
Melinacidin-Producing Acrostalagmus luteoalbus, a Major Constituent of Mixed Mycobiota Contaminating Insulation Material in an Outdoor Wall

 , ,
, ,
Abstract
:1. Introduction
2. Results
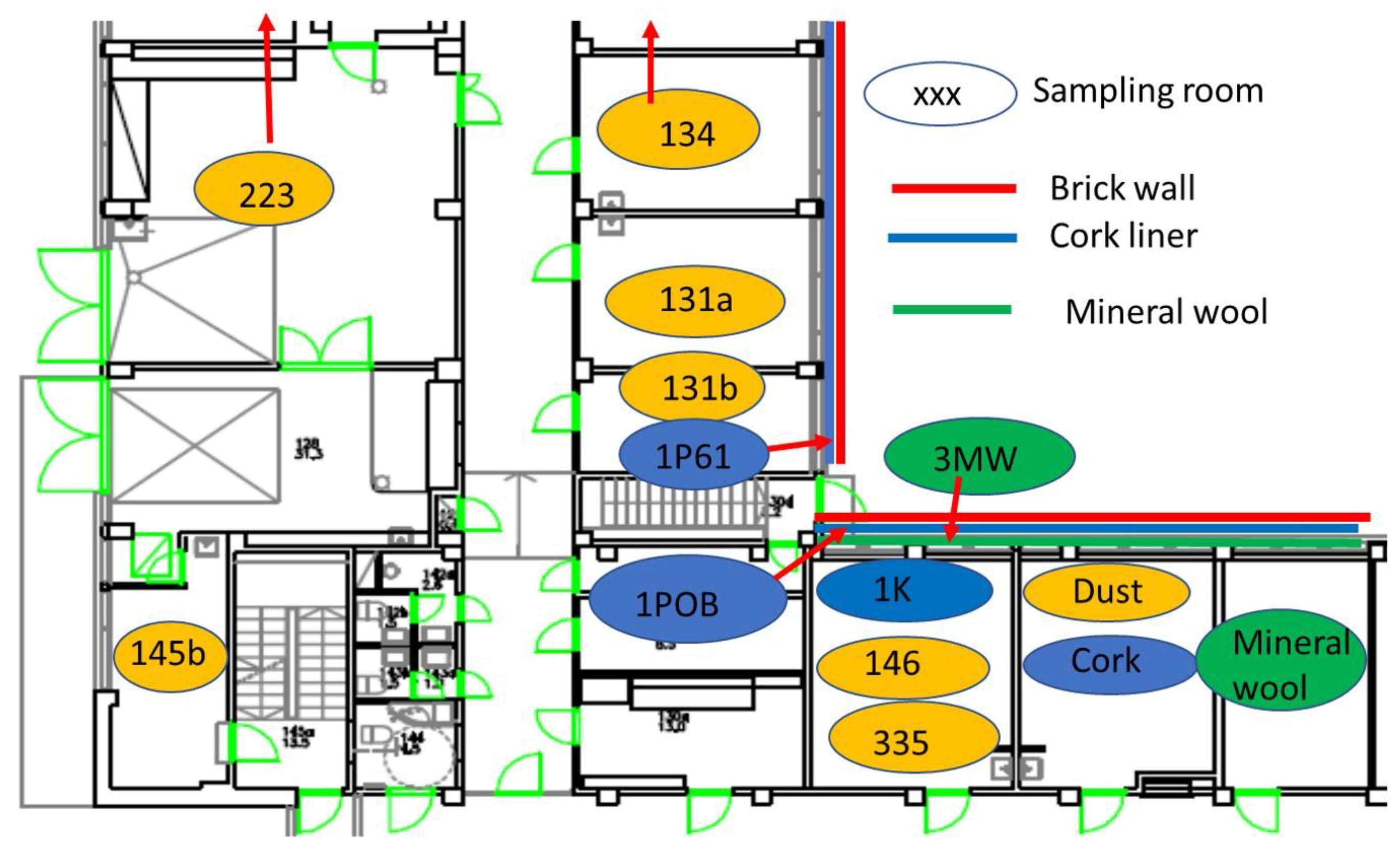
2.1. Building Inspection Connected to Five Problematic Rooms
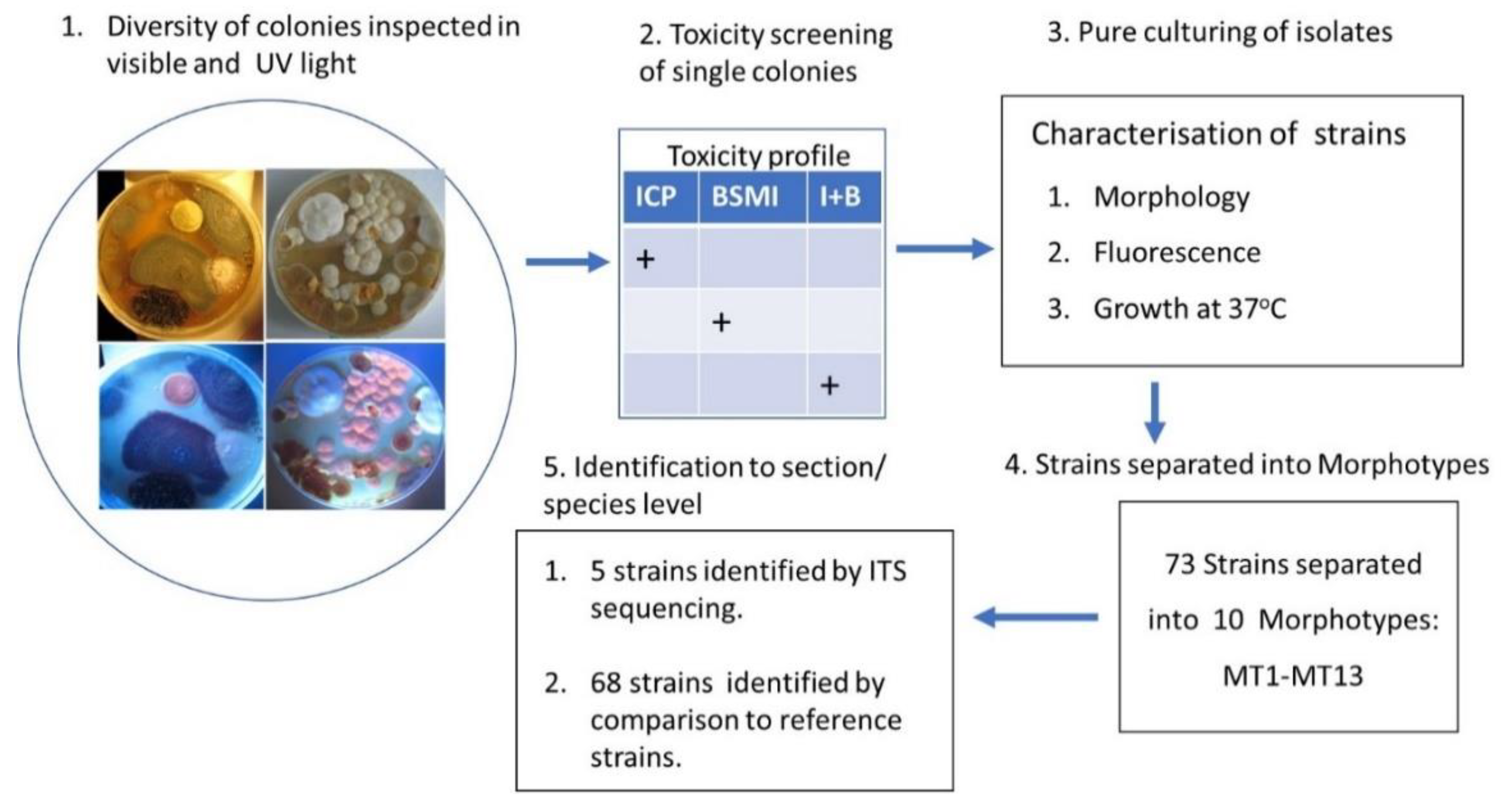
2.2. Diversity Tracking of Molds Cultivated from Wall Structures
2.3. Cultured Settled Dust from Problematic Rooms Revealed Five Major Morphotypes
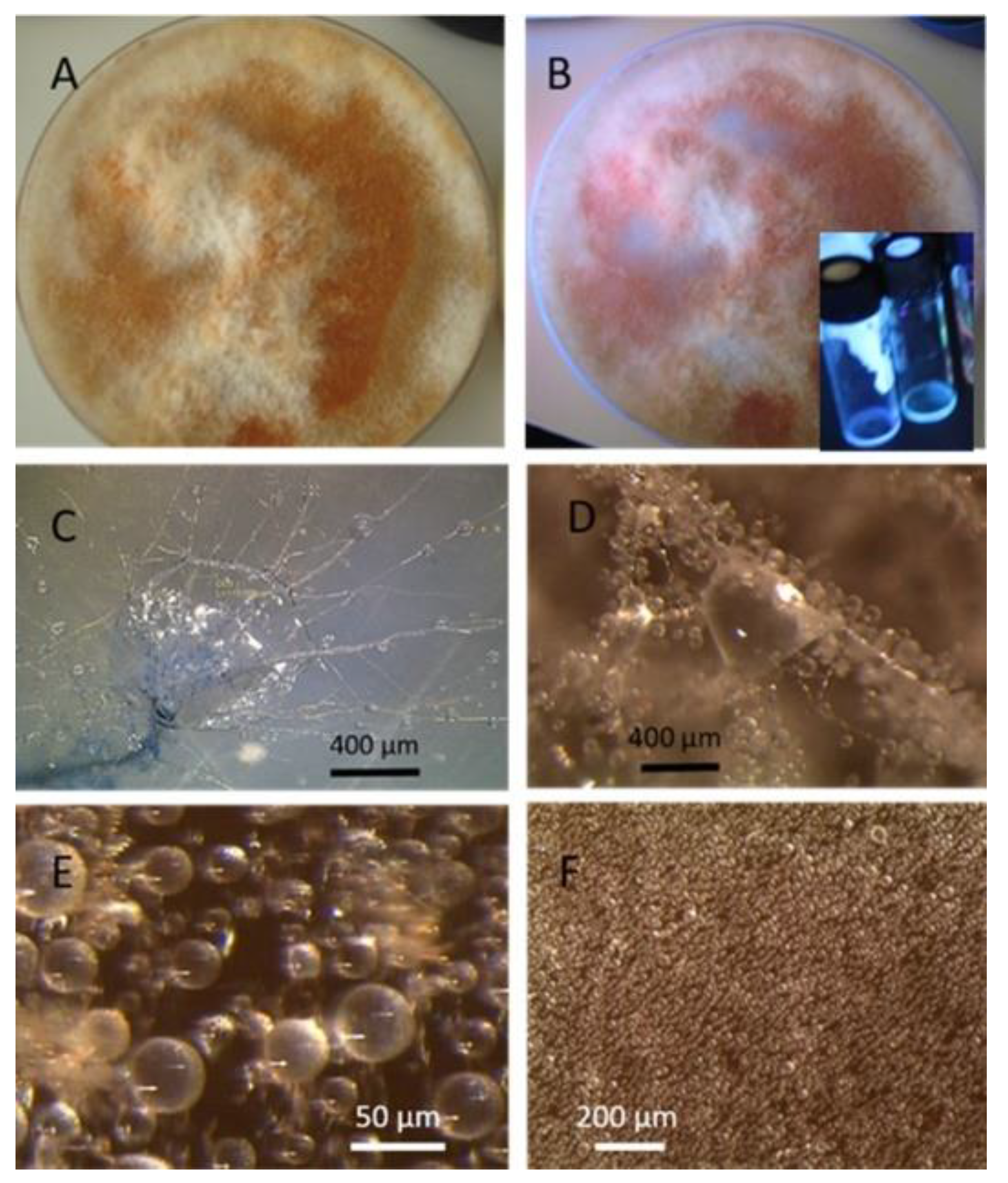
2.4. Characterization of Acrostalagmus luteoalbus Strain POB8
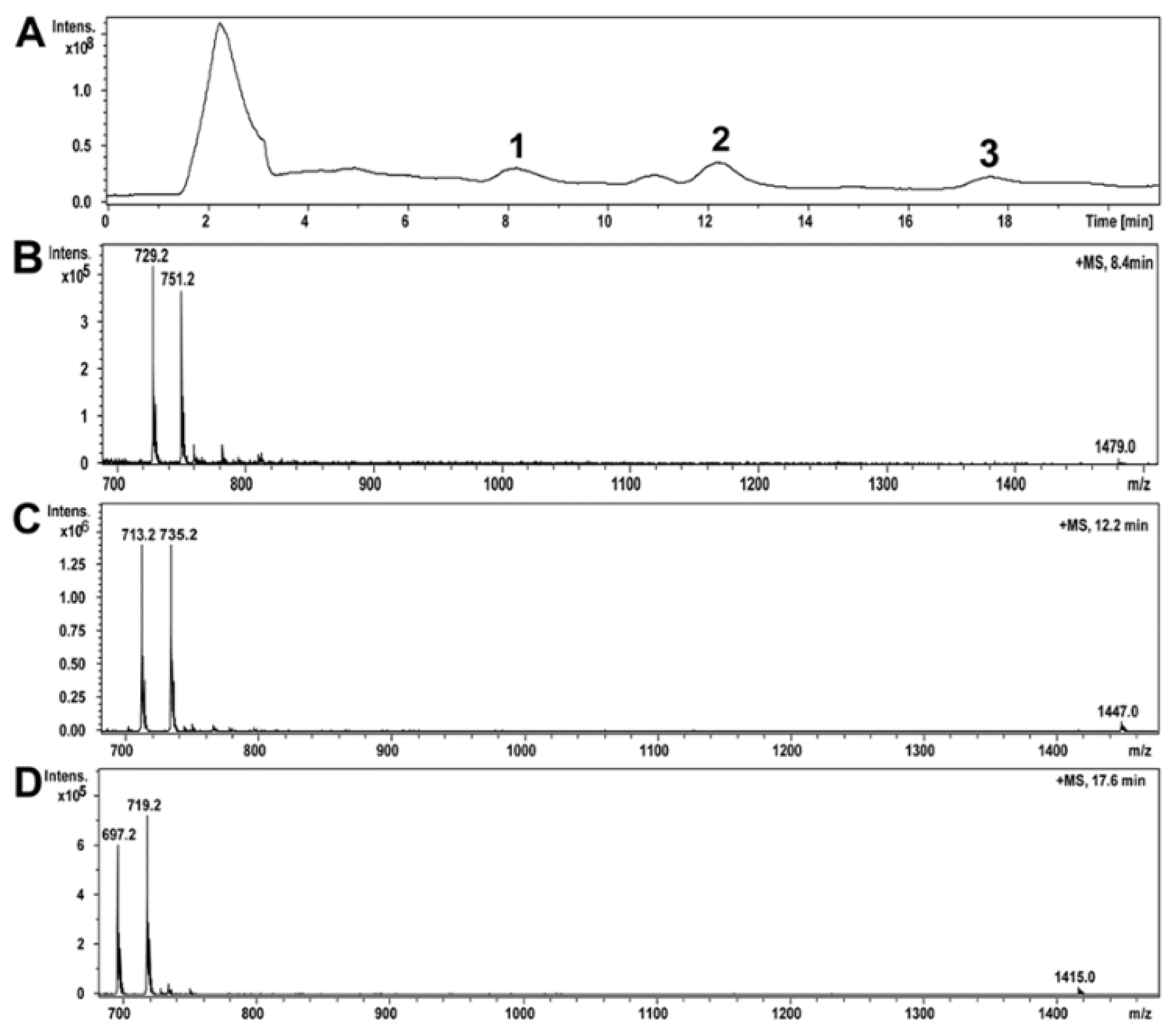
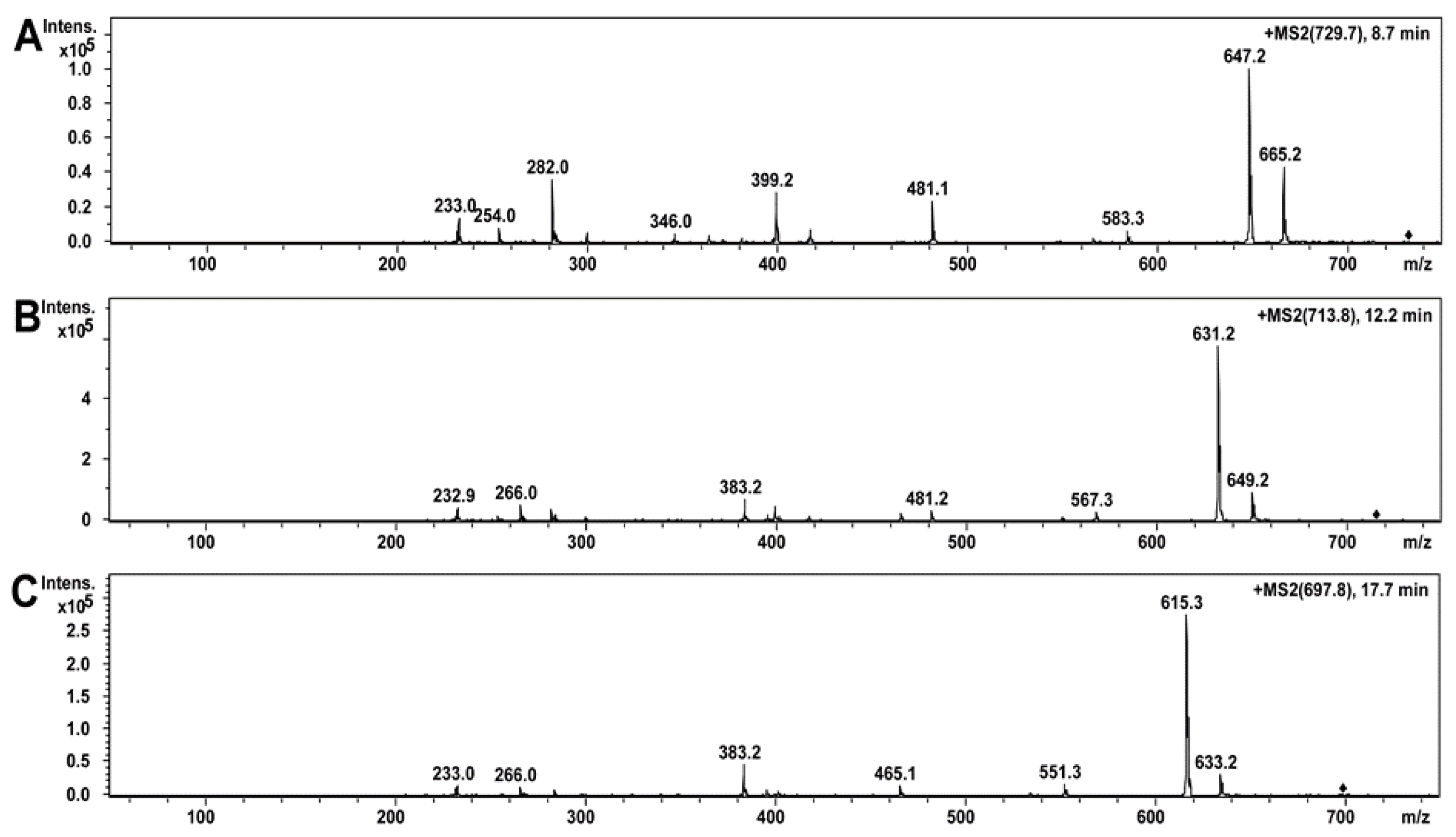
2.5. Compounds of Toxic A. luteoalbus POB8 Ethanol Extract Were Identified as Melinacidins II, III, and IV
3. Discussion
4. Materials and Methods
4.1. A Public Building from the 1960s Investigated for Mold Contamination
4.2. Collection of Samples and Diversity Tracking of Microbial Constituents in Building Materials and Settled Dust
4.3. Experimental Design for Cultivation and Identification of Microbial Isolates
4.4. Toxicity Assays
4.4.1. Rapid Screening Test of Single Colonies
4.4.2. Toxicity Assays for Ethanol-Extracted Pure Fungal Cultures
4.5. Chemical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andersen, B.; Frisvad, J.C.; Dunn, R.R.; Thrane, U. A pilot study on baseline fungi and moisture indicator fungi in Danish homes. J. Fungi 2021, 7, 71. [Google Scholar] [CrossRef]
- Shukla, S.; Budden, K.; Neal, R.; Hansbro, P. Microbiome effects on immunity, health and disease in the lung. Clin. Translat. Immunol. 2017, 6, e133. [Google Scholar] [CrossRef]
- Horve, P.; Lloyd, S.; Mhuireach, G.; Dietz, L.; Fretz, M.; MacCrone, G.; Van Den Wymelenberg, K.; Ishaq, S.L. Building upon current knowledge and techniques of indoor microbiology to construct the next era of theory into microorganisms, health, and the built environment. J. Expo. Sci. Environ. Epidemiol. 2020, 30, 219–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, M.H.Y.; Lee, P.K.H. The roles of the outdoors and occupants in contributing to a potential pan-microbiome of the built environment: A review. Microbiome 2016, 4, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haahtela, T.; Alenius, H.; Lehtimäki, J.; Sinkkonen, A.; Fyhrquist, N.; Hyöty, H.; Ruokolainen, L.; Mäkelä, M. Immunological resilience and biodiversity for prevention. Allergy 2021. [Google Scholar] [CrossRef]
- Andersson, M.A.; Salo, J.; Kedves, O.; Kredics, L.; Druzhinina, I.; Kurnitski, J.; Salonen, H. Bioreactivity, guttation and agents influencing surface tension of water emitted by actively growing indoor mould isolates. Microorganisms 2020, 8, 1940. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Prussin, A.; Marr, L.; Vikesland, P.; Edwards, M.A.; Pruden, A. Factors shaping the human exposome in the built environment: Opportunities for engineering control. Environ. Sci. Technol. 2017, 51, 7759–7774. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Yang, Z.; Hu, D.; Cao, L.; He, Q. Understanding building-occupant-microbiome interactions toward healthy built environments: A review. Environ. Sci. Eng. 2021, 15, 65. [Google Scholar] [CrossRef]
- Bloom, E.; Nyman, E.; Must, A.; Pehrson, C.; Larsson, L. Mold and mycotoxins in indoor environments—a survey in water damaged buildings. J. Occup. Environ. Hyg. 2009, 6, 671–678. [Google Scholar] [CrossRef]
- Gravesen, S.; Nielsen, P.A.; Iversen, R.; Nielsen, K.F. Microfungal contamination of damp buildings—Examples of risk constructions and risk materials. Environ. Health Perspect. 1999, 107, 505–508. [Google Scholar] [CrossRef]
- Jarvis, B.; Miller, J. Mycotoxins as harmful indoor air contaminants. Appl. Microbiol. Biotechnol. 2005, 66, 367–372. [Google Scholar] [CrossRef]
