
Effects of High Temperature-Triggered Transcriptomics on the Physiological Adaptability of Cenococcum geophilum, an Ectomycorrhizal Fungus
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Culture Conditions
2.2. Determination of Superoxide Dismutase (SOD), Peroxidase (POD) and Catalase (CAT) in Mycelium
2.3. Determination of the Types and Content of Organic Acids in Culture Medium
2.4. RNA-Seq Analysis
2.5. RT-qPCR Analysis
2.6. Statistical Analysis
3. Results
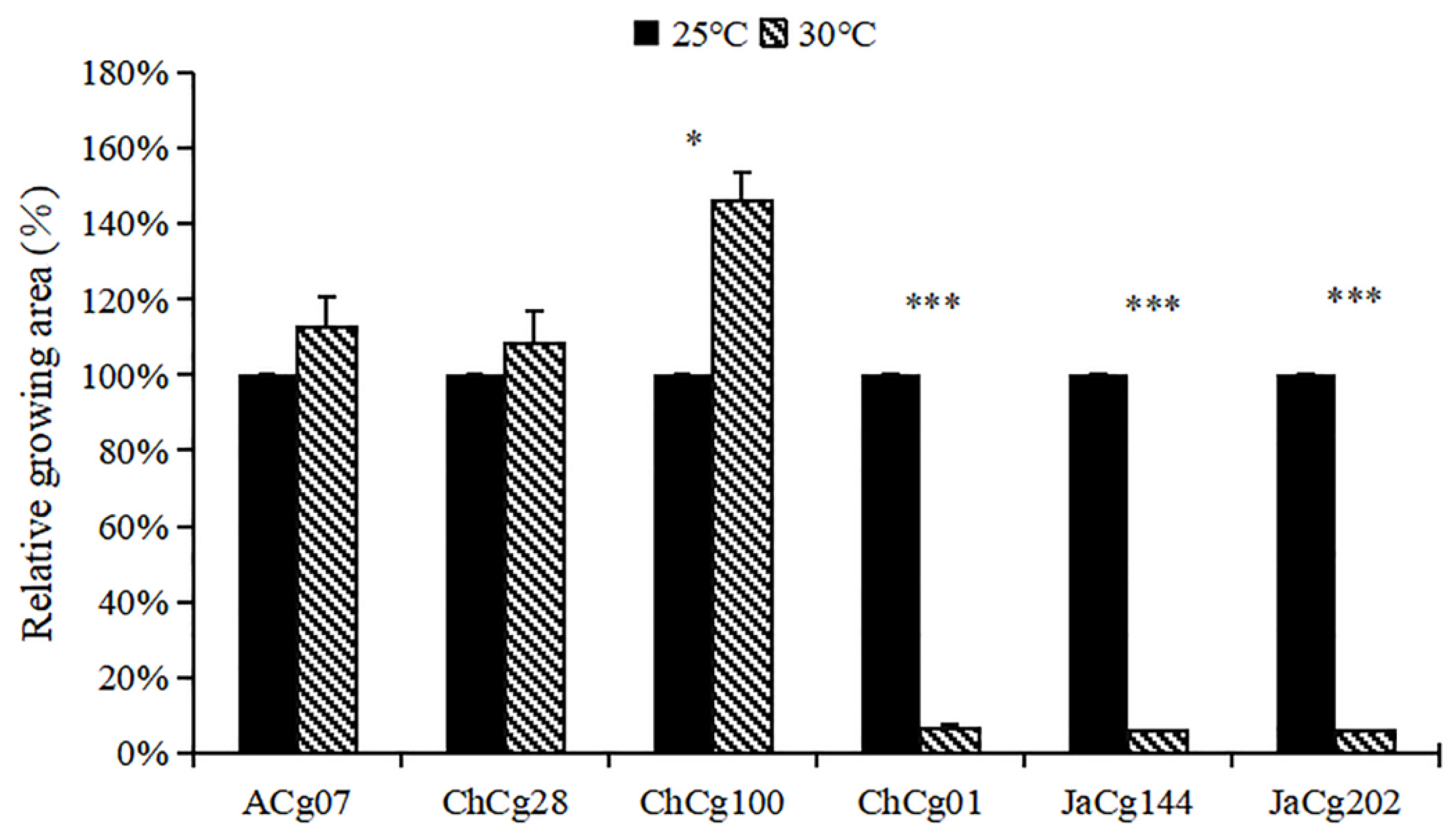
3.1. Effects of High Temperature Treatment on Mycelial Growth of C. geophilum
3.2. Effect of High Temperature on the Content of Organic Acids
3.3. Effect of High Temperature on the Activity of Antioxidative Enzymes
3.4. Transcriptome Response of C. geophilum Mycelia in Different Temperatures
3.4.1. Overview of Transcriptome Assembly and Functional Annotation
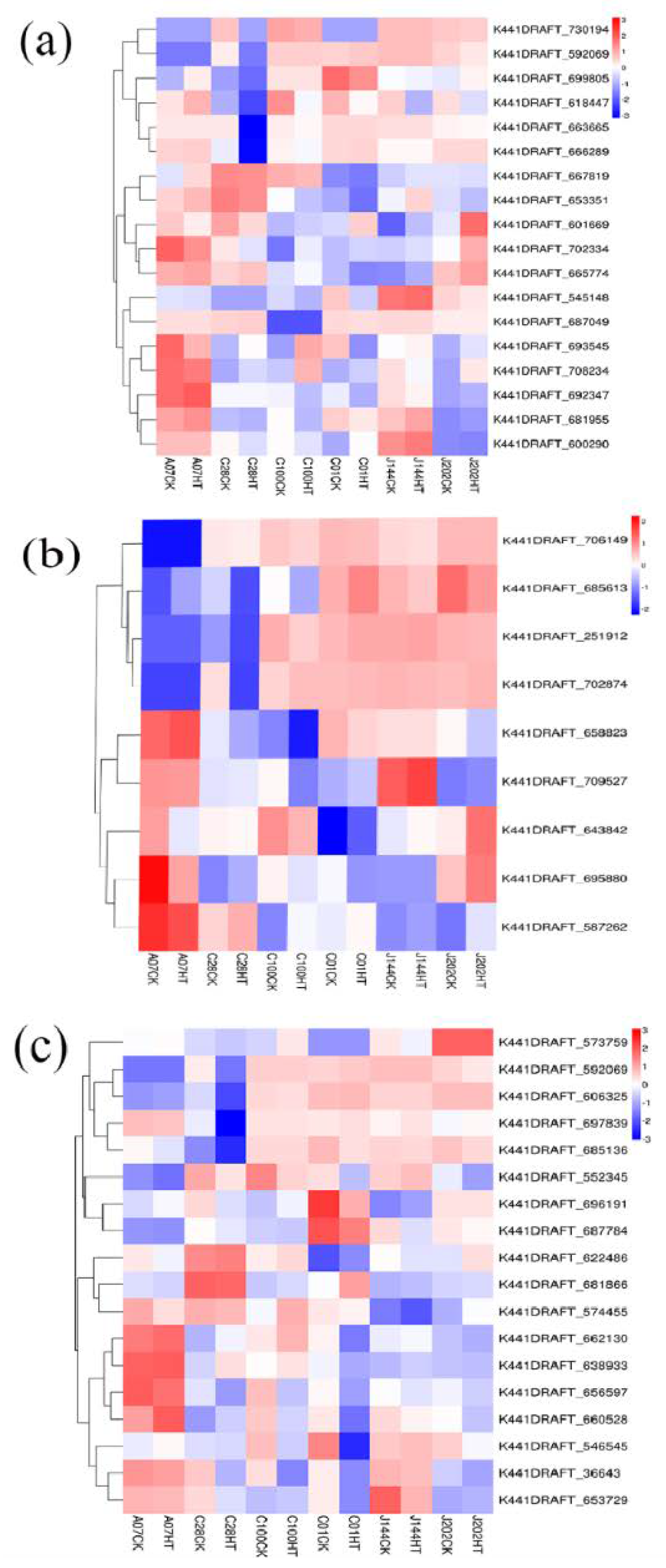
3.4.2. Differential Gene Screening
3.4.3. Gene Ontology (GO) Analysis of DEGs
3.4.4. Kyoto Encyclopedia of Genes and Genomes (KEGG) Analysis of DEGs
3.4.5. Genes Involved in Key and Related Metabolism Pathways for High Temperature Tolerance and Reduction in C. geophilum Isolates
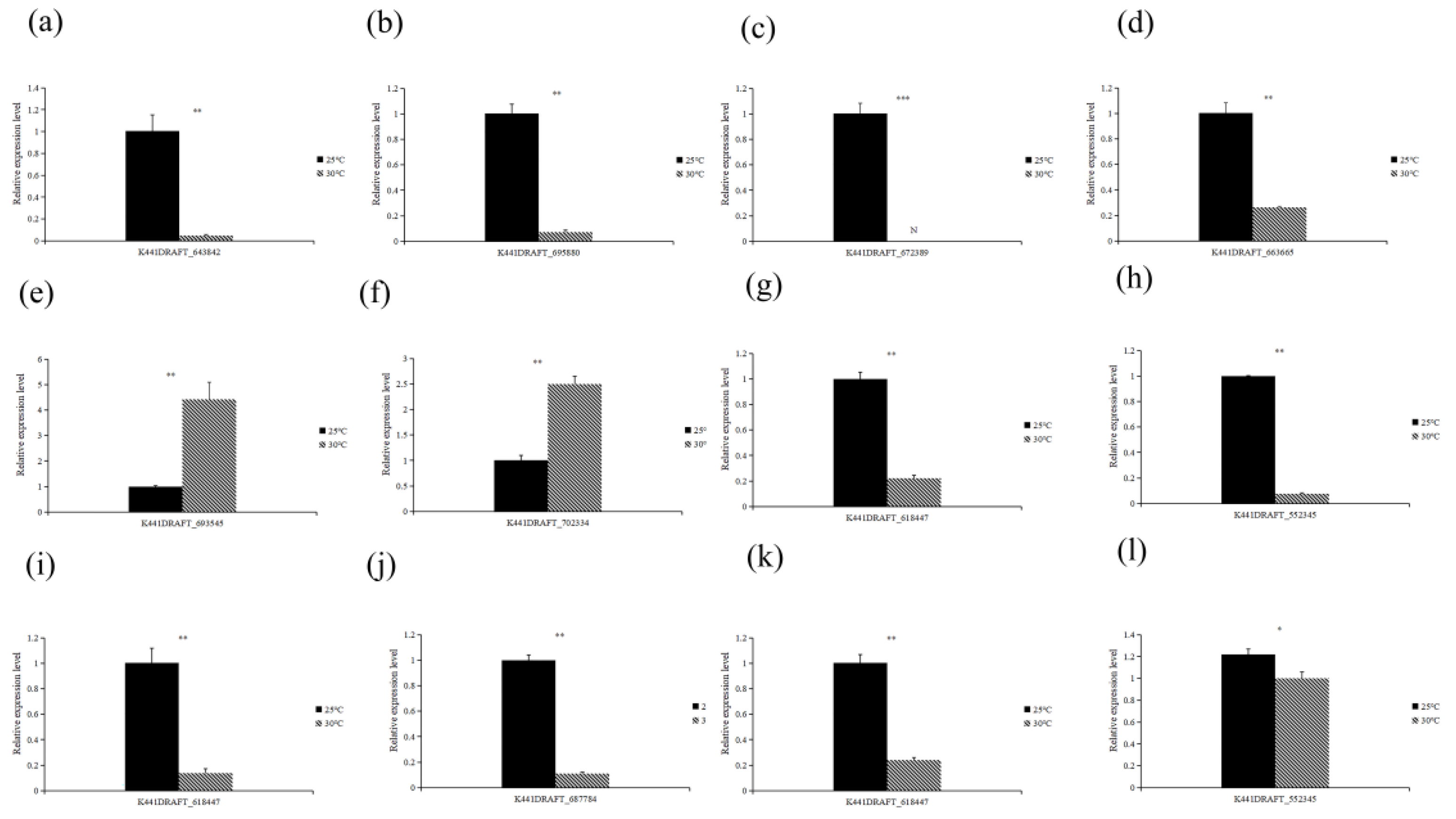
3.4.6. RT-qPCR Verification of DEGs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruno, M.; Michael, M.; Ülo, M.; Evelyn, U. Increased carbon assimilation and efficient water usage may not compensate for carbon loss in European forests. Commun. Earth Environ. 2022, 3, 194. [Google Scholar]
- Han, J.; Singh, V.P. Forecasting of droughts and tree mortality under global warming: A review of causative mechanisms and modeling methods. J. Water Clim. Chang. 2020, 11, 600–632. [Google Scholar] [CrossRef] [Green Version]
- Pengxia, Z.; Qing, Y.; Fang, O.; Longhui, P.; Xingping, L.; Yuehua, G.; Juping, Z. Global warming and droughts aggravates forest damage resulting from pests and diseases in Jiangxi Province. Acta Ecol. Sin. 2017, 37, 639–649. [Google Scholar]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef]
- Strullu-Derrien, C.; Selosse, M.A.; Kenrick, P.; Martin, F.M. The origin and evolution of mycorrhizal symbioses: From palaeomycology to phylogenomics. New Phytol. 2018, 220, 1012–1030. [Google Scholar] [CrossRef] [Green Version]
