3.1. Desription of the New Genus
Spinopygina gen. nov.
Type species: Camptochaeta uniceps Hippa & Vilkamaa, 1994.
LSID urn:lsid:zoobank.org:act: DBFE5207-337A-4817-B955-14847009D23B.
Etymology. The name is formed from the Latin word spine, thorn, and the Greek word pyge, rump, referring to the outstanding megasetae of the gonostylus of the species.
Diagnosis. Small to medium sized Sciaridae, wing length 1.6–2.2 mm. Antenna long, flagellomeres with long setae and necks. Mouth parts small, maxillary palpus with 2 segments, 2nd segment much reduced. Anal lobe of wing small, halter with long stalk. Body setosity dark, long and strong. Legs long and slender, fore tibial organ poorly differentiated. Intergonocoxal area long or moderate, without medial lobe. Gonocoxae united medially, setae at medial margins short. Gonostylus impressed or deeply excavated medially, with lambda-shaped basomedial sclerotization; with strong megasetae with basal bodies, including (in most species) strongly procurved basal megaseta; without apical tooth. Tegmen modified, with flat apicolateral parts, without detectable aedeagal teeth.
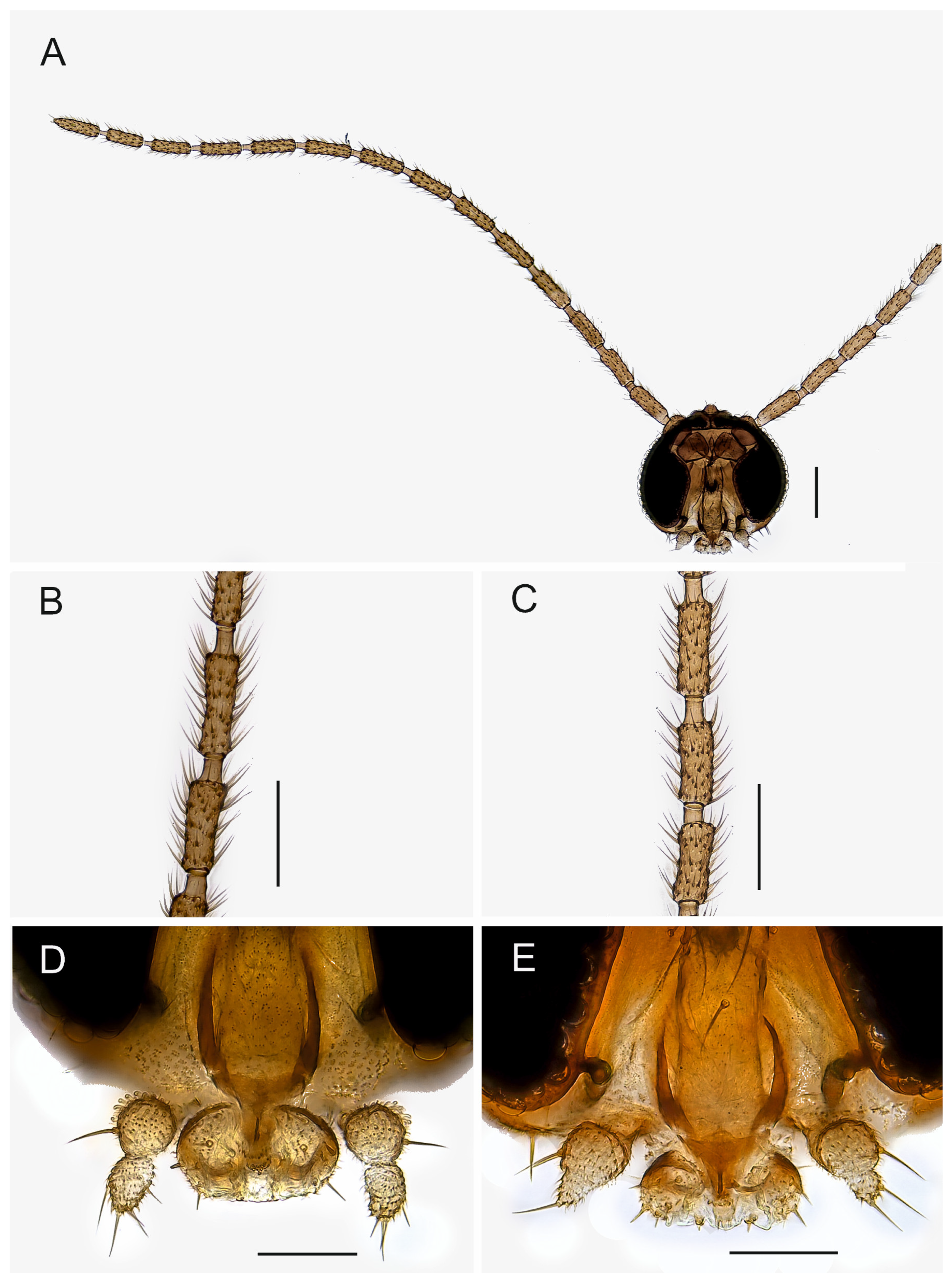
Description. Head (
Figure 1). Normal, roundish. Eye bridge 1 to 2 facets wide. Anterior vertex non-setose. Face setose. Clypeus with 1–2 setae or non-setose. Antennal scape and pedicel normal. Antennal flagellum long, with 14 flagellomeres, bodies of flagellomeres subcylindrical, with smooth surface, body of 4th flagellomere 2.25–3.20× as long as subapically wide, necks longer than wide. Flagellomeral vestiture rather sparse, setae longer than width of flagellomeres. Mouth parts small. Maxillary palpus with 2 segments, 2nd segment strongly reduced, variably between species and even specimens; 1st segment with 1, rarely with 2 sharp setae; with long sparse sensilla dorsally.