- Mendell, M.J.; Kumagai, K. Observation-based metrics for residential dampness and mold with dose-response relationships to health: A review. Indoor Air 2017, 27, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D.; McMullin, D.R. Fungal secondary metabolites as harmful indoor air contaminants: 10 years on. Appl. Microbiol. Biotechnol. 2014, 98, 9953–9966. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.F. Mold Growth on Building Materials. Secondary Metabolites, Mycotoxins and Biomarkers. Ph.D. Thesis, Technical University of Denmark, Lyngby, Denmark, 2002. [Google Scholar]
- Piontek, M.; Łuszczynska, K. Testing the toxicity of Stachybotrys chartarum in indoor environments—A case study. Energies 2021, 14, 1602. [Google Scholar] [CrossRef]
- Kankkunen, P.; Rintahaka, J.; Aalto, A.; Leino, M.; Majuri, M.L.; Alenius, H.; Wolff, H.; Matikainen, S. Trichothecene mycotoxins activate inflammatory response in human macrophages. J. Immunol. 2009, 182, 6418–6425. [Google Scholar] [CrossRef] [Green Version]
- Kankkunen, P.; Valimaki, E.; Rintahaka, J.; Palomäki, J.; Nyman, T.; Alenius, H.; Wolff, H.; Matikainen, S. Trichothecene mycotoxins activate NLRP3 inflammasome through a P2X7 receptor and Src tyrosine kinase dependent pathway. Hum. Immunol. 2014, 75, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Korkalainen, M.; Taubel, M.; Naarala, J.; Kirjavainen, P.; Kistinen, A.; Hyvärinen, A.; Komulainen, H.; Viluksela, M. Synergistic proinflammatory interactions of microbial toxins and structural components characteristic to moisture-damaged buildings. Indoor Air 2017, 27, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Annila, P.J.; Hellemaa, M.; Pakkala, T.A.; Lahdensivu, J.; Suonketo, J.; Pentti, M. Extent of moisture and mould damage in structures of public buildings. Case Stud. Constr. Mater. 2017, 6, 103–108. [Google Scholar] [CrossRef]
- Andersson, M.A.; Nikulin, M.; Köljalg, U.; Andersson, M.C.; Rainey, F.; Reijula, K.; Hintikka, E.L.; Salkinoja-Salonen, M. Bacteria, molds, and toxins in water-damaged building materials. Appl. Environ. Microbiol. 1997, 63, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.A.; Mikkola, R.; Kroppenstedt, R.M.; Rainey, F.A.; Salkinoja-Salonen, M.S. The mitochondrial toxin produced by Streptomyces griseus strains isolated from an indoor environment is valinomycin. Appl. Environ. Microbiol. 1998, 64, 4767–4773. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.A.; Mikkola, R.; Raulio, M.; Kredics, L.; Maijala, P.; Salkinoja-Salonen, M.S. Acrebol, a novel toxic peptaibol produced by an Acremonium exuviarum indoor isolate. J. Appl. Microbiol. 2009, 106, 909–923. [Google Scholar] [CrossRef] [PubMed]
- Bornehag, C.-G.; Blomquist, G.; Gyntelberg, F.; Jarvholm, B.; Malmberg, P.; Nordvall, L.; Nielsen, A.; Pershagen, G.; Sundell, J. Dampness in buildings and health, Nordic interdisciplinary review of the scientific evidence on associations between exposure to “dampness” in buildings and health effects (NORDDAMP). Indoor Air 2001, 11, 72–86. [Google Scholar] [CrossRef]
- Bornehag, C.-G.; Sundell, J.; Bonini, S.; Custovic, A.; Malmberg, P.; Skerfving, S.; Sigsgaard, T.; Verhoeff, A. Dampness in buildings as a risk factor for health effects, EUROEXPO: A multidisciplinary review of the literature (1998–2000) on dampness and mite exposure in buildings and health effects. Indoor Air 2004, 14, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, R.; Andersson, M.A.; Grigoriev, P.; Teplova, V.V.; Saris, N.-E.L.; Rainey, F.A.; Salkinoja-Salonen, M.S. Bacillus amyloliquefaciens strains isolated from moisture-damaged buildings produced surfactin and a substance toxic to mammalian cells. Arch. Microbiol. 2004, 181, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, R.; Andersson, M.A.; Teplova, V.; Grigoriev, P.; Kuehn, T.; Loss, S.; Tsitko, I.; Apetroaie, C.; Saris, N.-E.L.; Veijalainen, P.; et al. Amylosin from Bacillus amyloliquefaciens, a K+ and Na+ channel-forming toxic peptide containing a polyene structure. Toxicon 2007, 49, 1158–1171. [Google Scholar] [CrossRef]
