Cryptococcus: History, Epidemiology and Immune Evasion

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Classification and Habitat of the Cryptococcus spp.
3. First Historical Aspects of Cryptococcus spp.
4. Cryptococcus gattii
5. Structure and Virulence Factors
6. Cell Wall
7. Polysaccharide Capsule
7.1. Glucuronoxylomannan (GXM)
7.2. Glucuronoxylomannogalactan (GXMGal)
8. Cryptococcosis
9. Disease Development
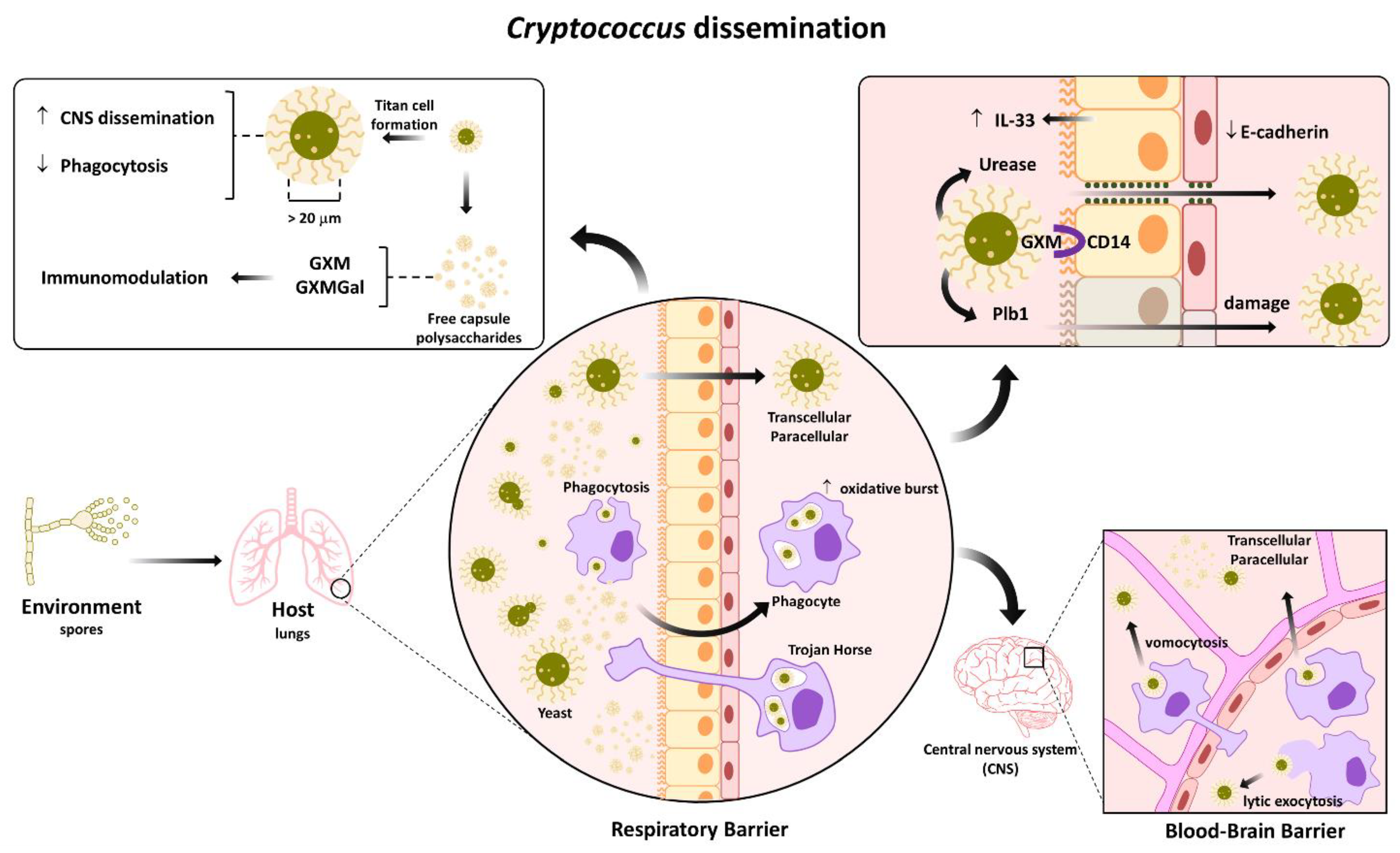
10. Cryptococcus Dissemination
11. Cryptococcosis Resistance
12. Immune Response against Cryptococcosis
12.1. The First Contact
12.2. Immunomodulation
12.3. Cell Response against the Infection
13. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maziarz, E.K.; Perfect, J.R. Cryptococcosis. Infect. Dis. Clin. N. Am. 2016, 30, 179–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Springer, D.J.; Chaturvedi, V. Projecting global occurrence of Cryptococcus gattii. Emerg. Infect. Dis 2010, 16, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J.; Fraser, J.A.; Doering, T.L.; Wang, Z.; Janbon, G.; Idnurm, A.; Bahn, Y.S. Cryptococcus neoformans and Cryptococcus gattii, the etiologic agents of cryptococcosis. Cold Spring Harb. Perspect. Med. 2014, 4, a019760. [Google Scholar] [CrossRef] [PubMed]
- Chau, T.T.; Mai, N.H.; Phu, N.H.; Nghia, H.D.; Chuong, L.V.; Sinh, D.X.; Duong, V.A.; Diep, P.T.; Campbell, J.I.; Baker, S.; et al. A prospective descriptive study of cryptococcal meningitis in HIV uninfected patients in Vietnam—High prevalence of Cryptococcus neoformans var grubii in the absence of underlying disease. BMC Infect. Dis. 2010, 10, 199. [Google Scholar] [CrossRef] [Green Version]
- Kielstein, P.; Hotzel, H.; Schmalreck, A.; Khaschabi, D.; Glawischnig, W. Occurrence of Cryptococcus spp. in excreta of pigeons and pet birds. Mycoses 2000, 43, 7–15. [Google Scholar] [CrossRef]
- Springer, D.J.; Billmyre, R.B.; Filler, E.E.; Voelz, K.; Pursall, R.; Mieczkowski, P.A.; Larsen, R.A.; Dietrich, F.S.; May, R.C.; Filler, S.G.; et al. Cryptococcus gattii VGIII isolates causing infections in HIV/AIDS patients in Southern California: Identification of the local environmental source as arboreal. PLoS Pathog. 2014, 10, e1004285. [Google Scholar] [CrossRef] [Green Version]
- Moller, L.; Kessler, K.D.; Steyn, A.; Valentine, A.J.; Botha, A. The role of Cryptococcus laurentii and mycorrhizal fungi in the nutritional physiology of Lupinus angustifolius L. hosting N2-fixing nodules. Plant. Soil 2016, 409, 345–360. [Google Scholar] [CrossRef]
- Diaz, J.H. The Disease Ecology, Epidemiology, Clinical Manifestations, and Management of Emerging Cryptococcus gattii Complex Infections. Wilderness Environ. Med. 2020, 31, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Paim, K.; Ferreira, T.B.; Andrade-Silva, L.; Mora, D.J.; Springer, D.J.; Heitman, J.; Fonseca, F.M.; Matos, D.; Melhem, M.S.; Silva-Vergara, M.L. Phylogenetic analysis of phenotypically characterized Cryptococcus laurentii isolates reveals high frequency of cryptic species. PLoS ONE 2014, 9, e108633. [Google Scholar] [CrossRef]
- Castro-Lainez, M.T.; Deliz-Aguirre, R.; Antunez, D.; Cruz-Codina, M.; Cahuayme-Zuniga, L.; Vitale, K.; Sierra-Hoffman, M.; Midturi, J.K. Cryptococcus laurentii meningitis in a non-HIV patient. IDCases 2019, 18, e00612. [Google Scholar] [CrossRef]
- Salazar-Leal, J.I.; Ramirez-Montelongo, S.M.; Lopez Luis, B.A. Clinical significance of nosocomial Cryptococcus laurentii in urine: A case series. Infect. Control Hosp. Epidemiol. 2019, 40, 1442–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, M.; Mishra, A.K.; Singh, S.K. Cryptococcus laurentii fungemia in a low birth weight preterm neonate: India. J. Infect. Public Health 2018, 11, 896–897. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, T.G.; Perfect, J.R. Cryptococcosis in the era of AIDS--100 years after the discovery of Cryptococcus neoformans. Clin. Microbiol. Rev. 1995, 8, 515–548. [Google Scholar] [CrossRef] [PubMed]
- Neiman, A.M. Ascospore formation in the yeast Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2005, 69, 565–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnett, J.A. A history of research on yeasts 14: Medical yeasts part 2, Cryptococcus neoformans. Yeast 2010, 27, 875–904. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.E.; Sorensen, L.J.; Walls, K.W. The antigenic composition of Cryptococcus neoformans. V. A survey of cross-reactions among strains of Cryptococcus and other antigens. J. Bacteriol. 1953, 66, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Heitman, J. The biology of the Cryptococcus neoformans species complex. Annu Rev. Microbiol. 2006, 60, 69–105. [Google Scholar] [CrossRef]
- Franzot, S.P.; Salkin, I.F.; Casadevall, A. Cryptococcus neoformans var. grubii: Separate varietal status for Cryptococcus neoformans serotype A isolates. J. Clin. Microbiol. 1999, 37, 838–840. [Google Scholar] [CrossRef] [Green Version]
- Litvintseva, A.P.; Thakur, R.; Vilgalys, R.; Mitchell, T.G. Multilocus sequence typing reveals three genetic subpopulations of Cryptococcus neoformans var. grubii (serotype A), including a unique population in Botswana. Genetics 2006, 172, 2223–2238. [Google Scholar] [CrossRef] [Green Version]
- Meyer, W.; Marszewska, K.; Amirmostofian, M.; Igreja, R.P.; Hardtke, C.; Methling, K.; Viviani, M.A.; Chindamporn, A.; Sukroongreung, S.; John, M.A.; et al. Molecular typing of global isolates of Cryptococcus neoformans var. neoformans by polymerase chain reaction fingerprinting and randomly amplified polymorphic DNA-a pilot study to standardize techniques on which to base a detailed epidemiological survey. Electrophoresis 1999, 20, 1790–1799. [Google Scholar] [CrossRef]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Tintelnot, K.; Hagen, F.; Han, C.O.; Seibold, M.; Rickerts, V.; Boekhout, T. Pitfalls in Serological Diagnosis of Cryptococcus gattii Infections. Med. Mycol. 2015, 53, 874–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iverson, S.A.; Chiller, T.; Beekmann, S.; Polgreen, P.M.; Harris, J. Recognition and diagnosis of Cryptococcus gattii infections in the United States. Emerg. Infect. Dis. 2012, 18, 1012–1015. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J.; Polacheck, I.; Bennett, J.E. Improved diagnostic medium for separation of Cryptococcus neoformans var. neoformans (serotypes A and D) and Cryptococcus neoformans var. gattii (serotypes B and C). J. Clin. Microbiol. 1982, 15, 535–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polacheck, I.; Kwon-Chung, K.J. Canavanine resistance in Cryptococcus neoformans. Antimicrob. Agents Chemother. 1986, 29, 468–473. [Google Scholar] [CrossRef] [Green Version]
- Klein, K.R.; Hall, L.; Deml, S.M.; Rysavy, J.M.; Wohlfiel, S.L.; Wengenack, N.L. Identification of Cryptococcus gattii by use of L-canavanine glycine bromothymol blue medium and DNA sequencing. J. Clin. Microbiol. 2009, 47, 3669–3672. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.C.; Meyer, W.; Sorrell, T.C. Cryptococcus gattii infections. Clin. Microbiol. Rev. 2014, 27, 980–1024. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, C.A.; Kronstad, J.W.; Taylor, G.; Warren, R.; Yuen, M.; Hu, G.; Jung, W.H.; Sham, A.; Kidd, S.E.; Tangen, K.; et al. Genome variation in Cryptococcus gattii, an emerging pathogen of immunocompetent hosts. mBio 2011, 2, e00342-10. [Google Scholar] [CrossRef] [Green Version]
- Springer, D.J.; Phadke, S.; Billmyre, B.; Heitman, J. Cryptococcus gattii, no longer an accidental pathogen? Curr. Fungal. Infect. Rep. 2012, 6, 245–256. [Google Scholar] [CrossRef]
- Da Silva, B.K.; Freire, A.K.; Bentes Ados, S.; Sampaio Ide, L.; Santos, L.O.; Dos Santos, M.S.; De Souza, J.V. Characterization of clinical isolates of the Cryptococcus neoformans-Cryptococcus gattii species complex from the Amazonas State in Brazil. Rev. Iberoam. Micol. 2012, 29, 40–43. [Google Scholar] [CrossRef]
- Dos Santos Bentes, A.; Wanke, B.; Dos Santos Lazéra, M.; Freire, A.K.L.; da Silva Júnior, R.M.; Rocha, D.F.S.; Pinheiro, S.B.; Zelski, S.E.; Matsuura, A.B.J.; da Rocha, L.C.; et al. Cryptococcus gattii VGII isolated from native forest and river in Northern Brazil. Braz. J. Microbiol. 2019, 50, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M.M.; Almeida-Paes, R.; Brito-Santos, F.; Nascimento, C.R.; Fialho, M.M.; Trilles, L.; Morales, B.P.; da Silva, S.A.; Santos, W.; Santos, L.O.; et al. Comparative antifungal susceptibility analyses of Cryptococcus neoformans VNI and Cryptococcus gattii VGII from the Brazilian Amazon Region by the Etest, Vitek 2, and the Clinical and Laboratory Standards Institute broth microdilution methods. Med. Mycol. 2019, 57, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.M.; Wanke, B.; Lazéra Mdos, S.; Trilles, L.; Barbosa, G.G.; Macedo, R.C.; Cavalcanti Mdo, A.; Eulálio, K.D.; Castro, J.A.; Silva, A.S.; et al. Genotypes of Cryptococcus neoformans and Cryptococcus gattii as agents of endemic cryptococcosis in Teresina, Piauí (northeastern Brazil). Mem Inst. Oswaldo Cruz 2011, 106, 725–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruyama, F.H.; de Paula, D.A.J.; Menezes, I.G.; Favalessa, O.C.; Hahn, R.C.; de Almeida, A.; Sousa, V.R.F.; Nakazato, L.; Dutra, V. Genetic Diversity of the Cryptococcus gattii Species Complex in Mato Grosso State, Brazil. Mycopathologia 2019, 184, 45–51. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Bennett, J.E. High prevalence of Cryptococcus neoformans var gattii in tropical and subtropical regions. Zentralbl. Bakteriol. Mikrobiol. Hyg. A 1984, 257, 213–218. [Google Scholar]