- Zong, K.; Huang, J.; Nara, K.; Chen, Y.; Shen, Z.; Lian, C. Inoculation of ectomycorrhizal fungi contributes to the survival of tree seedlings in a copper mine tailing. J. For. Res. 2017, 20, 493–500. [Google Scholar] [CrossRef]
- Seb, J.; Ajungla. T. Role of ectomycorrhiza in forest ecosystems: A review. Int. J. Adv. Res. 2018, 6, 866–873. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, T.; Zhou, Y.; Zou, X.; Yin, Y.; Li, H.; Liu, L.; Zhang, S. Ectomycorrhizal symbioses increase soil calcium availability and water use efficiency of Quercus acutissima seedlings under drought stress. Eur. J. For. Res. 2021, 140, 1039–1048. [Google Scholar] [CrossRef]
- Wen, Z.; Xing, J.; Liu, C.; Zhu, X.; Zhao, B.; Dong, J.; He, T.; Zhao, X.; Hong, L. The effects of ectomycorrhizal inoculation on survival and growth of Pinus thunbergii seedlings planted in saline soil. Symbiosis 2022, 86, 71–80. [Google Scholar] [CrossRef]
- Hachani, C.; Lamhamedi, M.S.; Zine El Abidine, A.; Abassi, M.; Khasa, D.P.; Béjaoui, Z. Water relations, gas exchange, chlorophyll fluorescence and electrolyte leakage of ectomycorrhizal Pinus halepensis seedlings in response to multi-heavy metal stresses (Pb, Zn, Cd). Microorganisms 2021, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Steidinger, B.S.; Bhatnagar, J.M.; Vilgalys, R.; Taylor, J.W.; Qin, C.; Zhu, K.; Bruns, T.D.; Peay, K.G. Ectomycorrhizal fungal diversity predicted to substantially decline due to climate changes in North American Pinaceae forests. J. Biogeogr. 2020, 47, 772–782. [Google Scholar] [CrossRef]
- Koizumi, T.; Nara, K. Ectomycorrhizal fungal communities in ice-age relict forests of Pinus pumila on nine mountains correspond to summer temperature. ISME J. 2020, 14, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.; Gao, T.; Li, G.; Xu, Y.; Xing, P.; Zhou, J.; Xie, M.; Li, J.; Li, Y. Correlation between ectomycorrhizal mycorrhizal fungal community and environmental factors in pure forest of Pinus red in Northeast China. J. Fungus 2020, 39, 743–754. [Google Scholar] [CrossRef]
- Parts, K.; Tedersoo, L.; Schindlbacher, A.; Sigurdsson, B.D.; Leblans, N.I.W.; Oddsdóttir, E.S.; Borken, W.; Ostonen, I. Acclimation of fine root systems to soil warming: Comparison of an experimental setup and a natural soil temperature gradient. Ecosystems 2018, 22, 457–472. [Google Scholar] [CrossRef] [Green Version]
