
Determination of Volatile Compounds in Blossoms of Rosa spinosissima, Rosa pendulina, Rosa gallica, and Their Cultivars

Department of Agronomy, Biotechnical Faculty, University of Ljubljana, Jamnikarjeva 101, 1000 Ljubljana, Slovenia
*
Author to whom correspondence should be addressed.
Agriculture 2024, 14(2), 253; https://doi.org/10.3390/agriculture14020253
Submission received: 18 December 2023
/
Revised: 24 January 2024
/
Accepted: 3 February 2024
/
Published: 4 February 2024
(This article belongs to the Special Issue Phenolic Compounds, Terpenoids and N-containing Substances Found in Fruits and Vegetables)
Abstract
:The aim of our study was to determine how the content and composition of volatile organic compounds (VOCs) change with crossbreeding. We included three autochthonous Slovenian species of roses (R. pendulina, R. spinosissima, and R. gallica) and several cultivars derived from them in the research. The results were compared with the content of the reference plant R. damascena, which is known worldwide for its rich VOC profile and is used in the perfume industry. We found that the total VOC content ranged from 2.579 µg/g FW (‘Fruhlingsmorgen’) to 168.24 µg/g FW (‘Violacea’). According to the content of total VOCs, the variety ‘Charles de Mills’ (43.12 µg/g DM) was the most similar to the variety R. damascena (44.55 µg/g DM). We can conclude that the flowers of the roses we included in the study are a rich source of VOCs. VOC content in hybrids varies depending on the purpose and goals of breeders. We also found that flowering time had no effect on VOC content and composition. With this study, we not only carefully examined the VOC profile of roses but also clearly showed which compounds were present in individual taxa. In this way, it is possible to select a suitable variety or cultivar depending on the desired use, for example, for flavoring food or for a floral fragrance in the perfume industry.
1. Introduction
Roses (Rosa spp.) have been popular throughout the world since ancient times and are considered the most widely cultivated ornamental plants and the queens of fragrant flowers [1,2]. Although they were originally popular for their fragrance and medicinal properties, they were later cultivated for their ornamental value. The best-known species at that time was the Damask rose (Rosa damascena Mill.), which has small flowers with an extremely strong fragrance. Even today, it is considered the rose with the strongest fragrance and the highest essential oil content [3,4]. It is a hybrid of R. gallica and R. phoenicea, but its origin is still not well understood and is disputed worldwide [5]. However, there are many old rose species that are very rich in VOCs and have great potential for use, but unfortunately, they are poorly studied [6].
The pleasant fragrance of roses is due to the essential oils present. Their content is influenced by various factors, such as the cultivar and the climatic conditions. The profile of the fragrance depends on the soil, heat, humidity, time of the day, etc. Most rose perfumes used in commerce are obtained from R. damascena and R. centifolia [7]. The VOCs are released from the above-ground parts of the plant to the atmosphere and from the underground parts to the soil. Their main functions are to protect plants from herbivores and pathogens, attract pollinators, transfer seeds, and communicate between plants. They are used as flavoring agents and preservatives and in alternative medicine [7]. More than 400 fragrant VOCs have been identified in rose flowers, which are classified into different chemical compounds, including hydrocarbons, alcohols, esters, aromatic ethers, and other similar compounds such as geranial and nonanal aldehydes, rose oxide, and norisoprenes such as β-ionone [8]. VOC esters such as geranyl acetate and 2-phenylethyl acetate contribute significantly to rose aroma [8]. It should be emphasized that many VOCs are odorless, which means that their content cannot elicit an olfactory response in humans [9]. The proportion of the major constituents is an important factor that determines the quality of rose essential oil. The most common constituents of the R. damascena essential oil are β-citronellol, nerol, phenylethyl alcohol, geraniol, heptadecane, nonadecane, methyl eugenol, eugenol, and geranyl acetate, which account for 85 to 90% of the total rose oil VOCs. β-damascone, β-damascenone, and β-ionone also appear as less represented components that contribute significantly to the characteristic fragrance of Damask rose [5,10]. β-damascone and β-damascenone are used as markers for the determination of Damask rose essential oil [5,10]. Demir et al. [11] reported that the VOCs in rosehip are a mixture of 52 components, including alcohols, aldehydes, ketones, terpenoids, and esters. Aldehydes and alcohols are the predominant components of the VOC fractions of fruit and herbs. Shamspur and Mostafavi [2] analyzed the VOCs; henicosane, tricosane, (Z)-5-nonadecene, and methyl eugenol were listed as major compounds in R. damascena among all 76 compounds identified. In R. gallica, 49 compounds were determined, which accounted for 99.4% of all detected components. The most important of these were citronellol, henicosane, tricosane, pentacosane, and phenylethyl alcohol.
Roses are used not only for decorative purposes in parks and public and private gardens but also in perfumery, the cosmetics industry, and medicinal purposes due to the above-mentioned ingredients. They have anti-aging, anti-tyrosinase, and depression effects; they also have a positive effect on the intestine with antioxidant, anti-cancer, anti-inflammatory, antimicrobial, anti-epileptic, anti-migraine, anti-allergic, and other effects [12,13]. Abudayeh et al. [12] reported a study in which rose absolute and essential oils were found to be active against E. coli, P. aeruginosa, B. subtilis, and S. aureus. Flowers are also widely used in the food industry for making jams, syrup, rose water, vinegar, spices, yogurt additives, tea, etc. [14,15,16,17,18,19].
Rubtsova et al. [20] wrote that rose morphological characteristics and flowering time have changed significantly due to many years of cultivation and breeding and that further breeding should be focused even more on improving biological traits, such as winter, drought, disease, and pest resistance. Gaudin [21] also reports that throughout the history of rose breeding, since the 18th century, the goals of breeding have not changed drastically, only the ways to achieve the desired results. Datta [22] describes those roses, along with chrysanthemums, as the plants where the newest breeding methods have been applied. R. gallica contributed to cold resistance, R. chinensis provided repeated flowering, and R. foetida provided a yellow color. VOCs in rose flowers exhibit great diversity while providing a wealth of information that could be used to infer taxonomic relationships. Therefore, VOCs in flowers are also an important tool for species identification and could be useful in defining new species/cultivars.
There are some studies on the composition and content of VOCs in R. damascena and R. gallica, but almost nothing is known about the VOCs of R. spinosissima and R. pendulina. In this research, we describe the content and composition of VOCs of the three mentioned autochthonous Slovenian species and compare them with the reference plant R. damascena. In addition, we will find out how the composition and VOC content change among cultivars (‘Fruhlingsmorgen’, ‘Fruhlingsduft’, ‘Marigold’, ‘Fruhlingsgold’, ‘Karl Foerster’, ‘Mon Amie Claire’, ‘Bourgogne’, ‘Harstad’, ‘Mount Everest’, ‘Violacea’, ‘Splendens’, ‘Complicata’ and ‘Charles de Mills’) derived from autochthonous roses. We will also examine whether rose cultivars with different flowering times differ from each other in terms of composition and VOC content in flowers. Finally, we determine if the roses we are studying are a sufficiently rich source of VOCs that could potentially be used in the pharmaceutical, food, and cosmetic industries.
2. Materials and Methods
2.1. Plant Material
Blossoms (Figure 1 and Figure 2) of Rosa damascena, Rosa pendulina, Rosa spinosissima, Rosa gallica, and their derived cultivars, ‘Fruhlingsmorgen’ (deriving from R. spinosissima), ‘Fruhlingsduft’ (deriving from R. spinosissima), ‘Maigold’ (deriving from R. spinosissima), ‘Fruhlingsgold’ (deriving from R. spinosissima), ‘Karl Foerster’ (deriving from R. spinosissima), ‘Mon Amie Claire’ (deriving from R. spinosissima), ‘Bourgogne’ (deriving from R. pendulina), ‘Harstad’ (deriving from R. pendulina), ‘Mount Everest’ (deriving from R. pendulina), ‘Violacea’ (deriving from R. gallica), ‘Splendens’ (deriving from R. gallica), ‘Complicata’ (deriving from R. gallica), and ‘Charles de Mills’ (deriving from R. gallica), were collected in Arboretum Volčji potok (Slovenia) in the BBCH65 phenophase [23]. All plants from which the blossoms were collected were growing under the same climatic conditions and were maintained, pruned, fertilized, and irrigated in the same way all year round.