Thorax (
Figure 2). Brown; setae dark. Acrostichal setae few anteriorly, dorsocentral and lateral setae in indistinct rows of a few long and strong and short and fine setae. Scutellum with 2 long and strong setae and a few short and fine setae. Posterior pronotum non-setose. Anterior pronotum and prothoracic episternum with a few setae, other pleural sclerites non-setose. Mesothoracic katepisternum high with oblique anterior margin.
Wing (
Figure 3A–C). Fumose. Length 1.6–2.2 mm. Anal lobe small. Veins distinct, except stM. Membrane non-setose, veins C, R and R1 setose, R5 with only dorsal setae, bM, r-m, stM, M and stCuA non-setose. c/w 0.70–0.90, R1/R 0.70–1.15, R1 joining C well before level of base of M-fork. Halter yellow, with long stalk.
Legs (
Figure 3D,E). Yellow or pale brown, long, femora slender; setae dark; tibial spurs 1:2:2, fore tibial spur as long as tibial width or slightly longer, middle and hind left and right tibial spurs subequal in size, longer than tibial width. Fore tibia without spinose setae, middle tibia rarely with 1 spinose seta, hind tibia with dorsal row of strong spinose setae. The retrolateral apical setae of hind tibia fine. Fore tibial organ not impressed or proximally bordered, with small patch of a few setae. Tarsi unmodified, long, length of fore basitarsomere/length of fore tibia 0.47–0.63. Tarsal claws unmodified, without teeth.
Abdomen (
Figure 4,
Figure 5,
Figure 6,
Figure 7,
Figure 8 and
Figure 9). Pale brown or yellowish, normal, slender; setae dark, long and strong. Hypopygium brown or yellowish, as abdomen. Intergonocoxal area long or moderate, without lobe(s). Gonocoxa unmodified, longer than gonostylus, gonocoxae fused ventromedially; rather richly setose, setae at ventromedial margin short, medial membrane non-setose; with 1 elongated seta ventrally and dorsally at apicomedial corner. Gonostylus elongated, widest medially and distinctly narrowed towards apex or evenly wide and slightly narrowed towards apex, strongly excavated or at least slightly impressed medially, with lambda-shaped basomedial sclerotization; without apical tooth; with apical, subapical and medial megasetae, or only apical or subapical megasetae present; megasetae strong and with distinct basal bodies, in some species basalmost megaseta distinctly procurved; with some elongated setae medially and apically. Tegmen variable in shape, truncate or slightly curved apically, slightly curved basolaterally, constricted subapically, membranous or with sclerotized rim apically, with flat apicolateral parts; parameres sclerotized, approaching each other and joining to sclerotized rim or ending at apex separately. Apodemes of tegmen and aedeagal apodeme usually short, aedeagal teeth not detectable. Tergite 9 rather narrow, with long and strong setae. Tergite 10 (cercus) normal.
Female unknown.
Distribution. USA: California, Oregon; Canada: British Columbia.
Figure 4.
Hypopygia, ventral view. (A). Holotype MZH GE.1928 of Spinopygina acerfalx sp. nov. (B). Holotype MZH GE.1932 of S. aurifera sp. nov. Scale bars = 0.1 mm.
Figure 4.
Hypopygia, ventral view. (A). Holotype MZH GE.1928 of Spinopygina acerfalx sp. nov. (B). Holotype MZH GE.1932 of S. aurifera sp. nov. Scale bars = 0.1 mm.
Figure 5.
Hypopygia, ventral view. (A). Holotype MZH GE.1935 of Spinopygina camura sp. nov. (B). Holotype MZH GE.1937 of S. edura sp. nov. Scale bars = 0.1 mm.
Figure 5.
Hypopygia, ventral view. (A). Holotype MZH GE.1935 of Spinopygina camura sp. nov. (B). Holotype MZH GE.1937 of S. edura sp. nov. Scale bars = 0.1 mm.
Figure 6.
Hypopygia, ventral view. (A). Holotype MZH GE.1938 of Spinopygina peltata sp. nov. (B). Holotype MZH GE.1941 of S. plena sp. nov. Scale bars = 0.1 mm.
Figure 6.
Hypopygia, ventral view. (A). Holotype MZH GE.1938 of Spinopygina peltata sp. nov. (B). Holotype MZH GE.1941 of S. plena sp. nov. Scale bars = 0.1 mm.
Figure 7.
Hypopygium of Spinopygina quadracantha sp. nov. (A). Ventral view (holotype MZH GE.1944). (B). Dorsal view (paratype MZH GE.1956). Scale bars = 0.1 mm.
Figure 7.
Hypopygium of Spinopygina quadracantha sp. nov. (A). Ventral view (holotype MZH GE.1944). (B). Dorsal view (paratype MZH GE.1956). Scale bars = 0.1 mm.
Figure 8.
Hypopygium of Spinopygina uniceps (Hippa & Vilkamaa, 1994). (A). Ventral view (holotype CNC). (B). Dorsal view (specimen MZH GE.1962). Scale bars = 0.1 mm.
Figure 8.
Hypopygium of Spinopygina uniceps (Hippa & Vilkamaa, 1994). (A). Ventral view (holotype CNC). (B). Dorsal view (specimen MZH GE.1962). Scale bars = 0.1 mm.