- Kwatcho Kengdo, S.; Persoh, D.; Schindlbacher, A.; Heinzle, J.; Tian, Y.; Wanek, W.; Borken, W. Long-term soil warming alters fine root dynamics and morphology, and their ectomycorrhizal fungal community in a temperate forest soil. Glob. Chang. Biol. 2022, 28, 3441–3458. [Google Scholar] [CrossRef]
- Kilpeläinen, J.; Aphalo, P.J.; Lehto, T. Temperature affected the formation of arbuscular mycorrhizas and ectomycorrhizas in Populus angustifolia seedlings more than a mild drought. Soil Biol. Biochem. 2020, 146, 107798. [Google Scholar] [CrossRef]
- Liu, J.Y.; Men, J.L.; Chang, M.C.; Feng, C.P.; Yuan, L.G. iTRAQ-based quantitative proteome revealed metabolic changes of Flammulina velutipes mycelia in response to cold stress. J. Proteom. 2017, 156, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Leyval, C.; Berthelin, J. Interactions betweenLaccaria laccata, Agrobacterium radiobacter and beech roots: Influence on P, K, Mg, and Fe mobilization from minerals and plant growth. Plant Soil 1989, 117, 103–110. [Google Scholar] [CrossRef]
- Wallander, H. Uptake of P from apatite by Pinus sylvestris seedlings colonised by different ectomycorrhizal fungi. Plant Soil 2000, 218, 249–256. [Google Scholar] [CrossRef]
- Bandou, E.; Lebailly, F.; Muller, F.; Dulormne, M.; Toribio, A.; Chabrol, J.; Courtecuisse, R.; Plenchette, C.; Prin, Y.; Duponnois, R.; et al. The ectomycorrhizal fungus Scleroderma bermudense alleviates salt stress in seagrape (Coccoloba uvifera L.) seedlings. Mycorrhiza 2006, 16, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Chen, M.Y.; Liang, W.; Chen, Y.; Guo, M.Y. Zinc Deficiency aggravation of ROS and inflammatory injury leading to renal fibrosis in mice. Biol. Trace Elem. Res. 2020, 199, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Li, J.; Sun, J.; Zhang, X.; Chen, S. Changes of antioxidant enzyme activities in ectomycorrhizal fungi Paxillus under salt stress. Anhui Agric. Sci. 2012, 40, 8401–8403+8405. [Google Scholar] [CrossRef]
- Dachuan, Y.; Jinyu, Q. The physiological response of ectomycorrhizal fungus Lepista sordida to Cd and Cu stress. PeerJ 2021, 9, e11115. [Google Scholar] [CrossRef] [PubMed]
