collection, Commonwealth Scientific and Industrial Research Organisation, GPO Box 1700, Canberra, ACT 2601, Australia
7
Division of Ecology and Evolution, Research School of Biology, Australian National University, Canberra ACT 2601, Australia
8
Negaunee Integrative Research Center, Field Museum of Natural History, 1400 S Lake Shore Drive, Chicago, IL 60605, USA
*
Author to whom correspondence should be addressed.
Insects 2022, 13(9), 768; https://doi.org/10.3390/insects13090768
Submission received: 23 July 2022
/
Revised: 22 August 2022
/
Accepted: 23 August 2022
/
Published: 25 August 2022
(This article belongs to the Special Issue Beetle Diversity)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Lymexylidae is a small beetle family, with some members exhibiting strongly reduced elytra and largely exposed functional hind wings. Previously, four species from Kachin (Myanmar), Baltic, and Rovno ambers were assigned to the extant lymexylid genus Raractocetus. Our new examination suggests that these fossils are morphologically separated from the extant Raractocetus, primarily in the hind wing venation, and should be removed from Raractocetus.
Abstract
The fossils once assigned to Raractocetus Kurosawa from the Mesozoic and Cenozoic amber deposits differ from extant Raractocetus in the longer elytra, the more strongly projecting metacoxae, and the hind wing with vein 2A forked. Thus, these fossils should be removed from Raractocetus. Cretoquadratus engeli Chen from Kachin amber appears to be conspecific with R. fossilis Yamamoto. As a result, R. fossilis and R. extinctus Yamamoto from Kachin amber, R. balticus Yamamoto from Baltic amber, and R. sverlilo Nazarenko, Perkovsky & Yamamoto from Rovno amber are transferred to Cretoquadratus Chen, as C. fossilis (Yamamoto) comb. nov., C. extinctus (Yamamoto) comb. nov., C. balticus (Yamamoto) comb. nov., and C. sverlilo (Nazarenko, Perkovsky & Yamamoto) comb. nov., and C. engeli syn. nov. is suggested to be a junior synonym of C. fossilis.
1. Introduction
With 84 worldwide distributed species in 19 genera (including fossils), Lymexylidae is divided into four subfamilies: Atractocerinae (nine genera, with three extinct), Hylecoetinae (one genus), Lymexylinae (one genus), and Melittommatinae (five genera, with one extinct) [1,2,3,4,5,6,7]. In addition, one fossil genus was unable to be confidently assigned to any existing subfamily [8,9].
Historically, Lymexylidae had once been associated with Cleroidea or Cucujoidea [10,11], but molecular studies have recovered an affinity with Tenebrionoidea. In three of the studies, Lymexylidae appeared to be nested within basal Tenebrionoidea, with various positions [12,13,14], although only a few gene fragments were sampled in these studies. Other studies, including three recent phylogenomic ones, suggested Lymexylidae as the sister group of (the remaining) Tenebrionoidea [15,16,17,18,19]. Wheeler [20] concluded that the maxillary palporgan is the strongest autapomorphy for this family, and considered its loss in Australymexylon Wheeler as secondary.
The fossil record of Lymexylidae is relatively abundant. Many lymexylid fossils have been recently reported from several Mesozoic and Cenozoic deposits. Almost all known fossils from the family are from amber deposits [3,4,5,6,7,8,21]. As resiniferous deposits seem to entrap arthropod fauna living primarily in or near the resiniferous trees [22], a potential relationship between the family and gymnosperm trees, the source of Kachin, Baltic and Rovno ambers [23,24,25,26], can be postulated, given the close developmental association of modern lymexylids with trees (see Section 4).
Four species of fossil lymexylids from Mesozoic and Cenozoic deposits were assigned to the extant genus Raractocetus Kurosawa [5,6]. However, these Raractocetus fossils differ from extant members of Raractocetus in having longer elytra and more strongly projecting metacoxae. In this study, we examine additional specimens of R. fossilis Yamamoto from mid-Cretaceous Kachin amber, which provide new clues for the systematic placement of these Raractocetus fossils.
