Taxonomy
As the species of
Diplogelasinospora were located in a well-supported clade (clade I, 100% bs/1 pp;
Figure 1) distinct from the Sordariaceae (clade III), and because the morphology and ornamentation (i.e., inwardly pitted upper and lower cell) of the ascospores are unique in the Lasiosphaeriaceae s. lat., we propose the new family Diplogelasinosporaceae to accommodate this genus.
Diplogelasinosporaceae Y. Marin & Stchigel, fam. nov. MycoBank MB835495.
Etymology: Named for Diplogelasinospora, the type genus of this family.
Type genus: Diplogelasinospora Cain, Can. J. Bot. 39: 1669. 1961.
Ascomata non-ostiolate, immersed or superficial, scattered or in groups, dark brown to nearly black, spherical, non-stromatic, sometimes covered by hyphae-like setae; ascomatal wall membranaceous to slightly coriaceous, textura angularis, occasionally cephalothecoid or outer layer of textura intricata, dark brown, opaque. Asci unitunicate, eight-spored, cylindrical to cylindrical-clavate, short stipitate, lacking a distinct apical ring, arranged in parallel fascicles, evanescent. Paraphyses present, cylindrical to moniliform, septate and sometimes constricted at the septa. Ascospores at first hyaline, aseptate, ellipsoidal, becoming transversely septate and two-celled, one cell remaining hyaline, one becoming black and opaque, wall smooth or pitted showing an endodentate endosporium, sometimes with a thin gelatinous sheath. Asexual morph absent or present. Conidiophores lacking or not distinctly differentiated from vegetative hyphae. Conidia arthric, terminal or intercalary, hyaline, oblong to cylindrical, with both ends truncated, smooth-walled, sometimes sliding into a slimy mass; conidia blastic, borne singly and laterally on aerial hyphae, hyaline, ovate to elongate, occasionally subspherical to somewhat elongate, smooth-walled.
Notes: This family includes only species of the genus
Diplogelasinospora, which was originally in Neurosporaceae (syn. Sordariaceae) based on the similar ascospore ornamentation of the species belonging to the genus
Neurospora (formerly
Gelasinospora) [
37]. Cai et al. [
33] observed that this genus did not group in the Sordariaceae and demonstrated that it was more related to the Lasiosphaeriaceae. In our phylogenetic study, this genus formed a well-supported clade distinct from the other families of the Sordariales. Diplogelasinosporaceae can be distinguished from other members of the Lasiosphaeriaceae s. lat., Naviculisporaceae, Schizotheciaceae, and Podosporaceae by the presence of an asexual morph characterized by the production of arthroconidia. A similar asexual morph has been observed in Chaetomiaceae and Sordariaceae. However, the latter ones produce one-celled ascospores, while ascospores are two-celled in Diplogelasinosporaceae.
The type species of the genus Lasiosphaeria (L. ovina) was placed into a well-supported clade (clade V, 95 bs/1 pp) together with other spp. of that genus, Corylomyces selenospora and Bellojisia rhychostoma, and several species of Anopodium, Cercophora, Podospora, and Zopfiella. As the rest of the members of Lasiophaeriaceae fall into other distinct clades, we restrict the circumscription of the family Lasiosphaeriaceae as described below.
Lasiosphaeriaceae Nannf., Nova Acta R. Soc. Scient. upsal., Ser. 4 8(no. 2): 50, 1932, emend. Y. Marin & Stchigel. MycoBank MB80930.
Type genus: Lasiosphaeria Ces. & De Not., Comment. Soc. Crittog. Ital. 1 (fasc. 4): 229. 1863.
Ascomata mostly ostiolate, immersed to superficial, scattered or aggregated, olivaceous yellow to dark brown, pyriform, globose, subglobose or ovoid, covered with white to variously colored (but not dark) tomentum below the neck, or with hyaline, light brown or olive, sometimes rigid, septate hairs on the neck; neck short or long, black or nearly so, cylindrical to conical; non-ostiolate ascomata globose, covered with yellowish to olivaceous-brown hyphae-like hairs; ascomatal wall membranaceous to coriaceous, textura angularis to textura intricata, weakly cephalothecoid in non-ostiolate ascomata. Paraphyses absent or present, filiform. Asci unitunicate, four- or eight-spored, cylindrical to cylindrical-clavate or clavate, short- or long-stipitate, with an apical ring, distinct or inconspicuous, or with an apical differentiation refractive and non-amyloid, usually with a subapical globule, or without apical differentiations. Ascospores uniseriate or biseriate, one- or two-celled. One-celled ascospores hyaline to yellowish, sometimes turning pale brown with age, or at first hyaline and then becoming dark brown, cylindrical, irregularly ellipsoidal, reniform, or navicular, mostly sigmoid, geniculate or curved, sometimes with an umbonate germ pore, with or without gelatinous appendages, one end may become swollen and turn brown. Two-celled ascospores at first hyaline and one-celled, becoming transversely septate; upper cell olivaceous to dark brown, ellipsoidal or vermiform to clavate, with an apical or subapical germ pore, sometimes later developing a transversal septum; lower cell hyaline, cylindrical, collapsing or not; gelatinous cauda sometimes present, lash-like, attached at both ends. Asexual morph absent or present. Conidiophores reduced to conidiogenous cells. Conidiogenous cells phialidic, monophialidic, or sometimes polyphialidic, produced laterally or terminally, delimited by a basal septum, obclavate to lageniform, hyaline to pale brown, constricted below the collarette. Conidia enteroblastic, aggregated in slimy heads, hyaline, pyriform, truncate at base; conidia holoblastic absent or present, produced directly from hyphae or lateral branches not delimited by a basal septum and without collarette, hyaline, subglobose, pyriform, or obclavate.
Notes: Lasiosphaeriaceae s. str. was divided in two main clades, one (100% bs/1 pp) encompassing only species of the genus
Lasiosphaeria (including the type species of the genus,
L. ovina, and
L. glabrata,
L. lanuginosa,
L. rugulosa and
L. sorbina), and a second one (100% bs/1 pp) containing the type species of
Zopfiella and other taxa producing ascospores with a septate upper cell (with the exception of
Anopodium ampullaceum [
38]). Further studies are needed to clarify if the genus
Zopfiella should be restricted to this second clade. This family is characterized by the production of (mostly) ostiolate ascomata with a tomentose ascomatal wall or bearing septate hairs on or below the neck. The family includes taxa producing one- and two-celled ascospores. Within the first group, a high morphological variability of the ascospores can be found, those of
Lasiosphaeria being cylindrical and sigmoid, geniculate or curved [
10] vs. reniform to navicular in
Bellojisia rhynchostoma and
Corylomyces selenospora [
28,
31].
In order to accommodate three new genera, i.e., Areotheca, Naviculispora, and Rhypophila, as well as some species of Arnium, Triangularia, and Zopfiella, which form a monophyletic clade (clade IX, 100% bs/1 pp) distinct from Lasiosphaeriaceae s. str., we propose the erection of the new family Naviculisporaceae as follows.
Naviculisporaceae Y. Marin & Stchigel, fam. nov. MycoBank MB835496.
Etymology: Named after Naviculispora, the type genus of this family.
Type genus: Naviculispora Stchigel, Y. Marín, Cano & Guarro.
Ascomata ostiolate, less frequently non-ostiolate, immersed or superficial, scattered or aggregated, brown to dark brown or black, globose to subglobose, pyriform or ovate, glabrous or covered with flexous or stiff hairs; neck short or long, dark brown to blackish, conical or cylindrical, papillate, rarely glabrous, mostly with elongate tuberculated projections at the base or covered with small black papillae; ascomatal wall membranaceous, pseudoparenchymatous, semi-translucent, yellowish to light brown or olivaceous brown, textura angularis or irregularly-shaped cells, or blackish brown, opaque, areolate, cracking in polyhedral plates when crushed. Paraphyses absent or present. Asci unitunicate, 8- to 256-spored, cylindrical or clavate, evanescent, long- or short-stipitate, without apical ring, or apical ring indistinct or distinct, small, subapical globulus or chamber absent or present. Ascospores biseriate or irregularly arranged into the asci, mostly two-celled. One-celled ascospores at first hyaline, becoming ochraceous to brown or blackish brown when mature, ellipsoidal, with somewhat pointed ends, smooth-walled, provided at each end with a germ pore and a lash-like gelatinous cauda. Two-celled ascospores at first one-celled and hyaline, clavate, cylindrical or spatuliform, sometimes vermiform, becoming transversely septate and two-celled; upper cell olivaceous or brown to dark brown, navicular, conical, ellipsoidal, ellipsoidal-fusiform, limoniform, pyriform or obclavate, smooth-walled, septate, with an apical or subapical germ pore, sometimes with one or more transverse septa; lower cell hyaline or pale brown, rarely dark brown, hemispherical, cylindrical, cylindrical-obclavate, or cylindrical-conical, thick- or thin-walled, sometimes with the same length or longer than the upper cell, collapsing or not with age, rarely with 3-5 septa; several secondary appendages sometimes present, small, at the base; apical gelatinous cauda absent or present, lash-like, fibrillate or lamellate. Asexual morph absent or present. Conidiphores reduced to conidiogenous cells or sympodially proliferating. Conidiogenous cells phialidic. Conidia holoblastic, sometimes sessile, hyaline to subhyaline, ellipsoidal to subspherical, obovoid or clavate.