Blossoms of R. spinosissima, ‘Fruhlingsmorgen’, ‘Fruhlingsduft’, ‘Maigold’, ‘Fruhlingsgold’, ‘Karl Foerster’, ‘Mon Amie Claire’, ‘Bourgogne’, ‘Harstad’, R. pendulina and R. damascena were collected in the second half of May 2023. In the first half of June 2023, ‘Mount Everest’, R. gallica, ‘Violacea’, ‘Splendens’, ‘Complicata’, and ‘Charles de Mills’ were harvested. The collected material was placed on ice and transferred to the laboratory. The material was immediately ground with liquid nitrogen and stored at −80 °C until further analyses.
Table 1 shows the species and cultivars we included in the study, their origin, year of origin, and breeding company. Cultivars crossed before 1876 are classified as old cultivars, and cultivars crossed later belong to modern cultivars. The group of old cultivars includes: ‘Violacea’, ‘Splendens’, ‘Charles de Mills’, ‘Complicata’, and ‘Harstad’.
2.2. Identification and Semi-Quantification of Volatile Compounds
The profile of VOCs was determined by gas chromatography analysis (HS-GC-MS) following that of [24]. Five flowers were selected for each species/cultivar. Fresh flowers were frozen with liquid nitrogen immediately after harvest and ground into powder using a mortar and pestle. Three technical replicates were made for each species or cultivar. Then, 0.5 g of powder along with 10 μL of an internal standard (IS: 3-nonanone, 0.405 mg/mL in acetonitrile) was placed in 20 mL glass vials, sealed with a screw cap with PTFE silicone septum, and transferred to a Shimadzu AOC-20s autosampler (Shimadzu, Kyoto, Japan), where it was incubated at 50 °C for 10 min with constant shaking at 250 rpm. An injection of 1000 μL of the headspace portion was performed in splitless mode for 0.4 min into the injection port at 250 °C with an injection rate of 25 mL/min. A Shimadzu GC-MS QP2020 gas chromatograph connected to a single quadrupole MS with an EI detector was used, with a ZB Wax PLUS capillary column (30 m × 0.25 mm and 0.5 μm film thickness). The carrier gas was helium, with a flow rate of 1 mL/min. The temperature program was set as follows: first, the temperature was held at 45 °C for 3 min, then increased to 130 °C at a rate of 3 °C/min, then increased again to 220 °C at 25 °C/min, and finally held at 220 °C for 5 min. The interface and MS ion source temperature were set to 240 °C, the scan rate to 2.0 scan/s, the ionization energy to 70 eV, and the mass scan range to 35–500 m/z. The VOC of the taxons were identified based on their retention indices (RIs) and commercial spectral libraries (NIST 11 and FFNSC 4) and semi-quantified based on the peak areas of each compound and the internal standard, as well as the internal standard and taxons weight.
2.3. Statistical Analyses
Results were analyzed using the statistical program R Commander i386 4.1.2. One-way analysis of variance (ANOVA) and Tukey’s test were used to compare the results of each variable between taxons. Results are expressed as the compound concentration per the 3-nonanone standard and presented as mean with standard error (SE). When p-values were lower than 0.05, differences between treatments were considered statistically significant. A cluster dendrogram was obtained using the UPGMA method based on the total content of VOCs.
3. Results
Of all the taxons studied, R. pendulina had the highest number of individual VOCs (52), followed by R. damascena (50) (Figure 3). The lowest number of VOCs was determined in ‘Mon amie Claire’ (R. spinosissima group) (19).
The proportion of monoterpenes was highest for ‘Mount Everest’ (R. pendulina group) (Figure 3). Sesquiterpenes were highest in R. pendulina (38%) and lowest in the cultivar ‘Violacea’ (3%) (R. gallica group). As for the highest value of aldehydes, ‘Fruhlingsmorgen’ (R. spinosissima group) stood out (23%), while the lowest content was determined in R. pendulina (10%). The content of alcohols varied from 16% (‘Complicata’, R. gallica group) to 42% (‘Mon amie Claire’, R. spinosissima group). The proportion of identified ketones in the taxons varied between 0% and 3% and differentially to 14% in ‘Karl Foerster’ (R. spinosissima group). The percentage of hydrocarbons ranged from 15% (R. pendulina) to 36% (‘Fruhlingsduft’, R. spinosissima group). Only in nine analyzed taxons were phenylpropenes, determined in all groups except R. pendulina; the percentage of other compounds ranged from 2 to 8%.
Examining all the species and cultivars included in the study (Table 2), it can be seen that the cultivar ‘Violacea’, which was derived from R. gallica, had the highest content of VOCs overall. The lowest content was exhibited by the cultivar ‘Fruhlingsmorgen’ (R. spinosissima group).
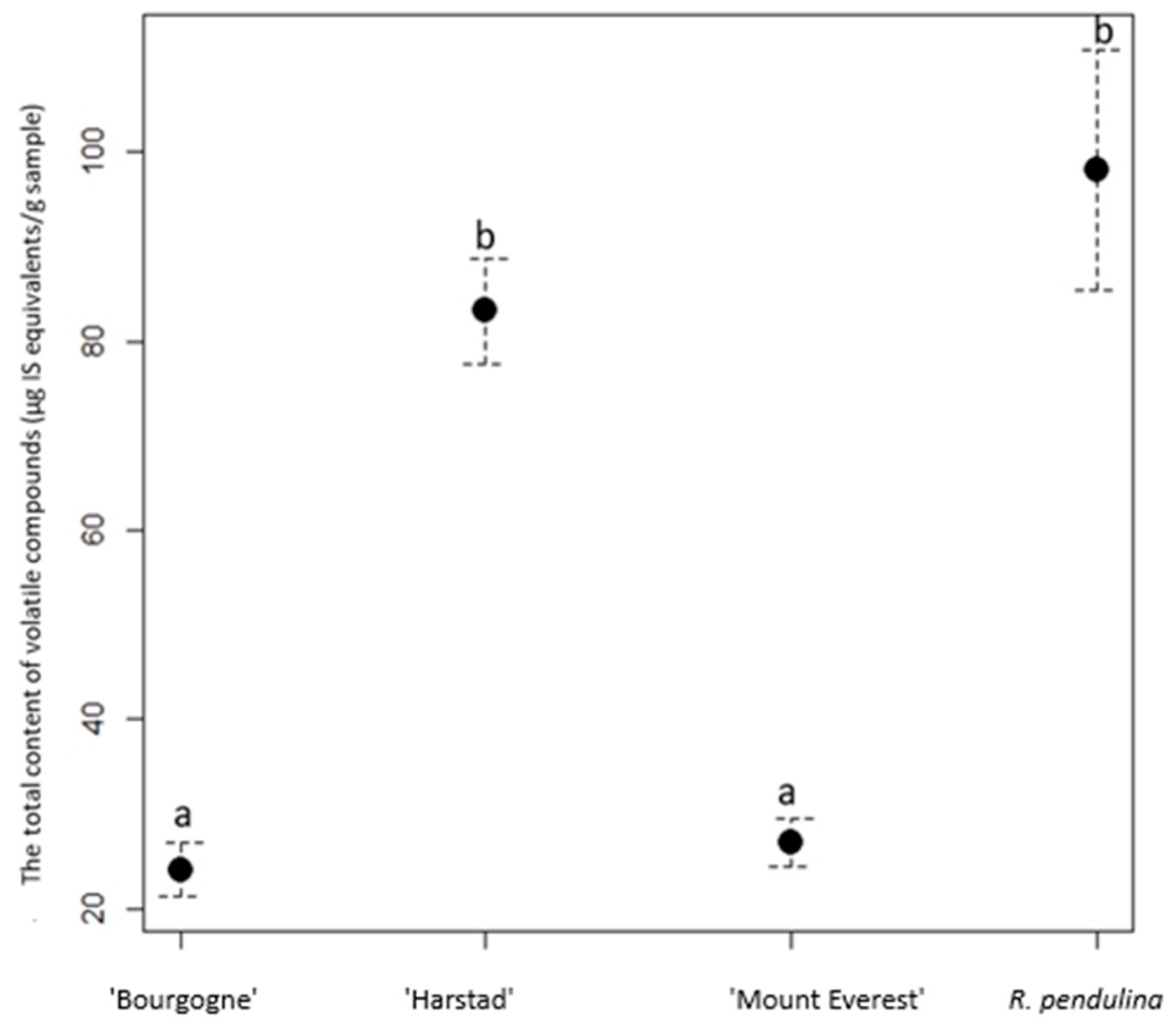
Table 2 and Table S1 show that there is no statistically significant difference between the content of monoterpenes and sesquiterpenes between the cultivars ‘Bourgogne’ and ‘Mount Everest’. We detect statistically significant differences in the content of aldehydes and alcohols between the studied species/cultivars. R. pendulina was statistically different from all cultivars except ‘Harstad’. Regarding the total content of VOCs, we found that R. pendulina was not statistically different from ‘Harstad’ and that there was no statistical difference between ‘Bourgogne’ and ‘Mount Everest’ (Figure 4).