2. Materials and Methods
The new specimens of Cretoquadratus fossilis (Yamamoto) (NIGP180657 and NIGP180658; Figure 1, Figure 2, Figure 3 and Figure 4) originated from amber mines near Noije Bum (26°20′ N, 96°36′ E), Hukawng Valley, Kachin State, northern Myanmar, and are deposited in the Nanjing Institute of Geology and Palaeontology (NIGP), Chinese Academy of Sciences, Nanjing, China. Amber pieces containing the new specimens were trimmed with a small table saw, ground with emery paper of different grit sizes, and finally polished with polishing powder.
For a comparative purpose, S.Y. examined the holotypes of Raractocetus fossilis Yamamoto, R. extinctus Yamamoto, and R. balticus Yamamoto, which are deposited in the Gantz Family Collections Center, Field Museum of Natural History (FMNH), Chicago, IL, USA; Y.H. examined specimens of extant R. kreusleri (Pascoe), which are deposited in the Australian National Insect Collection (ANIC), Commonwealth Scientific and Industrial Research Organisation (CSIRO), Canberra, Australia.
Photographs under incident light were taken with a Zeiss Discovery V20 stereo microscope. Confocal images were obtained with a Zeiss LSM710 confocal laser scanning microscope, using the 561 nm (DPSS 561-10) laser excitation line [27]. Images under incident light were stacked in Helicon Focus 7.0.2. Confocal images were semi-manually stacked with Helicon Focus 7.0.2 and Adobe Photoshop CC. Images were further processed in Adobe Photoshop CC to adjust brightness and contrast.
The terms for hindwing venation follow Wheeler [20], without regard to whether they are anatomically correct.
This published work has been registered in ZooBank, the official registry of Zoological Nomenclature. The LSID for this publication is urn:lsid:zoobank.org:pub:16BAE184-E1DE-4976-A0DB-68698CED438C.
3. Systematic Palaeontology
- Order Coleoptera Linnaeus, 1758
- Suborder Polyphaga Emery, 1886
- Superfamily Lymexyloidea Fleming, 1821
- Family Lymexylidae Fleming, 1821
- Subfamily Atractocerinae Laporte, 1840
- Genus Cretoquadratus Chen, 2019
- Type species. Cretoquadratus engeli Chen, 2019.
Composition. Cretoquadratus fossilis (Yamamoto, 2019) comb. nov. (=Cretoquadratus engeli Chen, 2019 syn. nov.), Cretoquadratus extinctus (Yamamoto, 2019) comb. nov., Cretoquadratus balticus (Yamamoto, 2019) comb. nov., Cretoquadratus sverlilo (Nazarenko, Perkovsky & Yamamoto, 2022) comb. nov.
Revised diagnosis. Eyes large, occupying almost entire frons, nearly contiguous anteriorly (Figure 2A,B, Figure 3B, and Figure 4A; Yamamoto [5]: supplementary Figures S4b,c, S6c,e and S8c,d; Yamamoto et al., 2022: Figure 4A,C). Antennae more or less fusiform (Figure 2 and Figure 4B; Yamamoto [5]: supplementary Figures S7b and S8e; Yamamoto et al. [6]: Figure 3A). Pronotal disc subquadrate or slightly elongate (Figure 3A; Yamamoto [5]: supplementary Figures S4 and S8b; Chen [4]: Figure 1.3). Elytra reduced, but relatively oblong, each more than three times as long as wide (Figure 2C and Figure 3D; Yamamoto [5]: Figure 2g–i; Yamamoto et al. [6]: Figure 4D). Hind wings with M+Cu fork absent, 1A unforked, and 2A forked (Figure 2D–G and Figure 3E; see also Yamamoto et al. [6]: Figure 2). Metacoxae relatively strongly projecting posteriorly (Yamamoto [5]: supplementary Figures S5b, S7a and S9b; Yamamoto et al. [6]: Figure 3D). Tibial spurs 0-0-0 in male [4], 0-1-1 in female (Figure 4D–F; see also [5]).
