Natural Fungicolous Regulators of Biscogniauxia destructiva sp. nov. That Causes Beech Bark Tarcrust in Southern European (Fagus sylvatica) Forests


Abstract
:1. Introduction
2. Materials and Methods
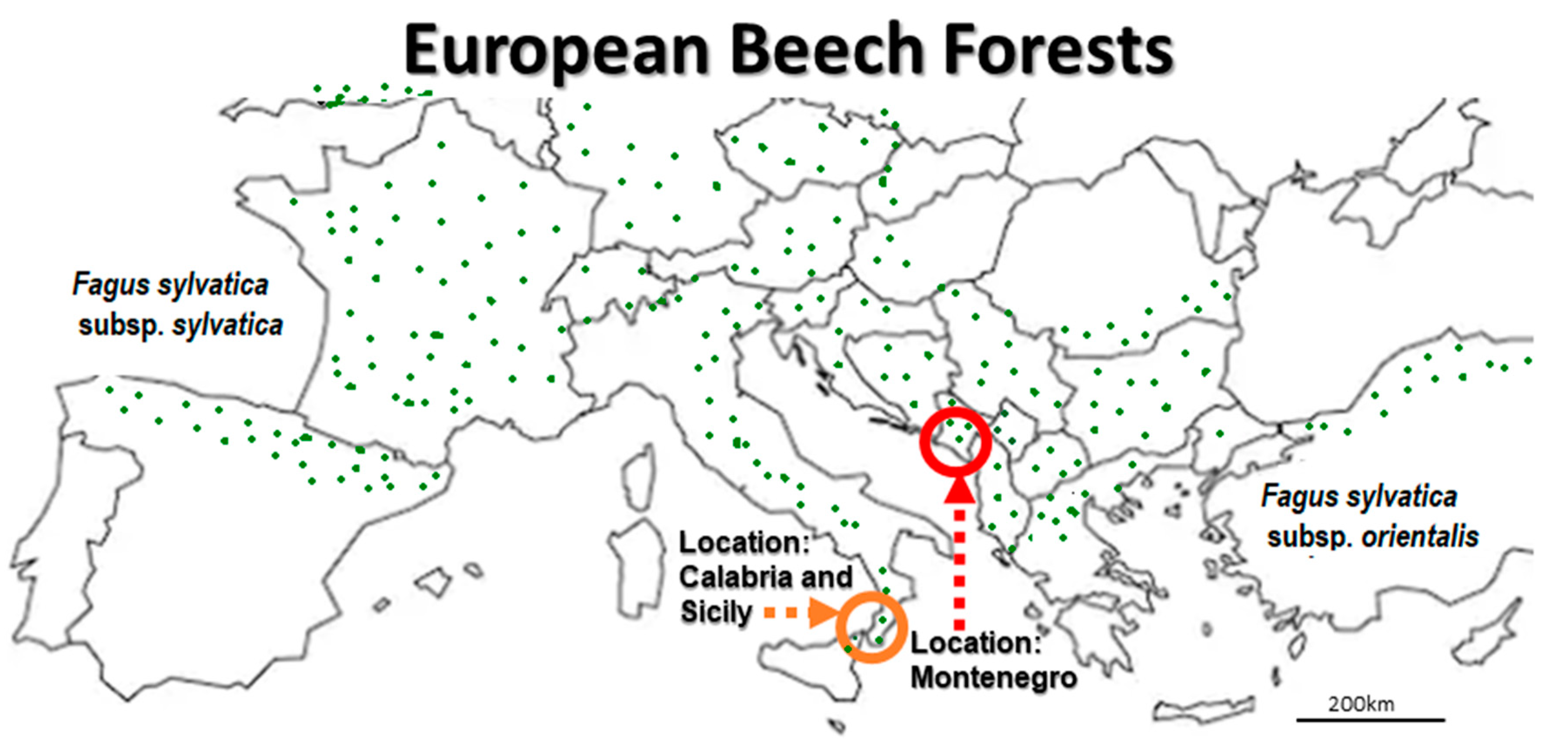
2.1. Location and Sampling
2.2. DNA Extraction, Sequencing Methods and Microscopy
3. Results
3.1. Biscogniauxia sp. Taxonomy and BTC Identity
3.2. Unique Biscogniauxia-BTC Taxon on Fagus in Montenegro
3.3. Endophytic Fungicolous Fungi, Mycoparasites and Hyperparasites
4. Discussion
4.1. Tarcrust Identity, Pathogenicity and Symptomatology
4.2. Biscogniauxia sp. nov. Evolutionary Affinity
4.3. Fungicolous Fungi
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gams, W.; Diederich, P.; Põldmaa, K. Fungicolos Fungi. In Biodiversity of Fungi: Inventory and Monitoring Methods; Bills, G.F., Foster, M.S., Eds.; Academic Press: Burlington, NJ, USA, 2004; Chapter 17; pp. 343–392. [Google Scholar] [CrossRef]
- Goh, Y.K.; Vujanovic, V. Sphaerodes quadrangularis biotrophic mycoparasitism on Fusarium avenaceum. Mycologia 2010, 102, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Chenthamara, K.; Druzhinina, I.S. 12 Ecological Genomics of Mycotrophic Fungi. In Environmental and Microbial Relationships; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2016; pp. 215–246. [Google Scholar]
- Buras, A.; Menzel, A. Projecting Tree Species Composition Changes of European Forests for 2061–2090 Under RCP 4.5 and RCP 8.5 Scenarios. Front. Plant Sci. 2019, 9, 1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vujanovic, V.; Brisson, J. Microfungal biodiversity on Fagus grandifolia in an old-growth forest of Eastern North-America. Phyton-Ann. Rei Bot. 2002, 42, 315–327. [Google Scholar]
- Granata, G.; Sidoti, A. Biscogniauxia nummularia: Pathogenic agent of a beech decline. For. Pathol. 2004, 34, 363–367. [Google Scholar] [CrossRef]
- Peters, R. Beech Forests; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1997. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi; CABI Europe: Wallingford, UK, 2008. [Google Scholar]
- Vujanovic, V.; Vuckovic, M. Die Lage Gefährdung der Wälder im National Park Lovćen. Acad. Sci. Arts Monten. J. Sci. 1994, 34, 175–183. [Google Scholar]
- Thomas, S. Fungi and Their Potential as Biological Control Agents of Beech Bark Disease. Ph.D. Thesis, Royal Holloway, University of London, London, UK, 2014. [Google Scholar]
- Herrera, C.S.; Hirooka, Y.; Chaverri, P. Pseudocospeciation of the mycoparasite Cosmospora with their fungal hosts. Ecol. Evol. 2016, 6, 1504–1514. [Google Scholar] [CrossRef] [Green Version]
- Vujanovic, V.; Neumann, P.; Laflamme, G. A new record of Cosmospora episphaeria hyperparasitic on hypoxylon species from American beech. Mycotaxon 2003, 87, 249–253. [Google Scholar]