Notes: The family shows a high morphological variability:
Arnium caballinum,
A. japonense, and
A. mendax produce one-celled ascospores [
9,
39], while in the rest of the species of the family, these are two-celled. Taxa with both kinds of ascospores are also observed in the family Lasiosphaeriaceae s.str. as well as in Podosporaceae and Schizotheciaceae. The ascomata are mostly ostiolate, except in
Z. marina,
Z. pilifera, and
Z. submersa, in which they are non-ostiolate [
40,
41,
42]. In our phylogenetic study, these three species with non-ostiolate ascomata grouped in a well-supported clade (100 bs/1 pp) together with
T. mangenotii, which produces ostiolate ascomata [
43], suggesting that these could represent a new genus. Further molecular and phenotypic studies are needed to confirm this hypothesis. The ascomatal wall is mostly membranaceous, except in the new genus
Areotheca (introduced below), which is characterized by an areolate ascomatal wall. Among the Sordariales, this type of ascomatal wall is only seen in two species of
Cladorrhinum, i.e.,
Cl. coprophilum and
Cl. tomentosum. It is also found in a few other
Cercophora species, i.e.,
C. caerulea,
C. septentrionalis, and
C. sylvatica, but their walls are not as carbonaceous as in
Areotheca, in which the fully developed carbonaceous wall breaks into polyhedral plates when crushed.
Within the Naviculisporaceae, reference strains of
C. ambigua and
C. areolata formed a well-supported clade (100 bs/1 pp;
Figure 2). Both species are characterized by a well-developed carbonaceous, areolate ascomatal wall that cracks into polyhedral plates when crushed (cephalothecoid). The new genus
Areotheca is proposed here to accommodate these two species.
Areotheca Y. Marín & Stchigel, gen. nov. MycoBank MB835497. (
Figure 3A–D).
Type species: Areotheca ambigua (R. Hilber) Y. Marín & Stchigel.
Etymology: The name refers to the areolate ascomatal wall.
Ascomata ostiolate, scattered or aggregated, superficial, ovoid, glabrous or covered with flexuous, septate hairs; neck conical, ridged, carbonaceous; ascomatal wall pseudoparenchymatous, blackish brown, carbonaceous, opaque, areolate, cephalothecoid, 2–3 layered; outer ascomatal wall cells prismatic; intern layers composed of flattened cells. Paraphyses filiform-venticrose. Asci unitunicate, eight-spored, narrowly clavate or cylindrical, non-amyloid, apical ring thickened, with or without subapical globulus. Ascospores biseriate or multiseriate, at first one-celled, hyaline, cylindrical, vermiform, slightly sigmoid or bent in one end, later swelling above, becoming transversely uniseptate; upper cell brown, ellipsoidal to pyriform, more or less equilateral, smooth-walled, truncate at base, with a subapical germ pore, sometimes with one or more transverse septum; lower cell hyaline to pale brown, cylindrical, geniculate below, collapsing with age, without septa or 3–5-septate; gelatinous cauda absent or present, lash-like, attached at one or both ends of the ascospore. Asexual morph absent or present. Conidiophores reduced to conidiogenous cells. Conidiogenous cells phialidic, hyaline, arising directly from ascospores, normally from the lower cell, rarely branched. Conidia hyaline, globose, smooth-walled.
Notes: The main distinctive feature of Areotheca is its carbonaceous, areolate, cephalothecoid ascomatal wall. In this regard, other taxa in the Sordariales produce this type of ascomatal wall: Cladorrhinum coprophilum (syn. C. coprophila) shows indistinct areoles, whereas they are more obvious in C. caerulea, C. septentrionalis, C. silvatica, and T. striata (syn. C. striata). However, these particular structures are fully developed and carbonaceous in Ar. areolata. Another important morphological feature is the septation of the upper cell, present only in the genus Naviculispora and some species (Bellojisia rhynchostoma, C. sulphurella, Corylomyces selenospora, Podospora dydima, and Zopfiella tabulata) included in the Lasiosphaeriaceae s. str., and Zopfiella pleuropora (of clade VI). Finally, Areotheca is distinguished from other genera in the family by its ascospores with a basal cell up to twice as long as the upper cell. The two species of Areotheca are mainly differentiated by the size of their ascospores (the upper cell measures 20–27 × 8–11 µm in Ar. ambigua and 15–18 × 7–10 µm in Ar. areolata; the lower cell measures 35–55 × 6–7 µm in Ar. ambigua and 25–32 × 4–5 µm in Ar. aerolata), and by the presence of several septa in the lower ascospore cell of Ar. ambigua, absent in Ar. areolata.
Areotheca areolata (N. Lundq.) Y. Marín, A.N. Mill. & Stchigel, comb. nov. MycoBank MB 835498. (
Figure 3A–D).
Basionym: Cercophora areolata N. Lundq., Symb. Bot. Upsal. 20: 104. 1972.
Areotheca ambigua (Sacc.) Y. Marín & Stchigel, comb. nov. MycoBank MB835681.
Basionym: Lasiosphaeria hispida subsp. ambigua Sacc., Michelia 1: 46. 1877.
Synonym: Lasiosphaeria ambigua (Sacc.) Sacc., Sylloge Fungorum 2: 197. 1883.
Bombardia ambigua (Sacc.) G. Winter, Rabenhorst’s Kryptogamen-Flora, Pilze - Ascomyceten, Edn 2 1: 236. 1885.
Lasiosordaria ambigua (Sacc.) Chenant., Bull. Soc. Mycol. France 35: 79. 1919.
Cercophora ambigua (Sacc.) R. Hilber, in Hilber & Hilber, Z. Mykol. 45: 212. 1979.
As the strain CBS 13729 was placed in a clade far from Lasiosphaeriaceae s. str., and because it differs from all other members of Lasiosphaeriaceae s. lat. by the production of navicular ascospores whose upper cell is dark brown and septate and the lower cell is pale brown and non-collapsing with age, it is proposed as belonging to the new genus Naviculispora as follows:
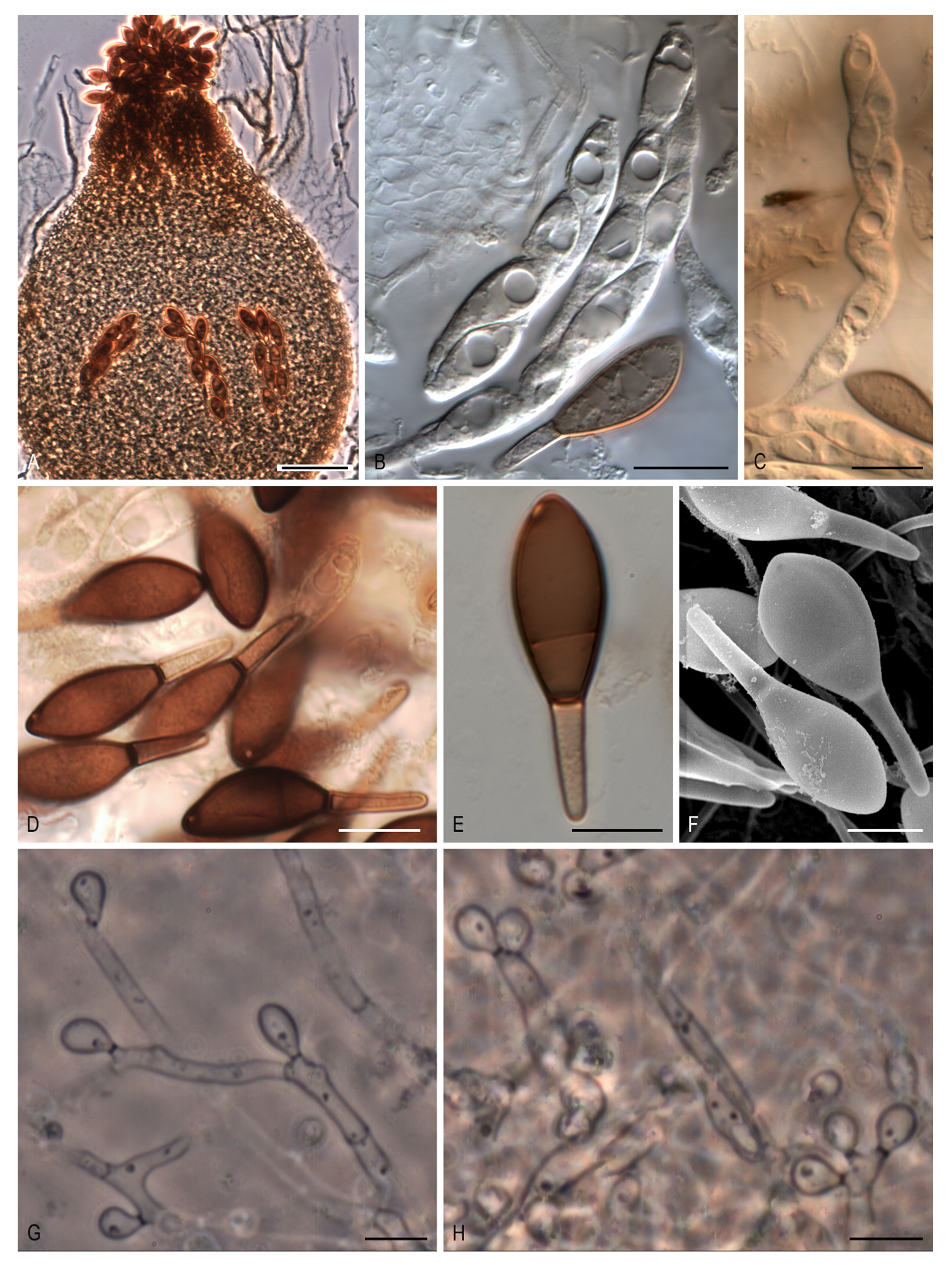
Naviculispora Stchigel, Y. Marín, Cano & Guarro, gen. nov. MycoBank MB 812135. (
Figure 4A–H).
Etymology: From Latin navicularibus-, navicular, and -sporarum, spore, referring to the shape of the ascospores.
Type species: Naviculispora terrestris Stchigel, Cano, Y. Marín & Guarro.
Ascomata ostiolate, brown, pyriform, covered with septate hyphae-like hairs; neck short, dark brown, papillate; ascomatal wall membranaceous, textura angularis. Asci unitunicate, eight-spored, cylindrical, short stipitate, with a small apical ring, evanescent. Ascospores at first one-celled, hyaline, clavate, becoming transversely septate and two-celled; upper cell dark brown, navicular, septate, with a subapical to lateral germ pore; lower cell pale brown, cylindrical-conical, thick-walled, not collapsing with age; gelatinous cauda absent. Conidia holoblastic, hyaline to subhyaline, ellipsoidal to obovoid or clavate, sessile or less commonly on sympodially proliferating conidiophores.