Remarks. Yamamoto [5] and Yamamoto et al. [6] reported several fossils of Atractocerinae from Mesozoic and Cenozoic amber deposits. Among them, four species were assigned to the extant genus Raractocetus, i.e., R. fossilis and R. extinctus from Kachin amber, R. balticus from Baltic amber, and R. sverlilo from Rovno amber of Ukraine. Raractocetus is closely related to Atractocerus Palisot de Beauvois, and contains only two extant species [28]. As characterized by Paulus [1], extant Raractocetus (and also Atractocerus) have very short elytra, which are only slightly longer than wide, and non-projecting metacoxae, which are not longer than the trochanters. However, the fossil species assigned to Raractocetus have longer elytra and more strongly projecting metacoxae [5,6]. Our new specimens of R. fossilis, NIGP180657 and NIGP180658, further provide additional information on the hind wing venation. Extant Raractocetus has hind wing venation essentially equal to Atractocerus, where vein 1A is forked (e.g., Wheeler [20]: Figures 127, 128; Casari & Albertoni [29]: Figure 48), whereas in R. fossilis vein 1A is unforked and, instead, vein 2A is forked (Figure 2D–G and Figure 3E). As such, it is well justified to place those fossil species formerly placed in Raractocetus in a separate genus.
Chen [4] reported another fossil from Burmese amber, which he assigned to a new genus and species, i.e., Cretoquadratus engeli. The most important character of Cretoquadratus claimed by Chen [4] is the presence of media branches M1–M4 originated from the cross point of the r-m and M+Cu veins, which, however, are not visible in the left hind wing (Chen [4]: Figure 1). Thus, we suggest that such media branches M1–M4 seemingly present on the right wing of C. engeli are likely to be artifacts, and should not be regarded as a diagnostic character. The paired longitudinal ridges on the scutellar shield (mesonotum actually; see [30]), suggested by Chen [4] as an additional diagnostic character for Cretoquadratus, are commonly present in the subfamily Atractocerinae, including the fossil and extant Raractocetus. Other character of Cretoquadratus, for example, the approximation of eyes and the shape of head and pronotum, are well comparable with the fossils assigned to Raractocetus. In fact, the body size, the antennal structure, the pronotal shape, and the darkened elytral apex of C. engeli are essentially identical to those of R. fossilis; therefore, we believe that C. engeli is conspecific with R. fossilis. Both of these names were (likely) first published in 2019, but according to their publishers, Yamamoto [5] was published on 20 March 2019, while Chen [4] was published no earlier than December 2019. Thus, C. fossilis has priority over C. engeli as the valid name of this species.
4. Discussion
In this study, we report the additional materials of “Raractocetus” fossilis from mid-Cretaceous Kachin amber from northern Myanmar. Our materials reveal several additional important character, such as wing veins, which allow further discussion on the generic identity with the recently described similar atractocerine taxa from both Mesozoic and Cenozoic deposits. Based on our new morphological data, with re-observation of some of the previously described material, we propose a total of four new generic combinations and a junior synonymy. As a result, the mid-Cretaceous genus Cretoquadratus is now known from not only the Mesozoic but also from the Cenozoic (Eocene), which indicates that Cretoquadratus successfully survived the K-Pg mass extinction, but likely went extinct later and does not last until the present day. This finding also reinforces the idea that the genus was once widely distributed, based on its presence in various fossil localities.
Larvae of all known lymexylid species develop in decaying wood or tree trunks [31]. This wood-boring habit of larvae and their feeding primarily on symbiotic fungi cultivated inside the galleries [20,31,32,33] is what led Peris et al. [34] to suggest that Lymexylidae might actually be considered as ambrosia beetles, following the criteria by Kirkendall et al. [35]. The estimated origin for the earliest groups of ambrosia fungi during the Cretaceous period concurs with a known high diversity of Lymexylidae from the amber record [34,36], which has been suggested to have an Early Jurassic or even Triassic origin based on molecular dating [18,19]. By contrast, more typical ambrosia beetles, Scolytinae and Platypodinae (Curculionidae), radiated later (although the earliest scolytine fossil is known from the Early Cretaceous [37]), developing a fungus-farming behavior associated with angiosperms during the Late Cretaceous [38], or perhaps later [34]. This fact points to lymexylids as potential early vectors of ambrosia fungi since the Early Cretaceous, before the close fungal relationship with Scolytinae and Platypodinae evolved [34]. However, the direct impact of wood-borer or ambrosia beetles in resin production during the Cretaceous has not yet been properly demonstrated [36,39] and is a question that needs further attention.