Table 2 and Table S2 show the content and composition of the VOCs fraction of R. spinosissima group. We found that there was no statistically significant difference between species/cultivars in the content of monoterpenes and sesquiterpenes. R. spinosissima and ‘Fruhlingsmorgen’ did not significantly differ from each other in terms of total aldehyde content. We found that both ‘Maigold’ and ‘Fruhlingsgold’ differed significantly from the rest of the species/cultivars within the R. spinosissima group in terms of aldehyde content. ‘Fruhlingsduft’ was significantly different from R. spinosissima, ‘Fruhlingsmorgen’, and ‘Karl Foerster’ in total alcohol content. ‘Fruhlingsduft’ was also statistically different from all other species/cultivars in terms of the total content of ketones and hydrocarbons within the studied group of R. spinosissima. Statistically significant differences between certain species/cultivars were also observed in the total content of phenylpropenes. According to the total VOC content (Figure 5), we saw that there were significant differences between some of the studied species/cultivars.
Table 2 and Table S3 show the content and composition of the VOC fraction of the R. gallica group (Figure 6). We found that ‘Violacea’ statistically differed from all other samples in terms of total monoterpene content. We did not detect a statistically significant difference in the total content of sesquiterpenes. ‘Charles de Mills’ was not statistically different from R. gallica and ‘Violacea’ in total aldehyde content. We did not detect statistically significant differences in the total content of ketones and phenylpropenes either. However, we found that ‘Violacea’ was statistically significantly different from the other varieties in terms of hydrocarbon content and also in terms of total VOC content.
Table S4 shows the content and composition of the VOCs of R. damascena. Of all the groups of compounds analyzed, the monoterpenes were the most abundant, followed by aldehydes, alcohols, hydrocarbons, sesquiterpenes, ketones, phenylpropenes, and finally, the remaining compounds. Of the phenylpropenes, only methyl eugenol was determined, and among the compounds classified as other compounds, only acetic acid was found.
Figure 7 shows the cluster dendrogram for the total VOC content for all 17 species. It can be seen from the figure that cultivar ‘Violacea’ (R. gallica group) was very different from the other analyzed cultivars in terms of the total content of VOC compounds. The other cultivars were divided into two larger groups based on the VOCs. One group included R. pendulina, ‘Harstad’ (R. pendulina group), and ‘Maigold’ (R. spinosissima group). The second group was further divided into a group that included ‘Fruhlingsgold’ (R. spinosissima group), R. gallica, ‘Charles de Mills’ (R. gallica group), and R. damascena, and to the group then further divided into two groups. The first of the two groups included R. spinosissima, ‘Splendens’ (R. gallica group), ‘Bourgogne’ (R. pendulina group), ‘Fruhlingsduft’ (R. spinosissima group), ‘Mount Everest’ (R. pendulina), and ‘Complicata’ (R. gallica). In the second case, they were ‘Fruhlingsmorgen’ (R. spinosissima group), ‘Karl Foerster (R. spinosissima group), and ‘Mon amie Claire’ (R. spinosissima group).
4. Discussion
With the investigation, we determined the content and composition of the VOCs profile of three native Slovenian rosehips, their derived cultivars, and the reference plant R. damascena. So far, there are not many studies investigating the content of VOCs in the flowers of the above-mentioned original genotypes and their cultivars.
When we studied the R. pendulina group, we found that the content of monoterpenes in the flowers of cultivars in the group was lower compared to the flowers of the R. penduline species. Only the cultivar ‘Harstad’, which belongs to the old cultivars, had a similar content of monoterpenes in its flowers as R. pendulina. The lower VOC content in the modern cultivars is probably the consequence of the breeding objective of their breeding house, Interplant. The latter is known as a world leader in the development of spray roses and is among the first companies to introduce this product to the international rose market. One of its breeding goals is to breed compact roses in different colors that are resistant to diseases and pests. However, it does not focus on crossing roses whose flowers are rich in VOCs [25].
The content of R. spinosissima and its cultivars was quite different compared to R. pendulina. The total content of all VOCs determined in the flowers of R. spinosissima and its modern cultivars was lowest for the cultivar ‘Fruhlingsmorgen’ and highest for ‘Maigold’. The main goal of breeding in their breeding house, Kordes, is the production of resistant cultivars, not only to pests but also to cold. In addition, the breeding of cultivars with multiple flowering and rich fragrances is another goal of their strategy [26]. The latter was probably also observed in the cultivar ‘Maigold’, which had a higher content of VOCs in the flowers.
The total content of VOCs in the cultivar group of R. gallica showed a relatively wide range between the cultivars, with the lowest content in the flowers of the cultivar ‘Splendens’ and the highest content in the flowers of the cultivar ‘Violacea’. The content of monoterpenes in this group increased with crossing, which was probably due to the fact that mainly older cultivars were included in the study.
The inheritance of fragrance depends on several genes, and even when two fragrant roses are crossed, their crosses are not necessarily similar due to random segregation and unfavorable recombination of the genes involved. The genes responsible for fragrance are consistently associated with some undesirable traits that produce an unpleasant scent [27]. The rose crosses are performed solely for the purpose of production in the perfume industry. They concluded by stating that in order to develop roses, it was necessary to breed many new genetic variations by using conventional breeding techniques of population segregation because roses are very heterozygous and polyploid. Appropriate breeding techniques and skillful selection of hybridization parents would help to achieve the desired results by direct breeding. All this information confirms our findings that breeding objectives often go in directions quite different from the VOCs of rosehip flowers. In addition, Ryu et al. [28] analyzed 12 mutant genotypes of roses obtained by gamma irradiation and their six original cultivars. They identified 77 VOCs and concluded that a generation of new rose genotypes by irradiation increases the content of various bioactive phytochemicals, which may be new sources for future use in food, cosmetics, horticulture, and aromatherapy. It is also interesting to note that rose flower fragrance is a highly heritable quantitative trait [29]. When a scented cultivar is crossed with a non-scented rose cultivar, the progeny differ in the intensity of their aromas. Walls et al. [30] state that the content of the VOCs of different rose cultivars is critical for the sustainable development of the rose industry.