Notes:
Naviculispora spp. can be easily distinguished from the two phylogenetically related species of
Arnium (
Arnium caballinum and
A. japonense) by the production of two-celled ascospores (one-celled in
Arnium spp.).
Areotheca, as well as
Naviculispora, produces two-celled ascospores with a septum in the upper cell. However, these taxa differ in the nature of the ascomatal wall, being membranaceous in
Naviculispora and cephalothecoid in
Areotheca. As in
Naviculispora spp., the upper cell of the ascospores in
Bellojisia rhynchostoma,
Cercophora sulphurella,
Corylomyces selenospora,
Podospora dydima, and
Zopfiella tabulata is septate; however, the lower cell of the ascospores is pale brown to brown, thick-walled, and non-collapsing with age in
Naviculispora spp., whereas, in the other taxa, it is hyaline (or nearly so) and collapses very soon, except for
C. sulphurella.
Zopfiella attenuata and
Z. pleuropora, both located in a non-supported main clade (clade VI,
Figure 1), also produce ascospores with a septate upper cell, but differ from the Naviculisporaceae by the production of non-ostiolate ascomata.
Naviculispora terrestris Stchigel, Y. Marín, Cano & Guarro, sp. nov. MycoBank MB 812136. (
Figure 4A–H).
Etymology: Referring to the substrate from which the fungus has been isolated.
Type material: ARGENTINA: Tucumán province, Tafí del Valle, -26.8667, -65.6833, from soil, 17-V-2000, A. M. Stchigel, J. Cano, J. Guarro. (holotype CBS H-2159; ex-type cultures CBS 137295 = FMR 10060).
Mycelium composed of subhyaline to brown, septate, smooth-walled, branched hyphae, 1.5–4 µm wide. Ascomata ostiolate, superficial or immersed, scattered to aggregated, pale brown to brown, pyriform, 210–410 × 160–380 µm, covered with brown, septate hyphae-like hairs, occasionally with 2 necks; necks short, brown to dark brown, cylindrical to conical, papillate, 35–70 µm long, 62.5–105 µm wide; ascomatal wall membranaceous, 10–20 µm thick, composed of up to 10 layers of flattened cells of 2–10 µm in diam, textura angularis. Asci unitunicate, eight-spored, cylindrical, evanescent, 100–150 × 15–20 µm, short stipitate, with small apical ring. Ascospores uniseriate to biseriate, at first one-celled, hyaline, clavate, becoming transversely septate and two-celled; upper cell dark brown, navicular, 20–29 × 9–13.5 µm, truncate at the base, septate, with a subapical to apical germ pore of 0.5–1 µm; lower cell pale brown, cylindrical-conical, thick-walled, 10–14(–17) × (2–)3–4 µm, not collapsing; gelatinous cauda absent. Asexual morph present. Conidia holoblastic, 3–8 × 2–3 µm, hyaline to subhyaline, ellipsoidal to ovoid or clavate, with a small truncate base, sessile, produced laterally and terminally on the hyphae, or less commonly on sympodially proliferating conidiophores.
Culture characteristics: Colonies on PCA attaining a diam. of 40–47 mm in 14 d at 25 °C, velvety, slightly lobulate, margins fringed, radially zonate, greyish-brown (M 5D3 to 5E3) and brown to dark brown (6E3 to 6F3) at center; reverse greyish-yellow to olive brown (4B4 to 4D4). Colonies on OA attaining a diam. of 35–38 mm in 14 d at 25 °C, velvety, margins regular to slightly arachnoid, grey to greyish-brown (M 8F1 to 8F3); reverse grey to greyish brown (M 8F1 to 8F3). Ascomata produced after at least two months. Maximum and minimum temperatures of growth, 5 and 30 °C, respectively. Optimal temperature 25 °C.
Reference strains of
P. cochleariformis,
P. decipiens, P. myriaspora, and
P. pleiospora, classified by Lundqvist [
9] into the section
Rhypophila of the genus
Podospora, formed a well-supported clade (100% bs/ 1 pp) in the Naviculisporaceae. All four species are characterized by the production of an ascomatal neck with elongate tuberculated projections at the base, asci usually with more than eight ascospores, and ascospores with a lower cell usually longer than the upper cell. Based on their morphologically distinctive features and our phylogenetic results, we erect the genus
Rhypophila as follows:
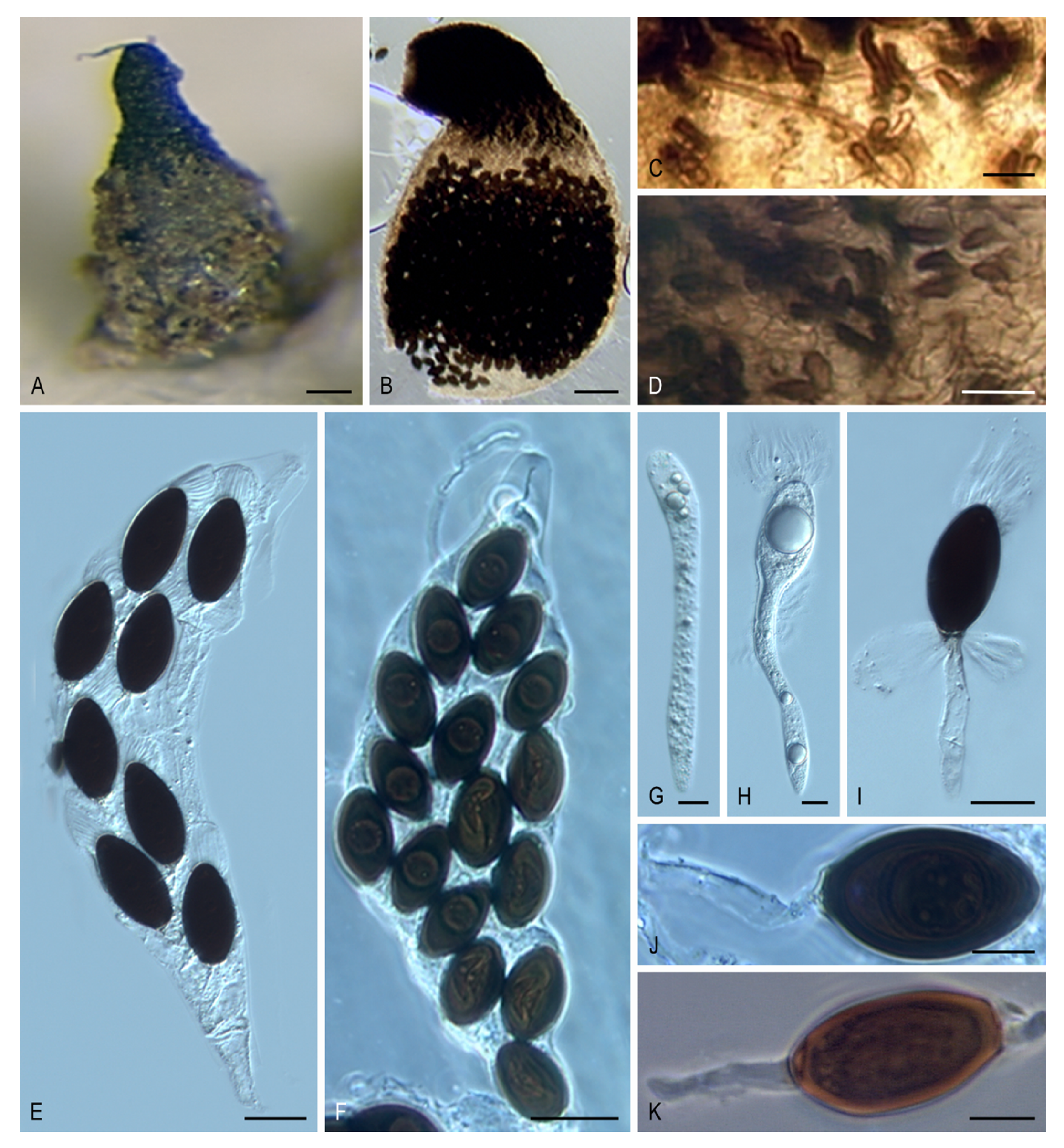
Rhypophila Y. Marín, A.N. Mill. & Guarro, gen. nov. MycoBank MB 812130. (
Figure 5A–K).
Etymology: Referring to the name of the section Rhypophila of the genus Podospora, proposed by Lundqvist (1972).
Type species: Rhypophila myriospora (P. Crouan & H. Crouan) Y. Marín, A.N. Mill. & Guarro.
Ascomata ostiolate, semi-immersed or superficial, scattered or aggregated, pyriform, glabrous or covered with flexous or stiff hairs; neck long, conical or cylindrical, with blackish, obtuse, straight or curved, elongate tuberculated projections at the base; ascomatal wall membranaceous, semi-translucent, yellowish to light brown. Paraphyses absent or present. Asci unitunicate, 8- to 128-spored, clavate, long or short stipitate, apical ring absent or indistinct. Ascospores biseriate or irregularly arranged, at first one-celled, hyaline, cylindrical or spatuliform, clavate, later swelling above, becoming transversely uniseptate and two-celled; upper cell dark brown, ellipsoidal to ellipsoidal-fusiform, with an apical or subapical germ pore; lower cell hyaline, cylindrical to cylindrical-obclavate, same length or longer than upper cell, frequently collapsing; secondary appendages small, at the base of the lower cell; apical gelatinous cauda absent or present, fibrillate or lamellate. Asexual morph absent.