- Nielsen, K.F.; Smedsgaard, J. Fungal metabolite screening: Database of 474 mycotoxins and fungal metabolites for dereplication by standardised liquid chromatography-UV-mass spectrometry methodology. J. Chromatogr. A 2003, 100, 111–136. [Google Scholar] [CrossRef]
- Paananen, A.; Järvinen, K.; Sareneva, T.; Salkinoja-Salonen, M.S.; Timonen, T.; Hölttä, E. Valinomycin-induced apoptosis of human NK cells is predominantly caspase independent. Toxicology 2005, 212, 37–45. [Google Scholar] [CrossRef]
- Peltola, J.; Andersson, M.A.; Haahtela, T.; Mussalo-Rauhamaa, H.; Rainey, F.A.; Kroppenstedt, R.M.; Samson, R.A.; Salkinoja-Salonen, M.S. Toxic-metabolite-producing bacteria and fungus in an indoor environment. Appl. Environ. Microbiol. 2001, 67, 3269–3274. [Google Scholar] [CrossRef] [Green Version]
- Peltola, J.; Andersson, M.A.; Kämpfer, P.; Auling, G.; Kroppenstedt, R.M.; Busse, H.-J.; Salkinoja-Salonen, M.S.; Rainey, F.A. Isolation of toxigenic Nocardiopsis strains fromindoor environments and description of two new Nocardiopsis species, N. exhalans sp. nov. and N. umidischolae sp. nov. Appl. Environ. Microbiol. 2001, 67, 4293–4304. [Google Scholar] [CrossRef] [Green Version]
- Rasimus-Sahari, S.; Teplova, V.V.; Andersson, M.A.; Mikkola, R.; Kankkunen, P.; Matikainen, S.; Gahmberg, C.G.; Andersson, L.C.; Salkinoja-Salonen, M. The peptide toxin amylosin of Bacillus amyloliquefaciens from moisture damaged buildings is immunotoxic, induces potassium efflux from mammalian cells, and has antimicrobial activity. Appl. Environ. Microbiol. 2015, 81, 2939–2949. [Google Scholar] [CrossRef] [Green Version]
- Tuomi, T.; Reijula, K.; Johnsson, T.; Hemminki, K.; Hintikka, E.L.; Lindroos, O.; Kalso, S.; Koukila-Kähkölä, P.; Mussalo-Rauhamaa, H.; Haahtela, T. Mycotoxins in crude building materials from water-damaged buildings. Appl. Environ. Microbiol. 2000, 66, 1899–1904. [Google Scholar] [CrossRef] [Green Version]
- Wahab, S.N.A.; Mohammed, N.I.; Khamidi, M.F.; Jamaluddin, N. Qualitative assessment of mould growth for higher education library building in Malaysia. Procedia Soc. Behav. Sci. 2015, 170, 252–261. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, G.L.V.; Oliveira, C.N.S.; Pinzan, C.F.; de Salis, L.V.V.; Cardoso, C.R.B. Microbiota modulation of the gut-lung axis in COVID-19. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Fitzgibbon, G.; Mills, K. The microbiota and immune-mediated diseases: Opportunities for therapeutic intervention. Eur. J. Immunol. 2020, 50, 326–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganal, S.C.; Sanos, S.L.; Kallfass, C.; Oberle, K.; Johner, C.; Kirschning, C.; Lienenklaus, S.; Weiss, S.; Staeheli, P.; Aichele, P.; et al. Priming of natural killer cells by nonmucosal mononuclear phagocytes requires instructive signals from commensal microbiota. Immunity 2012, 37, 171–186. [Google Scholar] [CrossRef] [Green Version]
- Haahtela, T.; Laatikainen, T.; Alenius, H. Hunt for the origin of allergy—Comparing the Finnish and Russian Karelia. Clin. Exp. Allergy 2015, 45, 891–901. [Google Scholar] [CrossRef]
- Haahtela, T.; Von Hertzen, L.; Anto, J.M.; Bai, C.; Baigenzhin, A.; Bateman, E.D.; Behera, D.; Bennoor, K.; Camargos, P.; Chavannes, N.; et al. Helsinki by nature: The nature step to respiratory health. Clin. Transl. Allergy 2019, 9, 57. [Google Scholar] [CrossRef]
- Rook, G.A. Regulation of the immune system by biodiversity from the natural environment: An ecosystem service essential to health. Proc. Natl. Acad. Sci. USA 2013, 110, 18360–18367. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Yu, Y.; Du, W.; Liu, Y.; Dai, R.; Tang, W.; Wang, P.; Zhang, C.; Shi, G. Fungal and bacterial microbiome dysbiosis and imbalance of trans-kingdom network in asthma. Clin. Transl. Allergy 2020, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.; Kauth, M.; Schwarze, J.; Körner-Rettberg, C.; Riedler, J.; Nowak, D.; Braun-Fahrländer, C.; von Mutius, E.; Bufe, A.; Holst, O. Inhalation of stable dust extract prevents allergen induced airway inflammation and hyperresponsiveness. Thorax 2006, 61, 134–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roslund, M.; Puhakka, R.; Grönroos, M.; Nurminen, N.; Oikarinen, S.; Gazali, A.M.; Cinek, O.; Kramná, L.; Siter, N.; Vari, H.K.; et al. Bio-diversity intervention enhances immune regulation and health-associated commensal microbiota among daycare children. Sci. Adv. 2020, 6, eaba2578. [Google Scholar] [CrossRef]