- Khullar, S.; Reddy, M.S. Cadmium induced glutathione bioaccumulation mediated by gamma-glutamylcysteine synthetase in ectomycorrhizal fungus Hebeloma cylindrosporum. Biometals 2019, 32, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Spatafora, J.W.; Owensby, C.A.; Douhan, G.W.; Boehm, E.W.A.; Schoch, C.L. Phylogenetic placement of the ectomycorrhizal genus Cenococcum in Gloniaceae (Dothideomycetes). Mycologia 2012, 104, 758–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massicotte, H.B.; Trappe, J.M.; Peterson, R.L.; Melville, L.H. Studies on Cenococcum geophilum. II. Sclerotium morphology, germination, and formation in pure culture and growth pouches. Can. J. Bot. 1992, 70, 125–132. [Google Scholar] [CrossRef]
- Kerner, R.; Delgado-Eckert, E.; Castillo, E.D.; Müller-Starck, G.; Peter, M.; Kuster, B.; Tisserant, E.; Pritsch, K. Comprehensive proteome analysis in Cenococcum geophilum Fr. as a tool to discover drought-related proteins. J. Proteom. 2012, 75, 3707–3719. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, C.; Tsuruta, M.; Matsushita, N.; Goto, S.; Shen, Z.; Tsugama, D.; Zhang, S.; Lian, C. Physiological and transcriptional responses of the ectomycorrhizal fungus Cenococcum geophilum to salt stress. Mycorrhiza 2022, 32, 327–340. [Google Scholar] [CrossRef]
- Shi, Y.; Yan, T.; Yuan, C.; Li, C.; Rensing, C.; Chen, Y.; Xie, R.; Zhang, T.; Lian, C. Comparative physiological and transcriptome analysis provide insights into the response of Cenococcum geophilum, an ectomycorrhizal fungus to cadmium Stress. J. Fungi 2022, 8, 724. [Google Scholar] [CrossRef] [PubMed]
- Obase, K.; Douhan, G.W.; Matsuda, Y.; Smith, M.E. Culturable fungal assemblages growing within Cenococcum sclerotia in forest soils. FEMS Microbiol. Ecol. 2014, 90, 708–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyamsanjaa, K.; Oyuntsetseg, B.; Takashima, Y.; Sakagami, N.; Watanabe, M. Characteristics of;Cenococcum geophilum;sclerotia found in steppe forest soil in Mongolia. J. For. Res. 2022, 27, 76–82. [Google Scholar] [CrossRef]
- Obase, K.; Douhan, G.W.; Matsuda, Y.; Smith, M.E. Genotypic diversity of the ectomycorrhizal fungus Cenococcum geophilum at a soil core scale. Present. Database Annu. Meet. Jpn. For. Soc. 2016, 127, 559. [Google Scholar]