Notes:
Rhypophila includes species formerly classified in a section with the same name in the genus
Podospora [
9]. Chang et al. [
12] also noted that
Podospora species of the section
Rhypophila grouped together in the phylogenies based on ITS and a fragment of the glyceraldehyde-3-phosphate dehydrogenase (
gpd) gene. Although the phylogenetic distance between
R. cochlearifomis/R. myriaspora and
R. decipiens/R. pleiospora suggests that these could represent two different genera, we decided to include all these species in
Rhypophila based on the morphological resemblances. Further studies including more taxa are needed to clarify it. The genus
Rhypophila is characterized by the production of ostiolate ascomata with a neck bearing elongated tuberculated projections at the base, asci normally containing more than eight ascospores, and ascospores with a lower cell as long as, or longer than, the upper cell. Ornamentation in the upper part of the ascomata is also present in
Pseudoechria, Pseudoschizothecium, and
Schizothecium, but these three genera, classified in the new family Schizotheciaceae (clade VIII,
Figure 2), differ in the nature of the ornamentation (see Notes on
Pseudoechria).
Rhypophila cochleariformis (Cailleux) Y. Marín, A.N. Mill., Guarro, comb. nov. MycoBank MB 812131.
Basionym: Podospora cochleariformis Cailleux, Cah. Maboké 7: 100. 1969.
Rhypophila decipiens (G. Winter ex Fuckel) Y. Marín, A.N. Mill., Guarro, comb. nov. MycoBank MB 812132. (
Figure 5E,G–I)
Basionym: Sordaria decipiens G. Winter, Abh. Naturf. Ges. Halle 13: 28. 1873.
Synonym: Podospora decipiens (G. Winter ex Fuckel) Niessl, Hedwigia 22: 156. 1883.
Rhypophila myriospora (P. Crouan & H. Crouan) Y. Marín, A.N. Mill., Guarro, comb. nov. MycoBank MB 812133. (
Figure 5B,D,K).
Basionym: Sordaria myriospora P. Crouan & H. Crouan, Florule Finistère (Paris): 22. 1867.
Synonym: Podospora myriospora (P. Crouan & H. Crouan) Niessl, Hedwigia 22: 156. 1883.
Rhypophila pleiospora (G. Winter) Y. Marín, A.N. Mill. & Guarro, comb. nov. MycoBank MB 812134. (
Figure 5A,C,F,J).
Basionym: Sordaria pleiospora G. Winter, Abh. Naturf. Ges. Halle 13: 13. 1873.
Synonym: Podospora pleiospora (G. Winter) Niessl, Hedwigia 22: 156. 1883.
Key to species of
Rhypophila.
| 1. Asci 8-spored | R. decipiens |
| 1. Asci 16 to 128-spored | 2 |
| 2. Asci 128-spored; upper and lower cells each shorter than 20 µm; absence of cauda and gelatinous appendages | R. cochleariformis |
| 2. Asci 16-, 32- or 64-spored; upper and lower cells each longer than 20 µm; fibrillate upper cauda and gelatinous appendages in the lower cell | 3 |
| 3. Asci 16 or 32 spored; upper cell 23–34 × 14–19 µm | R. myriospora |
| 3. Asci 64 spored; upper cell 25–37 × 18–23 µm | R. pleiospora |
The Podosporaceae (100% bs/1 pp; clade IV,
Figure 1) formed three fully supported clades (100% bs/1 pp) representing the genera
Cladorrhinum,
Podospora, and
Triangularia. The reference strains of
A. tomentosum and
C. coprophila were nested in the
Cladorrhinum clade. Consequently, the description of
Cladorrhinum is emended, and new combinations for
A. tomentosum and
C. coprophila are proposed as follows:
Cladorrhinum Sacc. & Marchal, Bull. Soc. Roy. Bot. Belgique 24: 64. 1885. emend. Y. Marin & Stchigel. MycoBank MB7678.
Type species: Cladorrhinum foecundissimum Sacc. & Marchal.
Ascomata ostiolate or non-ostiolate, superficial to immersed, solitary or aggregated, globose to subglobose, ovoid, conical, obpyriform or broadly obpyriform, covered with hypha-like ascomatal hairs or by aerial mycelium, or a dense tomentum of thick-walled hairs; ascomatal wall membranaceous, semi-translucent to opaque, textura angularis, textura intricata or textura epidermoidea in surface view, or sometimes composed of patches or indistinct areoles; neck cylindrical, black, opaque, carbonaceous, often covered by rigid, septate hairs. Paraphyses absent or present, filiform-vetricose. Asci unitunicate, eight-spored, cylindrical, narrowly clavate, pyriform, obovoid or fusiform, short- or long-stipitate, without or with a thin or thick apical ring, evanescent. Ascospores uniseriate into the asci, one- or two-celled. One-celled ascospores ochraceous or olivaceous brown to dark brown, ellipsoidal to broadly fusiform, ovoid or obovoid, smooth-walled, with a germ pore at each end or with an apical germ pore, a diffuse sheath surrounding the whole spore sometimes present; gelatinous cauda absent or present, attached to each end. Two-celled ascospores biseriate or triseriate, at first hyaline, one-celled, vermiform, slightly sigmoid or geniculate only below, smooth-walled, then swelling above, becoming transversely septate and two-celled; upper cell brown, ellipsoidal, truncate at the base, conical above with an apical germ pore; lower cell cylindrical, collapsing; gelatinous cauda attached to each end. Asexual morph absent or present. Conidiophores micronematous, reduced to conidiogenous cells. Conidiogenous cells mostly intercalary, occasionally terminal, originating as a lateral or terminal peg-like structure with a flaring collarette. Conidia enteroblastic, one-celled, hyaline, broad obovoid, ellipsoidal or subglobose, usually with a truncated base and a rounded apex, smooth-walled.
Notes:
Cladorrhinum previously included species that only produced an asexual morph characterized by the production of one-celled conidia in slimy masses from intercalary enteroblastic conidiogenous cells with lateral openings (integrated phialides; adelophialides). Fertile hyphae can develop more or less branched conidiophores, and terminal and lateral phialides and microsclerotia can sometimes also be observed in culture [
44,
45]. Recently, Wang et al. [
6] maintained in the genus only the type species,
Cl. foecundissimum, based on sequence data. They also observed that two species previously identified as
Thielavia hyalocarpa and
Th. intermedia, clustered together with
Cl. foecundissimum. Therefore, the description of
Cladorrhinum was emended to incorporate the presence of a sexual morph characterized by non-ostiolate ascomata and one-celled ascospores. In our phylogenetic study,
C. coprophila, which produces ostiolate ascomata and two-celled ascospores, as well as
A. tomentosum, which forms ostiolate ascomata and one-celled ascospores, also grouped in the
Cladorrhinum clade. Therefore, the new combinations are proposed here and the description of the genus is newly emended to incorporate these new morphological characteristics. In addition, a reference strain of
A. olerum, CBS 120012, occurred in a well-supported clade (100% bs/0.98 pp) together with the type strain of
Cl. foecundissimum, suggesting that the former species is the sexual stage of the asexual
Cl. foecundissimum; therefore, their synonymy is proposed here.
Cladorrhinum, Podospora, and Triangularia present the same asexual morph. However, these genera differ among the sexual morphs. The ascospores are mostly one-celled in
Cladorrhinum, whereas they are mostly two-celled in
Podospora and
Triangularia. Ostiolate and non-ostiolate ascomata can be found in all three genera, suggesting that the ascomatal morphology in Podosporaceae is not phylogenetically informative. Cai et al. [
46] had already observed this high variability and reached the same conclusion. Sequence data are necessary to properly delineate these genera, which display a high variability in the ascomata and ascospore morphology, with many overlapping features among them.
Cladorrhinum coprophilum and
Cl. tomentosum produce ascomata with a wall composed of patches or indistinct areoles, a feature also observed in
T. striata. However, this latter species can be distinguished from the former two by its tuberculate projections on the neck giving a striate appearance.
Cladorrhinum coprophilum (Fr.) Y. Marin, A.N. Mill. & Stchigel, comb. nov. MycoBank MB 835499. (
Figure 6A–G).
Basionym: Sphaeria coprophila Fr., in Kunze & Schmidt, Mykol. Hefte (Leipzig) 2: 38. 1823.
Synonym: Cercophora coprophila (Fr.) N. Lundq., Symb. Bot. Upsal. 20: 95. 1972.
Because the reference strain of Arnium olerum formed a well-supported terminal lineage with Cladorrhinum foecundissimum (100% bs/0.98 pp), the synonymy of A. olerum under C. foecundissimum is proposed here.
Cladorrhinum foecundissimum Sacc. & Marchal, Bull. Soc. R. Bot. Belg. 24: 64. 1885. MycoBank MB119723. (
Figure 7A–E).
Synonyms: Sphaeria olerum Fr., Elench. Fung. (Greifswald) 2: 98. 1828.
Arnium olerum (Fr.) N. Lundq. & J.C. Krug, Symb. Bot. Upsal. 20: 212. 1972.
For other synonyms see reference [
9].
Cladorrhinum tomentosum (Speg.) Y. Marin & Stchigel, comb. nov. MycoBank MB835500.
Basionym: Hypocopra tomentosa Speg., Anales Soc. Ci. Argent.10: 136. 1880.
Synonym: Arnium tomentosum (Speg.) N. Lundq. & J.C. Krug, Symb. Bot. Upsal. 20: 216. 1972.
Sordaria tomentosa (Speg.) Sacc., Syll. Fung. (Abellini) 1: 236. 1882.
Podospora tomentosa (Speg.) Niessl, Hedwigia 22: 156. 1883.
Key to species of
Cladorrhinum.
| 1. Two-celled ascospores | Cl. coprophilum |
| 1. One-celled ascospores | 2 |
| 2. Non-ostiolate ascomata | 3 |
| 2. Ostiolate ascomata | 4 |
| 3. Ascospores (22.5–)24.5–31(–36) × (11.5–)13–15.5(–16) μm | Cl. hyalocarpum |
| 3. Ascospores 12–17 × 9–12 μm | Cl. foecundissimum |
| 4. Ascomatal wall not areolate, ascospores 40–52 × 25–32 µm | Cl. olerum |
| 4. Ascomatal wall areolate, ascospores 47–58 × 22–28(–30) µm | Cl. tomentosum |
The type strains of Apiosordaria sacchari and Ap. striatispora, and a reference strain of C. costaricensis, formed independent lineages within Podospora. Therefore, the new combinations are proposed here, and the description of Podospora is emended:
Podospora Ces., Hedwigia 1: 103. 1856. emend. Y. Marin & Stchigel. MycoBank MB4284.