Author Contributions
Conceptualization, Y.-D.L. and C.-Y.C.; methodology, Y.-D.L. and C.-Y.C.; investigation, Y.-D.L., S.Y., Y.H., D.P. and C.-Y.C.; resources, C.-Y.C.; writing—original draft preparation, Y.-D.L., D.P. and S.Y.; writing—review and editing, Y.-D.L., D.P., S.Y., A.F.N. and C.-Y.C.; visualization, Y.-D.L.; project administration, C.-Y.C.; funding acquisition, C.-Y.C. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support was provided by the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB26000000), the National Natural Science Foundation of China (42288201, 42222201), and the Second Tibetan Plateau Scientific Expedition and Research project (2019QZKK0706).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original confocal data are available on the Zenodo repository (doi:10.5281/zenodo.7008334).
Acknowledgments
We are grateful to Rong Huang for technical help with confocal imaging. Alexander Kirejtshuk and an anonymous reviewer provided valuable comments on an early version of this paper. Y.-D.L. is supported by a scholarship granted by the China Scholarship Council (202108320010). S.Y. is supported by the Grant-in-Aid for JSPS Fellows (20J00159) from the Japan Society for the Promotion of Science (JSPS). This is contribution no. 8 of the postdoctoral fellowships program Beatriu de Pinós project 2020 BP 00015, The flowering plant success–Influence of beetles, funded to D.P. by the Secretary of Universities and Research (Government of Catalonia) and by the Horizon 2020 program of research and innovation of the European Union under the Marie Curie grant agreement No. 801370.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Paulus, H. Urtea graeca nov. gen. et nov. spec., der erste Vertreter der tropischen Atractocerinae in Europa sowie eine Beschreibung von Hymaloxylon aspoecki nov. spec. aus Yunnan (China) (Coleoptera, Cucujiformia, Lymexylidae, Atractocerinae nov. status). Denisia 2004, 13, 277–290. [Google Scholar]
- Wolf-Schwenninger, K. The oldest fossil record of Lymexylidae (Insecta: Coleoptera) from the Lower Cretaceous Crato formation of Brazil. Insect Syst. Evol. 2011, 42, 205–212. [Google Scholar] [CrossRef]
- Kirejtshuk, A. A new genus and species of timber beetle (Coleoptera: Lymexylidae) from the Baltic amber. Paleontol. J. 2008, 42, 63–65. [Google Scholar] [CrossRef]
- Chen, X. A new genus and species of Lymexylidae (Insecta: Coleoptera) from mid-Cretaceous amber of northern Myanmar. Paleontol. J. 2019, 53, 1203–1205. [Google Scholar] [CrossRef]
- Yamamoto, S. Fossil evidence of elytra reduction in ship-timber beetles. Sci. Rep. 2019, 9, 4938. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Nazarenko, V.Y.; Vasilenko, D.V.; Perkovsky, E.E. First fossil species of ship-timber beetles (Coleoptera, Lymexylidae) from Eocene Rovno amber (Ukraine). Foss. Rec. 2022, 25, 65–74. [Google Scholar] [CrossRef]
- Nazarenko, V.; Perkovsky, E.; Vasilenko, D. New genus and species of Lymexylidae (Coleoptera) from Baltic amber. Paleontol. J. 2020, 54, 1395–1399. [Google Scholar] [CrossRef]
- Chen, X.-Y.; Zhang, H.-C. A new fossil record of Lymexylidae (Insecta: Coleoptera) from mid-Cretaceous amber of northern Myanmar. Zootaxa 2020, 4878, 195–200. [Google Scholar] [CrossRef]
- Perkovsky, E. Anancites nom. nov., a replacement name for Adamas Chen et Wang, 2020 (Coleoptera, Lymexylidae). Paleontol. J. 2022, 56, 327. [Google Scholar] [CrossRef]