- Zhang, Z.; Reponen, T.; Hershey, G. Fungal exposure and asthma: IgE and non-IgE-mediated mechanisms. Curr. Allergy Asthma Rep. 2016, 16, 86. [Google Scholar] [CrossRef] [PubMed]
- Algetaa, H.; Mohammed, A.; Zhou, J.; Sing, N.; Nagarkatti, M.; Nagarkatti, P. Resveratrol-mediated attenuation of superantigen-driven acute respiratory distress syndrome is mediated by microbiota in the lungs and gut. Pharmacol. Res. 2021, 167, 105548. [Google Scholar] [CrossRef]
- Haahtela, T. A biodiversity hypothesis. Allergy 2019, 74, 1445–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuman, H.; Forsythe, P.; Uzan, A.; Avni, O.; Koren, O. Antibiotics in early life: Dysbiosis and the damage done. FEMS Microbiol. Rev. 2018, 42, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Versini, M.; Jeandel, P.Y.; Bashi, T.; Bizzaro, G.; Blank, M.; Shoenfeld, Y. Unraveling the hygiene hypothesis of helminthes and autoimmunity: Origins, pathophysiology, and clinical applications. BMC Med. 2015, 13, 8147. [Google Scholar] [CrossRef] [Green Version]
- Atosuo, J.; Karhuvaara, O.; Suominen, E.; Vilén, L.; Nuutila, J.; Putus, T.; Janne, A.; Outi, K.; Eetu, S.; Liisa, V.; et al. Indoor exposure to Streptomyces albus and Aspergillus versicolor elevates the levels of spore-specific IgG, IgG1 and IgG3 serum Abs in building users—A new ELISA-based assay for exposure assessment. Sci. Total Environ. 2020, 698, 134335. [Google Scholar] [CrossRef] [PubMed]
- Atosuo, J.; Karhuvaara, O.; Suominen, E.; Vilen, L.; Nuutila, J.; Putus, T. Indoor-related microbe damage induces complement system activation in building users. Innate Immun. 2021, 27, 15–22. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Corrier, D.E. Mycotoxicosis: Mechanisms of immunosuppression. Vet. Immunol. Immunopathol. 1991, 30, 73–87. [Google Scholar] [CrossRef]
- Fahimipour, A.K.; Ben Mamaar, S.; McFarland, A.G.; Blaustein, R.A.; Chen, J.; Glawe, A.J.; Kline, J.; Green, J.L.; Halden, R.U.; Van Den Wymelenberg, K.; et al. Antimicrobial chemicals associate with microbial function and antibiotic resistance indoors. mSystems 2018, 3, e00200-18. [Google Scholar] [CrossRef] [Green Version]
- Mendell, M.J.; Mirer, A.G.; Cheung, K.; Tong, M.; Douwes, J. Respiratory and allergic health effects of dampness, mold, and dampness related agents: A review of the epidemiologic evidence. Environ. Health Perspect. 2011, 119, 748–756. [Google Scholar] [CrossRef]
- Schutze, N.; Lehmann, I.; Bönisch, U.; Simon, J.; Polte, T. Exposure to mycotoxins increases the allergic immune response in a murine asthma model. Am. J. Respir. Crit. Care Med. 2010, 181, 1188–1199. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Guidelines for Indoor Air Quality: Dampness and Mould; World Health Organization: Copenhagen, Denmark, 2009. [Google Scholar]
- Wolff, H.; Mussalo-Rauhamaa, H.; Raitio, H.; Elg, P.; Orpana, A.; Piilonen, A.; Haahtela, T. Patients referred to an indoor air health clinic: Exposure to water-damaged buildings causes an increase of lymphocytes in bronchoalveolar lavage and a decrease of CD19 leucocytes in peripheral blood. Scand. J. Clin. Lab. Investig. 2009, 69, 537–544. [Google Scholar] [CrossRef]
- Wolff, H. Innate immunity and the pathogenicity of inhaled microbial particles. Int. J. Biol. Sci. 2011, 7, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yim, G.; Wang, H.H.; Davies, J. The truth about antibiotics. Int. J. Med. Microbiol. 2006, 296, 163–170. [Google Scholar] [CrossRef]