- Grizante Barião, P.H.; Tonani, L.; Cocio, T.A.; Martinez, R.; Nascimento, E.; von Zeska Kress, M.R. Molecular typing, in vitro susceptibility and virulence of Cryptococcus neoformans/Cryptococcus gattii species complex clinical isolates from south-eastern Brazil. Mycoses 2020, 63, 1341–1351. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, E.; Barião, P.H.G.; Kress, M.; Vilar, F.C.; Santana, R.C.; Gaspar, G.G.; Martinez, R. Cryptococcosis by Cryptococcus neoformans/Cryptococcus gattii Species Complexes in non-HIV-Infected Patients in Southeastern Brazil. Rev. Soc. Bras. Med. Trop. 2021, 54, e01692021. [Google Scholar] [CrossRef]
- McGill, S.; Malik, R.; Saul, N.; Beetson, S.; Secombe, C.; Robertson, I.; Irwin, P. Cryptococcosis in domestic animals in Western Australia: A retrospective study from 1995-2006. Med. Mycol. 2009, 47, 625–639. [Google Scholar] [CrossRef] [Green Version]
- Duncan, C.; Schwantje, H.; Stephen, C.; Campbell, J.; Bartlett, K. Cryptococcus gattii in wildlife of Vancouver Island, British Columbia, Canada. J. Wildl. Dis. 2006, 42, 175–178. [Google Scholar] [CrossRef] [Green Version]
- Stephen, C.; Lester, S.; Black, W.; Fyfe, M.; Raverty, S. Multispecies outbreak of cryptococcosis on southern Vancouver Island, British Columbia. Can. Vet. J. 2002, 43, 792–794. [Google Scholar]
- Kidd, S.E.; Hagen, F.; Tscharke, R.L.; Huynh, M.; Bartlett, K.H.; Fyfe, M.; Macdougall, L.; Boekhout, T.; Kwon-Chung, K.J.; Meyer, W. A rare genotype of Cryptococcus gattii caused the cryptococcosis outbreak on Vancouver Island (British Columbia, Canada). Proc. Natl. Acad. Sci. USA 2004, 101, 17258–17263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrnes, E.J., 3rd; Li, W.; Lewit, Y.; Perfect, J.R.; Carter, D.A.; Cox, G.M.; Heitman, J. First reported case of Cryptococcus gattii in the Southeastern USA: Implications for travel-associated acquisition of an emerging pathogen. PLoS ONE 2009, 4, e5851. [Google Scholar] [CrossRef] [PubMed]
- Bruner, K.T.; Franco-Paredes, C.; Henao-Martinez, A.F.; Steele, G.M.; Chastain, D.B. Cryptococcus gattii Complex Infections in HIV-Infected Patients, Southeastern United States. Emerg. Infect. Dis. 2018, 24, 1998–2002. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.R.; Lockhart, S.R.; Debess, E.; Marsden-Haug, N.; Goldoft, M.; Wohrle, R.; Lee, S.; Smelser, C.; Park, B.; Chiller, T. Cryptococcus gattii in the United States: Clinical aspects of infection with an emerging pathogen. Clin. Infect. Dis. 2011, 53, 1188–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, K.; Bartlett, K.H.; Baer, R.; Byrnes, E.; Galanis, E.; Heitman, J.; Hoang, L.; Leslie, M.J.; MacDougall, L.; Magill, S.S.; et al. Spread of Cryptococcus gattii into Pacific Northwest region of the United States. Emerg. Infect. Dis. 2009, 15, 1185–1191. [Google Scholar] [CrossRef]
- Souto, A.C.; Bonfietti, L.X.; Ferreira-Paim, K.; Trilles, L.; Martins, M.; Ribeiro-Alves, M.; Pham, C.D.; Martins, L.; Dos Santos, W.; Chang, M.; et al. Population Genetic Analysis Reveals a High Genetic Diversity in the Brazilian Cryptococcus gattii VGII Population and Shifts the Global Origin from the Amazon Rainforest to the Semi-arid Desert in the Northeast of Brazil. PLoS Negl. Trop. Dis. 2016, 10, e0004885. [Google Scholar] [CrossRef] [Green Version]
- Akins, P.T.; Jian, B. The Frozen Brain State of Cryptococcus gattii: A Globe-Trotting, Tropical, Neurotropic Fungus. Neurocrit. Care 2019, 30, 272–279. [Google Scholar] [CrossRef]
- May, R.C.; Stone, N.R.; Wiesner, D.L.; Bicanic, T.; Nielsen, K. Cryptococcus: From environmental saprophyte to global pathogen. Nat. Rev. Microbiol. 2016, 14, 106–117. [Google Scholar] [CrossRef]
- Casadevall, A.; Freij, J.B.; Hann-Soden, C.; Taylor, J. Continental Drift and Speciation of the Cryptococcus neoformans and Cryptococcus gattii Species Complexes. mSphere 2017, 2, e00103-17. [Google Scholar] [CrossRef] [Green Version]
- Farrer, R.A.; Desjardins, C.A.; Sakthikumar, S.; Gujja, S.; Saif, S.; Zeng, Q.; Chen, Y.; Voelz, K.; Heitman, J.; May, R.C.; et al. Genome Evolution and Innovation across the Four Major Lineages of Cryptococcus gattii. mBio 2015, 6, e00868-15. [Google Scholar] [CrossRef] [Green Version]
- Meyer, W. Cryptococcus gattii in the Age of Whole-Genome Sequencing. mBio 2015, 6, e01761-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litvintseva, A.P.; Mitchell, T.G. Population genetic analyses reveal the African origin and strain variation of Cryptococcus neoformans var. grubii. PLoS Pathog. 2012, 8, e1002495. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, K.L.; Murphy, J.W. What makes Cryptococcus neoformans a pathogen? Emerg. Infect. Dis. 1998, 4, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Srikanta, D.; Santiago-Tirado, F.H.; Doering, T.L. Cryptococcus neoformans: Historical curiosity to modern pathogen. Yeast 2014, 31, 47–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odom, A.; Muir, S.; Lim, E.; Toffaletti, D.L.; Perfect, J.; Heitman, J. Calcineurin is required for virulence of Cryptococcus neoformans. EMBO J. 1997, 16, 2576–2589. [Google Scholar] [CrossRef]
- Narasipura, S.D.; Chaturvedi, V.; Chaturvedi, S. Characterization of Cryptococcus neoformans variety gattii SOD2 reveals distinct roles of the two superoxide dismutases in fungal biology and virulence. Mol. Microbiol. 2005, 55, 1782–1800. [Google Scholar] [CrossRef]
- Ngamskulrungroj, P.; Himmelreich, U.; Breger, J.A.; Wilson, C.; Chayakulkeeree, M.; Krockenberger, M.B.; Malik, R.; Daniel, H.M.; Toffaletti, D.; Djordjevic, J.T.; et al. The trehalose synthesis pathway is an integral part of the virulence composite for Cryptococcus gattii. Infect. Immun. 2009, 77, 4584–4596. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Casadevall, A. Susceptibility of melanized and nonmelanized Cryptococcus neoformans to nitrogen- and oxygen-derived oxidants. Infect. Immun. 1994, 62, 3004–3007. [Google Scholar] [CrossRef] [Green Version]
- Blasi, E.; Barluzzi, R.; Mazzolla, R.; Tancini, B.; Saleppico, S.; Puliti, M.; Pitzurra, L.; Bistoni, F. Role of nitric oxide and melanogenesis in the accomplishment of anticryptococcal activity by the BV-2 microglial cell line. J. Neuroimmunol. 1995, 58, 111–116. [Google Scholar] [CrossRef]
- Doering, T.L.; Nosanchuk, J.D.; Roberts, W.K.; Casadevall, A. Melanin as a potential cryptococcal defence against microbicidal proteins. Med. Mycol. 1999, 37, 175–181. [Google Scholar] [CrossRef]
- Ikeda, R.; Sugita, T.; Jacobson, E.S.; Shinoda, T. Effects of melanin upon susceptibility of Cryptococcus to antifungals. Microbiol. Immunol. 2003, 47, 271–277. [Google Scholar] [CrossRef] [PubMed]
- van Duin, D.; Casadevall, A.; Nosanchuk, J.D. Melanization of Cryptococcus neoformans and Histoplasma capsulatum reduces their susceptibilities to amphotericin B and caspofungin. Antimicrob. Agents Chemother. 2002, 46, 3394–3400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khajo, A.; Bryan, R.A.; Friedman, M.; Burger, R.M.; Levitsky, Y.; Casadevall, A.; Magliozzo, R.S.; Dadachova, E. Protection of melanized Cryptococcus neoformans from lethal dose gamma irradiation involves changes in melanin’s chemical structure and paramagnetism. PLoS ONE 2011, 6, e25092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadachova, E.; Bryan, R.A.; Howell, R.C.; Schweitzer, A.D.; Aisen, P.; Nosanchuk, J.D.; Casadevall, A. The radioprotective properties of fungal melanin are a function of its chemical composition, stable radical presence and spatial arrangement. Pigment. Cell Melanoma Res. 2008, 21, 192–199. [Google Scholar] [CrossRef]
- Zhu, X.; Williamson, P.R. Role of laccase in the biology and virulence of Cryptococcus neoformans. FEMS Yeast Res. 2004, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Waterman, S.R.; Hacham, M.; Panepinto, J.; Hu, G.; Shin, S.; Williamson, P.R. Cell wall targeting of laccase of Cryptococcus neoformans during infection of mice. Infect. Immun. 2007, 75, 714–722. [Google Scholar] [CrossRef] [Green Version]
- Chrissian, C.; Camacho, E.; Fu, M.S.; Prados-Rosales, R.; Chatterjee, S.; Cordero, R.J.B.; Lodge, J.K.; Casadevall, A.; Stark, R.E. Melanin deposition in two Cryptococcus species depends on cell-wall composition and flexibility. J. Biol. Chem. 2020, 295, 1815–1828. [Google Scholar] [CrossRef]