- Sun, J.-Z.; Liu, X.; McKenzie, E.H.C.; Jeewon, R.; Liu, J.-K. (Jack); Zhang, X.-L.; Zhao, Q.; Hyde, K.D. Fungicolous fungi: Terminology, diversity, distribution, evolution, and species checklist. Fungal Divers. 2019, 95, 337–430. [Google Scholar] [CrossRef]
- Arnold, G.R.W. Cladobotryum campanisporum. Feddes Repert. Spec. Nov. Regni Veg. 1987, 98, 352. [Google Scholar]
- Samuels, G.J. Fungicolous, Lichenicolous, and Myxomyceticolous-Species of Hypocreopsis, Nectriopsis, Nectria, Peristomialis, and Trichonectria. Mem. N. Y. Bot. Gard. 1988, 48, 1–78. [Google Scholar]
- Bandoni, R.J. On some species of Mycogloea. Mycoscience 1998, 39, 31–36. [Google Scholar] [CrossRef]
- Chlebicki, A.; Skirgiełło, A. Some mycogenous fungi from Poland. Acta Mycol. 2014, 30, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Ellis, M.B.; Ellis, J.P. Microfungi on Miscellaneous Substrates: An Identification Handbook; Croom Helm: London, UK, 1988; p. vii+244. [Google Scholar]
- Helfer, W. Pilze auf Pilzfruchtkorpern. Untersuchungen zur Okologie, Systematik und Chemie. Libri Bot 1991, 1, 1–157. [Google Scholar]
- Karadžić, D.; Vujanović, V. Bolesti bukovih sastojina na području nacionalnog parka “Lovćen”. CANU Naučni Skupovi 1994, 34, 175–183. [Google Scholar]
- Vujanovic, V.; Goh, Y.K. Sphaerodes mycoparasitica sp. nov., a new biotrophic mycoparasite on Fusarium avenaceum, F. graminearum and F. oxysporum. Mycol. Res. 2009, 113, 1172–1180. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.; Taylor, J. 38-Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Vujanovic, V.; Hamelin, R.; Bernier, L.; Vujanovic, G.; St-Arnaud, M. Fungal diversity, dominance, and community structure in the rhizosphere of clonal Picea mariana plants throughout nursery production chronosequences. Microb. Ecol. 2007, 54, 672–684. [Google Scholar] [CrossRef]
- Jaklitsch, W.; Fournier, J.; Rogers, J.; Voglmayr, H. Phylogenetic and taxonomic revision of Lopadostoma. Persoonia Mol. Phylogeny Evol. Fungi 2014, 32, 52–82. [Google Scholar] [CrossRef] [Green Version]
- Rzhetsky, A.; Nei, M. A simple method for estimating and testing minimum-evolution trees. Mol. Biol. Evol. 1992, 9, 945. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Fournier, J.; Magni, J.F. Pyrenomycetes from southwestern France. Section: Systematics and Dichotomous Key to European Biscogniauxia. Available online: http://pyrenomycetes.free.fr/biscogniauxia/ (accessed on 20 January 2019).
- Rogers, J.; Ju, Y.-M.; Candoussau, F. Biscogniauxia anceps comb. nov. and Vivantia guadalupensis gen. et sp. nov. Mycol. Res. 1996, 100, 669–674. [Google Scholar] [CrossRef]
- U′ren, J.M.; Miadlikowska, J.; Zimmerman, N.B.; Lutzoni, F.; Stajich, J.E.; Arnold, A.E. Contributions of North American endophytes to the phylogeny, ecology, and taxonomy of Xylariaceae (Sordariomycetes, Ascomycota). Mol. Phylogenetics Evol. 2016, 98, 210–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, Y.M.; Rogers, J.D.; San Martín, F.; Granmo, A. The genus Biscogniauxia. Mycotaxon 1998, 66, 1–98. [Google Scholar]