- Chang, C.; Gershwin, M.E. Mold hysteria: Origin of the hoax. Clin. Dev. Immunol. 2005, 12, 151–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordin, S. Mechanisms underlying nontoxic indoor air health problems: A review. Int. J. Hyg. Environ. Health 2020, 226, 113489. [Google Scholar] [CrossRef] [PubMed]
- Tuuminen, T.; Andersson, A.; Hyvönen, S.; Lohi, J.; Vaali, K. Indoor air nontoxicity should be proven with special techniques prior claiming that it may cause a variety of mental disorders. J. Hyg. Environ. Health 2020, 229, 113545. [Google Scholar] [CrossRef] [PubMed]
- Tuuminen, T.; Lohi, J. Dampness and mold hypersensitivity syndrome is a biotoxicosis that should be diagnosed promptly. Adv. Clin. Toxicol. 2019, 4, 1–4. [Google Scholar] [CrossRef]
- Valtonen, V. Clinical diagnosis of the dampness and mold hypersensitivity syndrome: Review of the literature and suggested diagnostic criteria. Front. Immunol. 2017, 8, 951. [Google Scholar] [CrossRef] [Green Version]
- Hayward, J.; Mathur, A.; Ngo, C.; Man, S. Cytosolic recognition of microbes and pathogens: Inflammasome in action. Microbiol. Mol. Biol. Rev. 2018, 82, e00015-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salo, M.J.; Marik, T.; Mikkola, R.; Andersson, M.A.; Kredics, L.; Salonen, H.; Kurnitski, J. Penicillium expansum strain isolated from indoor building material was able to grow on gypsum board and emitted guttation droplets containing chaetoglobosins and communesins A, B and D. J. Appl. Microbiol. 2019, 127, 1135–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salo, M.J.; Marik, T.; Bencsik, O.; Mikkola, R.; Kredics, L.; Szekeres, A.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Screening mold colonies by using two toxicity assays revealed indoor strains of Aspergillus calidoustus producing ophiobolins G and K. Toxins 2019, 11, 683. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi, 2nd ed.; Westerdijk Laboratory Manual Series 2; Westerdijk Fungal Biodiversity Institute: Utrecht, The Netherlands, 2019. [Google Scholar]
- Cai, F.; Gao, R.; Zhao, Z.; Ding, M.; Jiang, S.; Yagtu, C.; Zhu, H.; Zhang, J.; Ebner, T. Evolutionary compromises in fungal fitness: Hydrophobins can hinder the adverse dispersal of conidiospores and challenge their survival. ISME J. 2020, 14, 2610–2624. [Google Scholar] [CrossRef] [PubMed]
- Meyer, V.; Basenko, E.Y.; Benz, J.P.; Braus, G.H.; Caddick, M.X.; Csukai, M.; de Vries, R.P.; Endy, D.; Frisvad, J.C.; Gunde- Cimerman, N.; et al. Growing a circular economy with fungal biotechnology: A white paper. Fungal Biol. Biotechnol. 2020, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Gareis, M.; Gottschalk, C. Stachybotrys spp. and the guttation phenomenon. Mycotox. Res. 2014, 30, 151–159. [Google Scholar] [CrossRef]
- Salo, J.; Andersson, M.A.; Mikkola, R.; Kredics, L.; Viljanen, M.; Salkinoja-Salonen, M. Vapor as a carrier oftoxicity in a health troubled building. In Proceedings of the Healthy Buildings 2015—Europe (ISIAQ International), Eindhoven, The Netherlands, 18–20 May 2015. [Google Scholar]
- Lu, R.; Pørneki, A.; Lindgreen, J.; Li, Y.; Madsen, A.M. Species of fungi and pollenin the PM1 and the inhalable fraction of indoor air in homes. Atmosphere 2021, 12, 404. [Google Scholar] [CrossRef]
- Salo, J. Developent of analytical methods for assaying metabolites of molds in buildings. Master Thesis, Department of Structural Engineering and Building Technology, Aalto University, Espoo, Finland, 2014. [Google Scholar]
- Argoudelis, A.; Mizsak, S.A. Melinacidins II, III, IV. Structural studies. J. Antibiot. 1977, 30, 468–473. [Google Scholar] [CrossRef] [Green Version]
- Ebead, G.A.; Overy, D.P.; Berrue, F.; Kerr, R.G. Westerdykella reniformis sp. nov., producing the antibiotic metabolites melinacidin IV and chetracin B. IMA Fungus 2012, 3, 189–201. [Google Scholar] [CrossRef]