- Baker, L.G.; Specht, C.A.; Donlin, M.J.; Lodge, J.K. Chitosan, the deacetylated form of chitin, is necessary for cell wall integrity in Cryptococcus neoformans. Eukaryot. Cell 2007, 6, 855–867. [Google Scholar] [CrossRef] [Green Version]
- Lam, W.C.; Upadhya, R.; Specht, C.A.; Ragsdale, A.E.; Hole, C.R.; Levitz, S.M.; Lodge, J.K. Chitosan Biosynthesis and Virulence in the Human Fungal Pathogen Cryptococcus gattii. mSphere 2019, 4, e00644-19. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, V.; Wong, B.; Newman, S.L. Oxidative killing of Cryptococcus neoformans by human neutrophils. Evidence that fungal mannitol protects by scavenging reactive oxygen intermediates. J. Immunol. 1996, 156, 3836–3840. [Google Scholar]
- Wong, B.; Perfect, J.R.; Beggs, S.; Wright, K.A. Production of the hexitol D-mannitol by Cryptococcus neoformans in vitro and in rabbits with experimental meningitis. Infect. Immun. 1990, 58, 1664–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, G.M.; McDade, H.C.; Chen, S.C.; Tucker, S.C.; Gottfredsson, M.; Wright, L.C.; Sorrell, T.C.; Leidich, S.D.; Casadevall, A.; Ghannoum, M.A.; et al. Extracellular phospholipase activity is a virulence factor for Cryptococcus neoformans. Mol. Microbiol. 2001, 39, 166–175. [Google Scholar] [CrossRef]
- Lev, S.; Desmarini, D.; Li, C.; Chayakulkeeree, M.; Traven, A.; Sorrell, T.C.; Djordjevic, J.T. Phospholipase C of Cryptococcus neoformans regulates homeostasis and virulence by providing inositol trisphosphate as a substrate for Arg1 kinase. Infect. Immun. 2013, 81, 1245–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.C.; Muller, M.; Zhou, J.Z.; Wright, L.C.; Sorrell, T.C. Phospholipase activity in Cryptococcus neoformans: A new virulence factor? J. Infect. Dis. 1997, 175, 414–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruvada, R.; Zhu, L.; Pearce, D.; Zheng, Y.; Perfect, J.; Kwon-Chung, K.J.; Kim, K.S. Cryptococcus neoformans phospholipase B1 activates host cell Rac1 for traversal across the blood-brain barrier. Cell Microbiol. 2012, 14, 1544–1553. [Google Scholar] [CrossRef] [Green Version]
- Bose, I.; Reese, A.J.; Ory, J.J.; Janbon, G.; Doering, T.L. A yeast under cover: The capsule of Cryptococcus neoformans. Eukaryot. Cell 2003, 2, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Doering, T.L. How sweet it is! Cell wall biogenesis and polysaccharide capsule formation in Cryptococcus neoformans. Annu. Rev. Microbiol. 2009, 63, 223–247. [Google Scholar] [CrossRef] [Green Version]
- Chrissian, C.; Lin, C.P.; Camacho, E.; Casadevall, A.; Neiman, A.M.; Stark, R.E. Unconventional Constituents and Shared Molecular Architecture of the Melanized Cell Wall of C. neoformans and Spore Wall of S. cerevisiae. J. Fungi 2020, 6, 329. [Google Scholar] [CrossRef]
- Hernandez-Chavez, M.J.; Perez-Garcia, L.A.; Nino-Vega, G.A.; Mora-Montes, H.M. Fungal Strategies to Evade the Host Immune Recognition. J. Fungi 2017, 3, 51. [Google Scholar] [CrossRef] [Green Version]
- Hopke, A.; Brown, A.J.P.; Hall, R.A.; Wheeler, R.T. Dynamic Fungal Cell Wall Architecture in Stress Adaptation and Immune Evasion. Trends Microbiol. 2018, 26, 284–295. [Google Scholar] [CrossRef]
- Kumar, P.; Heiss, C.; Santiago-Tirado, F.H.; Black, I.; Azadi, P.; Doering, T.L. Pbx proteins in Cryptococcus neoformans cell wall remodeling and capsule assembly. Eukaryot. Cell 2014, 13, 560–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, L.; Sood, P.; Lenardon, M.D.; Milne, G.; Olson, J.; Jensen, G.; Wolf, J.; Casadevall, A.; Adler-Moore, J.; Gow, N.A.R. The Viscoelastic Properties of the Fungal Cell Wall Allow Traffic of AmBisome as Intact Liposome Vesicles. mBio 2018, 9, e02383-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Meara, T.R.; Holmer, S.M.; Selvig, K.; Dietrich, F.; Alspaugh, J.A. Cryptococcus neoformans Rim101 is associated with cell wall remodeling and evasion of the host immune responses. mBio 2013, 4, e00522-12. [Google Scholar] [CrossRef] [Green Version]
- Esher, S.K.; Ost, K.S.; Kohlbrenner, M.A.; Pianalto, K.M.; Telzrow, C.L.; Campuzano, A.; Nichols, C.B.; Munro, C.; Wormley, F.L., Jr.; Alspaugh, J.A. Defects in intracellular trafficking of fungal cell wall synthases lead to aberrant host immune recognition. PLoS Pathog. 2018, 14, e1007126. [Google Scholar] [CrossRef]
- James, P.G.; Cherniak, R.; Jones, R.G.; Stortz, C.A.; Reiss, E. Cell-wall glucans of Cryptococcus neoformans Cap 67. Carbohydr. Res. 1990, 198, 23–38. [Google Scholar] [CrossRef]
- Banks, I.R.; Specht, C.A.; Donlin, M.J.; Gerik, K.J.; Levitz, S.M.; Lodge, J.K. A chitin synthase and its regulator protein are critical for chitosan production and growth of the fungal pathogen Cryptococcus neoformans. Eukaryot. Cell 2005, 4, 1902–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reese, A.J.; Yoneda, A.; Breger, J.A.; Beauvais, A.; Liu, H.; Griffith, C.L.; Bose, I.; Kim, M.J.; Skau, C.; Yang, S.; et al. Loss of cell wall alpha(1-3) glucan affects Cryptococcus neoformans from ultrastructure to virulence. Mol. Microbiol. 2007, 63, 1385–1398. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.A.; Li, L.X.; Doering, T.L. Unraveling synthesis of the cryptococcal cell wall and capsule. Glycobiology 2018, 28, 719–730. [Google Scholar] [CrossRef]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The Fungal Cell Wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2019, 10, 2993. [Google Scholar] [CrossRef]
- Cabib, E.; Blanco, N.; Grau, C.; Rodriguez-Pena, J.M.; Arroyo, J. Crh1p and Crh2p are required for the cross-linking of chitin to beta(1-6)glucan in the Saccharomyces cerevisiae cell wall. Mol. Microbiol. 2007, 63, 921–935. [Google Scholar] [CrossRef]
- Lu, C.F.; Montijn, R.C.; Brown, J.L.; Klis, F.; Kurjan, J.; Bussey, H.; Lipke, P.N. Glycosyl phosphatidylinositol-dependent cross-linking of alpha-agglutinin and beta 1,6-glucan in the Saccharomyces cerevisiae cell wall. J. Cell Biol. 1995, 128, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Macekova, D.; Farkas, V.; Kishida, E.; Takeo, K. Ecto-glycanases and metabolic stability of the capsule in Cryptococcus neoformans. J. Basic Microbiol. 2006, 46, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.E.; Esher, S.K.; Alspaugh, J.A. Chitin: A “Hidden Figure” in the Fungal Cell Wall. Curr Top. Microbiol. Immunol. 2020, 425, 83–111. [Google Scholar] [CrossRef]
- Aiba, S. Studies on chitosan: 4. Lysozymic hydrolysis of partially N-acetylated chitosans. Int. J. Biol. Macromol. 1992, 14, 225–228. [Google Scholar] [CrossRef]
- Baker, L.G.; Specht, C.A.; Lodge, J.K. Chitinases are essential for sexual development but not vegetative growth in Cryptococcus neoformans. Eukaryot. Cell 2009, 8, 1692–1705. [Google Scholar] [CrossRef] [Green Version]
- Eigenheer, R.A.; Jin Lee, Y.; Blumwald, E.; Phinney, B.S.; Gelli, A. Extracellular glycosylphosphatidylinositol-anchored mannoproteins and proteases of Cryptococcus neoformans. FEMS Yeast Res. 2007, 7, 499–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Q.Y.; de Groot, P.W.; Dekker, H.L.; de Jong, L.; Klis, F.M.; de Koster, C.G. Comprehensive proteomic analysis of Saccharomyces cerevisiae cell walls: Identification of proteins covalently attached via glycosylphosphatidylinositol remnants or mild alkali-sensitive linkages. J. Biol. Chem. 2005, 280, 20894–20901. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Yu, Z.; Jia, D.; Xie, Z.; Zhang, K.; Xia, Z.; Lei, L.; Qiao, M. The contribution of Pir protein family to yeast cell surface display. Appl. Microbiol. Biotechnol. 2014, 98, 2897–2905. [Google Scholar] [CrossRef]
- McClelland, E.E.; Bernhardt, P.; Casadevall, A. Estimating the relative contributions of virulence factors for pathogenic microbes. Infect. Immun. 2006, 74, 1500–1504. [Google Scholar] [CrossRef] [Green Version]
- Decote-Ricardo, D.; LaRocque-de-Freitas, I.F.; Rocha, J.D.B.; Nascimento, D.O.; Nunes, M.P.; Morrot, A.; Freire-de-Lima, L.; Previato, J.O.; Mendonca-Previato, L.; Freire-de-Lima, C.G. Immunomodulatory Role of Capsular Polysaccharides Constituents of Cryptococcus neoformans. Front. Med. 2019, 6, 129. [Google Scholar] [CrossRef] [Green Version]
- Araujo Gde, S.; Fonseca, F.L.; Pontes, B.; Torres, A.; Cordero, R.J.; Zancope-Oliveira, R.M.; Casadevall, A.; Viana, N.B.; Nimrichter, L.; Rodrigues, M.L.; et al. Capsules from pathogenic and non-pathogenic Cryptococcus spp. manifest significant differences in structure and ability to protect against phagocytic cells. PLoS ONE 2012, 7, e29561. [Google Scholar] [CrossRef] [PubMed]
- Cherniak, R.; Valafar, H.; Morris, L.C.; Valafar, F. Cryptococcus neoformans chemotyping by quantitative analysis of 1H nuclear magnetic resonance spectra of glucuronoxylomannans with a computer-simulated artificial neural network. Clin. Diagn Lab. Immunol. 1998, 5, 146–159. [Google Scholar] [CrossRef]