- Callan, B.E.; Rogers, J.D. Cultural characters and anamorphs of Biscogniauxia (= Nummularia) marginata, B. dennisii, and B. repanda. Can. J. Bot. 1986, 64, 842–847. [Google Scholar] [CrossRef]
- Chaverri, P.; Salgado, C.; Hirooka, Y.; Rossman, A.; Samuels, G. Delimitation of Neonectria and Cylindrocarpon (Nectriaceae, Hypocreales, Ascomycota) and related genera with Cylindrocarpon-like anamorphs. Stud. Mycol. 2011, 68, 57–78. [Google Scholar] [CrossRef]
- Sun, J.-Z.; Liu, X.-Z.; Hyde, K.D.; Zhao, Q.; Maharachchikumbura, S.S.N.; Camporesi, E.; Bhat, J.; Nilthong, S.; Lumyong, S. Calcarisporium xylariicola sp. nov. and introduction of Calcarisporiaceae fam. nov. in Hypocreales. Mycol. Prog. 2017, 16, 433–445. [Google Scholar] [CrossRef]
- Pouzar, Z. A key and conspectus of central European species of Biscogniauxia and Obalarina (Pyrenomycetes). Ceska Mykologie 1986, 40, 1–10. [Google Scholar]
- Wendt, L.; Sir, E.B.; Kuhnertm, E.; Heitkämper, S.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Luangsa-ard, J.J.; Srikitikulchai, P.; Peršoh, D.; et al. Resurrection and emendation of the Hypoxylaceae, recognised from a multi-gene genealogy of the Xylariales. Mycol. Prog. 2018, 17, 115–154. [Google Scholar] [CrossRef] [Green Version]
- Jong, S.C.; Rogers, J.D. Illustrations and descriptions of conidial states of some Hypoxylon species. Washington Agricultural Experiment Station. Tech. Bull. 1972, 71, 1–51. [Google Scholar]
- Collado, J.; Platas, G.; Peláez, F. Identification of an endophytic Nodulisporium sp. from Quercus ilex in Central Spain as the anamorph of Biscogniauxia mediterranea by rDNA sequence analysis and effect of different ecological factors on distribution of the fungus. Mycologia 2001, 93, 875. [Google Scholar] [CrossRef]
- Vujanović, V. Istraživanja nekih biotičkih i abiotičkih faktora kao mogućih prouzrokovača sušenja šuma u Crnoj Gori. Master’s Thesis, Belgrade University, Faculty of Forestry, Belgrade, Serbia, 1990. [Google Scholar]
- Luchi, N.; Capretti, P.; Feducci, M.; Vannini, A.; Ceccarelli, B.; Vettraino, A. Latent infection of Biscogniauxia nummularia in Fagus sylvatica: A possible bioindicator of beech health conditions. iFor. Biogeosci. For. 2016, 9, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Scholtysik, A.; Unterseher, M.; Otto, P.; Wirth, C. Spatio-temporal dynamics of endophyte diversity in the canopy of European ash (Fraxinus excelsior). Mycol. Prog. 2013, 12, 291–304. [Google Scholar] [CrossRef]
- Zabalgogeazcoa, I.; Pedro, J.; Canals, R.-M. Biscogniauxia nummulariainfecting beech (Fagus sylvatica) trees and sympatric plants of the sedge Carex brevicollis. For. Pathol. 2015, 45, 346–348. [Google Scholar] [CrossRef]