- Crowson, R. The phylogeny of Coleoptera. Annu. Rev. Entomol. 1960, 5, 111–134. [Google Scholar] [CrossRef]
- Lawrence, J.F.; Newton, A.F. Evolution and classification of beetles. Annu. Rev. Ecol. Syst. 1982, 13, 261–290. [Google Scholar] [CrossRef] [Green Version]
- Robertson, J.A.; Ślipiński, A.; Moulton, M.; Shockley, F.W.; Giorgi, A.; Lord, N.P.; Mckenna, D.D.; Tomaszewska, W.; Forrester, J.; Miller, K.B. Phylogeny and classification of Cucujoidea and the recognition of a new superfamily Coccinelloidea (Coleoptera: Cucujiformia). Syst. Entomol. 2015, 40, 745–778. [Google Scholar] [CrossRef]
- Mckenna, D.D.; Wild, A.L.; Kanda, K.; Bellamy, C.L.; Beutel, R.G.; Caterino, M.S.; Farnum, C.W.; Hawks, D.C.; Ivie, M.A.; Jameson, M.L. The beetle tree of life reveals that Coleoptera survived end-Permian mass extinction to diversify during the Cretaceous terrestrial revolution. Syst. Entomol. 2015, 40, 835–880. [Google Scholar] [CrossRef]
- Hunt, T.; Bergsten, J.; Levkanicova, Z.; Papadopoulou, A.; John, O.S.; Wild, R.; Hammond, P.M.; Ahrens, D.; Balke, M.; Caterino, M.S. A comprehensive phylogeny of beetles reveals the evolutionary origins of a superradiation. Science 2007, 318, 1913–1916. [Google Scholar] [CrossRef]
- Bocak, L.; Barton, C.; Crampton-Platt, A.; Chesters, D.; Ahrens, D.; Vogler, A.P. Building the Coleoptera tree-of-life for >8000 species: Composition of public DNA data and fit with Linnaean classification. Syst. Entomol. 2014, 39, 97–110. [Google Scholar] [CrossRef]
- Gunter, N.L.; Levkaničová, Z.; Weir, T.H.; Ślipiński, A.; Cameron, S.L.; Bocak, L. Towards a phylogeny of the Tenebrionoidea (Coleoptera). Mol. Phylogenet. Evol. 2014, 79, 305–312. [Google Scholar] [CrossRef]
- Zhang, S.-Q.; Che, L.-H.; Li, Y.; Liang, D.; Pang, H.; Ślipiński, A.; Zhang, P. Evolutionary history of Coleoptera revealed by extensive sampling of genes and species. Nat. Commun. 2018, 9, 205. [Google Scholar] [CrossRef]
- McKenna, D.D.; Shin, S.; Ahrens, D.; Balke, M.; Beza-Beza, C.; Clarke, D.J.; Donath, A.; Escalona, H.E.; Friedrich, F.; Letsch, H. The evolution and genomic basis of beetle diversity. Proc. Natl. Acad. Sci. USA 2019, 116, 24729–24737. [Google Scholar] [CrossRef]
- Cai, C.; Tihelka, E.; Giacomelli, M.; Lawrence, J.F.; Slipinski, A.; Kundrata, R.; Yamamoto, S.; Thayer, M.K.; Newton, A.F.; Leschen, R.A.; et al. Integrated phylogenomics and fossil data illuminate the evolution of beetles. R. Soc. Open Sci. 2022, 9, 211771. [Google Scholar] [CrossRef]
- Wheeler, Q.D. Revision of the genera of Lymexylidae (Coleoptera, Cucujiformia). Bull. Am. Mus. Nat. Hist. 1986, 183, 113–210. [Google Scholar]
- Kirejtshuk, A.G.; Azar, D. Current knowledge of Coleoptera (Insecta) from the Lower Cretaceous Lebanese amber and taxonomical notes for some Mesozoic groups. Terr. Arthropod Rev. 2013, 6, 103–134. [Google Scholar] [CrossRef]
- Solórzano Kraemer, M.M.; Delclòs, X.; Clapham, M.E.; Arillo, A.; Peris, D.; Jäger, P.; Stebner, F.; Peñalver, E. Arthropods in modern resins reveal if amber accurately recorded forest arthropod communities. Proc. Natl. Acad. Sci. USA 2018, 115, 6739–6744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poinar, G., Jr.; Lambert, J.B.; Wu, Y. Araucarian source of fossiliferous Burmese amber: Spectroscopic and anatomical evidence. J. Bot. Res. Inst. Tex. 2007, 1, 449–455. [Google Scholar]