When comparing the roses analyzed in our study with the reference plant R. damascena, we found that the cultivar ‘Charles de Mills’, which is derived from R. gallica, was the most similar to the mentioned cultivar. The total content of VOCs in R. damascena was 44.55 µg IS equivalents/g sample and in ‘Charles de Mills’ 43.12 µg IS equivalents/g sample. When we examined the compounds, we found that α-pinene was the most abundant monoterpene in both taxons; the content of alcohol and linalool was 0.07 µg IS equivalents/g sample in R. damascena, and 0.09 µg IS equivalents/g sample in ‘Charles de Mills’. Phenylethyl alcohol contents were also remarkably similar, with R. damascena containing 3.08 µg IS equivalents/g sample and ‘Charles de Mills’ containing 3.10 µg IS equivalents/g sample. The difference was found in the content of citronellol, which was present only in R. damascena (1.53 µg IS equivalents/g sample). ‘Violacea’ as the cultivar with the highest content of VOC compounds (168.24 µg IS equivalents/g sample), differed from R. damascena by the extremely high content of α-pinene (107.77 µg IS equivalents/g sample), which was only 9.01 µg IS equivalents/g sample in the flowers of R. damascena. Özdemir and Budak [19] studied the VOC profile of R. damascena in rose water and vinegar. Rose oxide, acetic acid, 2-phenethanol, octanoic acid, ethyl phenyl acetate, isoamyl acetate, phenylacetic acid, acetoin, linalool, ethyl acetate, and β-citronellol were listed as the major compounds. They concluded that this species was suitable for industrial cultivation. In our case, the main VOCs of the flowers of R. damascena were α-pinene, β-myrcene, hexanal, 2-hexanal, and phenylethyl alcohol. Abudayeh et al. [12] studied the composition and VOC content of R. damascena growing in Jordan. They mentioned henicosane as the main constituent in the petals, 0.12 µg/g FW. In our studies, the content of the mentioned compound in R. pendulina and its cultivars varied from 0.06 (R. pendulina) to 0.11 µg/g FW (‘Mount Everest’). The levels in the R. spinosissima group were even higher than those reported by Abudayeh et al. [12].
The cultivar ‘Mount Everest’ (group R. spinosissima) had the highest proportion of monoterpenes (28%). The highest total content was found in R. pendulina. Guterman et al. [31] reported that the floral scent of roses consists of volatile monoterpenes, isoprenoid alcohols, and their oxidized and glycosylated (non-volatile) derivatives. Plants form monoterpenes from geranyl diphosphate via the methylerythritol phosphate pathway localized in the plastid [32,33]. In addition to the methylerythritol phosphate pathway, rose flowers also have a unique cytosolic NUDIX hydrolase pathway that exclusively forms geraniol, an important acyclic monoterpene in hybrid roses [34,35]. Dani et al. [36], who studied Asian and American rose cultivars, wrote that cyclic monoterpenes in American roses (mainly α-pinene, β-pinene, limonene, cineole, and p-cymene) are low (comparable to linalool) and do not change during flower life. However, Asian roses showed a significant increase in cyclic monoterpenes in aging petals. In addition, flower color was found to affect monoterpene levels, with intense pink flowers having higher monoterpene levels compared to yellow flowers. Consistent with their results, we found that no monoterpenes were present in the cultivars ‘Mon amie Claire’, ‘Karl Foerster’, and ‘Fruhlingsduft’, with light pink, yellow with light pink spots, and white flowers, respectively, while the cultivar ‘Mount Everest’ had intense pink flowers and also the highest proportion of monoterpenes. Sparinska and Rostoks [37], who studied R. rugosa hybrids from Latvia, also mention phenylethyl alcohol, citronellol, nerol, and cis-geraniol as the main compounds. The VOCs found in the highest amounts in Latvian R. rugosa hybrids were phenylethyl alcohol (up to 79.9%), citronellol (up to 57%), and nerol (up to 39%). Karami et al. [38] wrote that the floral fragrance of Iranian R. damascena flowers contains 47% of β-citronellol. The content of citronellol in the cultivar ‘Raita’ (43% in fully opened flowers) was similar to our results in R. damascena, while the introduced cultivars ‘Plena’ and ‘Mrs. Dagmar Hastrup’ contained even higher content, up to 57% of this major constituent of rose oil. In the species R. pendulina, ‘Harstad’, ‘Violacea’ and ‘Complicata’ we identified α-pinene as the main constituent, which is used as a food additive and may be added directly to food for human consumption as a flavoring agent. It is characterized by intense woody and piney scents with camphor and turpentine notes. It also has herbaceous, spicy, and slightly tropical undertones [39]. Lijun et al. [9] studied the VOCs in extracts of R. odorata and R. chinensis and found that α-pinene was present among the compounds in lower amounts than the values we found. Kim et al. [39] reported that α-pinene is characterized by antimicrobial, apoptotic, antimetastatic, and antibiotic properties and is the main constituent of eucalyptus oils and oils of aromatic plants such as rosemary, mint, eucalyptus, hemp, and basil.
The highest proportion of sesquiterpenes was found in R. pendulina. Mai et al. [40] reported that sesquiterpenes are the largest class of terpene compounds, and many of them have exceptional biological activities due to their complex structure and are, therefore, useful in the flavor, pharmaceutical, and biofuel industries. They are believed to play an important role in determining the odor profile and have a significant impact on aroma, which is critical for marketing and quality [41]. It was also noted that floral scents are also the result of the accumulation of terpenoids, which play an important role in the interaction between pollinators, herbivores, and pathogens in their ecosystem. For example, β-ocimene is a universal terpenoid involved in pollinator attraction, and trans-β-ocimene is emitted most strongly by plants at night, coinciding with flower opening and pollinator activity [41]. In our study, β-ocimene was present only in ‘Fruhlingsmorgen’ (R. spinosissima group) and in the R. gallica group, except ‘Splendens’.
Aldehydes and ketones are known for their sweet and sometimes pungent odor. Benzaldehyde provided a strong almond odor and was present in our taxons at the highest content in R. pendulina. The highest proportion of aldehydes was found in ‘Fruhlingsgold’ (22%, R. spinosissima group), and the highest percentage of ketones in ‘Karl Foerster’ (14%, R. spinosissima group). Hexenals were the predominant VOCs in ‘Bourgogne’, ‘Mount Everest’, R. spinosissima, ‘Fruhlingsmorgen’, ‘Maigold’, ‘Fruhlingsgold’, R. gallica, ‘Charles de Mills’, ‘Splendens’, and in R. damascena. Hexanal is used for flavoring and perfuming. It has green, woody, herbaceous, apple, grass, citrus, and orange aromas with a fresh, lingering aftertaste [40,41,42,43]. Bergoulugunux et al. [43] studied VOCs in flowers of the modern cultivar Rosa × hybrid ‘Papa Meilland’ in different parts of the flower: petals, stamens, and sepals. The total VOC content was highest in petioles and was 443.1 µg/g FW; in stamens, it was 380.6 µg/g FW, and in sepals, it was 81.0 µg/g FW. When we compared the results obtained with the results of our research, we found that the values they obtained were higher than our results; trans-2-hexenal accounted for 58.4% of the total amount of VOCs.
Alcohols also contribute to the fruity aroma [44,45]. In the essential oil of R. damascena, the most abundant alcohols are phenethyl alcohol, citronellol, eugenol, methyleugenol, and geraniol. Demir et al. [11] also reported that in Rosa species (R. dumalis, R. canina, R. dumalis subsp. boissieri, R. gallica, and R. hirtissima), the most abundant compounds are alcohols in addition to aldehydes, monoterpenes, and sesquiterpenes. The main alcohols reported by Sparinska and Rostoks [37] in the taxons of six R. rugosa hybrids were phenylethyl alcohol, β-citronellol, nerol, and geraniol. In our taxons, the highest proportion of alcohols was present in ‘Mon amie Claire’ (42%), with the major alcohols being citronellol, phenylethyl alcohol, and geraniol. However, the highest content of alcohols was found in ‘Fruhlingsduft’ (9.74 µg/g FW), where the main alcohols were 1-hexanol, phenylethyl alcohol, geraniol and citronellol, in accordance with Sparinska and Rostoks [37]. Both mentioned cultivars rich in alcohol were derived from R. spinosissima. Bergougnoux et al. [43] reported that monoterpene alcohols such as geraniol, citronellol, and nerol are most abundant in the petals and stamens and are also responsible for the characteristic fragrance of roses, containing significant amounts of aromatic compounds such as 2-phenylethanol (rose notes) and 3,5-dioxytoluene (tea notes). 2-phenylethanol (phenylethyl alcohol) was also present in our taxons. We note that the cultivar ‘Fruhlingsgold’ exhibited the most pronounced rose note. 3,5-dimethoxytoluene was only present in R. gallica in the cultivars ‘Fruhlingsduft’ and ‘Fruhlingsgold’. In addition, Abudayeh et al. [12] also reported that linalool was not present in the petals. In our taxons, it was present in R. spinosissima and some of its cultivars, as well as in R. damascena (0.07 µg/g FW).
