
Longitudinal Distribution Map of the Active Components and Endophytic Fungi in Angelica sinensis (Oliv.) Diels Root and Their Potential Correlations
Abstract
:1. Introduction
2. Materials and Methods
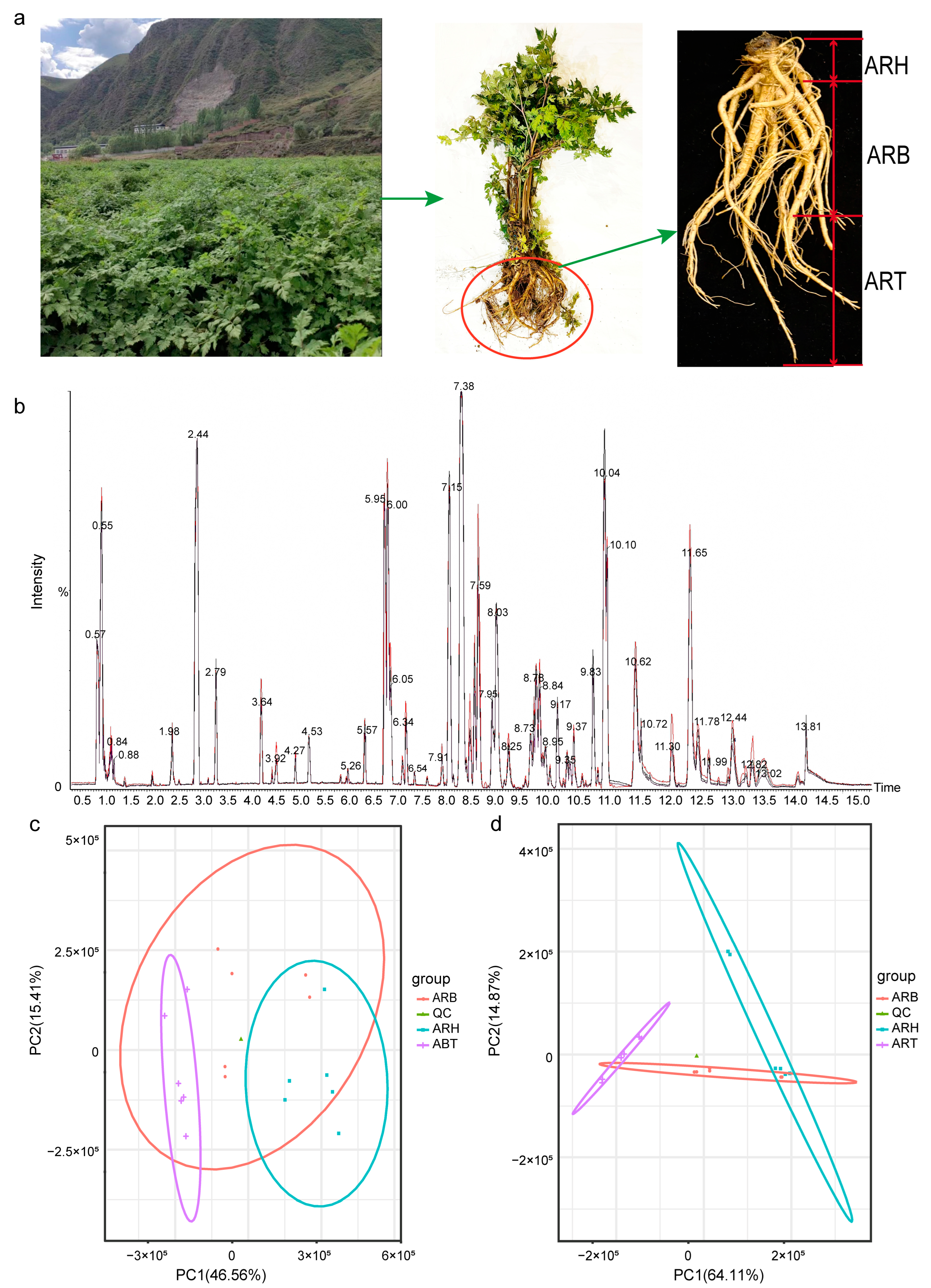
2.1. Collection of Fresh Plants and Preparation of Samples
2.2. Metabolite Extraction
2.3. Detection and Identification of Metabolites
2.4. Metabonomic Analysis
2.5. Metagenome Extraction, ITS Sequence Amplification, and Sequencing
2.6. Microbial Diversity Analysis
2.7. Correlation Analysis between the Microbiome and Metabolome
2.8. Data Analysis
3. Results