- Vannini, A.; Mazzaglia, A.; Anselmi, N. Use of random amplified polymorphic DNA (RAPD) for detection of genetic variation and proof of the heterothallic mating system in Hypoxylon mediterraneum. For. Pathol. 1999, 29, 209–218. [Google Scholar] [CrossRef]
- Lakatos, F.; Molnár, M. Mass mortality of Beech (Fagus sylvatica L.) in South-West Hungary. Acta Silv. Lignaria Hung. 2009, 5, 75–82. [Google Scholar]
- Stukenbrock, E.H. The Role of Hybridization in the evolution and emergence of new fungal plant pathogens. Phytopathology 2016, 106, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.G. Form and function of fungal spore appendages. Mycoscience 2006, 47, 167–183. [Google Scholar] [CrossRef]
- Kandemir, G.; Kaya, Z. Oriental beech—Fagus orientalis. In EUFORGEN Technical Guidelines for Genetic Conservation and Use; EUFORGEN—European Forest Genetic Resources Programme Bioversity International: Rome, Italy, 2009; p. 6. [Google Scholar]
- Boddy, L.; Heilmann-Clausen, J. Chapter 12 Basidiomycete Community Development in Temperate Angiosperm Wood; Elsevier Academic Press: London, UK, 2008; Volume 28, pp. 211–237. [Google Scholar]
- Tudor, D.; Robinson, S.C.; Cooper, P.A. The influence of moisture content variation on fungal pigment formation in spalted wood. AMB Express 2012, 2, 69. [Google Scholar] [CrossRef] [Green Version]
- Vujanovic, V.; Goh, Y.K. Mycoparasites of Fusarium Pathogens on Wheat: From Taxonomy, Genomics and Proteomics to Biotechnology; Almeida, M.T., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2011; pp. 297–314. [Google Scholar]
- Shim, S.; Sy, A.A.; Wicklow, D. Isolation of an isocoumarin and an isobenzofuran derivatives from a fungicolous isolate of Acremonium crotocinigenum. Bull. Korean Chem. Soc. 2008, 29, 863–865. [Google Scholar] [CrossRef] [Green Version]
- Summerbell, R.C.; Gueidan, C.; Guarro, J.; Eskalen, A.; Crous, P.; Gupta, A.K.; Gené, J.; Cano-Lira, J.F.; Van Iperen, A.; Starink, M.; et al. The protean acremonium. A. sclerotigenum/egyptiacum: Revision, food contaminant, and human disease. Microorganisms 2018, 6, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-Varas, V.V.; Freeman, T.A.; Gudmestad, N.; Secor, G. Mycoparasitism of Helminthosporium solani by Acremonium strictum. Phytopathology 2007, 97, 1331–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mijuskovic, M. Calonestria gymnosporangii Jaap [Bactridium gymnosporangii (Jaap) Wr.] kao superparazit na Gymnosporangium confusum Plowr. u Crnoj Gori. Zastita Bilja 1976, 27, 245–251. [Google Scholar]
- Knapp, D.G.; Kovács, G.; Zajta, E.; Groenewald, J.; Crous, P. Dark septate endophytic pleosporalean genera from semiarid areas. Persoonia Mol. Phylogeny Evol. Fungi 2015, 35, 87–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suryanarayanan, T.S.; Devarajan, P.T.; Girivasan, K.P.; Govindarajulu, M.B.; Kumaresan, V.; Murali, T.S.; Rajamani, T.; Thirunavukkarasu, N.; Venkatesan, G. The host range of multi-host endophytic fungi. Curr. Sci. 2018, 115, 1963–1969. [Google Scholar] [CrossRef]