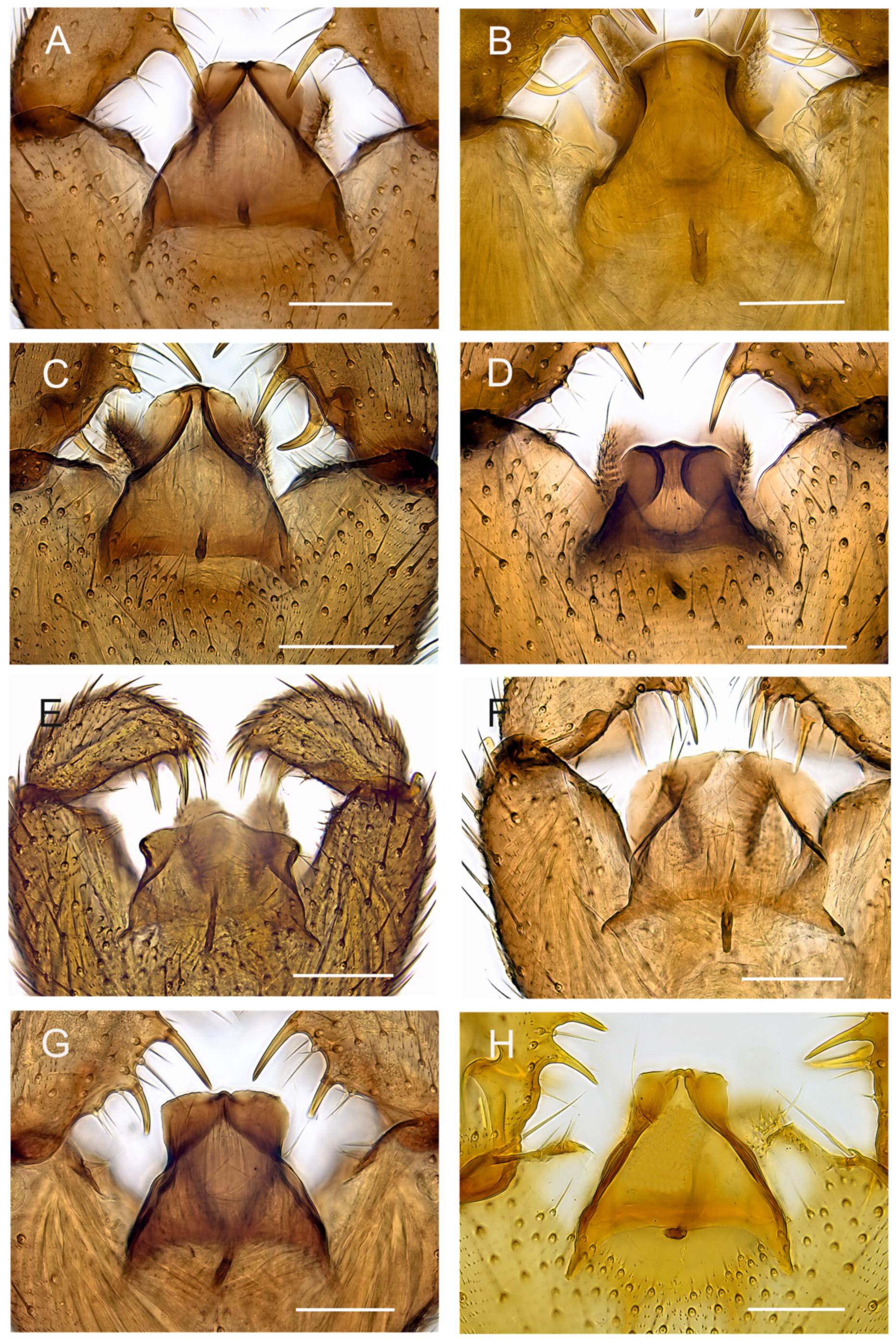
Figure 9.
Tegmina, ventral view. (A). Holotype MZH GE.1928 of Spinopygina acerfalx sp. nov. (B). Holotype MZH GE.1932 of S. aurifera sp. nov. (C). Paratype MZH GE.1936 of S. camura sp. nov. (D). Holotype MZH GE.1937 of S. edura sp. nov. (E). Holotype MZH GE.1938 of S. peltata sp. nov. (F). Holotype MZH GE.1941 of S. plena sp. nov. (G). Holotype MZH GE.1944 of S. quadracantha sp. nov. (H). Holotype CNC of S. uniceps (Hippa & Vilkamaa, 1994) Scale bars = 0.05 mm.
Figure 9.
Tegmina, ventral view. (A). Holotype MZH GE.1928 of Spinopygina acerfalx sp. nov. (B). Holotype MZH GE.1932 of S. aurifera sp. nov. (C). Paratype MZH GE.1936 of S. camura sp. nov. (D). Holotype MZH GE.1937 of S. edura sp. nov. (E). Holotype MZH GE.1938 of S. peltata sp. nov. (F). Holotype MZH GE.1941 of S. plena sp. nov. (G). Holotype MZH GE.1944 of S. quadracantha sp. nov. (H). Holotype CNC of S. uniceps (Hippa & Vilkamaa, 1994) Scale bars = 0.05 mm.
Figure 10.
Maximum likelihood hypothesis for relationships among selected taxa of Sciaridae (Diptera) based on DNA sequence data (18S, 28S, 16S, and COI), 4287 characters. Support numbers refer to ultrafast bootstrap values (ufboot) over 50.
Figure 10.
Maximum likelihood hypothesis for relationships among selected taxa of Sciaridae (Diptera) based on DNA sequence data (18S, 28S, 16S, and COI), 4287 characters. Support numbers refer to ultrafast bootstrap values (ufboot) over 50.
3.4. Descriptions of New Species
LSID urn:lsid:zoobank.org:act: 008D0F6D-919A-4915-99A8-8AB979A3DAD2.
Comparative diagnosis. By its tegmen, Spinopygina acerfalx sp. nov. is very similar to Spinopygina camura sp. nov., S. quadracantha sp. nov. and S. uniceps (Hippa & Vilkamaa). All have the tegmen narrowed subapically, with flat apicolateral parts and with the parameres joining a shortly sclerotized rim apically. Spinopygina acerfalx is similar to S. edura sp. nov. and differs from all above-mentioned species in having only three, not four or five, gonostylar megasetae. Moreover, in the form of the gonostylus and in the arrangement of the gonostylar megasetae Spinopygina acerfalx resembles S. edura, but the latter is distinct in having the tegmen much smaller and more sclerotized basally and in having the parameres wide apart, joining the broadly sclerotized rim separately at the apex.