- Vartivarian, S.E.; Reyes, G.H.; Jacobson, E.S.; James, P.G.; Cherniak, R.; Mumaw, V.R.; Tingler, M.J. Localization of mannoprotein in Cryptococcus neoformans. J. Bacteriol. 1989, 171, 6850–6852. [Google Scholar] [CrossRef] [Green Version]
- McFadden, D.C.; Fries, B.C.; Wang, F.; Casadevall, A. Capsule structural heterogeneity and antigenic variation in Cryptococcus neoformans. Eukaryot. Cell 2007, 6, 1464–1473. [Google Scholar] [CrossRef] [Green Version]
- McFadden, D.C.; De Jesus, M.; Casadevall, A. The physical properties of the capsular polysaccharides from Cryptococcus neoformans suggest features for capsule construction. J. Biol. Chem. 2006, 281, 1868–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherniak, R.; Jones, R.G.; Reiss, E. Structure determination of Cryptococcus neoformans serotype A-variant glucuronoxylomannan by 13C-n.m.r. spectroscopy. Carbohydr. Res. 1988, 172, 113–138. [Google Scholar] [CrossRef]
- Janbon, G.; Himmelreich, U.; Moyrand, F.; Improvisi, L.; Dromer, F. Cas1p is a membrane protein necessary for the O-acetylation of the Cryptococcus neoformans capsular polysaccharide. Mol. Microbiol. 2001, 42, 453–467. [Google Scholar] [CrossRef]
- Cherniak, R.; Sundstrom, J.B. Polysaccharide antigens of the capsule of Cryptococcus neoformans. Infect. Immun. 1994, 62, 1507–1512. [Google Scholar] [CrossRef] [Green Version]
- Villena, S.N.; Pinheiro, R.O.; Pinheiro, C.S.; Nunes, M.P.; Takiya, C.M.; DosReis, G.A.; Previato, J.O.; Mendonca-Previato, L.; Freire-de-Lima, C.G. Capsular polysaccharides galactoxylomannan and glucuronoxylomannan from Cryptococcus neoformans induce macrophage apoptosis mediated by Fas ligand. Cell Microbiol. 2008, 10, 1274–1285. [Google Scholar] [CrossRef]
- Nimrichter, L.; Frases, S.; Cinelli, L.P.; Viana, N.B.; Nakouzi, A.; Travassos, L.R.; Casadevall, A.; Rodrigues, M.L. Self-aggregation of Cryptococcus neoformans capsular glucuronoxylomannan is dependent on divalent cations. Eukaryot. Cell 2007, 6, 1400–1410. [Google Scholar] [CrossRef] [Green Version]
- Vaishnav, V.V.; Bacon, B.E.; O’Neill, M.; Cherniak, R. Structural characterization of the galactoxylomannan of Cryptococcus neoformans Cap67. Carbohydr. Res. 1998, 306, 315–330. [Google Scholar] [CrossRef]
- Wozniak, K.L.; Levitz, S.M. Isolation and purification of antigenic components of Cryptococcus. Methods Mol. Biol. 2009, 470, 71–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, F.L.; Reis, F.C.G.; Sena, B.A.G.; Jozefowicz, L.J.; Kmetzsch, L.; Rodrigues, M.L. The Overlooked Glycan Components of the Cryptococcus Capsule. Curr Top. Microbiol. Immunol. 2019, 422, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Heiss, C.; Skowyra, M.L.; Liu, H.; Klutts, J.S.; Wang, Z.; Williams, M.; Srikanta, D.; Beverley, S.M.; Azadi, P.; Doering, T.L. Unusual galactofuranose modification of a capsule polysaccharide in the pathogenic yeast Cryptococcus neoformans. J. Biol. Chem. 2013, 288, 10994–11003. [Google Scholar] [CrossRef] [Green Version]
- Previato, J.O.; Vinogradov, E.; Maes, E.; Fonseca, L.M.; Guerardel, Y.; Oliveira, P.A.V.; Mendonca-Previato, L. Distribution of the O-acetyl groups and beta-galactofuranose units in galactoxylomannans of the opportunistic fungus Cryptococcus neoformans. Glycobiology 2017, 27, 582–592. [Google Scholar] [CrossRef] [Green Version]
- Bryan, R.A.; Zaragoza, O.; Zhang, T.; Ortiz, G.; Casadevall, A.; Dadachova, E. Radiological studies reveal radial differences in the architecture of the polysaccharide capsule of Cryptococcus neoformans. Eukaryot. Cell 2005, 4, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Levitz, S.M.; Nong, S.; Mansour, M.K.; Huang, C.; Specht, C.A. Molecular characterization of a mannoprotein with homology to chitin deacetylases that stimulates T cell responses to Cryptococcus neoformans. Proc. Natl. Acad. Sci. USA 2001, 98, 10422–10427. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.L.; Alvarez, M.; Fonseca, F.L.; Casadevall, A. Binding of the wheat germ lectin to Cryptococcus neoformans suggests an association of chitinlike structures with yeast budding and capsular glucuronoxylomannan. Eukaryot. Cell 2008, 7, 602–609. [Google Scholar] [CrossRef] [Green Version]
- Jesus, M.D.; Nicola, A.M.; Chow, S.K.; Lee, I.R.; Nong, S.; Specht, C.A.; Levitz, S.M.; Casadevall, A. Glucuronoxylomannan, galactoxylomannan, and mannoprotein occupy spatially separate and discrete regions in the capsule of Cryptococcus neoformans. Virulence 2010, 1, 500–508. [Google Scholar] [CrossRef] [Green Version]
- Reuwsaat, J.C.V.; Motta, H.; Garcia, A.W.A.; Vasconcelos, C.B.; Marques, B.M.; Oliveira, N.K.; Rodrigues, J.; Ferrareze, P.A.G.; Frases, S.; Lopes, W.; et al. A Predicted Mannoprotein Participates in Cryptococcus gattii Capsular Structure. mSphere 2018, 3, e00023-18. [Google Scholar] [CrossRef] [Green Version]
- Cordero, R.J.; Pontes, B.; Guimaraes, A.J.; Martinez, L.R.; Rivera, J.; Fries, B.C.; Nimrichter, L.; Rodrigues, M.L.; Viana, N.B.; Casadevall, A. Chronological aging is associated with biophysical and chemical changes in the capsule of Cryptococcus neoformans. Infect. Immun. 2011, 79, 4990–5000. [Google Scholar] [CrossRef] [Green Version]
- Lian, T.; Simmer, M.I.; D’Souza, C.A.; Steen, B.R.; Zuyderduyn, S.D.; Jones, S.J.; Marra, M.A.; Kronstad, J.W. Iron-regulated transcription and capsule formation in the fungal pathogen Cryptococcus neoformans. Mol. Microbiol. 2005, 55, 1452–1472. [Google Scholar] [CrossRef]
- Cadieux, B.; Lian, T.; Hu, G.; Wang, J.; Biondo, C.; Teti, G.; Liu, V.; Murphy, M.E.; Creagh, A.L.; Kronstad, J.W. The Mannoprotein Cig1 supports iron acquisition from heme and virulence in the pathogenic fungus Cryptococcus neoformans. J. Infect. Dis. 2013, 207, 1339–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urai, M.; Kaneko, Y.; Ueno, K.; Okubo, Y.; Aizawa, T.; Fukazawa, H.; Sugita, T.; Ohno, H.; Shibuya, K.; Kinjo, Y.; et al. Evasion of Innate Immune Responses by the Highly Virulent Cr.ryptococcus gattii by Altering Capsule Glucuronoxylomannan Structure. Front. Cell Infect. Microbiol. 2015, 5, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, N.; Li, L.; McFadden, D.C.; Banarjee, U.; Wang, X.; Cook, E.; Fries, B.C. Phenotypic switching in a Cryptococcus neoformans variety gattii strain is associated with changes in virulence and promotes dissemination to the central nervous system. Infect. Immun. 2006, 74, 896–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diniz-Lima, I.; da Fonseca, L.M.; Dos Reis, J.S.; Rodrigues da Costa Santos, M.A.; da Costa, K.M.; do Nascimento Santos, C.A.; Barcelos, P.M.; Guimaraes-Pinto, K.; Filardy, A.A.; Freire-de-Lima, M.E.; et al. The Sweet Side of Fungal Infections: Structural Glycan Diversity and Its Importance for Pathogenic Adaptation. Medicines 2022, 9, 37. [Google Scholar] [CrossRef]
- Goldman, D.L.; Lee, S.C.; Casadevall, A. Tissue localization of Cryptococcus neoformans glucuronoxylomannan in the presence and absence of specific antibody. Infect. Immun. 1995, 63, 3448–3453. [Google Scholar] [CrossRef] [Green Version]
- Rocha, J.D.; Nascimento, M.T.; Decote-Ricardo, D.; Corte-Real, S.; Morrot, A.; Heise, N.; Nunes, M.P.; Previato, J.O.; Mendonca-Previato, L.; DosReis, G.A.; et al. Capsular polysaccharides from Cryptococcus neoformans modulate production of neutrophil extracellular traps (NETs) by human neutrophils. Sci. Rep. 2015, 5, 8008. [Google Scholar] [CrossRef]
- Cleare, W.; Casadevall, A. Scanning electron microscopy of encapsulated and non-encapsulated Cryptococcus neoformans and the effect of glucose on capsular polysaccharide release. Med. Mycol. 1999, 37, 235–243. [Google Scholar]
- Nosanchuk, J.D.; Casadevall, A. Cellular charge of Cryptococcus neoformans: Contributions from the capsular polysaccharide, melanin, and monoclonal antibody binding. Infect. Immun. 1997, 65, 1836–1841. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.S.; Filler, S.G.; Alcouloumre, M.S.; Kozel, T.R.; Edwards, J.E., Jr.; Ghannoum, M.A. Adherence to and damage of endothelial cells by Cryptococcus neoformans in vitro: Role of the capsule. Infect. Immun. 1995, 63, 4368–4374. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Pirofski, L.A. Opsonization of Cryptococcus neoformans by human anticryptococcal glucuronoxylomannan antibodies. Infect. Immun. 1996, 64, 3446–3450. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Lee, S.C.; Casadevall, A. Antibodies to Cryptococcus neoformans glucuronoxylomannan enhance antifungal activity of murine macrophages. Infect. Immun. 1995, 63, 573–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaragoza, O.; Taborda, C.P.; Casadevall, A. The efficacy of complement-mediated phagocytosis of Cryptococcus neoformans is dependent on the location of C3 in the polysaccharide capsule and involves both direct and indirect C3-mediated interactions. Eur. J. Immunol. 2003, 33, 1957–1967. [Google Scholar] [CrossRef]