- Samuels, G.J.; Candoussau, F.; Magni, J.-F. Fungicolous Pyrenomycetes 1. Helminthosphaeria and the new family Helminthosphaeriaceae. Mycolology 1997, 89, 141. [Google Scholar] [CrossRef] [Green Version]
- Kirschner, R.; Piepenbring, M. A new fungicolous species of Titaea and new reports of Bahusaganda indica and Exosporium ampullaceum (hyphomycetes) from tropical rainforests in Panama. Fungal Divers 2006, 21, 93–103. [Google Scholar]
- Guatimosim, E.; Pinto, H.J.; Barreto, R.W. Passalora acrocomiae sp. nov. and Exosporium acrocomiae from the palm Acrocomia aculeata in Puerto Rico. Mycotaxon 2013, 122, 61–67. [Google Scholar] [CrossRef]
- Crous, P.; Crous, P.W.; Nakashima, C. Cercosporoid fungi (Mycosphaerellaceae) 2. Species on monocots (Acoraceae to Xyridaceae, excluding Poaceae). IMA Fungus 2014, 5, 203–390. [Google Scholar] [CrossRef]
- El Shafie, A.; Webster, J. Curvularia species as parasites of Rhizopus and other fungi. Trans. Br. Mycol. Soc. 1979, 73, 352–353. [Google Scholar] [CrossRef]
- Põldmaa, K.; Samuels, G.J. Fungicolous hypocreaceae (Ascomycota: Hypocreales) from Khao Yai National Park, Thailand. Sydowia 2004, 56, 79–130. [Google Scholar]
- Döbbeler, P. Ascospore diversity of bryophilous Hypocreales and two new hepaticolous Nectria species. Mycologia 2005, 97, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Barr, M.E. On the family Tubeufiaceae (Pleosporales). Mycotaxon 1980, 12, 137–167. [Google Scholar]
- Boom, P.P.G.V.D.; Palice, Z. Some interesting lichens and lichenicolous fungi from the Czech Republic. Czech Mycol. 2006, 58, 105–116. [Google Scholar] [CrossRef]
- Hashimoto, A.; Matsumura, M.; Hirayama, K.; Tanaka, K. Revision of Lophiotremataceae (Pleosporales, Dothideomycetes): Aquasubmersaceae, Cryptocoryneaceae, and Hermatomycetaceae fam. nov. Persoonia Mol. Phylogeny Evol. Fungi 2017, 39, 51–73. [Google Scholar] [CrossRef] [PubMed]
- Mathiassen, G.; Granmo, A.; Stensrud, Ø. Lophiotrema borealiforme, a new species close to L. boreale. Karstenia 2017, 57, 11–15. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, H.K.; Fournier, J.; Crous, P.W.; Jeewon, R.; Pointing, S.B.; Hyde, K.D. Towards a phylogenetic clarification of Lophiostoma/Massarina and morphologically similar genera in the Pleosporales. Fungal Divers. 2009, 38, 225–251. [Google Scholar]
- Tane, C.; Barbu, L.; Rodriguez, S.; Cojanu, D.; Andreea, C. One strain of endophytic Preussia, a potential biological partner of tomato seedlings against alternariosis. Biologie 2019, 28, 15–19. [Google Scholar]