- Wu, Z.J.; Li, G.Y.; Fang, D.M.; Qi, H.Y.; Ren, W.J.; Zhang, G.L. Analysis of epipolythiodioxopiperazines in fungus Chaetomium cochliodes using HPLC-ESI-MS/MS/MS. Anal. Chem. 2008, 80, 217–226. [Google Scholar] [CrossRef]
- Li, L.; Li, D.; Luan, Y.; Gu, Q.; Zhu, T. Cytotoxic metabolites from the antarctic psychrophilic fungus Oidiodendron truncatum. J. Nat. Prod. 2012, 75, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhu, X.; Kolli, S.; Wang, H.; Pearce, C.J.; Oberlies, N.H.; Phelps, M.A. Plasma pharmacokinetics and bioavailability of verticillin A following different routes of administration in mice using liquid chromatography tandem mass spectrometry. J. Pharm. Biomed. Anal. 2017, 139, 187–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagnoli, E.; Marik, T.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Indoor Trichoderma strains emitting peptaibols in guttation droplets. J. Appl. Microbiol. 2018, 125, 1408–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagnoli, E.; Salo, J.; Toivonen, M.S.; Marik, T.; Mikkola, R.; Kredics, L.; Vicente-Carrillo, A.; Nagy, S.; Andersson, M.T.; Andersson, M.A.; et al. An evaluation of boar spermatozoa as a biosensor for the detection of sublethal and lethal toxicity. Toxins 2018, 10, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraldo, A.; Crous, P.W. Inside Plectosphaerellaceae. Stud. Mycol. 2019, 92, 227–286. [Google Scholar] [CrossRef]
- Mohammadi, A.; Amini, Y. Molecular characterization and identification of Acrostalagmus luteoalbus from saffron in Iran. Agric. Sci. Dev. 2015, 4, 16–18. [Google Scholar]
- Zare, R.; Gams, W.; Schroers, H.-J. The type species of Verticillium is not congeneric with the plant-pathogenic species placed in Verticillium and it is not the anamorph of ‘Nectria’ inventa. Mycol. Res. 2004, 108, 576–582. [Google Scholar] [CrossRef]
- Zare, R.; Asgari, B. Report of two new hyperparasitic species from Golestan Province. Rostaniha 2008, 8, 116–117. [Google Scholar]
- Alvárez-Rodríguez, M.L.; López-Ocaña, L.; López-Coronado, J.M.; Rodríguez, E.; Martínez, M.J.; Larriba, G.; Coque, J.-J.R. Cork taint of wines: Role of filamentous fungi isolated from cork in the formation of 2,4,6-trichloroanisole by O methylation of 2,4,6-trichlorophenol. Appl. Environ. Microbiol. 2002, 68, 5860–5869. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, D.; Waring, P.; Howlett, B. The epipolythiodioxopiperazine (ETP) class of fungal toxins: Distribution, mode of action, functions and biosynthesis. Microbiology 2005, 151, 1021–1032. [Google Scholar] [CrossRef] [Green Version]
- Argoudelis, A.; Reusser, F. Melinacidins, a new family of antibiotics. J. Antibiot. 1971, 6, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Bugli, F.; Sterbini, F.P.; Cacaci, M.; Martini, C.; Lancellotti, S.; Stigliano, E.; Torelli, R.; Arena, V.; Caira, M.; Posteraro, P.; et al. Increased production of gliotoxin is related to the formation of biofilm by Aspergillus fumigatus: An immunological approach. Pathog. Dis. 2014, 70, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Dagenais, T.R.T.; Keller, N.P. Pathogenesis of Aspergillus fumigatus in invasive aspergillosis. Clin. Microbiol. Rev. 2009, 22, 447–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grovel, O.; Kerzaon, I.; Petit, K.; Du Pont, T.; Pouchus, Y.-F. A new and rapid bioassay for the detection of gliotoxin and related epipolythiodioxopiperazines produced by fungi. J. Microbiol. Meth. 2006, 66, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.; Frisvad, J.; Søndergaard, I.; Rasmussen, I.; Larsen, L. Associations between fungal species and water-damaged building materials. J. Appl. Environ. Microbiol. 2011, 77, 4180–4188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salonen, P. Homeiden Toksiinien Tuotto ja Sen Seuranta Metaboliitti ja RNA-Tasolla. Bachelor’s Thesis, Aalto University, Espoo, Finland, 2014. [Google Scholar]