Hydrocarbons are commonly used in industry as insecticides and pharmaceuticals but not as flavoring agents. They include the esters, which have a generally pleasant odor and are also responsible for the fruity and floral aroma [39,42,46]. The highest proportion of hydrocarbons was found in ‘Fruhlingsmorgen’, where n-nonadecane dominated and accounted for 56.8% of all hydrocarbons in the taxon. The highest amount of hydrocarbons was determined in ‘Violacea’, where n-nonadecane also dominated and accounted for 39.6% of all hydrocarbons. The hydrocarbon n-nonadecane was also predominant in the cultivars ‘Fruhlingsduft’, ‘Karl Foerster’, and ‘Mon amie Claire’. Mostafa et al. [46] also reported a high content of 3,5-domethoxytoluene, but in our case, the values were considerably lower.
Among the phenylpropenes, we determined the presence of methyl eugenol, which is characterized by a pungent odor reminiscent of cloves. Synthetic methyl eugenol is widely used as a flavoring agent in many types of processed foods, soft drinks, and sauces, in perfumery, and as an essential oil in aromatherapy [47]. Its content in our taxons ranged from 0.04 µg/kg FW (‘Harstad’, R. pendulina group) to 0.28 µg/kg FW (R. spinosissima). Rusanov et al. [48] reported that methyl eugenol is a carcinogenic compound present in many essential oils, including rose oil distilled from the flowers of R. damascena. They found that the level of methyl eugenol decreased fivefold in rose oil distilled from flower buds before the petals opened. We harvested the flowers at full maturity, and it was suggested that this was the reason for the increase in methyl eugenol content in some species/cultivars. Mileva et al. [49] wrote that the low content of methyl eugenol and the high content of geraniol ensure safe use, with proven biological effects of R. alba essential oil, and that both R. gallica and R. centifolia are generally characterized by a low content of methyl eugenol. In our taxons, R. gallica and its modern cultivars did not contain methyl eugenol, except for the cultivar ‘Complicata’.
According to the results obtained by grouping all the species/cultivars we included with the research, it can be concluded that not all varieties that come from the same breeding companies have a similar VOC content, and the dendrogram does not group them together. We note, however, that only ‘Fruhlingsmorgen’, ‘Karl Foerster’, and ‘Mon amie Claire’, modern cultivars, come from the same breeding company and are grouped together in the dendrogram. R. pendulina and its old cultivar ‘Harstad’ are grouped together, without the other two cultivars derived from R. pendulina. The dendrogram also groups together ‘Mount Everest’ and ‘Complicata’, which are of completely different origins. The last comes from R. gallica, whose breeding company is unknown, and the year of origin is before 1873. While ‘Mount Everest’ is a modern cultivar created in 1956 by the breeding company Interplant. We also found that R. pendulina, ‘Harstad’, and ‘Maigold’, which are grouped together, differed not only in terms of the original species but also in terms of the quality of the flowers. The last of the listed roses has yellow-orange flowers, while the other two are distinctly pink-purple, which is also seen in the group that includes ‘Fruhlingsgold’ and also differs from the others in its yellow flowers. The fact that flower color does not affect the grouping of studied species/cultivars is also confirmed by other classifications of roses and their flower color.
Staudt and Bertin [50] found that VOC content was also influenced by the time of flowering. Roses blooming in April, i.e., cool spring, generally have higher levels of some components (e.g., geraniol) than species blooming in July, i.e., hot summer. However, the time period of our study was too broad (late May and early June) to observe any influence of flowering time on VOCs.
5. Conclusions
This research determined the VOC profile of three native Slovenian rosehips (R. pendulina, R. spinosissima, and R. gallica) and the cultivars derived from them and compared the results with the reference plant R. damascena. We found that the VOC content changes with crossbreeding, depending on the variety and the goal that the breeders are pursuing with the crossbreeding. Recently, their goals have been focused mainly on the resistance of plants against diseases and pests rather than rich and fragrant flowers and repeated flowering. We can conclude that the flowers of the roses we included in the study are a rich source of VOCs, quite comparable to roses such as R. damascena, widely known for their fragrance. With this research, we not only carefully examined the volatile profile of roses but also clearly showed which compounds were present in individual samples. In this way, it is possible to choose a suitable variety depending on the desired application goals, for example, for flavoring food or for a floral scent in the perfume industry. For the further use of species and varieties, more research would be needed to determine exactly for which use certain species or varieties are suitable. In addition, we also found that the flowering time did not affect the composition and content of VOCs. No effect was observed between flowers harvested at the end of May and those harvested at the beginning of June.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/agriculture14020253/s1. Table S1: Content and composition of the volatile phase (µg IS equivalents/g sample) in R. pendulina and its derived cultivars. Table S2: Content and composition of the volatile phase (µg IS equivalents/g sample) in R. spinosissima and its derived cultivars. Table S3: Content and composition of the volatile phase (µg IS equivalents/g sample) in R. gallica and its derived cultivars. Table S4: The content and composition of VOCs (µg IS equivalents/g sample) in R. damascena.
Author Contributions
Conceptualization, N.K., G.O. and M.C.G.; methodology, N.K., G.O. and M.C.G.; formal analysis, N.K., G.O. and M.C.G.; investigation, N.K.; data curation, N.K. and M.C.G.; writing—original draft preparation, N.K. and G.O.; writing—review and editing, N.K. and M.H.; G.O. and M.C.G.; visualization, N.K., G.O. and M.C.G.; supervision, M.H., G.O. and M.C.G.; funding acquisition, M.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work is part of the Horticulture P4-0013-0481 program supported by the Slovenian Research and Innovation Agency (ARIS).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data will be made available on request.
Acknowledgments
The authors thank the whole staff of the Arboretum Volčji Potok, especially Matjaž Mastnak, for making the plant material available.
Conflicts of Interest
The authors declare no conflicts of interest. The funder had no role in the study’s design, in the collection, analysis, or interpretation of the data, in the writing of the manuscript, or in the decision to publish the results.