Etymology. The name is a Latin noun in apposition, formed from acer, sharp, and falx, sickle, referring to the sharp and curved gonostylar megasetae.
Material examined. Holotype: USA, Oregon, ♂; Benton County; Corvallis; 1460 SW Allen St.; 44.550860° N, 121.270189° W; 4 March–20 April 2015; S. Fitzgerald leg.; Malaise trap; in Euparal; MZH GE.1928. Paratypes: USA, Oregon, 2 ♂♂; Same collection data as for holotype; in Euparal; MZH GE.1929, GE.1945. Oregon, 1 ♂; same collection data as for holotype but 5 April–3 June 2015; in Euparal; MZH GE.1976. Oregon, 2 ♂♂; same collection data as for holotype but 11 December 2014–14 January 2015; used for DNA extraction (samples No. SCI80, SCI81); in Euparal; MZH GE.1978, GE.1979. Oregon, 1 ♂; Benton County; 5 miles up Woods Creek Road from jct Highway 20; 44.544° N, 123.50° W; 25 April–15 May 2014; S. Fitzgerald leg.; Malaise trap; mixed forest; in Euparal; MZH GE.1930. Oregon, 1 ♂; Benton County; Corvallis; 6.4 miles up Woods Creek Road from jct Highway 20; 3 March–12 April 2015; Malaise trap; fir/alder/maple; in Euparal; MZH GE.1946. Oregon, 1 ♂; Coos County; Seven Devils’ Road; 43.3109° N, 124.3484° W; 7 February–15 May 2016; E. Boyd leg; Malaise trap; in Euparal; MZH GE.1931.
Description. Male. Head. Face and antenna uniformly brown, maxillary palpus pale brown. Eye bridge 1–2 facets wide. Body of 4th antennal flagellomere 2.60–3.20× as long as subapically wide, the neck longer than wide, the longest setae longer than the width of flagellomere. Face with 5–9 dark long and short setae. Clypeus non-setose or with 1 dark seta. Maxillary palpus with 2 segments; 2nd segment reduced; 1st segment with 1 sharp seta, with large indistinct dorsal patch of sensilla, 2nd segment with 1–5 setae.
Thorax (
Figure 2B). Brown; setae dark. Anterior pronotum with 2 setae. Prothoracic episternum with 2–3 setae.
Wing (
Figure 3A). Fumose. Length 1.8–2.0 mm. Anal lobe small. Width/length 0.40–0.45. R1/R 0.75–0.85. c/w 0.75–0.80. stM slightly longer than M-fork, bM shorter than or as long as r-m, stCuA shorter or as long as bM. bM and r-m non-setose. Halter yellow, with long stalk.
Legs. Yellow, long. Fore femur slender. Fore tibial organ not impressed or proximally bordered, with small patch of a few setae. Fore tibial spur longer than tibial width. Length of fore basitarsomere/length of fore tibia 0.63.
Abdomen. Pale brown; setae dark, long and strong. Hypopygium (
Figure 4A) brown, as abdomen. Intergonocoxal area long, with short setosity. Gonocoxa longer than gonostylus; setae rather short, shorter towards medial margin. Gonostylus elongated, curved, the medial side strongly impressed; with a few elongated setae apically; without apical tooth, with 3 megasetae, 1 apical in ventral position, 1 subapical in dorsal position and 1 medial in ventral position, the megasetae long and strong, slightly recurved, with long basal bodies. Tegmen (
Figure 9A) slightly shorter than wide, truncate apically, sharply narrowed subapically; with parameres joining short sclerotized rim apically; with flat apicolateral parts. Aedeagal apodeme short, aedeagal teeth not detectable.
LSID urn:lsid:zoobank.org:act: B9089867-2FA5-4A03-92E4-5C99E60BD592.
Comparative diagnosis. In having the basal megaseta of the gonostylus strongly procurved, Spinopygina aurifera sp. nov. resembles S. camura sp. nov. and S. uniceps (Hippa and Vilkamaa) but by its form of the gonostylus and the tegmen, S. aurifera is not very similar to any other known species of the genus: the apicalmost megaseta is shifted to a subapical position and the two medial megasetae are in a lobe-like extension of the ventromedial margin of the gonostylus. The tegmen is characteristic in being strongly constricted at the middle, and in having the apex wide and roundish and in having the flat apicolateral parts small and triangular. Spinopygina aurifera differs from all other species of the genus in having a peculiar conical lobe apicodorsally in the gonocoxa.
Etymology. The name is a Latin noun in apposition, from the words auris, ear, and the suffix -fer, bearing, referring to the ear-like apicolateral parts of the tegmen.
Material examined. Holotype: USA, Oregon, ♂; Linn County; Hackleman Creek; 0.6 miles E of Tombstone Pass; 44.397501° N, 122.131401° E; 29 June–1 August 2016; S. Fitzgerald leg; Malaise trap; in Euparal; MZH GE.1932. Paratypes: USA, Oregon, 2 ♂♂; same collection data as for holotype; in Euparal; MZH GE.1933 and GE.1934. Oregon, 1 ♂; same collection data as for holotype; in Euparal; MZH GE.1970.
Description. Male. Head. Face and antenna uniformly brown, maxillary palpus pale brown. Eye bridge 1–2 facets wide. Body of 4th antennal flagellomere 2.25–2.55× as long as subapically wide, the neck longer than wide, the longest setae longer than the width of flagellomere. Face with 5–7 dark long and short setae. Clypeus non-setose or with 1 dark seta. Maxillary palpus (
Figure 1D) with 2 segments; 2nd segment reduced; 1st segment with 1(2) sharp setae, with an indistinct dorsal patch of sensilla, 2nd segment with 3–5 setae.
Thorax. Brown; setae dark. Anterior pronotum with 2 setae. Prothoracic episternum with 2–4 setae.