- Mikkola, R.; Andersson, M.A.; Kredics, L.; Grigoriev, P.A.; Sundell, N.; Salkinoja-Salonen, M.S. 20-residue and 11-residue peptaibols from the fungus Trichoderma longibrachiatum are synergistic in forming Na+/K+-permeable channels and adverse action towards mammalian cells. FEBS J. 2012, 279, 4172–4190. [Google Scholar] [CrossRef]
- Täubel, M.; Karvonen, A.M.; Reponen, T.; Hyvärinen, A.; Vesper, S.; Pekkanena, J. Application of the environmental relative moldiness index in Finland. Appl. Environ. Microbiol. 2016, 82, 578–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, D.; Habib, J.; Galler, H.; Buzina, W.; Schlacher, R.; Marth, E.; Reinthaler, F.F. Assessment of indoor air in Austrian apartments with and without visible mold growth. Atmos. Environ. 2007, 41, 5192–5201. [Google Scholar] [CrossRef]
- Pessi, A.-M.; Suonketo, J.; Pentti, M.; Kurkilahti, M.; Peltola, K.; Rantio-Lehtimäki, A. Microbial growth inside insulated external walls as an indoor air biocontamination source. Appl. Environ. Microbiol. 2002, 68, 963–967. [Google Scholar] [CrossRef] [Green Version]
- Vornanen-Winqvist, C.; Salonen, H.; Järvi, K.; Andersson, M.A.; Mikkola, R.; Marik, T.; Kredics, L.; Kurnitski, J. Effects of ventilation improvement on measured and perceived indoor quality in a school building with a hybrid ventilation system. Int. J. Environ. Res. Publ. Health 2018, 15, 1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birjukovski, M.; Apine, I.A.; Jakovics, A. Numerical models for long-term performance assessment of lightweight insulating assemblies. IOP Conf. Ser. Mater. Sci. Eng. 2019, 660, 012024. [Google Scholar] [CrossRef] [Green Version]
- Chao, C.; Hu, J.S. Development of a dual-mode demand control ventilation strategy for indoor air quality control and energy saving. Build. Environ. 2004, 39, 385–397. [Google Scholar] [CrossRef]
- Montgomery, J.; Storey, S.; Bartlett, K. Comparison of the indoor air quality in an office operating with natural or mechanical ventilation using short-term intensive pollutant monitoring. Indoor Built Environ. 2015, 24, 777–787. [Google Scholar] [CrossRef]
- Ahmad, A.; Othman, M.R.; Latif, M.T. Early study of surfactants in indoor dust and their connection with street dust. Int. J. Environ. Res. 2009, 3, 403–410. [Google Scholar] [CrossRef]
- Veirup, K.; Wolkoff, P. Linear alkylbenzene sulfonates in indoor floor dust. Sci. Total. Environ. 2003, 300, 51–58. [Google Scholar] [CrossRef]
- Williams, P.; Barnes, C.; Portnoy, J. Innate and adaptive immune response to fungal products and allergens. J. Allergy Clin. Immunol. Pract. 2016, 4, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.; Magun, B.E.; Wood, L.J. Lung inflammation caused by inhaled toxicants: A review. Int. J. Chronic Obstr. Pulm. Dis. 2016, 11, 1391–1401. [Google Scholar] [CrossRef] [Green Version]
- West, A.P. Mitochondrial dysfunction as a trigger of innate immune responses and inflammation. Toxicology 2017, 391, 54–63. [Google Scholar] [CrossRef]
- Araki, A.; Kawai, T.; Eitaki, Y.; Kanazawa, A.; Morimoto, K.; Nakayama, K.; Shibata, E.; Tanaka, M.; Takigawa, T.; Yoshimura, T.; et al. Relationship between selected indoor volatile organic compounds, so-called microbial VOC, and the prevalence of mucous membrane symptoms in single family homes. Sci. Total Environ. 2010, 408, 2208–2215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wålinder, R.; Ernstgard, L.; Norback, D.; Wieslander, G.; Johanson, G. Acute effects of 1-octen-3-ol, a microbial volatile organic compound (MVOC)—An experimental study. Toxicol. Lett. 2008, 181, 141–147. [Google Scholar] [CrossRef] [PubMed]
- STM 2003, Asumis Terveysohje Helsinki: Sosiaali-ja Terveysministeriön Oppaita 2003:1.23 s. Helsinki 19:th February 2003. ISSN 1236-116X. ISBN 952-00-1301-6.
- Asumisterveysopas: Sosiaali- ja terveysministeriön Asumisterveysohjeen (STM:n oppaita 2003:1) soveltamisopas. Ympäristö ja terveyslehti 2009. ISBN 9789529637386.