Type species: Podospora fimicola (Corda) Ces.
Ascomata usually superficial, solitary, ostiolate and pyriform, or less frequently non-ostiolate and globose, covered by erect or flexuous, hyphae-like hairs; neck short, conical; ascomatal wall membranaceous to coriaceous, sometimes pseudo-bombardioid and containing a central layer composed of thick-walled, gelatinized cells. Paraphyses absent or present. Asci unitunicate, eight-spored, clavate or cylindrical, with an apical ring, usually persistent until ascospore maturity. Ascospores uniseriate or biseriate, two-celled; upper cell olivaceous brown to brown or dark brown, ellipsoidal to ovoid or obovoid, or five-angled in side view, smooth-walled, spinulose (spines up to 3 μm long), or with several longitudinal striations under light microscopy or by SEM exhibiting several longitudinal ribs that result in a coalescence of warts, with an apical germ pore; lower cell hyaline to pale brown, clavate or cylindrical, smooth-walled; apical and basal mucilaginous appendages mostly present. Conidiophores micronematous, reduced to conidiogenous cells. Conidiogenous cells usually intercalary, originating from a lateral peg-like structure with a flaring collarette. Conidia blastic, single-celled, hyaline, smooth-walled, globose or ellipsoidal to elongate, usually with a truncated base and a rounded apex.
Notes:
Podospora was one of the largest and morphologically most diverse genera in Lasiosphaeriaceae [
9]. Recently, this was reduced to the type species,
P. fimicola, and the asexual species,
P. bulbillosa, previously known as
Cl. bulbillosum [
6]. In the same study, it was designated as the type genus of the family Podosporaceae, which includes only two genera,
Cladorrhinum and
Triangularia. For morphological comparison, see Notes of
Cladorrhinum.
Podospora costaricensis (G.C. Carroll & Munk) Y. Marín, A.N. Mill. & Stchigel, comb. nov. MycoBank MB835501. (
Figure 8A–G).
Basionym: Bombardia costaricensis G.C. Carroll & Munk, Mycologia 56: 80. 1964.
Synonym: Cercophora costaricensis (G.C. Carroll & Munk) O. Hilber & R. Hilber, Z. Mykol. 45: 217. 1979.
Podospora sacchari (B.M. Robison) Y. Marín & Stchigel, comb. nov. MycoBank MB835502.
Basionym: Echinopodospora sacchari B.M. Robison, Trans. Brit. Mycol. Soc. 54: 320. 1970.
Synonym: Apiosordaria sacchari (B.M. Robison) J.C. Krug, Udagawa & Jeng, Mycotaxon 17: 546. 1983.
Podospora striatispora (Furuya & Udagawa) Y. Marín & Stchigel, comb. nov. MycoBank MB835503.
Basionym: Triangularia striatispora Furuya & Udagawa, J. Jap. Bot. 51: 406. 1976.
Synonym: Apiosordaria striatispora (Furuya & Udagawa) Guarro & Cano, Trans. Brit. Mycol. Soc. 91: 589. 1988.
Key to species of
Podospora.
| 1. Only asexual morph present | P. bulbillosa |
| 1. Sexual morph present | 2 |
| 2. Ascospores smooth-walled | 3 |
| 2. Ascospores ornamented | 4 |
| 3. Upper cell 44.5–55 × 27.5–46 μm, lower cell 25–37 × 4.5–6.5(–8) μm | P. fimicola |
| 3. Upper cell 18–22 × 8–9(–10) μm, lower cell 22–27 × 4–5 μm | P. costaricensis |
| 4. Upper cell obovoid, spinulose, 33.2–40.5 × 24.8–31.3 μm | P. sacchari |
| 4. Upper cell five-angled in side view, ornamented with longitudinal ribs, 10–12(–15) × 8–9 μm | P. striatispora |
Because the reference strains of A. arizonensis, C. striata, and Z. tetraspora grouped in the Triangularia clade, new combinations are proposed, and the description of Triangularia is emended:
Triangularia Boedijn, Ann. Mycol. 32: 302. 1934, emend. Y. Marin & Stchigel. MycoBank MB5534.
Synonym: Apiosordaria Arx & W. Gams, Nova Hedwigia 13: 201. 1967.
Type species: Triangularia bambusae (J.F.H. Beyma) Boedijn, Ann. Mycol. 32: 302. 1934.
Ascomata ostiolate or non-ostiolate, often superficial, sometimes semi-immersed to immersed, solitary or aggregated, globose to subglobose, ovoid, or obpyriform to ampulliform, glabrous or with hypha-like or seta-like hairs; neck papillate, sometimes with rigid, agglutinated, cylindrical, septate, thick hairs, sometimes striate with 5–8 striations; ascomatal wall membranaceous to coriaceous, translucent, semitranslucent, or opaque, textura angularis to textura globulosa, sometimes composed of areoles of textura angularis. Periphyses absent or present. Paraphyses usually present, hyaline, filiform or filiform-ventricose. Asci unitunicate, four-, eight-, or multi-spored, cylindrical to elongated clavate or fusiform, short or long stipitate, without or with an inconspicuous or thick apical ring, evanescent or persistent until ascospores mature. Ascospores one- or two-celled. One-celled ascospores uniseriate, at first hyaline, becoming yellowish, olivaceous to brown-black, ellipsoidal, smooth-walled, with an apical germ pore; gelatinous cauda at each end. Two-celled ascospores uniseriate, biseriate, triseriate, or irregularly arranged; upper cell olivaceous brown, brown, or dark brown, ellipsoidal, ovoid, fusiform, navicular, or conical and triangular in lateral, smooth-walled, or spinulose to warty, with an apical or subapical pore, rarely 1-septate; lower cell short or long, hyaline to pale brown, occasionally brown, elongate fusiform, cylindrical, conical, triangular to hemispherical, rarely early evanescent, frequently collapsing, rarely up to 3-sepate; gelatinous appendages absent or present. Asexual morph absent or present. Conidiophores micronematous, reduced to conidiogenous cells. Conidiogenous cells intercalary or occasionally terminal, sometimes phialidic, originating lateral or terminal peg-like structure with a flaring collarette. Conidia blastic, single-celled, hyaline, globose to subglobose or pyriform, smooth-walled, usually with a truncated base and a rounded apex.
Notes:
Triangularia included species traditionally characterized by ascospores with a conical, triangular in lateral view upper cell, and triangular or hemispherical lower cell [
43,
47]. Recently, this genus was restricted to the type species of the genus,
T. bambusae, together with several species of
Apiosordaria and
Podospora that occurred in the same monophyletic clade [
6]. All the species at that time included in
Triangularia presented two-celled ascospores, although the lower cell of
T. allahabadensis is soon evanescent. In our phylogenetic study,
Ar. arizonense was nested in the
Triangularia clade, and, therefore, a new combination is proposed here and, consequently, the description of the genus is emended to incorporate the possibility of finding one-celled ascospores. Moreover,
C. striata and
Z. tetraspora, both characterized by two-celled ascospores, were also located in the same clade with
Triangularia spp., and new combinations are proposed here for these two taxa. Miller and Huhndorf [
48] observed that
T. striata produces a cladorrhinum-like asexual morph, which is one of the main characteristics that defines the recently described family Podosporaceae, where
Triangularia belongs. See Notes of
Cladorrhinum for morphological comparison with related genera.
Triangularia arizonensis (Griffiths) Y. Marín, A.N. Mill. & Stchigel, comb. nov. MycoBank MB835504. (
Figure 9A–F).
Basionym: Pleurage arizonensis Griffiths, Mem. Torrey Bot. Club 11: 57. 1901.
Synonyms: Sordaria arizonensis (Griffiths) Sacc. & D. Sacc., Syll. Fung. (Abellini) 17: 601. 1905.
Podospora arizonensis (Griffiths) Cain, Canad. J. Bot. 40: 459. 1962.
Arnium arizonense (Griffiths) N. Lundq. & J.C. Krug, Symb. Bot. Upsal. 20: 232. 1972.
Triangularia striata (Ellis & Everh.) Y. Marín, A.N. Mill. & Stchigel, comb. nov. MycoBank MB835505. (
Figure 10A–H).
Basionym: Sordaria striata Ellis & Everh., J. Mycol. 4: 79 [‘67’]. 1888.
Synonyms: Lasiosordaria striata (Ellis & Everh.) Chenant., Bull. Soc. Mycol. France 35: 78. 1919.
Podospora striata (Ellis & Everh.) Ellis & Everh., N. Amer. Pyren. (Newfield): 131. 1892.
Cercophora striata (Ellis & Everh.) N. Lundq., Symb. Bot. Upsal. 20: 105. 1972.
Triangularia tetraspora (J.N. Rai, Mukerji & J.P. Tewari) Y. Marín & Stchigel, comb. nov. MycoBank MB835506.
Basionym: Tripterospora tetraspora J.N. Rai, Mukerji & J.P. Tewari, Canad. J. Bot. 41: 327. 1963.
Synonyms: Zopfiella tetraspora (J.N. Rai, Mukerji & J.P. Tewari) S. Ahmad, Monogr. Biol. Soc. Pakistan 7: 78. 1978.
Podospora buffonii Cailleux, Cah. Maboké 7: 12. 1969.