Wing (
Figure 3B). Fumose. Length 1.8–2.0 mm. Width/length 0.40–0.45. Anal lobe small. R1/R 0.75–0.90. c/w 0.70–0.75. stM slightly longer than M-fork, bM longer than r-m, stCuA as long as or shorter than bM. bM and r-m non-setose. Halter yellow, long.
Legs. Yellow, long. Fore femur slender. Fore tibial organ not impressed or proximally bordered, with small patch of a few setae. Fore tibial spur as long as tibial width. Length of fore basitarsomere/length of fore tibia 0.47.
Abdomen. Pale brown; setae dark, long and strong. Hypopygium (
Figure 4B) brown, as abdomen. Intergonocoxal area long, with short setosity. Gonocoxa wide, slightly longer than gonostylus; setosity normal, at medial margin shorter. Gonostylus elongated, slightly curved, strongly excavated medially; with a few elongated setae apically and medially; without apical tooth, with 4 megasetae, 1 subapical and 2 medial at ventral margin and 1 basally arising from medial excavation, the megasetae long and strong, nearly straight except the basalmost one procurved, all with basal bodies. Tegmen (
Figure 9B) longer than wide, curved apically, constricted laterally, with parameres ending wide apart at apex; with small triangular apicolateral parts. Aedeagal apodeme rather short, aedeagal teeth not detectable.
LSID urn:lsid:zoobank.org:act: 144A1DDD-712D-4B81-862A-735709641561.
Comparative diagnosis. Spinopygina camura sp. nov. is distinguished from all other species of the genus in having five instead of three or four gonostylar megasetae, the basalmost of which is strongly procurved (a paratype of S. quadracantha has five megasetae on one of its gonostyli) Spinopygina uniceps (Hippa and Vilkamaa) and S. aurifera sp. nov. also have a procurved basal megaseta on their gonostylus but they have only four megasetae. See also under S. acerfalx sp. nov. and S. quadracantha sp. nov.
Etymology. The name is a Latin adjective, camura, curved, referring to the curved basal megaseta of the gonostylus.
Material examined. Holotype: USA, Oregon, ♂; Benton County; 6.4 miles up Woods Creek Road from jct Highway 20; 6 March–12 April 2015; S. Fitzgerald leg.; Malaise trap; fir/alder/maple forest; in Euparal; MZH GE.1935. Paratype: USA, Oregon, 1 ♂; same collection data as for holotype; in Euparal; MZH GE.1936.
Description. Male. Head. Face and antenna uniformly brown, maxillary palpus pale brown. Eye bridge 2 facets wide. Body of 4th antennal flagellomere (
Figure 1C) 2.75–2.90× as long as subapically wide, the neck longer than wide, the longest setae longer than the width of flagellomere. Face with 8–10 dark long and short setae. Clypeus with 1 dark seta. Maxillary palpus with 2 segments; 2nd segment reduced; 1st segment with 1 sharp seta, with an indistinct dorsal patch of sensilla, 2nd segment with 2–3 setae.
Thorax. Brown; setae dark. Anterior pronotum with 2 setae. Prothoracic episternum with 2–4 setae.
Wing. Fumose. Length 2.0 mm. Width/length 0.40–0.45. Anal lobe small. R1/R 0.80. c/w 0.80. stM slightly longer than M-fork, bM shorter than or as long as r-m, stCuA shorter than bM. bM and r-m non-setose. Halter yellow, long.
Legs. Yellow, long. Fore femur slender. Fore tibial organ not impressed or proximally bordered, with small patch of a few setae. Fore tibial spur longer than tibial width. Length of fore basitarsomere/length of fore tibia 0.60.
Abdomen. Pale brown; setae dark, long and strong. Hypopygium (
Figure 5A) brown, as abdomen. Intergonocoxal area long, with short setosity. Gonocoxa longer than gonostylus; setae rather short, at medial margin shorter. Gonostylus elongated, curved laterally, strongly impressed medially; with a few elongated setae apically; without apical tooth, with 5 megasetae, 1 apical in ventral position, 1 subapical in dorsal position, 2 medially in ventral position and 1 basally arising from the medial excavation, megasetae long and strong, slightly recurved, the basalmost megaseta strongly procurved, all with long basal bodies. Tegmen (
Figure 9C) slightly longer than wide, truncate apically, roundish laterally, with parameres joining to short sclerotized rim apically; with flat apicolateral parts. Aedeagal apodeme short, aedeagal teeth not detectable.
LSID urn:lsid:zoobank.org:act: AE559992-0563-426C-B08C-7CEB91256997.
Etymology. The name is a Latin adjective, edura, hard, referring to the strongly sclerotized tegmen.
Comparative diagnosis. By its gonostylus with a sharp apex and three megasetae Spinopygina edura sp. nov. is very similar to S. acerfalx sp. nov. although the former has the medial megaseta slightly closer to apex of the gonostylus. Spinopygina edura differs from all the species of the genus in its small and more sclerotized tegmen with concave parameres joining a broadly sclerotized rim apically.
Material examined. Holotype: USA, Oregon, ♂; Benton County; Corvallis; 1460 SW Allen St.; 44.550860° N, 121.270189° W; ex larva, 19 October 2014, emerged 14 November 2014; S. Fitzgerald; in Euparal; MZH GE.1937.
Description. Male. Head. Face and antenna uniformly brown, maxillary palpus pale brown. Eye bridge 1–2 facets wide. Antennae in poor condition in the specimen studied, the neck longer than wide. Face with 10 dark long and short setae. Clypeus with 1 dark seta. Maxillary palpus with 2 segments; 2nd segment reduced; 1st segment with 1 sharp seta, with an indistinct dorsal patch of sensilla, 2nd segment with 3–4 setae.
Thorax. Brown; setae dark. Anterior pronotum with 1 seta. Prothoracic episternum with 3 setae.
Wing. Fumose. Length 2.2 mm. Width/length 0.40. Anal lobe small. R1/R 0.70. c/w 0.80. stM slightly longer than M-fork, bM shorter than r-m, stCuA shorter than bM. bM and r-m non-setose. Halter yellow, long.