- Jany, J.-L.; Martin, F.; Garbaye, J. Respiration activity of ectomycorrhizas from Cenococcum geophilum and Lactarius sp. in relation to soil water potential in five beech forests. Plant Soil 2003, 255, 487–494. [Google Scholar] [CrossRef]
- Herzog, C.; Peter, M.; Pritsch, K.; Egli, S. Drought and air warming affects abundance and exoenzyme profiles of Cenococcum geophilum associated with Quercus robur, Q. petraea and Q. pubescens. Plant Biol. 2013, 15 (Suppl. 1), 230–237. [Google Scholar] [CrossRef]
- Du, S.; Sang, Y.; Liu, X.; Xing, S.; Li, J.; Tang, H.; Sun, L. Transcriptome profile analysis from different sex types of Ginkgo biloba L. Front. Plant Sci. 2016, 7, 871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods Companion Methods Enzymol. 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shinohara, M.L.; LoBuglio, K.F.; Rogers, S.O. Group-I intron family in the nuclear ribosomal RNA small subunit genes of Cenococcum geophilum isolates. Curr. Genet. 1996, 29, 377–387. [Google Scholar] [CrossRef]
- Bai, Z.; Yuan, Z.-Q.; Wang, D.-M.; Fang, S.; Ye, J.; Wang, X.-G.; Yuan, H.-S. Ectomycorrhizal fungus-associated determinants jointly reflect ecological processes in a temperature broad-leaved mixed forest. Sci. Total Environ. 2020, 703, 135475. [Google Scholar] [CrossRef]
- Ming, W.; Ling, Y.; Zhi, F.; Hong, Y.; Jian, S. Effects of aluminum on oxalate secretion and NPK uptake by ectomycorrhizal fungi. For. Sci. 2012, 48, 82–88. [Google Scholar] [CrossRef]
- Hu, Y.; Wu, S.; Sun, Y.; Li, T.; Zhang, X.; Chen, C.; Lin, G.; Chen, B. Arbuscular mycorrhizal symbiosis can mitigate the negative effects of night warming on physiological traits of Medicago truncatula L. Mycorrhiza 2015, 25, 131–142. [Google Scholar] [CrossRef]
- Alberto, A.; Georg, G.; Sarmite, K.; Robert, M.; Leopold, S.; Jens, B. Production of organic acids by arbuscular mycorrhizal fungi and their contribution in the mobilization of phosphorus bound to iron oxides. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Adeleke, R.; Cloete, T.E.; Bertrand, A.; Khasa, D.P. Relationship between plant growth and organic acid exudates from ectomycorrhizal and non-ectomycorrhizal Pinus patula. S. Afr. J. Plant Soil 2015, 32, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Van Hees, P.A.W.; Rosling, A.; Essén, S.; Godbold, D.L.; Jones, D.L.; Finlay, R.D. Oxalate and ferricrocin exudation by the extramatrical mycelium of an ectomycorrhizal fungus in symbiosis with Pinus sylvestris. New Phytol. 