Key to species of
Triangularia.
| 1. Only asexual morph produced | T. phialophoroides |
| 1. Sexual morph produced | 2 |
| 2. Ascospores one-celled | T. arizonensis |
| 2. Ascospores two-celled | 3 |
| 3. Asci 128-spored | T. setosa |
| 3. Asci 4–8-spored | 4 |
| 4. Ascospores spinulose to warty | T. verruculosa |
| 4. Ascospores smooth-walled under light microscope | 5 |
| 5. Lower cell early evanescent | T. allahabadensis |
| 5. Lower cell not evanescent, frequently collapsing | 6 |
| 6. Lower cell short, conical, or triangular to hemispherical | 7 |
| 6. Lower cell long, elongate fusiform to cylindrical | 8 |
| 7. Lower cell conical, ascospores with mucilaginous small appendages at both ends | T. backusii |
| 7. Lower cell triangular to hemispherical, ascospores without mucilaginous appendages | T. bambusae |
| 8. Ascomata non-ostiolate | 9 |
| 8. Ascomata ostiolate | 10 |
| 9. Upper cell (11–)12–14.5(–15) × (7.5–)8–9(–9.5) µm | T. longicaudata |
| 9. Upper cell 23.5–35.5 × 17.5–23.5 µm | T. tetraspora |
| 10. Striate ascomata composed of areoles, upper cell up to 23 µm | T. striata |
| 10. Above characters of the ascomata not present, upper cell up to 38 µm | T. anserina/comata/pauciseta complex
|
Clade VIII (80% bs/1 pp;
Figure 2) includes species of
Schizothecium,
Echria,
Immersiella,
Jugulospora,
Rinaldiella,
Strattonia, and
Zygopleurage, plus several species of
Arnium,
Apiosordaria,
Cercophora,
Triangularia, and
Zopfiella. This clade represents a new family, the Schizotheciaceae, which is proposed below.
Schizotheciaceae Y. Marin & Stchigel, fam. nov. MycoBank MB835507.
Etymology: Based on the genus Schizothecium, the oldest representative genus in the family.
Type genus: Schizothecium Corda, Icon. Fung. (Prague) 2: 29. 1838.
Ascomata ostiolate, immersed, semi-immersed, or superficial, scattered or aggregated, light to dark brown or black, globose to subglobose, pyriform, pyriform-conical, ovoid or obpyriform, glabrous or covered with flexous hairs, sometimes upper part of the ascomata adorned with groups of swollen agglutinated hairs, or with prominent protruding ascomatal cells; neck short or long, dark brown to black, opaque, conical, cylindrical, or elongate, papillate, sometimes with short, thick-walled, swollen cells, or swollen agglutinated hairs, or long, sometimes agglutinated, rigid hairs not forming fascicles, or with tufts of brown to black, rigid, agglutinated, septate, long, thick hairs; ascomatal wall pseudoparenchymatous, membranaceous, occasionally semi-coriaceous, semi-transparent, occasionally dark brown to black and opaque, yellowish to light brown, textura angularis, textura globulosa, or textura epidermoidea. Paraphyses and periphyses absent or present. Asci unitunicate, 4- to 2048-spored, clavate, cylindrical, broadly ellipsoid, or broadly fusiform, apically rounded, long or short stipitate, apical ring absent or present, dehiscing below the apex. Ascospores uniseriate, biseriate, or irregularly arranged, one- or two-celled. One-celled ascospores uniseriate or biseriate, hyaline, turning pale brown, golden, ochraceous, dark brown, or blackish brown at mature, cylindrical, fusiform or ellipsoidal, sigmoid or geniculate, one end may become swollen and turn brown, smooth-walled, without or with a germ pore at one or both ends, sometimes with a gelatinous appendage at each end, sometimes with a striated gelatinous sheath surrounding whole ascospore. Two-celled ascospores at first one-celled, hyaline, clavate, cylindrical or spatuliform, mostly becoming transversely uniseptate, occasionally developing two terminal ellipsoid black cells that remain connected with the long, often undulating, hyaline middle part of the spore, with a germ-pore at the free end and secondary appendages gelatinous arranged in rings around the top and base of the colored cells; upper cell olivaceous to brown to dark brown, globose, navicular, ellipsoidal, ellipsoidal-fusiform, fusiform, fusiform-obovoid, obovoid, or polygonal with five angles in side view, smooth-walled, finely granulated, pitted, ornamented with spines, or warted, sometimes with warts arranged forming ridges or large spots, with an apical or subapical germ pore; lower cell hyaline to pale brown, conical, cylindrical, cylindrical-obclavate, obclavate or obconical, collapsing or long persistent, smooth-walled to slightly warted; apical gelatinous appendages absent or present, attached at each end, lash-like; gelatinous sheaths rarely present, striated, swelling in water. Asexual morph absent or present. Conidiophores reduced to conidiogenous cells. Conidiogenous cells sometimes present, phialidic. Conidia hyaline to pale brown, ovoid to clavate or elongate, almost smooth-walled, usually produced laterally or terminally on undifferentiated hyphae, solitary.
Notes: This family is introduced to accommodate lasiosphaeriaceous taxa located in a well-supported clade (80% bs/1 pp; clade VIII,
Figure 2), distinct from Lasiosphaeriaceae s. str. (clade V,
Figure 1). It includes the genera
Echria,
Immersiella,
Jugulospora,
Pseudoechria,
Pseudoschizothecium,
Rinaldiella,
Rhypophila,
Schizothecium, and
Zygopleurage, but also several species of
Apiosordaria and
Zopfiella. However, the type species of these latter genera are located in the Podosporaceae (clade IV) and in the Lasiopshaeriaceae s. str. (clade V), respectively. Consequently, those species that fall into clade VIII must be reviewed before they are moved to other (new) genera. Two species of
Arnium, i.e.,
A. cirriferum and
A. hirtum, are also included in this family.
Arnium is a polyphyletic genus scattered throughout the order Sordariales, and its type species (
A. lanuginosum) has not been sequenced. Finally, a strain identified as
Cercophora mirabilis (which is the type species of the genus) was also located in the Schizotheciaceae, but its morphological study was not possible to confirm the identification, because the colonies remained sterile on different culture media. Due to this inconvenience, further studies are needed for the correct placement and delimitation of the genus
Cercophora. The Schizotheciaceae are characterized by the production of mostly ornate, ostiolate ascomata with different kinds of structures:
Schizothecium presents short swollen agglutinated hairs or prominent protruding ascomatal wall cells;
Echria and
Pseudoechria produce tufts of long rigid agglutinated hairs, while
Pseudoschizothecium produces warts composed of small polygonal cells. The ascomatal wall is mostly pseudoparenchymatous, membranaceous, and semi-transparent. Ascospores display a large amount of variability including taxa with one- or two-celled ascospores, as well as ascospores with two independent cells that remain connected by a long, often undulating, hyaline middle part.
In the Schizotheciaceae, a well-supported clade (100% bs/1 pp;
Figure 2) was composed of a reference strain of
Jugulospora rotula, ATCC 38359; a reference strain of
Strattonia carbonaria, ATCC 34567; the type strains of
Apiosordaria antarctica, Apiosordaria globosa, Apiosordaria hispanica, and
Apiosordaria vestita, CBS 381338, CBS 110113, CBS 110112, and IMI 381338, respectively; and an isolate morphologically identified by us as
Rhexosporium terrestre, FMR 12428. All these taxa produce similar ascomata (with a translucent ascomatal wall and dark brown papillate neck), and clavate and early septate ascospores with the upper cell warted or finely granulate. Therefore, all these taxa are transferred to the genus
Jugulospora, which is also emended.
Jugulospora N. Lundq., Symb. Bot. Upsal. 20(no. 1): 256. 1972, emend. Y. Marin & Stchigel. MycoBank MB2538. (
Figure 11A–N).
Synonym: Rhexosporium Udagawa & Furuya, Trans. Mycol. Soc. Japan 18: 302. 1977.
Type species: Jugulospora rotula (Cooke) N. Lundq.
Ascomata ostiolate, covered with pale brown, flexuous, septate, thick-walled hairs; neck dark brown to black, composed of papillate cells disposed around the ostiole; ascomatal wall membranaceous, semi-transparent, 3–9-layered, outer cells isodiametric. Paraphyses filiform. Asci unitunicate, eight-spored, cylindrical, with a thin apical ring. Ascospores at first one-celled, hyaline, clavate, early septate; upper cell brown, navicular, globose, or obovoid, warted, finely granulated, or pitted, sometimes with warts arranged forming ridges or large spots, with an apical germ pore; lower cell hyaline, conical to cylindrical, smooth-walled to slightly warted, collapsing; gelatinous sheath rarely present, hyaline, thin. Asexual morph absent or present. Conidia hyaline to pale brown, almost smooth-walled, ovate to elongate, produced laterally or terminally on undifferentiated hyphae, solitary.
Notes: The genus
Jugulospora was introduced by Lundqvist [
9] to accommodate
Sphaeria rotula based on its verrucose ascospores with anastomosing warts arranged in large spots. The molecular data demonstrated that
A. globosa,
A. hispanica, and
R. terrestre, also with warted ascospores, is synonymous with
J. rotula.
Rhexosporium was proposed as a new genus by Udagawa and Furuya [
49] due to the presence of the longitudinal ridges in the upper cell of the ascospores, even though these authors cited a similar ascospore ornamentation in the genera
Apiosordaria and
Jugulospora. We noticed that such ascospore ridges seen under SEM (from the pictures taken by those authors) were really in a linear arrangement of warts and, consequently, not differing with respect to the ornamentation of those of
J. rotula. Based on the early ascospore septation, the absence of staining of the ascal apical ring by cotton blue, and the presence of paraphyses, Lundqvist [
9] highlighted the close similarity between
Jugulospora and
Strattonia carbonaria. Our study confirmed the phylogenetic relationship among them, already demonstrated in previous phylogenetic studies [
11,
26]. Therefore, we transferred
Strattonia carbonaria to
Jugulospora as
J. carbonaria. Although
Strattonia is an older name than Jugulospora, the type species of that genus,
S. tetraspora, has an ascomatal neck with rigid, cylindrical, septate, long hairs and ascospores with a gelatinous sheath, features that do not match with those of Jugulospora. Hence, the genus
Strattonia should be reduced to those species that share the morphology of the type species. Moreover, the type strains of
A. antarctica and
A. vestita clustered in the
Jugulospora clade but formed independent lineages. Therefore, both species are transferred to
Jugulospora.