Legs. Yellow, long. Fore femur slender. Fore tibial organ not impressed or proximally bordered, with small patch of a few setae. Fore tibial spur slightly longer than tibial width. Length of fore basitarsomere/length of fore tibia 0.60.
Abdomen. Pale brown; setae dark, long and strong. Hypopygium (
Figure 5B) brown, as abdomen. Intergonocoxal area long, with short setosity. Gonocoxa longer than gonostylus; setae rather short, at medial margin shorter. Gonostylus elongated, curved, the medial side impressed; with a few elongated setae apically; without apical tooth, with 3 megasetae, 1 at very apex in dorsal position, 1 subapically in ventral position and 1 medially in ventral position; the megasetae long and strong, slightly recurved, with long basal bodies. Tegmen (
Figure 9D) slightly shorter than wide, truncate apically, concave laterally, with strong basolateral sclerotizations, with concave parameres joining separately broad sclerotized rim apically; with large flat apicolateral parts. Aedeagal apodeme short, aedeagal teeth not detectable.
LSID urn:lsid:zoobank.org:act: E7434F57-4965-4501-B26C-3A0C49D11302.
Comparative diagnosis. In having four gonostylar megasetae all at the apical half of half of the gonostylus and in having a wide tegmen, Spinopygina peltata sp. nov. resembles S. plena sp. nov. but differs in having the tegmen constricted subapically and with acuminate flat apicolateral parts. Moreover, Spinopygina plena has the dorsal side of its gonostylus greatly enlarged.
Etymology. The name is formed from the Latin word pelta, shield, referring to the wide tegmen.
Material examined. Holotype: USA, Oregon, ♂; Benton County; Corvallis; 1460 SW Allen St.; 44.550860° N, 121.270189° W; 4 March–20 April 2015; S. Fitzgerald leg.; Malaise trap; in Euparal; MZH GE.1938. Paratypes: USA, Oregon, 6 ♂♂; same collection data as for holotype; MZH GE.1939, GE.1947–GE.1951. Oregon, 1 ♂; same collection data; in Euparal; MZH GE.1971. Oregon, 1 ♂; same collection data but 11 December 2014–15 January 2015; in Euparal; MZH GE.1980. Oregon, 2 ♂♂; same data as previous, used for DNA extraction (samples No. SCI82, SCI83), MZH GE.1973, GE.1974. Oregon, 1 ♂; Coos County; Seven Devils’ Road; 43.3109° N, 124.3484° W; 7 February–15 May 2016; E. Boyd leg; Malaise trap; in Euparal; MZH GE.1940. Oregon, 1 ♂; Coos County; Charleston; Seven Devils’ Road; 43.3132° N, 124.3485° W; 7 February–15 May 2016; S. Fitzgerald leg; Malaise trap; in Euparal; MZH GE.1972.
Description. Male. Head. Face brown, darker than antenna, maxillary palpus pale brown. Eye bridge 2 facets wide. Body of 4th antennal flagellomere 2.75–3.2× (
Figure 1B) as long as subapically wide, the neck longer than wide, the longest setae longer than the width of flagellomere. Face with 5–9 dark long and short setae. Clypeus with 1 dark seta. Maxillary palpus with 2 segments; 2nd segment reduced; 1st segment with 1–2 sharp setae, with an indistinct dorsal patch of sensilla, 2nd segment with 2 setae.
Thorax. Brown; setae dark. Anterior pronotum with 1–2 setae. Prothoracic episternum with 2–5 setae.
Wing. Fumose. Length 1.7–2.1 mm. Width/length 0.40–0.45. Anal lobe small. R1/R 0.80–1.15. c/w 0.75–0.90. stM slightly longer than M-fork, bM longer than or as long as r-m, stCuA shorter than r-m. bM and r-m non-setose. Halter yellow, long.
Legs (
Figure 3D). Yellow, long. Fore femur slender. Fore tibial organ not impressed or proximally bordered, with small patch of a few setae. Fore tibial spur longer than tibial width. Length of fore basitarsomere/length of fore tibia 0.58.
Abdomen. Pale brown; setae dark, long and strong. Hypopygium (
Figure 6A) brown, as abdomen. Intergonocoxal area long, with short setosity. Gonocoxa rather narrow, much longer than gonostylus; setae rather short, at medial margin shorter. Gonostylus elongated, rather straight, the medial side impressed; with a few elongated setae apically; without apical tooth, with 4 megasetae, 1 at apex in dorsal position, 1 subapically in ventral position and 2 slightly more mediad in dorsal and ventral positions, the megasetae long and strong, nearly straight, with short basal bodies. Tegmen (
Figure 9E) shorter than wide, hyaline and slightly produced apically, roundish basolaterally and slightly constricted subapically, with parameres ending separately wide apart at apex; with flat apicolateral parts. Aedeagal apodeme long, aedeagal teeth not detectable.
LSID urn:lsid:zoobank.org:act: 8F5CAF1D-108C-4008-A048-35CA415C8E19.
Comparative diagnosis. Spinopygina plena sp. nov. resembles S. peltata sp. nov., see under the latter.
Etymology. The name is a Latin adjective, plena, plump, referring to the tumid gonostylus.
Material examined. Holotype: USA, California, ♂; Los Angeles County; 9 km N of La Canada; 34°25′ N, 118°19′ W; 900 m; 14 December 1994; B.V. Brown leg.; Malaise trap; ravine with oak forest; in Euparal; MZH GE.1941. Paratypes: USA, California, 2 ♂♂; same collection data as for holotype; in Euparal; MZH GE.1942, GE.1943. California, 1 ♂; same collection data as for holotype but 20 January–2 February 1995; B.V. Brown leg.; in Euparal; MZH GE.1981.