- Rossbacher, J.; Shlomchik, M.J. The B cell receptor itself can activate complement to provide the complement receptor 1/2 ligand required to enhance B cell immune responses in vivo. J. Exp. Med. 2003, 198, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Ahearn, J.M.; Fischer, M.B.; Croix, D.; Goerg, S.; Ma, M.; Xia, J.; Zhou, X.; Howard, R.G.; Rothstein, T.L.; Carroll, M.C. Disruption of the Cr2 locus results in a reduction in B-1a cells and in an impaired B cell response to T-dependent antigen. Immunity 1996, 4, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Taborda, C.P.; Casadevall, A. CR3 (CD11b/CD18) and CR4 (CD11c/CD18) are involved in complement-independent antibody-mediated phagocytosis of Cryptococcus neoformans. Immunity 2002, 16, 791–802. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.M.; Murphy, J.W. Cryptococcal polysaccharides bind to CD18 on human neutrophils. Infect. Immun. 1997, 65, 557–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syme, R.M.; Spurrell, J.C.; Amankwah, E.K.; Green, F.H.; Mody, C.H. Primary dendritic cells phagocytose Cryptococcus neoformans via mannose receptors and Fcgamma receptor II for presentation to T lymphocytes. Infect. Immun. 2002, 70, 5972–5981. [Google Scholar] [CrossRef] [Green Version]
- Yauch, L.E.; Mansour, M.K.; Levitz, S.M. Receptor-mediated clearance of Cryptococcus neoformans capsular polysaccharide in vivo. Infect. Immun. 2005, 73, 8429–8432. [Google Scholar] [CrossRef] [Green Version]
- Shoham, S.; Huang, C.; Chen, J.M.; Golenbock, D.T.; Levitz, S.M. Toll-like receptor 4 mediates intracellular signaling without TNF-alpha release in response to Cryptococcus neoformans polysaccharide capsule. J. Immunol. 2001, 166, 4620–4626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, F.M.; Fonseca, F.L.; Figueiredo, R.T.; Bozza, M.T.; Casadevall, A.; Nimrichter, L.; Rodrigues, M.L. Binding of glucuronoxylomannan to the CD14 receptor in human A549 alveolar cells induces interleukin-8 production. Clin. Vaccine Immunol 2007, 14, 94–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vecchiarelli, A.; Retini, C.; Pietrella, D.; Monari, C.; Tascini, C.; Beccari, T.; Kozel, T.R. Downregulation by cryptococcal polysaccharide of tumor necrosis factor alpha and interleukin-1 beta secretion from human monocytes. Infect. Immun. 1995, 63, 2919–2923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walenkamp, A.M.; Chaka, W.S.; Verheul, A.F.; Vaishnav, V.V.; Cherniak, R.; Coenjaerts, F.E.; Hoepelman, I.M. Cryptococcus neoformans and its cell wall components induce similar cytokine profiles in human peripheral blood mononuclear cells despite differences in structure. FEMS Immunol. Med. Microbiol. 1999, 26, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiapello, L.S.; Baronetti, J.L.; Aoki, M.P.; Gea, S.; Rubinstein, H.; Masih, D.T. Immunosuppression, interleukin-10 synthesis and apoptosis are induced in rats inoculated with Cryptococcus neoformans glucuronoxylomannan. Immunology 2004, 113, 392–400. [Google Scholar] [CrossRef]
- Scriven, J.E.; Graham, L.M.; Schutz, C.; Scriba, T.J.; Wilkinson, K.A.; Wilkinson, R.J.; Boulware, D.R.; Urban, B.C.; Lalloo, D.G.; Meintjes, G. A Glucuronoxylomannan-Associated Immune Signature, Characterized by Monocyte Deactivation and an Increased Interleukin 10 Level, Is a Predictor of Death in Cryptococcal Meningitis. J. Infect. Dis 2016, 213, 1725–1734. [Google Scholar] [CrossRef] [Green Version]
- Chiapello, L.S.; Aoki, M.P.; Rubinstein, H.R.; Masih, D.T. Apoptosis induction by glucuronoxylomannan of Cryptococcus neoformans. Med. Mycol. 2003, 41, 347–353. [Google Scholar] [CrossRef]
- Chiapello, L.S.; Baronetti, J.L.; Garro, A.P.; Spesso, M.F.; Masih, D.T. Cryptococcus neoformans glucuronoxylomannan induces macrophage apoptosis mediated by nitric oxide in a caspase-independent pathway. Int. Immunol. 2008, 20, 1527–1541. [Google Scholar] [CrossRef] [Green Version]
- Monari, C.; Paganelli, F.; Bistoni, F.; Kozel, T.R.; Vecchiarelli, A. Capsular polysaccharide induction of apoptosis by intrinsic and extrinsic mechanisms. Cell Microbiol. 2008, 10, 2129–2137. [Google Scholar] [CrossRef]
- Retini, C.; Vecchiarelli, A.; Monari, C.; Bistoni, F.; Kozel, T.R. Encapsulation of Cryptococcus neoformans with glucuronoxylomannan inhibits the antigen-presenting capacity of monocytes. Infect. Immun. 1998, 66, 664–669. [Google Scholar] [CrossRef] [Green Version]
- Mariano Andrade, R.; Monteiro Almeida, G.; Alexandre DosReis, G.; Alves Melo Bento, C. Glucuronoxylomannan of Cryptococcus neoformans exacerbates in vitro yeast cell growth by interleukin 10-dependent inhibition of CD4+ T lymphocyte responses. Cell Immunol. 2003, 222, 116–125. [Google Scholar] [CrossRef]
- Yauch, L.E.; Lam, J.S.; Levitz, S.M. Direct inhibition of T-cell responses by the Cryptococcus capsular polysaccharide glucuronoxylomannan. PLoS Pathog. 2006, 2, e120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albuquerque, P.C.; Fonseca, F.L.; Dutra, F.F.; Bozza, M.T.; Frases, S.; Casadevall, A.; Rodrigues, M.L. Cryptococcus neoformans glucuronoxylomannan fractions of different molecular masses are functionally distinct. Future Microbiol. 2014, 9, 147–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monari, C.; Pericolini, E.; Bistoni, G.; Casadevall, A.; Kozel, T.R.; Vecchiarelli, A. Cryptococcus neoformans capsular glucuronoxylomannan induces expression of fas ligand in macrophages. J. Immunol. 2005, 174, 3461–3468. [Google Scholar] [CrossRef] [Green Version]
- Heiss, C.; Klutts, J.S.; Wang, Z.; Doering, T.L.; Azadi, P. The structure of Cryptococcus neoformans galactoxylomannan contains beta-D-glucuronic acid. Carbohydr. Res. 2009, 344, 915–920. [Google Scholar] [CrossRef]
- Dorokhova, V.S.; Gerbst, A.G.; Komarova, B.S.; Previato, J.O.; Previato, L.M.; Dmitrenok, A.S.; Shashkov, A.S.; Krylov, V.B.; Nifantiev, N.E. Synthesis and conformational analysis of vicinally branched trisaccharide beta-d-Galf-(1 ≥ 2)-[beta-d-Galf-(1 ≥ 3)-]-alpha-Galp from Cryptococcus neoformans galactoxylomannan. Org. Biomol. Chem. 2021, 19, 2923–2931. [Google Scholar] [CrossRef]
- Pericolini, E.; Gabrielli, E.; Cenci, E.; De Jesus, M.; Bistoni, F.; Casadevall, A.; Vecchiarelli, A. Involvement of glycoreceptors in galactoxylomannan-induced T cell death. J. Immunol. 2009, 182, 6003–6010. [Google Scholar] [CrossRef] [Green Version]
- De Jesus, M.; Nicola, A.M.; Frases, S.; Lee, I.R.; Mieses, S.; Casadevall, A. Galactoxylomannan-mediated immunological paralysis results from specific B cell depletion in the context of widespread immune system damage. J. Immunol. 2009, 183, 3885–3894. [Google Scholar] [CrossRef]
- Pericolini, E.; Gabrielli, E.; Bistoni, G.; Cenci, E.; Perito, S.; Chow, S.K.; Riuzzi, F.; Donato, R.; Casadevall, A.; Vecchiarelli, A. Role of CD45 signaling pathway in galactoxylomannan-induced T cell damage. PLoS ONE 2010, 5, e12720. [Google Scholar] [CrossRef] [Green Version]
- LaRocque-de-Freitas, I.F.; Rocha, J.D.B.; Nunes, M.P.; Oliveira, P.A.V.; Nascimento, D.O.; Freire-de-Lima, L.; Takiya, C.M.; Morrot, A.; Decote-Ricardo, D.; Previato, J.O.; et al. Involvement of the capsular GalXM-induced IL-17 cytokine in the control of Cryptococcus neoformans infection. Sci. Rep. 2018, 8, 16378. [Google Scholar] [CrossRef]
- Delfino, D.; Cianci, L.; Lupis, E.; Celeste, A.; Petrelli, M.L.; Curro, F.; Cusumano, V.; Teti, G. Interleukin-6 production by human monocytes stimulated with Cryptococcus neoformans components. Infect. Immun. 1997, 65, 2454–2456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snarr, B.D.; Qureshi, S.T.; Sheppard, D.C. Immune Recognition of Fungal Polysaccharides. J. Fungi 2017, 3, 47. [Google Scholar] [CrossRef] [PubMed]
- Pericolini, E.; Gabrielli, E.; Alunno, A.; Bartoloni Bocci, E.; Perito, S.; Chow, S.K.; Cenci, E.; Casadevall, A.; Gerli, R.; Vecchiarelli, A. Functional improvement of regulatory T cells from rheumatoid arthritis subjects induced by capsular polysaccharide glucuronoxylomannogalactan. PLoS ONE 2014, 9, e111163. [Google Scholar] [CrossRef] [PubMed]
- Pericolini, E.; Alunno, A.; Gabrielli, E.; Bartoloni, E.; Cenci, E.; Chow, S.K.; Bistoni, G.; Casadevall, A.; Gerli, R.; Vecchiarelli, A. The microbial capsular polysaccharide galactoxylomannan inhibits IL-17A production in circulating T cells from rheumatoid arthritis patients. PLoS ONE 2013, 8, e53336. [Google Scholar] [CrossRef]
- Vecchiarelli, A.; Pericolini, E.; Gabrielli, E.; Chow, S.K.; Bistoni, F.; Cenci, E.; Casadevall, A. Cryptococcus neoformans galactoxylomannan is a potent negative immunomodulator, inspiring new approaches in anti-inflammatory immunotherapy. Immunotherapy 2011, 3, 997–1005. [Google Scholar] [CrossRef] [Green Version]
- Garelnabi, M.; May, R.C. Variability in innate host immune responses to cryptococcosis. Mem Inst. Oswaldo Cruz 2018, 113, e180060. [Google Scholar] [CrossRef]
- Jha, M.K.; Mohanty, A.; Gupta, P. Cryptococcus gattii meningitis in a diabetic adult in South India. J. Family Med. Prim. Care 2019, 8, 1253–1256. [Google Scholar] [CrossRef]