References
- Raymond, O.; Gouzy, J.; Just, J.; Badouin, H.; Verdenaud, M.; Lemainque, A.; Vergne, P.; Moja, S.; Choisne, N.; Pont, C.; et al. The Rosa genome provides new insights into the domestication of modern roses. Nat. Genet. 2018, 50, 772–777. [Google Scholar] [CrossRef]
- Shamspur, T.; Mostafavi, A. Chemical composition of the volatile oil of Rosa kazanlik and Rosa gallica from Kerman province in Iran. J. Essent. Oil-Bear. Plants 2010, 13, 78–84. [Google Scholar] [CrossRef]
- Cottini, P. Vrtnice, sorte in način gojenja (izbira, sajenje, nega, obrezovanje). Rože in vrt 2003, 1–34. [Google Scholar]
- Gudin, S. Rose: Genetics and Breeding. Plant Breed. Rev. 2000, 17, 319–334. [Google Scholar] [CrossRef]
- Pal, P.K. Evaluation, genetic diversity, recent development of distillation method, challenges and opportunities of Rosa damascena: A review. J. Essent. Oil-Bear. Plants 2013, 16, 1–10. [Google Scholar] [CrossRef]
- Antonelli, A.; Fabbri, C.; Giorgioni, M.E.; Bazzocchi, R. Characterization of 24 old garden roses from their volatile compositions. J. Agric. Food Chem. 1997, 45, 4435–4439. [Google Scholar] [CrossRef]
- Maffei, M. Sites of synthesis, biochemistry and functional role of plant volatiles. S. Afr. J. Bot. 2010, 76, 612–631. [Google Scholar] [CrossRef]
- Bianchi, G.; Nuzzi, M.; Avitabile Leva, A.; Rizzolo, A. Development of a headspace-solid phase micro extraction method to monitor changes in volatile profile of rose (Rosa hybrida, cv David Austin) petals during processing. J. Chromatogr. A 2007, 1150, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Lijun, Z.; Chao, Y.; Cheng, B.; Wan, H.; Le, L.; Huitanga, P.; Qixianga, Z. Volatile compound analysis and aroma evaluation of tea-scented roses in China. Ind. Crops Prod. 2020, 155, 112735. [Google Scholar] [CrossRef]
- Loghmani-Khouzani, H.; Fini, O.S.; Safari, J. Essential oil composition of Rosa damascena Mill. cultivated in Central Iran. Iran J. Sci. Technol. Trans. A Sci. 2007, 14, 316–319. [Google Scholar]
- Demir, N.; Yildiz, O.; Alpaslan, M.; Hayaloglu, A.A. Evaluation of volatiles, phenolic compounds and antioxidant activites of rose hip (Rosa L.) fruits in Turkey. LWT Food Sci. Technol. 2013, 57, 126–133. [Google Scholar] [CrossRef]
- Abudayeh, Z.; Karpiuk, U.; Armoon, N.; Abualassal, Q.; Mallah, E.; Hassouneh, D.L.; Aldalahmeh, Y. Phytochemical, Physiochemical, macroscopic, and microscopic analysis of Rosa damascena flower petals and buds. J. Food Qual. 2022, 2022, 5079964. [Google Scholar] [CrossRef]
- Kunc, N.; Hudina, M.; Bavcon, J.; Vreš, B.; Luthar, Z.; Gostinčar, K.; Mikulič-Petkovšek, M.; Osterc, G.; Ravnjak, B. Characterization of the Slovene autochthonous rose hybrid Rosa pendulina × spinosissima (Rosa reversa Waldst. and Kit) using biochemical patterns of the plant blossoms. Plants 2023, 12, 505. [Google Scholar] [CrossRef] [PubMed]
- Baydar, N.; Baydar, H. Phenolic compounds, antiradical activity and antioxidant capacity of oil-bearing rose (Rosa damascena Mill.) extracts. Ind. Crops Prod. 2013, 41, 375–380. [Google Scholar] [CrossRef]
- Boskabady, M.H.; Shafei, M.N.; Saberi, Z.; Amini, S. Pharmacological effects of Rosa damascena. Iran. J. Basic Med. Sci. 2011, 14, 295–307. [Google Scholar] [CrossRef]
- Hongratanaworakit, T. Relaxing effect of rose oil on humans. Nat. Prod. Commun. 2009, 4, 2. [Google Scholar] [CrossRef]
- Li, M.; Xie, J.; Bai, X. Anti-aging potential, anti-tyrosinase and antibacterial activities of extracts and compounds isolated from Rosa chinensis cv. ‘JinBian’. Ind. Crops Prod. 2021, 159, 113059. [Google Scholar] [CrossRef]
- Younis, I.; El-Hawary, S.; Eldahshan, O.; Abdel-Aziz, M.; Ali, Z. Green synthesis of magnesium nanoparticles mediated from Rosa floribunda charisma extract and its antioxidant, antiaging and antibiofilm activities. Sci. Rep. 2021, 11, 16868. [Google Scholar] [CrossRef]
- Özdemir, N.; Budak, N. Bioactive compounds and volatile aroma compounds in rose (Rosa damascena Mill.) vinegar during the aging period. Food Biosci. 2022, 50, 102062. [Google Scholar] [CrossRef]
- Rubtsova, O.; Kolesnichenko, O.; Shumyk, M.; Shynder, O.; Chyzhankova, V.; Dzyba, A.; Hrysiuk, S. Scots roses in Northern Ukraine. Hortic. Sci. 2021, 48, 144–148. [Google Scholar] [CrossRef]
- Gudin, S. Rose breeding technologies. Acta Hortic. 2001, 547, 23–33. [Google Scholar] [CrossRef]
- Datta, S.K. Breeding of new ornamental varieties: Rose. Curr. Sci. 2018, 114, 1194–1206. [Google Scholar] [CrossRef]
- Meier, U.; Graf, H.; Hack, H.; Hess, M.; Kennel, W.; Klose, R.; Mappes, D.; Seipp, D.; Stauß, R.; Streif, J.; et al. Phanologische Entwicklungsstadien des Kernobstes (Malus domestica Borkh. und Pyrus communis L.), des Steinobstes (Prunus-Arten), der Johannisbeere (Ribes-Arten) und der Erdbeere (Fragaria × ananassa Duch.). Nachr. Dtsch. Pflanzenschutzd. 1994, 46, 141–153. [Google Scholar]
- Baluszynska, U.B.; Licznar-Malanczuk, M.; Medic, A.; Veberic, R.; Grohar, M.C. Influence of living mulch and nitrogen dose on yield and fruit quality parameters of Malus domestica Borkh. cv. ‘Sampion’. Agriculture 2023, 13, 921. [Google Scholar] [CrossRef]
- Interplant. 2023. Available online: https://www.interplantroses.nl/company (accessed on 10 September 2023).
- Kordes. 2023. Available online: https://southamptonrose.org/wp-content/uploads/2012/pdf/Brochure_Kordes.pdf (accessed on 10 September 2023).
- Jang, S.S.; Choi, J.S.; Kim, J.H.; Kim, N.; Ference, E.H. Discordance between subjective and objective measures of smell and taste in US adults. Otolaryngol.–Head Neck Surg. 2022, 166, 572–579. [Google Scholar] [CrossRef]
- Ryu, J.; Lyu, J.I.; Kim, D.-G.; Kim, J.-M.; Jo, Y.D.; Kang, S.-Y.; Kim, J.-B.; Ahn, J.-W.; Kim, S.H. Comparative analysis of volatile compounds of gamma-irradiated mutants of rose (Rosa hybrida). Plants 2020, 9, 1221. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Cheng, X.; Lu, Y.; Wang, H.; Chen, D.; Luo, C.; Liu, H.; Gao, S.; Lei, T.; Huang, C.; et al. Gas chromatography-mass spectrometry analysis of floral fragrance-related compounds in scented rose (Rosa hybrida) varieties and a subsequent evaluation on the basis of the analytical hierarchy process. Plant Physiol. Biochem. 2022, 185, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Miranda, M.C.; Schäfer, A.; Pham, M.N.; Greaney, A.; Arunachalam, P.S.; Navarro, M.J.; Tortorici, M.A.; Rogers, K.; O’Connor, M.A.; et al. Elicitation of broadly protective sarbecovirus immunity by receptor-binding domain nanoparticle vaccines. Plant Cell 2021, 184, 5432–5447.e16. [Google Scholar] [CrossRef] [PubMed]
- Guterman, I.; Shalit, M.; Menda, N.; Piestun, D.; Dafny-Yelin, M.; Shalev, G.; Weiss, D. Rose scent genomics approach to discovering novel foral fragrance-related genes. Plant Cell 2002, 14, 2325–2338. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, J.; Meyer-Gauen, G.; Croteau, R. Plant terpenoid synthases: Molecular biology and phylogenetic analysis. Proc. Nat. Acad. Sci. USA 1998, 95, 4126–4133. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, J.; Köllner, T.G.; Gershenzon, J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar] [CrossRef]
- Magnard, J.L.; Roccia, A.; Caissard, J.C.; Vergne, P.; Sun, P.; Hecquet, R.; Baudino, S. Biosynthesis of monoterpene scent compounds in roses. Science 2015, 349, 81–83. [Google Scholar] [CrossRef]
- Sun, P.; Schuurink, R.C.; Caissard, J.C.; Hugueney, P.; Baudino, S. My way: Noncanonical biosynthesis pathways for plant volatiles. Trends Plant Sci. 2016, 21, 884–894. [Google Scholar] [CrossRef] [PubMed]
- Dani, K.G.S.; Fineschi, S.; Michelozzi, M.; Trivellini, A.; Pollastri, S.; Loreto, F. Diversification of petal monoterpene profiles during floral development and senescence in wild roses: Relationships among geraniol content, petal colour, and floral lifespan. Oecologia 2021, 197, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Sparinska, A.; Rostoks, N. Volatile organic compounds of hybrid Rugosa roses in Latvia. Proc. Latv. Acad. Sci. Sect. B. Nat. Exact Appl. Sci. 2015, 69, 57–61. [Google Scholar] [CrossRef]
- Karani, L.W.; Tolo, F.M.; Karanja, S.M.; Khayeka−Wandabwa, C. Safety of Prunus africana and Warburgia ugandensis in Asthma Treatment. S. Afr. J. Bot. 2013, 88, 183–190. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Mai, J.; Li, W.; Ledesma-Amaro, R.; Ji, X.J. Engineering plant sesquiterpene synthesis into yeasts: A review. J. Agric. Food Chem. 2021, 69, 9498–9510. [Google Scholar] [CrossRef] [PubMed]
- Abbas, F.; Zhou, Y.; O’Neill Rothenberg, D.; Alam, I.; Ke, Y.; Wang, H.-C. Aroma components in horticultural crops: Chemical diversity and usage of metabolic engineering for industrial applications. Plants 2023, 12, 1748. [Google Scholar] [CrossRef] [PubMed]
- Libre Text. 2023. Available online: https://chem.libretexts.org/Bookshelves/General_Chemistry/ChemPRIME_(Moore_et_al.)/08%3A_Properties_of_Organic_Compounds/8.09%3A_Aromatic_Hydrocarbons (accessed on 10 September 2023).