- Menkis, A.; Povilaitienė, A.; Marčiulynas, A.; Lynikienė, J.; Gedminas, A.; Marčiulynienė, D. Occurrence of common phyllosphere fungi of horse-chestnut (Aesculus hippocastanum) is unrelated to degree of damage by leafminer (Cameraria ohridella). Scand. J. For. Res. 2018, 34, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Din, Z.U.; De Medeiros, L.; Abreu, L.; Pfenning, L.H.; Jymeni, D.L.; Rodrigues-Filho, E. Differential metabolism of diastereoisomeric diterpenes by Preussia minima, found as endophytic fungus in Cupressus lusitanica. Bioorganic Chem. 2018, 78, 436–443. [Google Scholar] [CrossRef]
- Zaferanloo, B.; Bhattacharjee, S.; Ghorbani, M.M.; Mahon, P.J.; Palombo, E.A. Amylase production by Preussia minima, a fungus of endophytic origin: Optimization of fermentation conditions and analysis of fungal secretome by LC-MS. BMC Microbiol. 2014, 14, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mapperson, R.R.; Kotiw, M.; Davis, R.A.; Dearnaley, J.D.W. The Diversity and antimicrobial activity of Preussia sp. endophytes isolated from Australian dry rainforests. Curr. Microbiol. 2013, 68, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Currah, R.; Sherburne, R. Septal ultrastructure of some fungal endophytes from boreal orchid mycorrhizas. Mycol. Res. 1992, 96, 583–587. [Google Scholar] [CrossRef]
- Bartnik, C.; Michalcewicz, J.; Ledwich, D.; Ciach, M. Mycobiota of dead Ulmus glabra wood as breeding material for the endangered Rosalia alpina (Coleoptera: Cerambycidae). Pol. J. Ecol. 2020, 68, 13–22. [Google Scholar] [CrossRef]
- Bubák, F. Ein beitrag zur Pilzflora von Montenegro; Sitzungsberichte der Königlichen Böhmischen Gesellschaft der Wissenschaften: Praque, Czech Republic, 1903; p. 22. [Google Scholar]
- Bubák, F. Zweiter Beitrag zur Pilzflora von Montenegro. Bull. l’Herbier Boissier Série 1906, 26, 393–408, 473–488. [Google Scholar]
- Bubák, F. Dritter Beitrag zur Pilzflora von Montenegro [Third work on the fungus flora of Montenegro]. Botanikai Közlemények 1915, 14, 39–83. [Google Scholar]
- Bubák, F. Adatok Montenegro gombaflóràjahóz (Dritter Beitrag zur Pilzflora von Montenegro). Botanikai Közlemények 1915, 14, 98–99. [Google Scholar]
- Jaap, O. Beiträge zur Kenntniss der Pilze Dalmatiens. Ann. Mycol. 1916, 14, 1–44. [Google Scholar]
- Mijušković, M.; Vučinić, Z. Hyperparasitic Fungi in Mycopopulation of Montenegro. In Series: Mycologia Applicata; Montenegrien Mycological Center: Podgorica, Montenegro, 2001; pp. 1–108. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Xylariaceous Fungus | Mycoparasite/Fungicolous Fungus | Origin [Ref] |
|---|---|---|
| Xylariaceae Tul. & C. Tul. (Hypoxylon, Daldinia, Rosellinia, and Xylaria) | Acanthostigmella brevispina M.E. Barr & Rogerson | USA, UK [13] |
| Cladobotryum campanisporum G.R.W. Arnold | Cuba [4,14] | |
| Hypocrea xylariicola Henn | Brazil [5,15] | |
| Muroia nipponica I. Hino & Katum. | Japan [16] | |
| Cytoplea parasitica Petr., Feddes Repert | Pakistan [13] | |
| Tubeufia brevispina (M.E. Barr & Rogerson) J.L. Crane | USA [3] | |
| Tubeufia cerea (Berk. & M.A. Curtis) Höhn. | China [13], Poland [17] | |