- Prasad, K.T.; Sehgal, I.S.; Shivaprakash, M.R.; Dhooria, S. Uncommon mycosis in a patient with diabetes. BMJ Case Rep. 2016, 2016, bcr2016214453. [Google Scholar] [CrossRef]
- Paccoud, O.; Bougnoux, M.E.; Desnos-Ollivier, M.; Varet, B.; Lortholary, O.; Lanternier, F. Cryptococcus gattii in Patients with Lymphoid Neoplasms: An Illustration of Evolutive Host-Fungus Interactions. J. Fungi 2021, 7, 212. [Google Scholar] [CrossRef]
- Bartlett, K.H.; Cheng, P.Y.; Duncan, C.; Galanis, E.; Hoang, L.; Kidd, S.; Lee, M.K.; Lester, S.; MacDougall, L.; Mak, S.; et al. A decade of experience: Cryptococcus gattii in British Columbia. Mycopathologia 2012, 173, 311–319. [Google Scholar] [CrossRef]
- MacDougall, L.; Fyfe, M.; Romney, M.; Starr, M.; Galanis, E. Risk factors for Cryptococcus gattii infection, British Columbia, Canada. Emerg. Infect. Dis. 2011, 17, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Xie, J.X.; Zeng, Q.S.; Chen, G.Q.; Zhong, S.Q.; Zhong, N.S. Retrospective analysis of 76 immunocompetent patients with primary pulmonary cryptococcosis. Lung 2012, 190, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Deok-jong Yoo, S.; Worodria, W.; Davis, J.L.; Cattamanchi, A.; den Boon, S.; Kyeyune, R.; Kisembo, H.; Huang, L. The prevalence and clinical course of HIV-associated pulmonary cryptococcosis in Uganda. J. Acquir. Immune Defic. Syndr. 2010, 54, 269–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, J.R.; Lindsley, M.D.; Henchaichon, S.; Poonwan, N.; Naorat, S.; Prapasiri, P.; Chantra, S.; Ruamcharoen, F.; Chang, L.S.; Chittaganpitch, M.; et al. High prevalence of cryptococcal infection among HIV-infected patients hospitalized with pneumonia in Thailand. Clin. Infect. Dis. 2012, 54, e43–e50. [Google Scholar] [CrossRef] [PubMed]
- Suwatanapongched, T.; Sangsatra, W.; Boonsarngsuk, V.; Watcharananan, S.P.; Incharoen, P. Clinical and radiologic manifestations of pulmonary cryptococcosis in immunocompetent patients and their outcomes after treatment. Diagn Interv. Radiol. 2013, 19, 438–446. [Google Scholar] [CrossRef]
- Herkert, P.F.; Hagen, F.; Pinheiro, R.L.; Muro, M.D.; Meis, J.F.; Queiroz-Telles, F. Ecoepidemiology of Cryptococcus gattii in Developing Countries. J. Fungi 2017, 3, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damasceno-Escoura, A.H.; de Souza, M.L.; de Oliveira Nunes, F.; Pardi, T.C.; Gazotto, F.C.; Florentino, D.H.; Mora, D.J.; Silva-Vergara, M.L. Epidemiological, Clinical and Outcome Aspects of Patients with Cryptococcosis Caused by Cryptococcus gattii from a Non-endemic Area of Brazil. Mycopathologia 2019, 184, 65–71. [Google Scholar] [CrossRef]
- Mitchell, D.H.; Sorrell, T.C.; Allworth, A.M.; Heath, C.H.; McGregor, A.R.; Papanaoum, K.; Richards, M.J.; Gottlieb, T. Cryptococcal disease of the CNS in immunocompetent hosts: Influence of cryptococcal variety on clinical manifestations and outcome. Clin. Infect. Dis. 1995, 20, 611–616. [Google Scholar] [CrossRef]
- Anzai, M.C.; Lazéra Mdos, S.; Wanke, B.; Trilles, L.; Dutra, V.; de Paula, D.A.; Nakazato, L.; Takahara, D.T.; Simi, W.B.; Hahn, R.C. Cryptococcus gattii VGII in a Plathymenia reticulata hollow in Cuiabá, Mato Grosso, Brazil. Mycoses 2014, 57, 414–418. [Google Scholar] [CrossRef]
- Shirley, R.M.; Baddley, J.W. Cryptococcal lung disease. Curr. Opin. Pulm. Med. 2009, 15, 254–260. [Google Scholar] [CrossRef]
- Velagapudi, R.; Hsueh, Y.P.; Geunes-Boyer, S.; Wright, J.R.; Heitman, J. Spores as infectious propagules of Cryptococcus neoformans. Infect. Immun. 2009, 77, 4345–4355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Hermoso, D.; Janbon, G.; Dromer, F. Epidemiological evidence for dormant Cryptococcus neoformans infection. J. Clin. Microbiol. 1999, 37, 3204–3209. [Google Scholar] [CrossRef] [Green Version]
- MacDougall, L.; Fyfe, M. Emergence of Cryptococcus gattii in a novel environment provides clues to its incubation period. J. Clin. Microbiol. 2006, 44, 1851–1852. [Google Scholar] [CrossRef] [Green Version]
- Kerkering, T.M.; Duma, R.J.; Shadomy, S. The evolution of pulmonary cryptococcosis: Clinical implications from a study of 41 patients with and without compromising host factors. Ann. Intern. Med. 1981, 94, 611–616. [Google Scholar] [CrossRef] [PubMed]
- da Silva-Junior, E.B.; Firmino-Cruz, L.; Guimaraes-de-Oliveira, J.C.; De-Medeiros, J.V.R.; de Oliveira Nascimento, D.; Freire-de-Lima, M.; de Brito-Gitirana, L.; Morrot, A.; Previato, J.O.; Mendonca-Previato, L.; et al. The role of Toll-like receptor 9 in a murine model of Cryptococcus gattii infection. Sci. Rep. 2021, 11, 1407. [Google Scholar] [CrossRef] [PubMed]
- Hommel, B.; Mukaremera, L.; Cordero, R.J.B.; Coelho, C.; Desjardins, C.A.; Sturny-Leclere, A.; Janbon, G.; Perfect, J.R.; Fraser, J.A.; Casadevall, A.; et al. Titan cells formation in Cryptococcus neoformans is finely tuned by environmental conditions and modulated by positive and negative genetic regulators. PLoS Pathog. 2018, 14, e1006982. [Google Scholar] [CrossRef] [Green Version]
- Diniz-Lima, I.; da Rosa, P.R.; da Silva-Junior, E.B.; Guimarães-de-Oliveira, J.C.; de Freitas, E.O.; de Oliveira Nascimento, D.; Morrot, A.; Nimrichter, L.; Previato, J.O.; Mendonça-Previato, L.; et al. X-linked immunodeficient (XID) mice exhibit high susceptibility to Cryptococcus gattii infection. Sci. Rep. 2021, 11, 18397. [Google Scholar] [CrossRef]
- Dos Santos, M.H.; Machado, M.P.; Kumaresan, P.R.; da Silva, T.A. Titan Cells and Yeast Forms of Cryptococcus neoformans and Cryptococcus gattii Are Recognized by GXMR-CAR. Microorganisms 2021, 9, 1886. [Google Scholar] [CrossRef]
- Denham, S.T.; Brown, J.C.S. Mechanisms of Pulmonary Escape and Dissemination by Cryptococcus neoformans. J. Fungi 2018, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Trevijano-Contador, N.; Pianalto, K.M.; Nichols, C.B.; Zaragoza, O.; Alspaugh, J.A.; Pirofski, L.A. Human IgM Inhibits the Formation of Titan-Like Cells in Cryptococcus neoformans. Infect. Immun 2020, 88, e00046-20. [Google Scholar] [CrossRef]
- Voelz, K.; May, R.C. Cryptococcal interactions with the host immune system. Eukaryot. Cell 2010, 9, 835–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strickland, A.B.; Shi, M. Mechanisms of fungal dissemination. Cell Mol. Life Sci. 2021, 78, 3219–3238. [Google Scholar] [CrossRef] [PubMed]
- Sorrell, T.C.; Juillard, P.G.; Djordjevic, J.T.; Kaufman-Francis, K.; Dietmann, A.; Milonig, A.; Combes, V.; Grau, G.E. Cryptococcal transmigration across a model brain blood-barrier: Evidence of the Trojan horse mechanism and differences between Cryptococcus neoformans var. grubii strain H99 and Cryptococcus gattii strain R265. Microbes Infect. 2016, 18, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, D.C.; Filler, S.G. Host cell invasion by medically important fungi. Cold Spring Harb. Perspect. Med. 2014, 5, a019687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, H.K.; Huang, T.Y.; Wu, A.Y.; Chen, H.H.; Liu, C.P.; Jong, A. How Cryptococcus interacts with the blood-brain barrier. Future Microbiol. 2015, 10, 1669–1682. [Google Scholar] [CrossRef]
- Chang, Y.C.; Stins, M.F.; McCaffery, M.J.; Miller, G.F.; Pare, D.R.; Dam, T.; Paul-Satyaseela, M.; Kim, K.S.; Kwon-Chung, K.J. Cryptococcal yeast cells invade the central nervous system via transcellular penetration of the blood-brain barrier. Infect. Immun. 2004, 72, 4985–4995. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Beckerman, K.P.; Reznik, S.; Madan, R.P.; Goldman, D.L. Transplacental transmission of Cryptococcus neoformans to an HIV-exposed premature neonate. J. Perinatol. 2012, 32, 235–237. [Google Scholar] [CrossRef]
- Taylor-Smith, L.M. Cryptococcus-Epithelial Interactions. J. Fungi 2017, 3, 53. [Google Scholar] [CrossRef] [Green Version]
- Ganendren, R.; Carter, E.; Sorrell, T.; Widmer, F.; Wright, L. Phospholipase B activity enhances adhesion of Cryptococcus neoformans to a human lung epithelial cell line. Microbes Infect. 2006, 8, 1006–1015. [Google Scholar] [CrossRef]
- Na Pombejra, S.; Salemi, M.; Phinney, B.S.; Gelli, A. The Metalloprotease, Mpr1, Engages AnnexinA2 to Promote the Transcytosis of Fungal Cells across the Blood-Brain Barrier. Front. Cell Infect. Microbiol. 2017, 7, 296. [Google Scholar] [CrossRef] [Green Version]
- Jong, A.; Wu, C.H.; Chen, H.M.; Luo, F.; Kwon-Chung, K.J.; Chang, Y.C.; Lamunyon, C.W.; Plaas, A.; Huang, S.H. Identification and characterization of CPS1 as a hyaluronic acid synthase contributing to the pathogenesis of Cryptococcus neoformans infection. Eukaryot. Cell 2007, 6, 1486–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyen, L.; Muller, U.; Siegemund, S.; Schulze, B.; Protschka, M.; Alber, G.; Piehler, D. Lung epithelium is the major source of IL-33 and is regulated by IL-33-dependent and IL-33-independent mechanisms in pulmonary cryptococcosis. Pathog. Dis. 2016, 74, ftw086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutherford, J.C. The emerging role of urease as a general microbial virulence factor. PLoS Pathog. 2014, 10, e1004062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olszewski, M.A.; Noverr, M.C.; Chen, G.H.; Toews, G.B.; Cox, G.M.; Perfect, J.R.; Huffnagle, G.B. Urease expression by Cryptococcus neoformans promotes microvascular sequestration, thereby enhancing central nervous system invasion. Am. J. Pathol. 2004, 164, 1761–1771. [Google Scholar] [CrossRef] [Green Version]