- Wolfe, A.P.; Tappert, R.; Muehlenbachs, K.; Boudreau, M.; McKellar, R.C.; Basinger, J.F.; Garrett, A. A new proposal concerning the botanical origin of Baltic amber. Proc. R. Soc. B 2009, 276, 3403–3412. [Google Scholar] [CrossRef]
- Wolfe, A.P.; McKellar, R.C.; Tappert, R.; Sodhi, R.N.; Muehlenbachs, K. Bitterfeld amber is not Baltic amber: Three geochemical tests and further constraints on the botanical affinities of succinite. Rev. Palaeobot. Palynol. 2016, 225, 21–32. [Google Scholar] [CrossRef]
- Perkovsky, E.E.; Zosimovich, V.Y.; Vlaskin, A.P. Rovno amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 116–136. [Google Scholar]
- Fu, Y.-Z.; Li, Y.-D.; Su, Y.-T.; Cai, C.-Y.; Huang, D.-Y. Application of confocal laser scanning microscopy to the study of amber bioinclusions. Palaeoentomology 2021, 4, 266–278. [Google Scholar] [CrossRef]
- Kurosawa, Y. Revisional notes on the family Lymexylonidae (Coleoptera) in eastern and southeastern Asia. Bull. Natl. Sci. Mus. Ser. A 1985, 11, 109–119. [Google Scholar]
- Casari, S.A.; Albertoni, F.F. First instar larva of Atractocerus brasiliensis (Lepeletier & Audinet-Serville, 1825) (Lymexylidae, Atractocerinae). Pap. Avulsos Zool. 2013, 53, 359–372. [Google Scholar]
- Lawrence, J.F. The Australian Lymexylidae (Coleoptera: Tenerionoidea) with one new genus and two genera new to Australia. Zootaxa 2020, 4895, 211–238. [Google Scholar] [CrossRef]
- Francke-Grosmann, H. Ectosymbiosis in wood-inhabiting insects. In Symbiosis. Volume II. Associations of Invertebrates, Birds, Ruminants, and Other Biota; Henry, S., Ed.; Academic Press: New York, NY, USA, 1967; pp. 141–205. [Google Scholar]
- Toki, W. A single case study of mycetangia-associated fungi and their abilities to assimilate wood-associated carbon sources in the ship timber beetle Elateroides flabellicornis (Coleoptera: Lymexylidae) in Japan. Symbiosis 2021, 83, 173–181. [Google Scholar] [CrossRef]
- Casari, S.A.; Teixeira, É.P. Larva of Atractocerus brasiliensis (Lepeletier & Audinet-Serville, 1825) (Lymexylidae, Atractocerinae). Pap. Avulsos Zool. 2011, 51, 197–205. [Google Scholar]
- Peris, D.; Delclòs, X.; Jordal, B. Origin and evolution of fungus farming in wood-boring Coleoptera—A palaeontological perspective. Biol. Rev. 2021, 96, 2476–2488. [Google Scholar] [PubMed]
- Kirkendall, L.R.; Biedermann, P.H.; Jordal, B.H. Evolution and diversity of bark and ambrosia beetles. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: London, UK, 2015; pp. 85–156. [Google Scholar]
- Peris, D. Coleoptera in amber from Cretaceous resiniferous forests. Cretac. Res. 2020, 113, 104484. [Google Scholar] [CrossRef]
- Kirejtshuk, A.G.; Azar, D.; Beaver, R.A.; Mandelshtam, M.Y.; Nel, A. The most ancient bark beetle known: A new tribe, genus and species from Lebanese amber (Coleoptera, Curculionidae, Scolytinae). Syst. Entomol. 2009, 34, 101–112. [Google Scholar] [CrossRef]
- Pistone, D.; Gohli, J.; Jordal, B.H. Molecular phylogeny of bark and ambrosia beetles (Curculionidae: Scolytinae) based on 18 molecular markers. Syst. Entomol. 2018, 43, 387–406. [Google Scholar] [CrossRef]
- Peris, D.; Rust, J. Cretaceous beetles (Insecta: Coleoptera) in amber: The palaeoecology of this most diverse group of insects. Zool. J. Linn. Soc. 2020, 189, 1085–1104. [Google Scholar] [CrossRef]
Figure 1.