| Phragmogibbera xylariicola Samuels & Rogerson | Venezuela [13] | |
| Capronia moravica (Petr.) E. Müll., Petrini, P.J. Fisher, Samuels & Rossman | USA [13] | |
| Capronia parasitica (Ellis & Everh E. Müll., Petrini, P.J. Fisher, Samuels & Rossman | USA [13] | |
| Calcarisporium xylariicola Jing Z. | Italy [3] | |
| Neobarya xylariicola Cand., J.D. | USA [3] | |
| Polydesmia pruinosa (Berk. & Broome) Boud. | France [18] | |
| Hyphozyma lignicola L.J. Hutchison | Canada [3] | |
| Orbilia inflatula (P. Karst.) P. Karst. | Germany [19] | |
| Hydropisphaera hypoxantha (Penz. & Sacc.) Rossman & Samuels | China [3] | |
| Atkinsonella hypoxylon (Peck) Diehl | USA [3] | |
| Neobarya lutea Samuels & Lodge | Puerto Rico [3] | |
| Neobarya xylariicola Cand., J.D. Rogers & Samuels | USA, France [3] | |
| Hypocreopsis xylariicola Samuels | Europe [3] | |
| Trichoderma hypoxylon Jing Z. | Guyana [13] | |
| Trichoderma stilbohypoxyli Samuels & Schroers | Thailand [13] | |
| Cosmospora arxii (W. Gams) Gräfenhan & Schroers | Puerto Rico [13] | |
| Cosmospora vilior (Starbäck) Rossman & Samuels | Canada [20] | |
| Cosmospora episphaeria (Tode) Rossman & Samuels Nectria geastroides Samuels | Germany [3] | |
| Nectria viliuscula Samuels, Yoshim. Doi & Rogerson | Worldwide [21] | |
| Phaeoacremonium sp. W. Gams, Crous et M. J. Wingf. | Amazonas [13] | |
| Chlorostroma cyaninum Læssøe, Srikit. & J. Fourn. | Indonesia [3] | |
| Chlorostroma subcubisporum A.N. Mill., Lar. N. | USA [3] | |
| Vassiljeva & J.D. Rogers | Thailand [3] | |
| Acrostaphylus hypoxyli G. Arnaud | USA [3] | |
| Refractohilum mycophilum Castañeda, W.B. Kendr. & Guarro | France [3] | |
| Xenasma aculeatum C.E. Gómez | Cuba [3] | |
| Tremella flava Chee J. Chen | Argentina [3] | |
| Tremella resupinata Chee J. Chen | China [13] | |
| Tremella menglunensis Y.B. Peng | China [13] | |
| Mycogloea nipponica Bandoni | China [13] | |
| Immotthia hypoxylon (Ellis & Everh.) M.E. Barr. | Japan [3], Poland [17] | |
| Diatrypaceae Nitschke (Diatrype, Diatrypetala, Eutype and Eutypella) | Capronia nigerrima (R.R. Bloxam) M.E. Barr | UK [13] |
| Deltosperma infundibuliforme W.Y. Zhuang | Trinidad-Tobago [13] | |
| Chaetosphaeria phaeostroma (Durieu & Mont.) Fuckel | Europe [18] | |
| Chaetosphaerella fusca (Fuckel) E. Müll. & C. Booth | Europe [18] | |
| Chaetosphaerella phaeostroma (Durieu & Mont.) E. Müll. & C. Booth | Europe [8] | |
| Nectria pseudepisphaeria Samuels | USA [13] | |
| Nectria triqua Samuels | French Guiana [3] | |
| Ophiostoma grande Samuels & E. Müll. | Brazil [3] | |
| Diplococcium heterosporum L. Zeller & Tóth | Hungary [13] | |
| Endophragmiella eboracensis B. Sutton | Canada [13] | |
| Annellodochium ramulisporum Deighton | Sierra Leone [13] | |
| Tomentella badiofusca f. diatrypicola Svrček, | Czechoslovakia [13] | |
| Sirobasidium sandwicense Gilb. & Adask. | USA [13] | |