- Siafakas, A.R.; Sorrell, T.C.; Wright, L.C.; Wilson, C.; Larsen, M.; Boadle, R.; Williamson, P.R.; Djordjevic, J.T. Cell wall-linked cryptococcal phospholipase B1 is a source of secreted enzyme and a determinant of cell wall integrity. J. Biol. Chem. 2007, 282, 37508–37514. [Google Scholar] [CrossRef] [Green Version]
- Charlier, C.; Nielsen, K.; Daou, S.; Brigitte, M.; Chretien, F.; Dromer, F. Evidence of a role for monocytes in dissemination and brain invasion by Cryptococcus neoformans. Infect. Immun. 2009, 77, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Chang, Z.L.; Netski, D.; Thorkildson, P.; Kozel, T.R. Binding and internalization of glucuronoxylomannan, the major capsular polysaccharide of Cryptococcus neoformans, by murine peritoneal macrophages. Infect. Immun. 2006, 74, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Levitz, S.M.; Nong, S.H.; Seetoo, K.F.; Harrison, T.S.; Speizer, R.A.; Simons, E.R. Cryptococcus neoformans resides in an acidic phagolysosome of human macrophages. Infect. Immun. 1999, 67, 885–890. [Google Scholar] [CrossRef] [Green Version]
- Kronstad, J.W.; Hu, G.; Jung, W.H. An encapsulation of iron homeostasis and virulence in Cryptococcus neoformans. Trends Microbiol. 2013, 21, 457–465. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.J.; Moyer, S.; Hoke, E.S.; Sionov, E.; Mayer-Barber, K.D.; Barber, D.L.; Cai, H.; Jenkins, L.; Walter, P.J.; Chang, Y.C.; et al. Pulmonary Iron Limitation Induced by Exogenous Type I IFN Protects Mice from Cryptococcus gattii Independently of T Cells. mBio 2019, 10, e00799-19. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.M.; Dixon, E.F.; May, R.C. The fungal pathogen Cryptococcus neoformans manipulates macrophage phagosome maturation. Cell Microbiol 2015, 17, 702–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamil, K.; Polyak, M.J.; Feehan, D.D.; Surmanowicz, P.; Stack, D.; Li, S.S.; Ogbomo, H.; Olszewski, M.; Ganguly, A.; Mody, C.H. Phagosomal F-Actin Retention by Cryptococcus gattii Induces Dendritic Cell Immunoparalysis. mBio 2020, 11, e01821-20. [Google Scholar] [CrossRef] [PubMed]
- Eastman, A.J.; He, X.; Qiu, Y.; Davis, M.J.; Vedula, P.; Lyons, D.M.; Park, Y.D.; Hardison, S.E.; Malachowski, A.N.; Osterholzer, J.J.; et al. Cryptococcal heat shock protein 70 homolog Ssa1 contributes to pulmonary expansion of Cryptococcus neoformans during the afferent phase of the immune response by promoting macrophage M2 polarization. J. Immunol. 2015, 194, 5999–6010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, S.; Olszewski, M.A.; Tsang, T.M.; McDonald, R.A.; Toews, G.B.; Huffnagle, G.B. Effect of cytokine interplay on macrophage polarization during chronic pulmonary infection with Cryptococcus neoformans. Infect. Immun. 2011, 79, 1915–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huffnagle, G.B. Role of cytokines in T cell immunity to a pulmonary Cryptococcus neoformans infection. Biol. Signals 1996, 5, 215–222. [Google Scholar] [CrossRef]
- Haddow, L.J.; Colebunders, R.; Meintjes, G.; Lawn, S.D.; Elliott, J.H.; Manabe, Y.C.; Bohjanen, P.R.; Sungkanuparph, S.; Easterbrook, P.J.; French, M.A.; et al. Cryptococcal immune reconstitution inflammatory syndrome in HIV-1-infected individuals: Proposed clinical case definitions. Lancet Infect. Dis. 2010, 10, 791–802. [Google Scholar] [CrossRef] [Green Version]
- Seaton, R.A.; Verma, N.; Naraqi, S.; Wembri, J.P.; Warrell, D.A. The effect of corticosteroids on visual loss in Cryptococcus neoformans var. gattii meningitis. Trans. R Soc. Trop. Med. Hyg. 1997, 91, 50–52. [Google Scholar] [CrossRef]
- Perfect, J.R.; Dismukes, W.E.; Dromer, F.; Goldman, D.L.; Graybill, J.R.; Hamill, R.J.; Harrison, T.S.; Larsen, R.A.; Lortholary, O.; Nguyen, M.H.; et al. Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the infectious diseases society of america. Clin. Infect. Dis. 2010, 50, 291–322. [Google Scholar] [CrossRef] [Green Version]
- Trilles, L.; Fernández-Torres, B.; Lazéra Mdos, S.; Wanke, B.; Guarro, J. In vitro antifungal susceptibility of Cryptococcus gattii. J. Clin. Microbiol. 2004, 42, 4815–4817. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, G.F.; Santos, J.R.; Costa, M.C.; Holanda, R.A.; Denadai, Â.M.; Freitas, G.J.; Santos, Á.R.; Tavares, P.B.; Paixão, T.A.; Santos, D.A. Heteroresistance to Itraconazole Alters the Morphology and Increases the Virulence of Cryptococcus gattii. Antimicrob. Agents Chemother. 2015, 59, 4600–4609. [Google Scholar] [CrossRef] [Green Version]
- Gibson, J.F.; Johnston, S.A. Immunity to Cryptococcus neoformans and C. gattii during cryptococcosis. Fungal. Genet. Biol. 2015, 78, 76–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishore, U.; Bernal, A.L.; Kamran, M.F.; Saxena, S.; Singh, M.; Sarma, P.U.; Madan, T.; Chakraborty, T. Surfactant proteins SP-A and SP-D in human health and disease. Arch. Immunol. Ther. Exp. 2005, 53, 399–417. [Google Scholar]
- Wright, J.R. Host defense functions of pulmonary surfactant. Biol. Neonate 2004, 85, 326–332. [Google Scholar] [CrossRef]
- Giles, S.S.; Zaas, A.K.; Reidy, M.F.; Perfect, J.R.; Wright, J.R. Cryptococcus neoformans is resistant to surfactant protein A mediated host defense mechanisms. PLoS ONE 2007, 2, e1370. [Google Scholar] [CrossRef] [PubMed]
- Holmer, S.M.; Evans, K.S.; Asfaw, Y.G.; Saini, D.; Schell, W.A.; Ledford, J.G.; Frothingham, R.; Wright, J.R.; Sempowski, G.D.; Perfect, J.R. Impact of surfactant protein D, interleukin-5, and eosinophilia on Cryptococcosis. Infect. Immun. 2014, 82, 683–693. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, R.; Zoellner, H.; Sorrell, T.; Wilson, C.; Donald, C.; Djordjevic, J.; Shounan, Y.; Wright, L. Role of extracellular phospholipases and mononuclear phagocytes in dissemination of cryptococcosis in a murine model. Infect. Immun. 2004, 72, 2229–2239. [Google Scholar] [CrossRef] [Green Version]
- Coelho, C.; Bocca, A.L.; Casadevall, A. The intracellular life of Cryptococcus neoformans. Annu. Rev. Pathol. 2014, 9, 219–238. [Google Scholar] [CrossRef] [Green Version]
- Cross, C.E.; Bancroft, G.J. Ingestion of acapsular Cryptococcus neoformans occurs via mannose and beta-glucan receptors, resulting in cytokine production and increased phagocytosis of the encapsulated form. Infect. Immun. 1995, 63, 2604–2611. [Google Scholar] [CrossRef] [Green Version]
- Giles, S.S.; Dagenais, T.R.; Botts, M.R.; Keller, N.P.; Hull, C.M. Elucidating the pathogenesis of spores from the human fungal pathogen Cryptococcus neoformans. Infect. Immun. 2009, 77, 3491–3500. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Kinjo, T.; Saijo, S.; Miyazato, A.; Adachi, Y.; Ohno, N.; Fujita, J.; Kaku, M.; Iwakura, Y.; Kawakami, K. Dectin-1 is not required for the host defense to Cryptococcus neoformans. Microbiol. Immunol. 2007, 51, 1115–1119. [Google Scholar] [CrossRef]
- Means, T.K.; Mylonakis, E.; Tampakakis, E.; Colvin, R.A.; Seung, E.; Puckett, L.; Tai, M.F.; Stewart, C.R.; Pukkila-Worley, R.; Hickman, S.E.; et al. Evolutionarily conserved recognition and innate immunity to fungal pathogens by the scavenger receptors SCARF1 and CD36. J. Exp. Med. 2009, 206, 637–653. [Google Scholar] [CrossRef] [PubMed]
- Biondo, C.; Midiri, A.; Messina, L.; Tomasello, F.; Garufi, G.; Catania, M.R.; Bombaci, M.; Beninati, C.; Teti, G.; Mancuso, G. MyD88 and TLR2, but not TLR4, are required for host defense against Cryptococcus neoformans. Eur. J. Immunol. 2005, 35, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Dan, J.M.; Kelly, R.M.; Lee, C.K.; Levitz, S.M. Role of the mannose receptor in a murine model of Cryptococcus neoformans infection. Infect. Immun. 2008, 76, 2362–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.; Chen, M.; Fa, Z.; Lu, A.; Fang, W.; Sun, B.; Chen, C.; Liao, W.; Meng, G. Acapsular Cryptococcus neoformans activates the NLRP3 inflammasome. Microbes Infect. 2014, 16, 845–854. [Google Scholar] [CrossRef]
- Guillot, L.; Carroll, S.F.; Badawy, M.; Qureshi, S.T. Cryptococcus neoformans induces IL-8 secretion and CXCL1 expression by human bronchial epithelial cells. Respir. Res. 2008, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.C.; Santos, J.R.; Ribeiro, M.J.; Freitas, G.J.; Bastos, R.W.; Ferreira, G.F.; Miranda, A.S.; Arifa, R.D.; Santos, P.C.; Martins Fdos, S.; et al. The absence of microbiota delays the inflammatory response to Cryptococcus gattii. Int. J. Med. Microbiol. 2016, 306, 187–195. [Google Scholar] [CrossRef]