Description. Male. Head (
Figure 1A). Face brown, darker than antenna, maxillary palpus pale brown. Eye bridge 2 facets wide. Body of 4th antennal flagellomere 2.3–2.9× as long as subapically wide, the neck longer than wide, the longest setae longer than the width of flagellomere. Face with 6–10 dark long and short setae. Clypeus non-setose or with 1 dark seta. Maxillary palpus (
Figure 1E) with 2 segments; 2nd segment reduced; 1st segment with 1–2 sharp setae, with an indistinct dorsal patch of sensilla, 2nd segment with 1–2 setae.
Thorax. Brown; setae dark. Anterior pronotum with 2–4 setae. Prothoracic episternum with 3–5 setae.
Wing. Fumose. Length 1.6–1.7 mm. Width/length 0.40. Anal lobe small. R1/R 0.80–0.95. c/w 0.75–0.80. stM slightly longer than M-fork, bM longer than r-m, stCuA shorter than r-m. bM and r-m non-setose. Halter yellow, long.
Legs. Yellow, long. Fore femur slender. Fore tibial organ (
Figure 3E) not impressed or proximally bordered, with indistinct patch of a few setae. Fore tibial spur longer than tibial width. Length of fore basitarsomere/length of fore tibia 0.50.
Abdomen. Pale brown; setae dark, long and strong. Hypopygium (
Figure 6B) brown, as abdomen. Intergonocoxal area long, with short setosity. Gonocoxa longer than gonostylus; setosity rather sparse, at medial margin shorter. Gonostylus wide, curved laterally, strongly impressed medially, distinctly lobe-like produced dorsomedially, with a few elongated setae apically without apical tooth, with 4 megasetae, 1 at the very apex, 2 subapically, 1 medially, the megasetae rather long and strong, nearly straight, with distinct basal bodies. Tegmen (
Figure 9F) slightly shorter than wide, truncate apically, roundish laterally, with lateral sclerotizations (parameres) joining wide apart at apex; with large flat apicolateral parts. Aedeagal apodeme short, aedeagal teeth not detectable.
LSID urn:lsid:zoobank.org:act: E8ADE6BD-FF9F-4F09-931A-3DAD514FDCEA.
Comparative diagnosis. In having all gonostylar megasetae long and slightly recurved,
Spinopygina quadracantha sp. nov. resembles
S. acerfalx sp. nov. and
S. edura sp. nov. but differs in having four instead of three megasetae. However, two of the studied specimens have an additional procurved megaseta at the basal part in one of their gonostyli (
Figure 7B). These gonostyli with 5 megasetae are difficult to distinguish from those of
Spinopygina camura sp. nov. but the tegmen of
S. quadracantha is bigger with larger apicolateral flat parts. A large tegmen with convex parameres joining into a short sclerotized rim apically is fairly similar to that of
S. acerfalx and differs distinctly from that of
S. edura sp. nov. See also under the latter.
Etymology. The name is a Latin noun in apposition, from the words quadrus, four, and acantha, spine, referring to the four gonostylar megasetae.
Material examined. Holotype: Canada, British Columbia, ♂; Vancouver Island; Rocky Point; 19 March 1995; N. Winchester leg.; Malaise trap; in Euparal; MZH GE.1944.
Paratypes: Canada, British Columbia, 7 ♂♂; same collection data as for holotype but 29 October 1995; MZH GE.1952 to GE.1958. British Columbia, 2 ♂♂; same collection data as previous; in Euparal, MZH GE.1974, GE.1975. British Columbia, 2 ♂♂; same collection data as previous but 15 January 1995; in Euparal; MZH GE.1959, GE.1960.
Description. Male. Head. Face and antenna uniformly brown, maxillary palpus pale brown. Eye bridge 2 facets wide. Body of 4th antennal flagellomere 2.40–3× as long as subapically wide, the neck longer than wide, the longest setae longer than the width of flagellomere. Face with 4–10 dark long and short setae. Clypeus with 1 dark seta. Maxillary palpus with 2 segments; 2nd segment reduced; 1st segment with 1–2 sharp setae, with an indistinct dorsal patch of sensilla, 2nd segment with 1–3 setae.
Thorax. Brown; setae dark. Anterior pronotum with 2 setae. Prothoracic episternum with 3 setae.
Wing. Fumose. Length 1.8–2.1 mm. Width/length 0.40–0.45. Anal lobe small. R1/R 0.85–1.0. c/w 0.70–0.90. stM slightly longer, as long as or shorter than M-fork, bM longer or shorter than r-m, stCuA shortest. bM and r-m non-setose. Halter yellow, long.
Legs. Yellow, long. Fore femur slender. Fore tibial organ with weak vestiture, forming an indistinct patch of few setae. Fore tibial spur longer than tibial width. Length of fore basitarsomere/length of fore tibia 0.60.
Abdomen. Pale brown; setae dark, long and strong. Hypopygium (
Figure 7) brown, as abdomen. Intergonocoxal area long, with short setosity. Gonocoxa longer than gonostylus; setae rather short, at medial margin shorter. Gonostylus elongated, curved, strongly impressed medially; with a few elongated setae apically and medially; without apical tooth, with 4 (rarely 5) megasetae, 1 at the very apex, 1 subapically and 2 medially wide apart, the megasetae long and strong, slightly recurved, with long basal bodies. Tegmen (
Figure 9D) longer than wide, truncate apically, constricted laterally, with parameres joining to short sclerotized rim apically; with large flat apicolateral parts. Aedeagal apodeme short, aedeagal teeth not detectable.