- Bergougnoux, V.; Caissard, J.C.; Jullien, F.; Magnard, J.L.; Scalliet, G.; Cock, J.M.; Hugueney, P.; Baudino, S. Both the adaxial and abaxial epidermal layers of the rose petal emit volatile scent compounds. Planta 2007, 226, 853–866. [Google Scholar] [CrossRef]
- Riu-Aumatell, M.; Castellari, M.; López-Tamames, E.; Galassi, S.; Buxaderas, S. Characterisation of volatile compounds of fruit juices and nectars by HS/SPME and GC/MS. Food Chem. 2004, 87, 627–637. [Google Scholar] [CrossRef]
- Chemical Book. 2023. Available online: https://m.chemicalbook.com/ (accessed on 10 September 2023).
- Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Floral scents and fruit aromas: Functions, compositions, biosynthesis, and regulation. Front. Plant Sci. 2022, 13, 860157. [Google Scholar] [CrossRef]
- Tan, K.H.; Nishida, R. Methyl eugenol: Its occurrence, distribution, and role in nature, especially in relation to insect behavior and pollination. J. Insect Sci. 2012, 12, 56. [Google Scholar] [CrossRef] [PubMed]
- Rusanov, K.; Kovacheva, N.; Rusanova, M.; Atanassov, I. Low variability of flower volatiles of Rosa damascena Mill. plants from rose plantations along the Rose Valley, Bulgaria. Ind. Crops Prod. 2012, 37, 6–10. [Google Scholar] [CrossRef]
- Mileva, M.; Ilieva, Y.; Jovtchev, G.; Gateva, S.; Zaharieva, M.M.; Georgieva, A.; Dimitrova, L.; Dobreva, A.; Angelova, T.; Vilhelmova-Ilieva, N.; et al. Rose flowers-a delicate perfume or a natural healer? Biomolecules 2021, 11, 127. [Google Scholar] [CrossRef] [PubMed]
- Staudt, M.; Bertin, M. Light and temperature dependence of emission of cyclic and acyclic monoterpenes from holm oak (Quercus ilex L.) leaves. Plant Cell Environ. 1998, 21, 385–395. [Google Scholar] [CrossRef]
Figure 1.
Blossoms of (A) R. spinosissima, (B) ‘Fruhlingsmorgen’, (C) ‘Fruhlingsduft’, (D) ‘Maigold’, (E) ‘Fruhlingsgold’, (F) ‘Karl Foerster’, (G) ‘Mon Amie Claire’, (H) ‘Bourgogne’, (I) ‘Harstad’, (J) R. pendulina, (K) ‘Mount Everest’, (L) R. gallica, (M) ‘Violacea’, (N) ‘Splendens’, (O) ‘Complicata’, and (P) ‘Charles de Mills’.
Figure 1.
Blossoms of (A) R. spinosissima, (B) ‘Fruhlingsmorgen’, (C) ‘Fruhlingsduft’, (D) ‘Maigold’, (E) ‘Fruhlingsgold’, (F) ‘Karl Foerster’, (G) ‘Mon Amie Claire’, (H) ‘Bourgogne’, (I) ‘Harstad’, (J) R. pendulina, (K) ‘Mount Everest’, (L) R. gallica, (M) ‘Violacea’, (N) ‘Splendens’, (O) ‘Complicata’, and (P) ‘Charles de Mills’.

Figure 2.
Blossom of R. damascena.

Figure 3.
The number of compounds within individual groups (a) and the proportion (%) of individual groups (b) of VOCs in the analyzed samples.
Figure 3.
The number of compounds within individual groups (a) and the proportion (%) of individual groups (b) of VOCs in the analyzed samples.


Figure 4.
The total content of VOCs (µg IS equivalents/g sample) in the taxons of R. pendulina and cultivars derived from it in 2023. Results are expressed as compound concentration per the 3-nonanone standard. Means with standard errors are presented, and different letters indicate significant differences.
Figure 4.
The total content of VOCs (µg IS equivalents/g sample) in the taxons of R. pendulina and cultivars derived from it in 2023. Results are expressed as compound concentration per the 3-nonanone standard. Means with standard errors are presented, and different letters indicate significant differences.

Figure 5.
The total content of VOCs (µg IS equivalents/g sample) in the taxons of R. spinosissima and cultivars derived from it in 2023. Results are expressed as compound concentration per the 3-nonanone standard. Means with standard errors are presented, and different letters indicate significant differences at p ≤ 0.05.
Figure 5.
The total content of VOCs (µg IS equivalents/g sample) in the taxons of R. spinosissima and cultivars derived from it in 2023. Results are expressed as compound concentration per the 3-nonanone standard. Means with standard errors are presented, and different letters indicate significant differences at p ≤ 0.05.

Figure 6.
The total content of VOCs (µg IS equivalents/g sample) in the taxons of R. gallica and cultivars derived from it in 2023. Results are expressed as compound concentration per the 3-nonanone standard. Means with standard errors are presented, and different letters indicate significant differences at p ≤ 0.05.
Figure 6.
The total content of VOCs (µg IS equivalents/g sample) in the taxons of R. gallica and cultivars derived from it in 2023. Results are expressed as compound concentration per the 3-nonanone standard. Means with standard errors are presented, and different letters indicate significant differences at p ≤ 0.05.

Figure 7.
Cluster dendrogram for all analyzed species and cultivars of Rose spp.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Data of origin for cultivars used in the study.
| Cultivar | Species of Origin | Breeding Company | Year of the Origin |
|---|---|---|---|
| ‘Violacea’ | R. gallica | unknown (The Netherlands) | 1795 |
| ‘Splendens’ | R. gallica | unknown | 1583 |
| ‘Charles de Mills’ | R. gallica | unknown | 1790 |
| ‘Complicata’ | R. gallica | unknown | before 1873 |
| ‘Karl Foerster’ | R. spinosissima | Kordes (Germany) | 1930 |
| ‘Fruhlingsduft’ | R. spinosissima | Kordes (Germany) | 1941 |
| ‘Fruhlingsmorgen’ | R. spinosissima | Kordes (Germany) | 1941 |
| ‘Fruhlingsgold’ | R. spinosissima | Kordes (Germany) | 1937 |
| ‘Mon amie Claire’ | R. spinosissima | Ivan Louette (Belgium) | 2005 |
| ‘Maigold’ | R. spinosissima | Kordes (Germany) | 1953 |
| ‘Harstad’ | R. pendulina | unknown | unknown (old cultivar) |
| ‘Mount Everest’ | R. pendulina | Interplant (Netherland) | 1956 |
| ‘Bourgogne’ | R. pendulina | Interplant (Netherland) | 1983 |
Table 2.