- Ikeda-Dantsuji, Y.; Ohno, H.; Tanabe, K.; Umeyama, T.; Ueno, K.; Nagi, M.; Yamagoe, S.; Kinjo, Y.; Miyazaki, Y. Interferon-γ promotes phagocytosis of Cryptococcus neoformans but not Cryptococcus gattii by murine macrophages. J. Infect. Chemother. 2015, 21, 831–836. [Google Scholar] [CrossRef]
- Herkert, P.F.; Dos Santos, J.C.; Hagen, F.; Ribeiro-Dias, F.; Queiroz-Telles, F.; Netea, M.G.; Meis, J.F.; Joosten, L.A.B. Differential In Vitro Cytokine Induction by the Species of Cryptococcus gattii Complex. Infect. Immun. 2018, 86, e00958-17. [Google Scholar] [CrossRef] [Green Version]
- Nicola, A.M.; Albuquerque, P.; Martinez, L.R.; Dal-Rosso, R.A.; Saylor, C.; De Jesus, M.; Nosanchuk, J.D.; Casadevall, A. Macrophage autophagy in immunity to Cryptococcus neoformans and Candida albicans. Infect. Immun. 2012, 80, 3065–3076. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Feng, F.; Wang, H.; Zhang, Y.; Chen, Y. The Inflammasome NLRC4 Protects against Cryptococcus gattii by Inducing the Classic Caspase-1 to Activate the Pyroptosis Signal. J. Healthc. Eng. 2022, 2022, 7355485. [Google Scholar] [CrossRef]
- Ueno, K.; Yanagihara, N.; Otani, Y.; Shimizu, K.; Kinjo, Y.; Miyazaki, Y. Neutrophil-mediated antifungal activity against highly virulent Cryptococcus gattii strain R265. Med. Mycol. 2019, 57, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.L.; Spurrell, J.C.; Wang, J.F.; Neely, G.G.; Epelman, S.; Krensky, A.M.; Mody, C.H. CD8 T cell-mediated killing of Cryptococcus neoformans requires granulysin and is dependent on CD4 T cells and IL-15. J. Immunol. 2002, 169, 5787–5795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniam, K.S.; Datta, K.; Quintero, E.; Manix, C.; Marks, M.S.; Pirofski, L.A. The absence of serum IgM enhances the susceptibility of mice to pulmonary challenge with Cryptococcus neoformans. J. Immunol. 2010, 184, 5755–5767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufaud, C.; Rivera, J.; Rohatgi, S.; Pirofski, L.A. Naive B cells reduce fungal dissemination in Cryptococcus neoformans infected Rag1(-/-) mice. Virulence 2018, 9, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Rohatgi, S.; Pirofski, L.A. Molecular characterization of the early B cell response to pulmonary Cryptococcus neoformans infection. J. Immunol. 2012, 189, 5820–5830. [Google Scholar] [CrossRef] [Green Version]
- Szymczak, W.A.; Davis, M.J.; Lundy, S.K.; Dufaud, C.; Olszewski, M.; Pirofski, L.A. X-linked immunodeficient mice exhibit enhanced susceptibility to Cryptococcus neoformans Infection. mBio 2013, 4, e00265-13. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, K.; Metzger, B.; Hanau, L.H.; Guh, A.; Rucker, L.; Badri, S.; Pirofski, L.A. IgM(+) memory B cell expression predicts HIV-associated cryptococcosis status. J. Infect. Dis. 2009, 200, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Rohatgi, S.; Nakouzi, A.; Carreno, L.J.; Slosar-Cheah, M.; Kuniholm, M.H.; Wang, T.; Pappas, P.G.; Pirofski, L.A. Antibody and B Cell Subset Perturbations in Human Immunodeficiency Virus-Uninfected Patients With Cryptococcosis. Open Forum. Infect. Dis. 2018, 5, ofx255. [Google Scholar] [CrossRef]
- Pieper, K.; Grimbacher, B.; Eibel, H. B-cell biology and development. J. Allergy Clin. Immunol. 2013, 131, 959–971. [Google Scholar] [CrossRef]
- Haas, K.M.; Poe, J.C.; Steeber, D.A.; Tedder, T.F. B-1a and B-1b cells exhibit distinct developmental requirements and have unique functional roles in innate and adaptive immunity to S. pneumoniae. Immunity 2005, 23, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Ghosn, E.E.; Russo, M.; Almeida, S.R. Nitric oxide-dependent killing of Cryptococcus neoformans by B-1-derived mononuclear phagocyte. J. Leukoc Biol. 2006, 80, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Noal, V.; Santos, S.; Ferreira, K.S.; Almeida, S.R. Infection with Paracoccidioides brasiliensis induces B-1 cell migration and activation of regulatory T cells. Microbes Infect. 2016, 18, 798–803. [Google Scholar] [CrossRef] [PubMed]
- Popi, A.F.; Godoy, L.C.; Xander, P.; Lopes, J.D.; Mariano, M. B-1 cells facilitate Paracoccidioides brasiliensis infection in mice via IL-10 secretion. Microbes Infect. 2008, 10, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Arcanjo, A.F.; Nunes, M.P.; Silva-Junior, E.B.; Leandro, M.; da Rocha, J.D.B.; Morrot, A.; Decote-Ricardo, D.; Freire-de-Lima, C.G. B-1 cells modulate the murine macrophage response to Leishmania major infection. World J. Biol. Chem. 2017, 8, 151–162. [Google Scholar] [CrossRef]
- Crane, D.D.; Griffin, A.J.; Wehrly, T.D.; Bosio, C.M. B1a cells enhance susceptibility to infection with virulent Francisella tularensis via modulation of NK/NKT cell responses. J. Immunol. 2013, 190, 2756–2766. [Google Scholar] [CrossRef] [Green Version]
- Firmino-Cruz, L.; Decote-Ricardo, D.; Gomes, D.C.O.; Morrot, A.; Freire-de-Lima, C.G.; de Matos Guedes, H.L. How to B(e)-1 Important Cell During Leishmania Infection. Front. Cell Infect. Microbiol. 2019, 9, 424. [Google Scholar] [CrossRef] [Green Version]
- Marquis, G.; Montplaisir, S.; Pelletier, M.; Mousseau, S.; Auger, P. Genetic resistance to murine cryptococcosis: Increased susceptibility in the CBA/N XID mutant strain of mice. Infect. Immun. 1985, 47, 282–287. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | Cryptococcus neoformans | Cryptococcus gattii | References |
|---|---|---|---|
| Geographic distribution | Found globally in bird feces, trees and soil. | Found in tropical and subtropical areas closely to pine and eucalyptus trees *. | [2,5] |
| Infection | Mostly immunocompromised hosts; secondary infection; incubation period around 110 months. | Mostly immunocompetent hosts; primary infection; incubation period from 6 to 7 months. | [3,6,182,183] |
| Serotypes | A, D and AD | B and C. | [17,18] |
| Genotypes | (A) VNI and VNII; (D) VNIV; (AD) VNIII; VNB. | (B) VGI and VGII; (C) VGIII and VGIV. | [17,19,20] |
| Cryptococcosis globally | Corresponds around 80% of infections. | Corresponds < 20% of infections. | [3] |
| Culture medium growth | Only uses urea as a nitrogen source, does not grow well in the presence of canavanine. | Can use glycine as a nitrogen source, grows well in the presence of canavanine. | [25,26] |
| Cell wall | H99 strain presents lower chitosan composition; less homogenous melanin distribution. | R265 strain presents higher chitosan composition; more homogenous melanin distribution. | [67] |
| Capsule composition, mannoprotein components, and structural difference | Proportion of (xylose:mannose:glucuronic acid) by serotypes: (D) 1:3:1 and (A) 2:3:1; mannoproteins: 46 predicted; Cig1; MP98; H99 strain GXM presents more O-acetylations. | Proportion of (xylose:mannose:glucuronic acid) by serotypes: (B) 3:3:1 and (C) 4:3:1; mannoproteins: 36 predicted; Krp1; JP02 strain GXM presents less O-acetylations. | [68,108,109,120,123,124] |
| Cryptococcus dissemination | More likely to disseminate to the CNS **; less severe disease than C. gattii. | More likely to present as pneumonia; a more severe disease. | [171] |
| Immune and pharmacological resistance | Cannot fully prevent phagolysosomal maturation; susceptibility to fluconazole. | R265 strain can highly prevent phagolysosomal maturation in dendritic cells leading to immune paralysis; adaptive heteroresistance to fluconazole and best treatment combining also amphotericin B and flucytocin. | [212,219] |
| Immune response | During phagocytosis macrophage IFN-γ production is more potent when compared to C. gattii; less severe cryptococcosis in the absence of autophagy | Reduced capacity for inducing IL-12 in vitro when compared to C. neoformans; R265 strain presents a lower production of TNF-a, IL-1b and IL-6; greater resistance to neutrophil oxidative stress than C. neoformans | [237,238,239,241] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diniz-Lima, I.; Fonseca, L.M.d.; Silva-Junior, E.B.d.; Guimarães-de-Oliveira, J.C.; Freire-de-Lima, L.; Nascimento, D.O.; Morrot, A.; Previato, J.O.; Mendonça-Previato, L.; Decote-Ricardo, D.; et al. Cryptococcus: History, Epidemiology and Immune Evasion. Appl. Sci. 2022, 12, 7086. https://doi.org/10.3390/app12147086
Diniz-Lima I, Fonseca LMd, Silva-Junior EBd, Guimarães-de-Oliveira JC, Freire-de-Lima L, Nascimento DO, Morrot A, Previato JO, Mendonça-Previato L, Decote-Ricardo D, et al. Cryptococcus: History, Epidemiology and Immune Evasion. Applied Sciences. 2022; 12(14):7086. https://doi.org/10.3390/app12147086
Chicago/Turabian StyleDiniz-Lima, Israel, Leonardo Marques da Fonseca, Elias Barbosa da Silva-Junior, Joyce Cristina Guimarães-de-Oliveira, Leonardo Freire-de-Lima, Danielle Oliveira Nascimento, Alexandre Morrot, Jose Osvaldo Previato, Lucia Mendonça-Previato, Debora Decote-Ricardo, and et al. 2022. "Cryptococcus: History, Epidemiology and Immune Evasion" Applied Sciences 12, no. 14: 7086. https://doi.org/10.3390/app12147086