2005, 169, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Cessna, S.G.; Sears, V.E.; Dickman, M.B.; Low, P.S. Oxalic acid, a pathogenicity factor for Sclerotinia sclerotiorum, suppresses the oxidative burst of the host plant. Plant Cell Online 2000, 12, 2191–2200. [Google Scholar] [CrossRef] [Green Version]
- Duan, S.-C.; Hu, Z.-Y.; Zhang, R.-Y. Oxalic acid is an important factor for Lentinus edodes to regulate environmental pH and promote hyphal growth. In Proceedings of the Colorful Fungi Beautiful China—2019 Academic Annual Meeting of Chinese Fungi Society, Xi’an, China, 3–5 August 2019; p. 112. [Google Scholar]
- Murphy, M.P.; O’Neill, L.A.J. Krebs Cycle reimagined: The emerging roles of succinate and itaconate as signal transducers. Cell 2018, 174, 780–784. [Google Scholar] [CrossRef] [Green Version]
- Vanderauwera, S.; Suzuki, N.; Miller, G.; van de Cotte, B.; Morsa, S.; Ravanat, J.L.; Hegie, A.; Triantaphylides, C.; Shulaev, V.; Van Montagu, M.C.; et al. Extranuclear protection of chromosomal DNA from oxidative stress. Proc. Natl. Acad. Sci. USA 2011, 108, 1711–1716. [Google Scholar] [CrossRef] [Green Version]
- Miteva-Staleva, J.; Stefanova, T.; Krumova, E.; Angelova, M. Growth-phase-related changes in reactive oxygen species generation as a cold stress response in Antarctic Penicillium strains. Biotechnol. Biotechnol. Equip. 2011, 25, 58–63. [Google Scholar] [CrossRef] [Green Version]
- van der Heide, T.; Poolman, B. Osmoregulated ABC-transport system of Lactococcus lactis senses water stress via changes in the physical state of the membrane. Proc. Natl. Acad. Sci. USA 2000, 97, 7102–7106. [Google Scholar] [CrossRef] [Green Version]
- Geisler, M.; Blakeslee, J.J.; Bouchard, R.; Lee, O.R.; Vincenzetti, V.; Bandyopadhyay, A.; Titapiwatanakun, B.; Peer, W.A.; Bailly, A.; Richards, E.L.; et al. Cellular efflux of auxin catalyzed by the Arabidopsis MDR/PGP transporter AtPGP1. Plant J. 2005, 44, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Herrou, J.; Willett, J.W.; Czyż, D.M.; Babnigg, G.; Kim, Y.; Crosson, S. Conserved ABC transport system regulated by the general stress response pathways of alpha- and Gamma proteobacteria. J. Bacteriol. 