Jugulospora antarctica (Stchigel & Guarro) Y. Marín & Stchigel, comb. nov. MycoBank MB835663. (
Figure 11N).
Basionym: Apiosordaria antarctica Stchigel & Guarro, in Stchigel, Guarro & Mac Cormack, Mycologia 95: 1219. 2003.
Jugulospora carbonaria (W. Phillips & Plowr.) Y. Marín, Stchigel, Guarro & A.N. Mill., comb. nov. MycoBank MB812137. (
Figure 11C,D,M).
Basionym: Sphaeria carbonaria W. Phillips & Plowr., Grevillea 2: 188. 1874.
Synonyms: Podospora carbonaria (W. Phillips & Plowr.) Niessl, Hedwigia 22: 156. 1883.
Psilosphaeria carbonaria (W. Phillips & Plowr.) Cooke & Plowr., Grevillea 7: 85. 1879.
Sordaria carbonaria (W. Phillips & Plowr.) Sacc., Syll. Fung. (Abellini) 1: 233. 1882.
Strattonia carbonaria (W. Phillips & Plowr.) N. Lundq., Symb. Bot. Upsal. 20: 269 (1972).
Zopfiella carbonaria (W. Phillips & Plowr.) Arx, Proc. Kon, Ned. Akad. Wetensch. C. 76: 291. 1973.
Jugulospora rotula (Cooke) N. Lundq., emend. Y. Marin, Dania García, Cano & Stchigel. MycoBank MB315972. (
Figure 11A,B,E–L).
Basionym: Sphaeria rotula Cooke, Handb. British Fungi 2: no. 2598. 1871.
Synonyms: Rhexosporium terrestre Udagawa & Furuya, Trans. Mycol. Soc. Japan 18: 303. 1977.
Apiosordaria globosa Dania García, Stchigel & Guarro, Mycologia 95: 137. 2003.
Apiosordaria hispanica Dania García, Stchigel & Guarro, Mycologia 95: 134. 2003.
Ascomata ostiolate, superficial or immersed, scattered to aggregated, pale brown to brown, pyriform, 350–770 × 200–540 µm, covered with pale brown, wide near the base, septate hyphae-like hairs of 1–5 µm diam; neck brown to dark brown, cylindrical to conical, papillate, 78–280 µm long, 90–250 µm wide; ascomatal wall membranaceous, semi-transparent, brownish-orange to brown, 3–9-layered, 15–45 µm thick; outer layers with textura angularis to textura intricata; inner layers with textura epidermoidea. Paraphyses and periphyses filiform, up to 2 µm in diam. Asci unitunicate, eight-spored, cylindrical, 146–250 × 14–28 µm, stipitate, with a thin apical ring, evanescent. Ascospores at first one-celled, hyaline, clavate, becoming transversely septate and two celled; upper cell dark brown, obovoid to globose, truncate at the base, ornamented with warts arranged uniformly or forming longitudinal ridges or large spots, 18–29 × 12–27 µm, with an apical to lateral germ pore of 0.5–3 µm; lower cell hyaline, conical, smooth-walled to slightly warted, 1–6 µm, collapsing; gelatinous cauda absent. Asexual morph absent or present. Conidia hyaline to pale-colored, almost smooth-walled, ovate to elongate, 2–6 × 1.5–2.5 µm, produced laterally or terminally on undifferentiated hyphae, solitary.
Specimens examined: USA, NORTH CAROLINA, Great Smoky Mountains National Park, from soil, 8-VIII-2008, M. Calduch, A.N. Miller & A.M. Stchigel, culture FMR 12690; USA, TENNESSEE, Great Smoky Mountains National Park, from soil, 9-VIII-2008, M. Calduch, A.N. Miller & A.M. Stchigel, culture FMR 12781; SPAIN, TARRAGONA, Els Gorgs de la Febró, from soil, 25-IX-1996, A.M. Stchigel & M. Calduch, culture CBS 110112, ex-type strain of Apiosordaria hispanica; SPAIN, TARRAGONA, Els Gorgs de la Febró, from soil, 25-IX-1996, A.M. Stchigel & M. Calduch, culture CBS 110113, ex-type strain of Apiosordaria globosa; ANTARCTICA, King George Island, Jubany Argentinian base, from soil, 11-XI-1996, W. Mac Cormack, culture IMI 381338, ex-type strain of Apiosordaria antarctica; JAPAN, from burned soil, Y. Horie, culture ATCC 34567.
Jugulospora vestita (Udagawa & Y. Horie) Y. Marín & Stchigel, comb. nov. MycoBank MB835664.
Basionym: Apiosordaria vestita Udagawa & Y. Horie, Reports on the Cryptogamic Study in Nepal (Tokyo): 97. 1982.
Key to species of
Jugulospora.
| 1. Ascospores pitted, with gelatinous sheath | J. vestita |
| 1. Ascospores warty or finely granulated, without gelatinous sheath | 2 |
| 2. Ascospores warty | J. rotula |
| 2. Ascospores finely granulated | 3 |
| 3. Ascomata with short neck | J. carbonaria |
| 3. Ascomata with long neck, up to 400 μm | J. antarctica |
The type strain of
T. tanzaniensis and a reference strain of
Z. karachiensis, which are characterized by the formation of ostiolate ascomata with membranaceous to semi-coriaceous ascomatal walls, and ascospores with a smooth-walled upper cell, were included in a well-supported clade (100% bs/1 pp;
Figure 2). Consequently, we propose the erection of the new genus
Lundqvistomyces as follows:
Lundqvistomyces Y. Marín & Stchigel, gen. nov. MycoBank MB835508.
Etymology: Named in honor of the mycologist Nils Lundqvist, who contributed greatly to the taxonomy of Lasiosphaeriaceae.
Type species: Lundqvistomyces karachiensis (S.I. Ahmed & Asad) Y. Marín & Stchigel.
Ascomata ostiolate, immersed to almost superficial, scattered or aggregated, brown to nearly black, pyriform, covered with brown, flexous hairs; neck black, conical to papilliform; ascomatal wall membranaceous to semi-coriaceous, dark brown to black, opaque, textura angularis. Periphyses absent or present, hyaline, filiform. Paraphyses hyaline, filiform. Asci unitunicate, eight-spored, clavate or cylindrical-clavate, thin-walled, short stipitate, with an indistinct apical ring. Ascospores biseriate, at first one-celled, ellipsoidal, becoming transversely uniseptate and two-celled; upper cell dark, ellipsoidal-fusiform to fusiform, sometimes slightly inequilateral, with a truncate base and an apical germ pore; lower cell hyaline to pale brown, conical, often collapsing; gelatinous appendages absent. Asexual morph absent or present. Conidia absent or delimited from peg-like non-proliferating structures on undifferentiated hyphae.
Notes: This genus includes two species previously identified as T. tanzaniensis and Z. karachiensis. In our phylogenetic study, both species were located in an independent terminal clade (100% bs/1 pp) in the Schizotheciaceae (clade VIII), far from the type species of Triangularia, located in the Podosporaceae (clade IV), and from Zopfiella, in the Lasiosphaeriaceae s. str. (clade V). Lundqvistomyces is characterized by its ostiolate ascomata with a membranaceous to semi-coriaceous ascomatal wall of textura angularis, asci with an indistinct apical ring, and ascospores at the first ellipsoidal, and with a smooth-walled upper cell. Lundqvistomyces karachiensis differs from L. tanzaniensis mainly in the size of the upper and lower cells of the ascospores (broader upper cell and longer lower cell in the latter species). Moreover, L. karachiensis also produces an asexual morph, which is absent in L. tanzaniensis.
Lundqvistomyces karachiensis (S.I. Ahmed & Asad) Y. Marín & Stchigel, comb. nov. MycoBank MB835509.
Basionym: Strattonia karachiensis S.I. Ahmed & Asad, Sydowia 21: 282. 1968.
Synonyms: Zopfiella karachiensis (S.I. Ahmed & Asad) Guarro, in Guarro & Cano, Trans. Brit. Mycol. Soc. 91: 589. 1988.
Triangularia karachiensis (S.I. Ahmed & Asad) Udagawa, in Udagawa, Tsuzaki & Uehara, Trans. Mycol. Soc. Japan 20: 362. 1979.
Podospora faurelii Mouch., Rev. Mycol. (Paris) 38: 109 and 112. 1975.
Lundqvistomyces tanzaniensis (R.S. Khan & J.C. Krug) Y. Marín & Stchigel, comb. nov. MycoBank MB835510.
Basionym: Triangularia tanzaniensis R.S. Khan & J.C. Krug, Mycologia 81: 865. 1990.
Synonym: Zopfiella tanzaniensis (R.S. Khan & J.C. Krug) Guarro, P.F. Cannon & Aa, Syst. Ascomycetum, Reprint of Volumes 1-4 (1982-1985) 10: 103. 1991.
Reference strains of
Podospora curvicolla, P. decidua, P. longicollis, and
P. prolifica, grouped in a well-supported independent clade (100% bs/1 pp;
Figure 2) within the family Schizothecaceae. These taxa are characterized by ascomata with tufts of long agglutinated hairs on the neck and asci usually containing more than eight ascospores. Consequently, we introduce the new genus
Pseudoechria as follows.
Pseudoechria Y. Marín & Stchigel, gen. nov. MycoBank MB835511. (
Figure 12A–I).
Etymology: Based on the morphological resemblance with Echria, which also presents tufts of rigid agglutinated long hairs on the neck of the ascomata.
Type species: Pseudoechria curvicolla (G. Winter) Y. Marín & Stchigel.
Ascomata ostiolate, immersed or superficial, scattered or aggregated, ovoid, broadly obpyriform or globose, sometimes with flexuous, light brown septate, thick-walled hairs; neck short or long, dark, opaque, with tufts of brown to black, rigid, agglutinated, septate, long, thick-walled hairs; ascomatal wall membranaceous, translucent, light olive brown to black, or yellowish to light brown with an olivaceous tinge, thin, of textura angularis. Paraphyses absent or present, more or less thread-like, soon evanescent. Asci 8- to 2048-spored, clavate to broadly ellipsoid, or broadly fusiform, short-stipitate, with or without a thick apical ring. Ascospores multiseriate, at first one-celled, hyaline, becoming transversely uniseptate; upper cell olivaceous to dark brown, ellipsoidal, sometimes with subacute ends, with an apical to subapical germ pore; lower cell hyaline, clavate, soon evanescent; gelatinous cauda attached at each end, lash-like. Asexual morph absent.