- Samson, R.A. Ecology and general characteristics of indoor fungi. In Fundamentals of Mold Growth in Indoor Environments and Strategies for Healthy Livinged; Adan, O.C.F., Samson, R.A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherland, 2011; pp. 101–116. [Google Scholar]
- Samson, R.A.; Hoekstra, E.S.; Frisvad, J.C.; Filtenborg, O. (Eds.) Introduction to Food and Air-Borne Fungi, 6th ed.; Centraalbureau voor Schimmelcultures: Utrecht, The Netherland, 2002. [Google Scholar]
- Salo, J.; Kedves, O.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Kurnitski, J.; Salonen, H. Detection of Chaetomium globosum, Ch. cochliodes and Ch. rectangulare during the diversity tracking of mycotoxin-producing Chaetomium-like isolates obtained in buildings in Finland. Toxins 2020, 12, 443. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomass Lysates | Morphology in Microscope | Origin Sample | ||||
|---|---|---|---|---|---|---|
| Morphotype | Toxicity * BSMI ICP | Fluorescence/Colony Color | Conidiophores | Conidia | ||
| MT1 | Acrostalagmus luteoalbus | |||||
| Isolates: POB8 1, A1/K, A2/K, A3/K, A4/K | − | + |  Brown/white |  |  | 1P61, 1K |
| MT2 | Aspergillussection Versicolores | |||||
| Isolates: AS1, AS2 AS3, AS4, AS5 | − | + |  Green/yellow |  |  | 1POB 1P61 1K |
| MT2 | Reference strain Aspergillus versicolor SL/3 6 | |||||
| − | + |  Green/yellow |  |  | ||
| MT3 | Aspergillus section Circumdati | |||||
| Isolates: AW7, AW8, AW9, AW10, AW11 | + | + |  Yellow |  |  | 3MW |
| MT3 | Reference strain Aspergillus westerdijkiae PP2 6 | |||||
 Yellow |  |  | ||||
| MT4 | Aspergillus section Nigri | |||||
| Isolates: AN1, AN2, AN3, AN7, AN11 | − | + |  Black |  |  | 3MW 1K |
| MT4 | Reference strain Aspergillus niger HAMBI 1271 | |||||
 Black |  |  | ||||
| MT5 | Trichoderma atroviride | |||||
| Isolates: Tri335 2 Tri335b, Tri335c Tri336, Tri337 | + | + |  Green |  |  | 3MW |
| MT6 | Trichoderma trixiae | |||||
| Isolates: Th1/kg, Th2/kg, Th4/kg, Th5/kg 3, Th6/kg 4 |  Green |  |  | 1K 3MW | ||
| MT7 | Penicillium expansum | |||||
| Isolates: Rc P61 5, RcP62 | (+) | + |  Green-yellow/gray |  |  | 1P61 |
| MT8–MT10 | Spore-forming Actinobacteria | |||||
| Isolates: Str3/KN, Str4/KN | + | + |  |  |  | 1K |
| Isolates: Str1/KN, Str2/KN | + |  |  |  | 1K | |
| Isolates: Str5/KN, Str6/KN | + |  |  |  | 1K | |
| Biomass Lysates | Morphology in Microscope | Origin | ||||
|---|---|---|---|---|---|---|
| Toxicity | Fluorescence/ Colony Color | Conidiophores | Conidia | Room | ||
| Morphotype | BSMI | ICP | ||||
| MT1 | Acrostalagmus sp. | |||||
| Isolates: POB1, PH20 | − | + |  Brown/white |  |  | 131b |
| MT2 | Aspergillus section Versicolores | |||||
| Isolates: A20, A11, A13, A14, A15 | − | + |  Green/yellow |  |  | 335 131b |
| MT3 | Aspergillus section Circumdati | |||||
| Isolates: AW1, AW2, AW3, AW4, Aw5 | + | + |  Yellow |  |  | 335 |
| MT4 | Aspergillus section Nigri | |||||
| Isolates:AN10, AN11, AN12 |  Black |  |  | 335 145b | ||
| MT5 | Trichoderma sp. | |||||
| Isolates: PO1, PO2, PO3, PO4, PO5 | + | + |  Green |  |  | 335 |
| MT6 | Trichoderma sp. | |||||
| Isolates: T351, T355, T337, T338, T330 | + | + |  Green |  |  | 335 |
| MT7 | Penicillium expansum | |||||
| Isolates: MH6 1, P3a, P32, P33 | (+) | + |  Green/yellow/gray |  |  | 131b 131a |
| MT11 | Chaetomium globosum | |||||
| Isolates: MH5, M13, M14, MH52 2 | + | + |  Black |  |  | 131b 145b 146 |
| MT12 | Rhizopus sp. | |||||
| Isolates: R1, R2, R3, R4 | − | − |  Black |  |  | 134 223 |
| Ethanol-Extracted Dry Substances | Exposure Concentrations EC50 µg mL−1 | |||
|---|---|---|---|---|
| Boar sperm assays | Somatic cell line | |||
| BSMIR | SMIDM | BSMR | ICP | |
| Exposure time and temperature | 20 min, 22 °C | 2 h, 37 °C | 1 d, 22 °C | 3 d, 37 °C |
| Strain code | ||||
| Acrostalagmus luteoalbus POB8 | 100 | 6 | 10 | 10 |
| Acrostalagmus sp. Ac1/KG | 100 | 6 | 10 | 10 |
| Acrostalagmus sp. Ac2/kg | 100 | 12 | 20 | 20 |
| Acrostalagmus sp. AC3/kg | 100 | 6 | 10 | 20 |
| Acrostalagmus sp. AC4/kg | 100 | 6 | 10 | 10 |
| Acrostalagmus sp. POB1 | 100 | 12 | 20 | 10 |
| Reference strains | Ethanol extracted dry substances | EC50 µg mL−1 | ||
| Aspergillus versicolor SL/3 | >100 | >100 | 20 | 1 |
| Chaetomium globosum MTAV 35 | ND | 450 | 3 | 40 |
| Trichoderma atroviride Tri335 | 5 | 2 | 5 | 60 |
| Liquid exudates | Exposure concentration EC50 µL mL−1 | |||
| Acrostalagmus luteoalbus POB8 | ND | ND | 25 | 50 |
| Reference strains | Liquid exudates | Exposure concentration EC50 µL mL−1 | ||
| Aspergillus versicolor SL/3 | ND | ND | >100 | >100 |
| Aspergillus westerdijkiae PP2 | ND | ND | >100 | >100 |
| Aspergillus calidoustus MH34 | ND | ND | >100 | >100 |
| Stachybotrys sp. HJ5 * | ND | ND | >50 | 20 |
| Penicillium expansum RCP61 * | ND | ND | 2.5 | 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andersson, M.A.; Salo, J.; Mikkola, R.; Marik, T.; Kredics, L.; Kurnitski, J.; Salonen, H. Melinacidin-Producing Acrostalagmus luteoalbus, a Major Constituent of Mixed Mycobiota Contaminating Insulation Material in an Outdoor Wall. Pathogens 2021, 10, 843. https://doi.org/10.3390/pathogens10070843
Andersson MA, Salo J, Mikkola R, Marik T, Kredics L, Kurnitski J, Salonen H. Melinacidin-Producing Acrostalagmus luteoalbus, a Major Constituent of Mixed Mycobiota Contaminating Insulation Material in an Outdoor Wall. Pathogens. 2021; 10(7):843. https://doi.org/10.3390/pathogens10070843
Chicago/Turabian StyleAndersson, (Aino) Maria A., Johanna Salo, Raimo Mikkola, Tamás Marik, László Kredics, Jarek Kurnitski, and Heidi Salonen. 2021. "Melinacidin-Producing Acrostalagmus luteoalbus, a Major Constituent of Mixed Mycobiota Contaminating Insulation Material in an Outdoor Wall" Pathogens 10, no. 7: 843. https://doi.org/10.3390/pathogens10070843