General habitus of Cretoquadratus fossilis (Yamamoto) comb. nov., NIGP180657, under incident light. (A) Dorsal view. (B) Ventral view. Scale bars: 1 mm.
Figure 1.
General habitus of Cretoquadratus fossilis (Yamamoto) comb. nov., NIGP180657, under incident light. (A) Dorsal view. (B) Ventral view. Scale bars: 1 mm.

Figure 2.
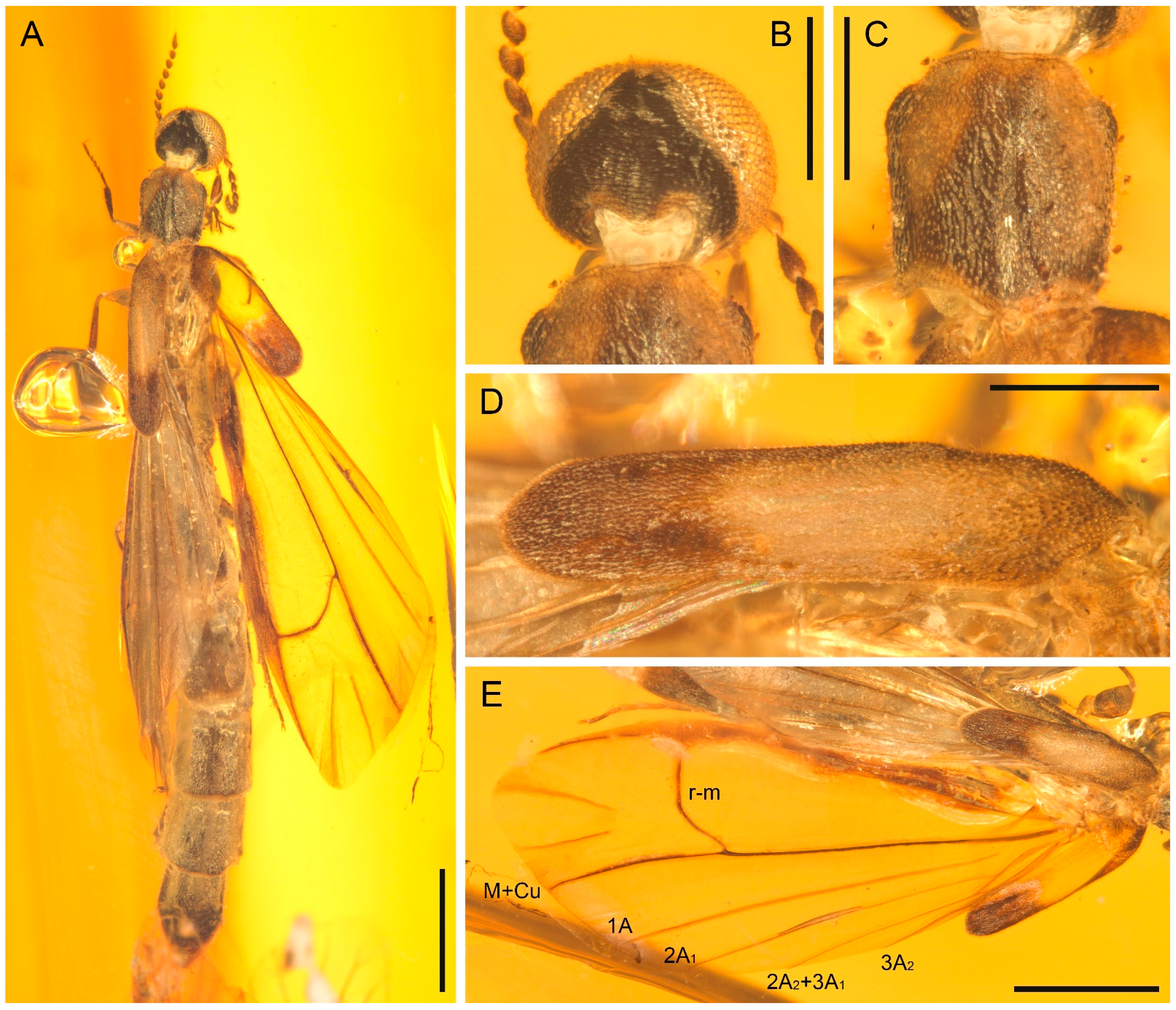
Details of Cretoquadratus fossilis (Yamamoto) comb. nov., NIGP180657, under incident light. (A) Head and prothorax, dorsal view. (B) Head, anterior view. (C) Elytron, dorsal view. (D,E) Right hind wing. (F,G) Left hind wing. Scale bars: 500 μm.