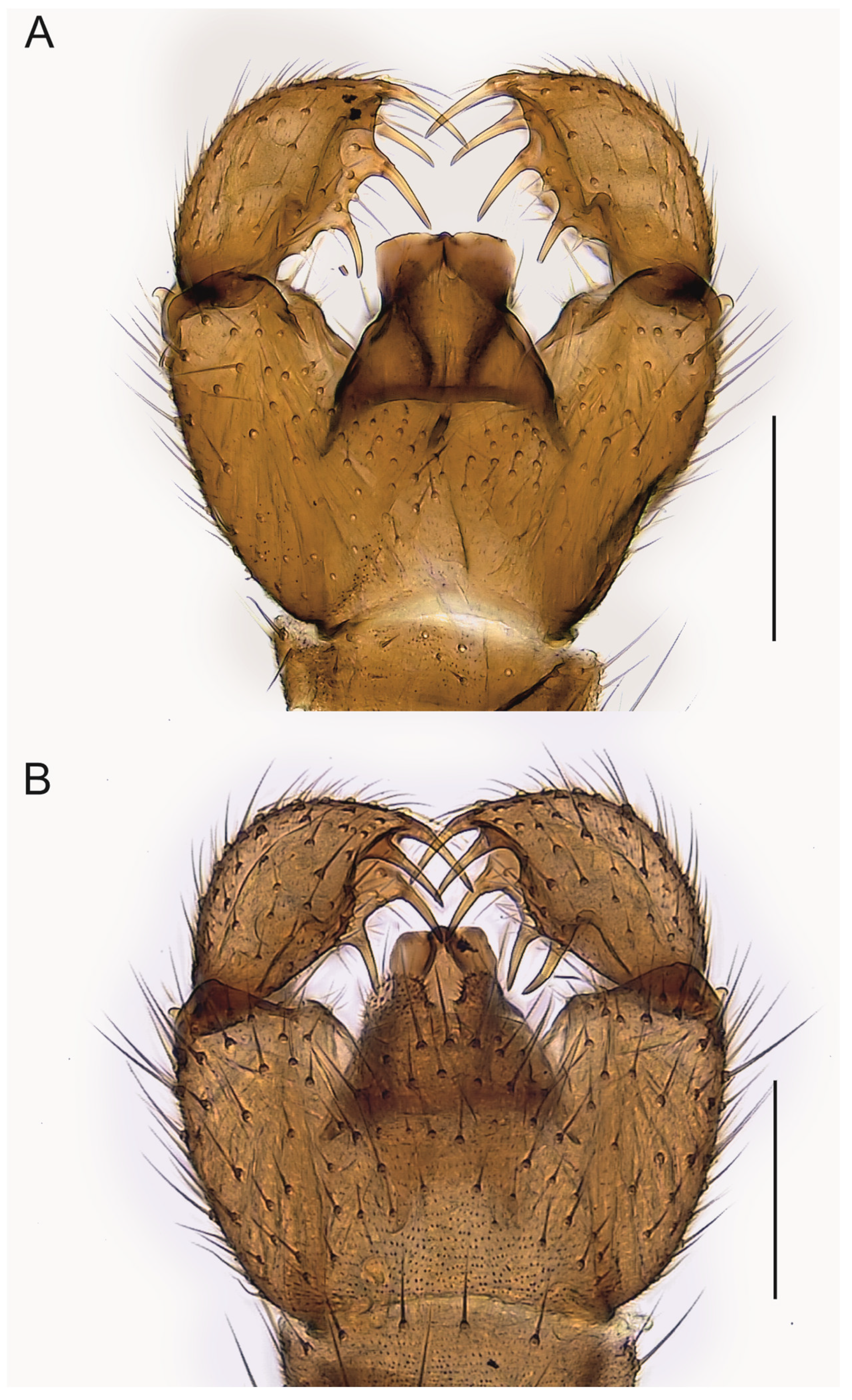
Spinopygina uniceps (Hippa & Vilkamaa, 1994) comb. nov.
Figure 2A,
Figure 8 and
Figure 9H.
Camptochaeta uniceps Hippa & Vilkamaa, 1994: 56, Figure 30A,B.
Corynoptera uniceps Mohrig et al. (2013): 195.
Short redescription. Male. Wing length 1.6–2.2 mm (
Figure 2A). 1st segment of maxillary palpus with 1 seta, 2nd segment of with 2–5 setae. Gonostylus (
Figure 8) widest medially, strongly narrowed towards apex, strongly excavated medially, apex with rather short apical megaseta; with two medial megasetae at dorsomedial margin, basal megaseta with long basal body arising from medial excavation. Tegmen (
Figure 9H) with flat apicolateral parts, parameres joining short sclerotized rim apically.
Comparative diagnosis. By its form of gonostylus with four megasetae and by the tegmen with flat apicolateral parts and the parameres joining a short apical rim,
Spinopygina uniceps (Hippa & Vilkamaa, 1994) most resembles
S. quadracantha sp. nov. but differs in having a shorter apical megaseta of the gonostylus, in having the basal megaseta distinctly procurved and with a long basal body and placed in the medial excavation instead at the ventromedial margin of the gonostylus.
Spinopygina uniceps has the apex of tegmen narrower and with smaller flat apicolateral parts (
Figure 7 and
Figure 8). See also under
Spinopygina acerfalx.
Material examined. Holotype: Canada, British Columbia, ♂; Vancouver Island; Upper Carmanah Valley; 30 July 1991; N. Winchester leg.; Malaise trap; forest floor; in Euparal; CNC. Paratypes. Canada, British Columbia, 3 ♂♂; same collection data as for holotype; in Euparal; MZH GE.1116 to GE.1118. British Columbia, 1 ♂; same collection data as for holotype; in Euparal; RBCM.
New material. Canada, British Columbia, 3 ♂♂; Vancouver Island; Upper Carmanah Valley; 10–29 September 1991; N. Winchester leg; Malaise trap, forest floor; in Euparal; MZH GE.1961 to GE.1963. British Columbia, 5 ♂♂; same data as previous but 17–26 October 1991; in Euparal; MZH GE.1964 to GE.1968. British Columbia, 1 ♂; same collection data as previous; in ethanol; MZH GE.1969. British Columbia, 1 ♂; same collection data as previous; in Euparal; GE.1977.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}