Notes:
Pseudoechria is introduced to accommodate four species of
Podospora, which form a monophyletic clade in the Schizotheciaceae (clade VIII), and were distantly related to
Podospora s. str., located in Podosporaceae (clade IV). This genus is characterized by ascomata with tufts of long agglutinated hairs on the neck and (mostly) more than eight-spored per asci. A similar morphology is observed in
Schizothecium, which also has ascomata with ornations at the apex and multi-spored asci. In fact,
Pse. longicollis was placed in
Schizothecium when it was introduced [
50]. Both genera differ in the type of ascomata ornamentation, with groups of swollen, agglutinated, short hairs or prominent protruding wall cells in
Schizothecium, while in
Pseudoechria, the vestiture comprises tufts of rigid, agglutinated, long hairs. Rigid long hairs can also be present in
Echria spp. and
Arnium hirtum, which are also located in the Schizotheciaceae. Both taxa can be easily distinguished from Pseudoechria by their one-celled ascospores. Moreover, the hairs in
A. hirtum are not grouped in tufts as in the other two genera, and this species presents also agglutinated hairs as in
Schizothecium, which is not observed in either
Echria or
Pseudoechria.
Pseudoschizothecium, located in Schizotheciaceae, and Rhypophila, located in Naviculisporaceae, also present similar morphology with an ornated ascomata apex and multi-spored asci.
Rhypophila differs in having tuberculate projections, whereas Pseudoschizothecium produces warts composed of smaller polygonal cells. Although
Triangularia setosa also presents multi-spored asci and ascomata with long and rigid hairs on the neck, as in the genus
Pseudoechria, the ascomatal hairs are mostly solitary. However, our phylogenetic analysis confirmed that this species, which was located in the family Podosporaceae (clade IV), is not related to
Pseudoechria.
Two strains of Pse. curvicolla included in the phylogenetic study grouped in a monophyletic clade (100% bs/0.99 pp) together with the ex-type strain of Pse. longicollis. However, only ITS and LSU sequences of Pse. curvicolla were available; unfortunately, these two loci display less variability. Consequently, rpb2 and tub2 sequences are necessary to confirm the potential synonymy of both species.
Pseudoechria curvicolla (G. Winter) Y. Marín, A.N. Mill. & Stchigel, comb. nov. MycoBank MB835512. (
Figure 12A–C,F,G)
Basionym: Sordaria curvicolla G. Winter, Abh. Naturf. Ges. Halle 13: 34. 1871.
Synonyms: Podospora curvicolla (G. Winter) Niessl, Hedwigia 22: 156. 1883.
Pseudoechria longicollis (L.M. Ames) Y. Marín & Stchigel, comb. nov. MycoBank MB835662.
Basionym: Schizothecium longicolle L.M. Ames, Sydowia 5: 121. 1951.
Synonyms: Pleurage longicollis (L.M. Ames) Boedijn, Persoonia 2: 312. 1962.
Podospora longicollis (L.M. Ames) J.H. Mirza & Cain, Canad. J. Bot. 47: 2028. 1970. (1969).
Pseudoechria decidua (Cailleux) Y. Marín, A.N. Mill. & Stchigel, comb. nov. MycoBank MB835513. (
Figure 12D,H)
Basionym: Podospora decidua Cailleux, Cah. Maboké 7: 102. 1969.
Pseudoechria prolifica (Cailleux) Y. Marín, A.N. Mill. & Stchigel, comb. nov. MycoBank MB835514. (
Figure 12E,I)
Basionym: Podospora prolifica Cailleux, Cah. Maboké 7: 102. 1969.
Key to species of
Pseudoechria.
| 1. Asci 8-spored | Pse. decidua |
| 1. Asci more than 8-spored | 2 |
| 2. Asci 1024-2048-spored, ascospores with upper cell up to 14 μm and lower cell up to 4 μm | Pse. prolifica |
| 2. Asci 256- or 512-spored, ascospores with upper and lower cell longer than 14 and 4 μm, respectively | 3 |
| 3. Asci about 256-spored, ascospores with upper cell up to 16 μm long and lower cell up to 9 μm | Pse. curvicolla |
| 3. Asci about 512-spored, ascospores with upper cell up to 22 μm and lower cell up to 8 μm | Pse. longicollis |
The genus Pseudoschizothecium is introduced to accommodate C. atropurpurea, which represents a basal independent lineage within the Schizotheciaceae (clade VIII), and that can be morphologically distinguished by the other members of the family by its dark purple warty ascomata and ascospores up to 5-septate.
Pseudoschizothecium Y. Marín, A.N. Mill. & Stchigel, gen. nov. MycoBank MB835515. (
Figure 13A–N).
Etymology: Based on the morphological resemblance to the genus Schizothecium.
Type species: Pseudoschizothecium atropurpureum (A.N. Mill. & Huhndorf) Y. Marín, A.N. Mill. & Stchigel.
Ascomata ostiolate, superficial, scattered or aggregated, dark purple to black, ovoid, papillate, warty toward the apex; neck conical, warty; warts small, hyaline to brownish, composed of polygonal cells; ascomatal wall pseudoparenchymatous, textura angularis. Periphyses present, filiform, hyaline, up to 2 µm in diam. Paraphyses filiform to venticrose, septate, soon evanescent. Asci unitunicate, eight-spored, elongate-clavate, long-stipitate, with a narrow and refractive apical ring, subapical globule large. Ascospores biseriate to triseriate into the asci, at first one-celled, hyaline, cylindrical, slightly sigmoid or geniculate, becoming transversely uniseptate; upper cell brown to dark brown, narrowly fusiform to ellipsoidal, occasionally 1-septate; lower cell hyaline to pale brown, sometimes brown, cylindrical, long, up to 3-septate. Asexual morph absent or present. Conidiophores reduced to conidiogenous cells. Conidiogenous cells phialidic, flask-shaped or elongate-clavate, some with distinct collarettes. Conidia hyaline, globose or ovoid.
Notes: Pseudoschizothecium is introduced to accommodate a single species, C. atropurpurea. This genus differs from all taxa included in the family by its dark purple warty ascomata and up to 5-septate ascospores (1-septate upper cell, and up to 3-septate lower cell). The same pattern of septation can be observed in T. striata, but this species can be distinguished by its areolate and striate ascomata. Ascospores with a septate lower cell are also seen in Ar. ambigua, which also has a septate upper cell. However, Ar. ambigua can be easily distinguished by its areolate cephalothecoid ascomatal wall. Our phylogenetic study demonstrated that these three taxa are not related, grouping in different families, i.e., Areotheca ambigua in Naviculisporaceae, Pseudoschizothecium atropurpureum in Schizotheciaceae, Triangularia striata in Podosporaceae. See Notes of Pseudoechria for morphological comparison with other genera with ornamented ascomata.
Pseudoschizothecium atropurpureum (A.N. Mill. & Huhndorf) Y. Marín, A.N. Mill. & Stchigel, comb. nov. MycoBank MB835516. (
Figure 13A–N).
Basionym: Cercophora atropurpurea A.N. Mill. & Huhndorf, Sydowia 53: 212. 2001.
Schizothecium Corda, Icon. Fungorum 2: 29. 1838. emend. Y. Marin & Stchigel. MycoBank MB4908.
Type species: Schizothecium fimicola Corda.
Ascomata ostiolate, immersed, semi-immersed or superficial, scattered or gregarious, light to dark brown, globose to subglobose, pyriform, or pyriform-conical, translucent, upper part ornate with groups of swollen agglutinated hairs, or with prominent protruding ascomatal wall cells; ascomatal wall membranaceous, pseudo-parenchymatous, textura angularis to textura globulosa. Paraphyses usually absent. Asci unitunicate, four-, eight-, or multi-spored, clavate to cylindrical, apically rounded, short- or long-stipitate, usually lacking apical structures, dehiscing below the apex. Ascospores at first one-celled, hyaline, becoming transversely uniseptate and two-celled; upper cell finally dark brown to blackish-brown, fusiform-obovoid, rarely heart-shaped to lunate and strongly curved and concave on one side, smooth-walled, with an upper germ pore; lower cell hyaline, rarely subhyaline, cylindrical, obclavate or obconical, usually long persistent; gelatinous appendages absent or present. Asexual morph absent or present. Conidiophores reduced to conidiogenous cells. Conidiogenous cells phialidic, flask-shaped or elongate-clavate, some with distinct collarettes. Conidia hyaline, globose or ovoid.
Notes:
Schizothecium was morphologically and phylogenetically studied by Cai et al. [
4]. As a result, the genus was delimited to 24 species, all them with ostiolate ascomata ornate with swollen agglutinated hairs or prominent protruding ascomatal wall cells. In our phylogenetic study,
P. selenospora, which produces similar ascomata, grouped in this monophyletic clade. Therefore, the new combination
S. selenosporum is proposed here, and the description of
Schizothecium is emended to incorporate its ascospore morphology. This species is distinguished from all other species of
Schizothecium by the heart-shaped to lunate shape of the ascospore upper cell. A key to distinguish all species of Schizothecium is available in Cai et al. [
4].
Because the type strain of
P. selenospora falls within the
Schizothecium clade (100% bs/1 pp;
Figure 2), the new combination for this taxon is proposed here.
Schizothecium selenosporum (Stchigel, Guarro & M. Calduch) Y. Marín & Stchigel, comb. nov. MycoBank MB835517.
Basionym: Podospora selenospora Stchigel, Guarro & M. Calduch, Mycologia 94: 554. 2002.


 ,
, 














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}