Content (µg IS equivalents/g sample) of individual groups of VOCs in the analyzed species and cultivars. Results are expressed as the compound concentration per the 3-nonanone standard. Means with standard errors are presented, and different letters indicate significant differences between species and cultivars at p ≤ 0.05.
Table 2.
Content (µg IS equivalents/g sample) of individual groups of VOCs in the analyzed species and cultivars. Results are expressed as the compound concentration per the 3-nonanone standard. Means with standard errors are presented, and different letters indicate significant differences between species and cultivars at p ≤ 0.05.
| MONOTERPENES | SESQUITERPENES | ALDEHYDES | ALCOHOLS | KETONES | HYDROCARBONS | PHENYLPROPENES | OTHER COMPOUNDS | Total | |
|---|---|---|---|---|---|---|---|---|---|
| R. pendulina | 59.15 ± 5.322 e | 13.04 ± 2.543 c | 15.44 ± 1.092 bd | 4.44 ± 1.935 ade | - | 6.17 ± 0.876 ce | - | - | 98.24 ± 11.768 e |
| ‘Bourgogne’ | 0.28 ± 0.07 a | 1.24 ± 0.216 a | 17.01 ± 1.501 bd | 1.32 ± 0.215 ab | - | 2.04 ± 0.51 ab | - | 2.19 ± 0.38 c | 24.08 ± 2.892 ac |
| ‘Harstad’ | 46.09 ± 3.049 d | 5.00 ± 0.627 b | 23.40 ± 1.87 de | 2.74 ± 0.193 ade | 0.09 ± 0.007 ac | 4.48 ± 0.404 abe | 0.04 ± 0.006 a | 1.40 ± 0.17 b | 83.24 ± 6.272 de |
| ‘Mount Everest’ | 2.04 ± 0.596 a | 0.54 ± 0.075 a | 22.04 ± 1.263 de | 0.96 ± 0.42 a | - | 1.32 ± 0.208 a | - | 0.10 ± 0.004 a | 27.00 ± 2.548 ac |
| R. spinosissima | 0.08 ± 0.02 a | 0.03 ± 0.007 a | 12.79 ± 1.53 bc | 1.50 ± 0.41 ab | 0.01 ± 0.01 ac | 5.81 ± 1.05 bce | 0.28 ± 0.04 e | 0.12 ± 0.02 a | 20.62 ± 3.087 ac |
| ‘Fruhlingsmorgen’ | 0.23 ± 0.03 a | 0.24 ± 0.043 a | 19.30 ± 1.26 be | 2.23 ± 0.42 ad | 0.08 ± 0.01 c | 3.65 ± 0.283 acd | 0.03 ± 0.003 a | 0.03 ± 0.005 a | 2.579 ± 2.054 a |
| ‘Fruhlingsduft’ | - | 0.61 ± 0.05 a | 1.04 ± 0.16 a | 9.74 ± 2.113 f | 0.19 ± 0.01 d | 12.30 ± 1.874 f | 0.11 ± 0.03 bd | 0.09 ± 0.01 a | 24.08 ± 4.247 ac |
| ‘Karl Foerster’ | - | 0.51 ± 0.022 a | 1.07 ± 0.15 a | 1.47 ± 0.302 ab | 0.52 ± 0.017 a | 3.45 ± 0.362 acd | - | 0.03 ± 0.002 a | 7.05 ± 0.855 ab |
| ‘Mon amie Claire’ | - | 0.28 ± 0.02 a | 0.30 ± 0.03 a | 5.95 ± 0.655 cdf | - | 3.32 ± 0.43 acd | 0.15 ± 0.02 d | 0.11 ± 0.01 a | 10.11 ± 1.165 ab |
| ‘Maigold’ | 2.98 ± 0.40 ab | 0.62 ± 0.02 a | 69.57 ± 3.20 g | 6.38 ± 1.356 df | 0.19 ± 0.003 bc | 8.60 ± 0.864 e | 0.05 ± 0.03 ab | 1.76 ± 0.227 bc | 90.15 ± 6.10 de |
| ‘Fruhlingsgold’ | 1.12 ± 0.27 a | 0.34 ± 0.027 a | 40.96 ± 1.612 f | 6.04 ± 1.70 df | 0.15 ± 0.008 a | 5.22 ± 0.437 abe | 0.06 ± 0.007 ab | 0.12 ± 0.01 a | 54.40 ± 4.071 cd |
| R. gallica | 5.26 ± 1.204 ab | 0.03 ± 0.005 a | 21.84 ± 1.27 cde | 2.45 ± 0.423 ad | 0.09 ± 0.003 ac | 6.98 ± 0.828 de | - | 0.14 ± 0.007 a | 36.84 ± 3.745 ac |
| ‘Charles de Mills’ | 3.60 ± 1.28 ab | 0.21 ± 0.032 a | 28.8 ± 3.521 e | 5.47 ± 1.203 bde | 0.10 ± 0.008 ac | 4.91 ± 0.544 abe | - | 0.03 ± 0.02 a | 43.12 ± 6.59 bc |
| ‘Violacea’ | 126.32 ± 19.24 f | 0.08 ± 0.01 a | 20.13 ± 2.605 cde | 4.45 ± 1.75 ade | 0.11 ± 0.01 ac | 17.01 ± 2.86 g | - | 0.14 ± 0.02 a | 168.24 ± 26.495 f |
| ‘Splendens’ | 0.58 ± 0.10 a | 0.48 ± 0.05 a | 18.00 ± 1.91 bd | 1.60 ± 0.199 abc | 0.11 ± 0.009 ac | 1.93 ± 0.097 ab | - | 0.40 ± 0.043 a | 23.10 ± 2.408 ac |
| ‘Complicata’ | 14.75 ± 3.22 bc | 0.18 ± 0.026 a | 10.74 ± 1.43 b | 1.45 ± 0.486 ab | 0.05 ± 0.008 ab | 2.91 ± 0.234 ac | 0.11 ± 0.02 cd | 0.07 ± 0.014 a | 30.26 ± 5.438 ac |
| R. damascena | 18.07 ± 2.71 c | 0.66 ± 0.16 a | 16.64 ± 3.164 bd | 6.76 ± 1.231 ef | 0.19 ± 0.03 c | 2.09 ± 0.302 ab | 0.06 ± 0.01 abc | 0.08 ± 0.002 a | 44.55 ± 7.609 bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kunc, N.; Hudina, M.; Osterc, G.; Grohar, M.C. Determination of Volatile Compounds in Blossoms of Rosa spinosissima, Rosa pendulina, Rosa gallica, and Their Cultivars. Agriculture 2024, 14, 253. https://doi.org/10.3390/agriculture14020253
AMA Style
Kunc N, Hudina M, Osterc G, Grohar MC. Determination of Volatile Compounds in Blossoms of Rosa spinosissima, Rosa pendulina, Rosa gallica, and Their Cultivars. Agriculture. 2024; 14(2):253. https://doi.org/10.3390/agriculture14020253
Chicago/Turabian StyleKunc, Nina, Metka Hudina, Gregor Osterc, and Mariana Cecilia Grohar. 2024. "Determination of Volatile Compounds in Blossoms of Rosa spinosissima, Rosa pendulina, Rosa gallica, and Their Cultivars" Agriculture 14, no. 2: 253. https://doi.org/10.3390/agriculture14020253
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.