Figure 2.
Details of Cretoquadratus fossilis (Yamamoto) comb. nov., NIGP180657, under incident light. (A) Head and prothorax, dorsal view. (B) Head, anterior view. (C) Elytron, dorsal view. (D,E) Right hind wing. (F,G) Left hind wing. Scale bars: 500 μm.

Figure 3.
Cretoquadratus fossilis (Yamamoto) comb. nov., NIGP180658, under incident light. (A) Habitus, dorsal view. (B) Head, dorsal view. (C) Prothorax, dorsal view. (D) Elytron, dorsal view. (E) Right hind wing. Scale bars: 1 mm in (A,E), 400 μm in (B–D).
Figure 3.
Cretoquadratus fossilis (Yamamoto) comb. nov., NIGP180658, under incident light. (A) Habitus, dorsal view. (B) Head, dorsal view. (C) Prothorax, dorsal view. (D) Elytron, dorsal view. (E) Right hind wing. Scale bars: 1 mm in (A,E), 400 μm in (B–D).

Figure 4.
Details of Cretoquadratus fossilis (Yamamoto) comb. nov., NIGP180657, under confocal microscopy. (A) Head, dorsal view. (B) Antenna and maxillary palps. (C) Ovipositor, dorsal view. (D) Fore leg. (E) Mid legs. (F) Hind leg. Abbreviations: an, antenna; ey, compound eye; gc, gonocoxite; gs, gonostylus; mxp, maxillary palp; mstb, mesotibia; msts, mesotarsus; mttb, metatibia; mtts, metatarsus; ptb, protibia; pts, protarsus. Scale bars: 200 μm.
Figure 4.
Details of Cretoquadratus fossilis (Yamamoto) comb. nov., NIGP180657, under confocal microscopy. (A) Head, dorsal view. (B) Antenna and maxillary palps. (C) Ovipositor, dorsal view. (D) Fore leg. (E) Mid legs. (F) Hind leg. Abbreviations: an, antenna; ey, compound eye; gc, gonocoxite; gs, gonostylus; mxp, maxillary palp; mstb, mesotibia; msts, mesotarsus; mttb, metatibia; mtts, metatarsus; ptb, protibia; pts, protarsus. Scale bars: 200 μm.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, Y.-D.; Peris, D.; Yamamoto, S.; Hsiao, Y.; Newton, A.F.; Cai, C.-Y. Revisiting the Raractocetus Fossils from Mesozoic and Cenozoic Amber Deposits (Coleoptera: Lymexylidae). Insects 2022, 13, 768. https://doi.org/10.3390/insects13090768
AMA Style
Li Y-D, Peris D, Yamamoto S, Hsiao Y, Newton AF, Cai C-Y. Revisiting the Raractocetus Fossils from Mesozoic and Cenozoic Amber Deposits (Coleoptera: Lymexylidae). Insects. 2022; 13(9):768. https://doi.org/10.3390/insects13090768
Chicago/Turabian StyleLi, Yan-Da, David Peris, Shûhei Yamamoto, Yun Hsiao, Alfred F. Newton, and Chen-Yang Cai. 2022. "Revisiting the Raractocetus Fossils from Mesozoic and Cenozoic Amber Deposits (Coleoptera: Lymexylidae)" Insects 13, no. 9: 768. https://doi.org/10.3390/insects13090768
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.