| Tremella episphaerica Rick | China [13] | |
| Tremella nivalis Chee J. Chen | China [13] | |
| Achroomyces henricii P. Roberts | USA [3] | |
| Chloridium clavaeformae (Pr.) Gams et Hol.-Jech. | Poland [17] | |
| Other | Ampelomyces sp. Ces. | Canada [5] |
| Aureobasidium pullulans (de Bary) G. Arnaud | ||
| Gliocladium sp. Corda | ||
| Cladosporium herbarum (Pers.: Fr.) Link | ||
| Cladosporum cladosporioides (Pers.) Link | ||
| Epicoccum nigrum Link | ||
| Leptographium microsporum R.W. Davidson | ||
| Paecilomyces farinosus (Holm.) Brown & SM. | ||
| Trichoderma harzianum Rifai | ||
| Trichoderma viride Pers.: Fr. | ||
| Verticillium lecanii (Zimm.) Viegas | ||
| Scolicosporium sp. Lib. ex Roum. |
| Biscogniauxia Taxon | Anceps | Nummularia | Destructiva |
|---|---|---|---|
| Stroma shape | applanate, irregularly orbicular 5–25 mm diam | applanate, discoid 5–20 mm diam | applanate, discoid 5–30 mm diam |
| Stroma colour | dull black, carbonaceous | black, carbonaceous | black, carbonaceous |
| Ostioles | umbilicate | umbilicate or papillate | umbilicate or papillate |
| Ascospore septation | two-celled | one-celled | one or two-celled |
| Ascospore colour | hyaline (abundant) and brown or blackish brown | hyaline (rare) and brown | Non-hyaline, brown and blackish brown |
| Ascospore shape | Sub-globose, broadly ellipsoid to deltoid (larger-dark cell) with cuneate extrusion (inferior, smaller-less colored membranous cell) | narrowly ellipsoid, ellipsoid to fusiform, without appendage | ovoid, ellipsoid to deltoid (larger-dark cell) and heap shaped extrusion (inferior, smaller-less colored membranous cell) |
| Ascospore size | 13–16 (18) × 7–8 (9) µm | 11.5–13.5(16) × 8.–9.5 (11) µm | 12–16 (18) × 8–11 (12) µm |
| Appendage | Straight, stick-like appendage with narrow-hyaline apex, 2–3 µm long | no appendage | Curved, thread-like appendage with a broadened-dark apex, 3–5 µm long. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vujanovic, V.; Kim, S.H.; Latinovic, J.; Latinovic, N. Natural Fungicolous Regulators of Biscogniauxia destructiva sp. nov. That Causes Beech Bark Tarcrust in Southern European (Fagus sylvatica) Forests. Microorganisms 2020, 8, 1999. https://doi.org/10.3390/microorganisms8121999
Vujanovic V, Kim SH, Latinovic J, Latinovic N. Natural Fungicolous Regulators of Biscogniauxia destructiva sp. nov. That Causes Beech Bark Tarcrust in Southern European (Fagus sylvatica) Forests. Microorganisms. 2020; 8(12):1999. https://doi.org/10.3390/microorganisms8121999
Chicago/Turabian StyleVujanovic, Vladimir, Seon Hwa Kim, Jelena Latinovic, and Nedeljko Latinovic. 2020. "Natural Fungicolous Regulators of Biscogniauxia destructiva sp. nov. That Causes Beech Bark Tarcrust in Southern European (Fagus sylvatica) Forests" Microorganisms 8, no. 12: 1999. https://doi.org/10.3390/microorganisms8121999