2017, 199, e00746-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.X.; Tang, H.; Wang, E.T.; Chen, W.X. An ABC transporter is required for alkaline stress and potassium transport regulation in Sinorhizobium meliloti. FEMS Microbiol. Lett. 2009, 293, 35–41. [Google Scholar] [CrossRef]
- Shabani, L.; Sabzalian, M.R.; Pour, S.M. Arbuscular mycorrhiza affects nickel translocation and expression of ABC transporter and metallothionein genes in Festuca arundinacea. Mycorrhiza 2016, 26, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Kovalchuk, A.; Kohler, A.; Martin, F.; Asiegbu, F.O. Diversity and evolution of ABC proteins in mycorrhiza-forming fungi. BMC Evol. Biol. 2015, 15, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosuke, K.; Masashi, M. ABC Transporter subfamily D: Distinct differences in behavior between ABCD1–3 and ABCD4 in subcellular localization, function, and human Disease. BioMed Res. Int. 2016, 2016, 6786245. [Google Scholar]
- Roermund, C.; Visser, W.F.; Ijlst, L.; Waterham, H.R.; Wanders, R. Differential substrate specificities of human ABCD1 and ABCD2 in peroxisomal fatty acid β-oxidation. BBA—Mol. Cell Biol. Lipids 2011, 1811, 148–152. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, T.; Zhang, P.; Pang, W.; Zhang, X.; Lian, C.; Zhang, T. Effects of High Temperature-Triggered Transcriptomics on the Physiological Adaptability of Cenococcum geophilum, an Ectomycorrhizal Fungus. Microorganisms 2022, 10, 2039. https://doi.org/10.3390/microorganisms10102039
Yan T, Zhang P, Pang W, Zhang X, Lian C, Zhang T. Effects of High Temperature-Triggered Transcriptomics on the Physiological Adaptability of Cenococcum geophilum, an Ectomycorrhizal Fungus. Microorganisms. 2022; 10(10):2039. https://doi.org/10.3390/microorganisms10102039
Chicago/Turabian StyleYan, Tianyi, Panpan Zhang, Wenbo Pang, Xiaohui Zhang, Chunlan Lian, and Taoxiang Zhang. 2022. "Effects of High Temperature-Triggered Transcriptomics on the Physiological Adaptability of Cenococcum geophilum, an Ectomycorrhizal Fungus" Microorganisms 10, no. 10: 2039. https://doi.org/10.3390/microorganisms10102039