3.1. Metabolite Information for Ang Root and the Metabolic Patterns of the Different Medicinal Parts
3.2. Differential Metabolites between Different Medicinal Parts
3.3. Dominant Metabolites in the Different Medicinal Parts of Ang Root and Their Medicinal Efficacy Characteristics
3.4. Functional Enrichment Patterns of Differential Metabolites
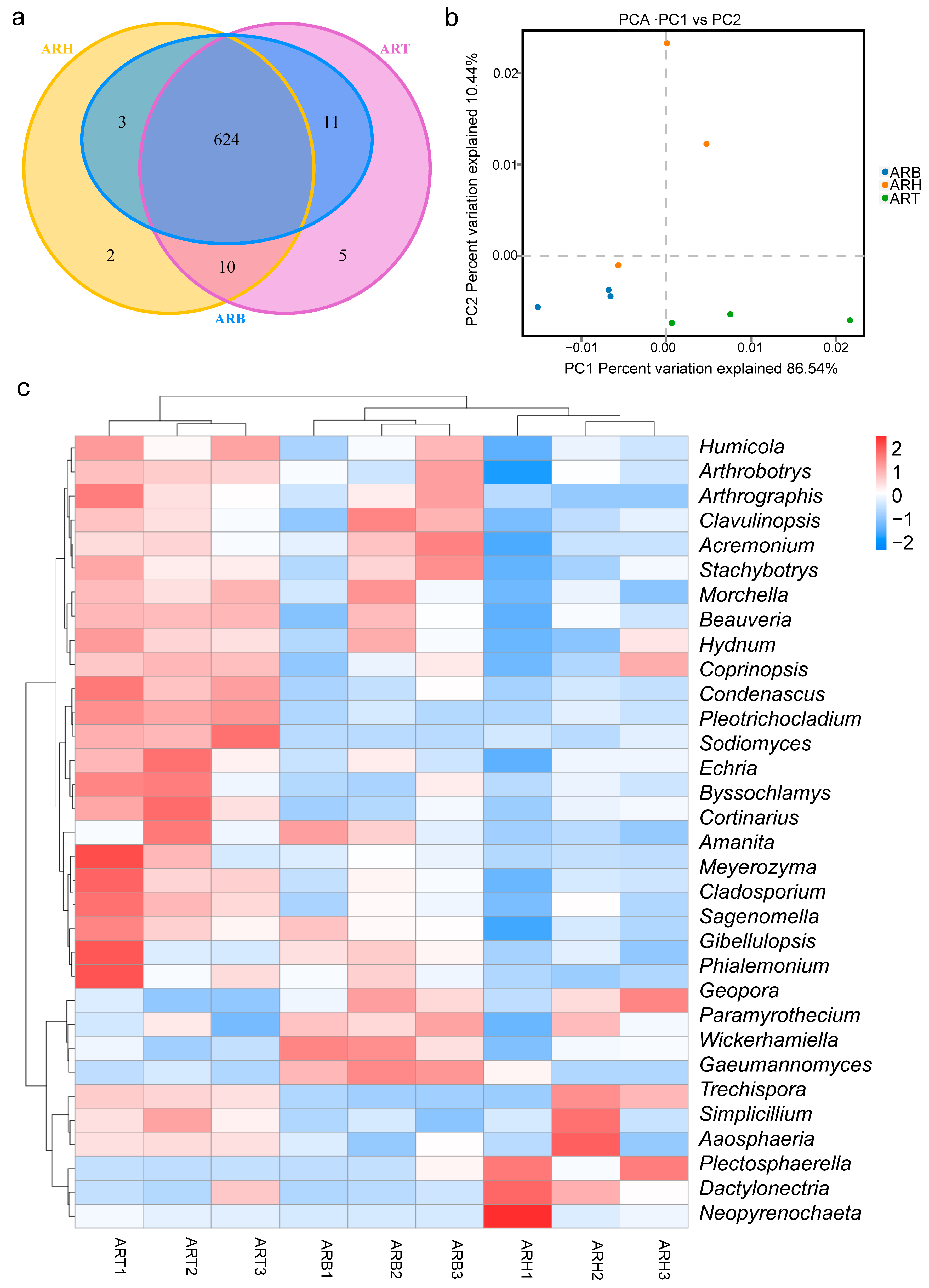
3.5. Community Structures of Endophytic Fungi in Ang Root and Distribution Characteristics
3.6. Endophytic Fungi with Significant Differences between the Different Medicinal Parts of Ang Root
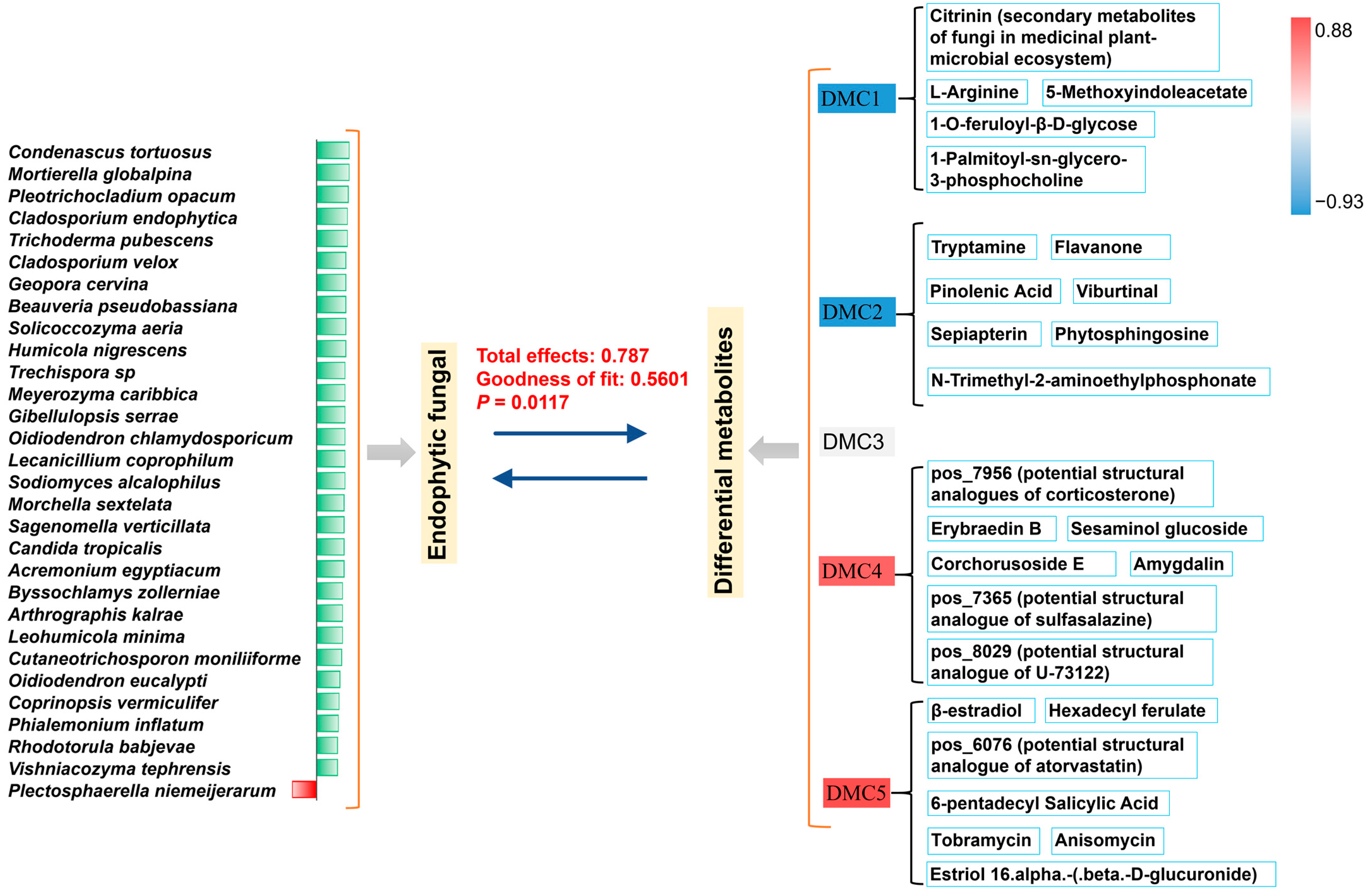
3.7. Key Endophytic Fungi and Active Components Distributed across Different Medicinal Parts and Their Interactions
4. Discussion
4.1. Supplement Showing Distribution Map of Active Compounds in Different Medicinal Parts of Ang Root
4.2. Spatial Distribution Patterns and Interaction Mechanisms of Endophytic Fungi and Active Metabolites
4.3. Important Research and Application Values of Key Metabolites and Endophytic Fungi Closely Related across Adjacent Medicinal Parts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hou, C.Y.; Yin, M.S.; Lan, P.; Wang, H.R.; Nie, H.; Ji, X.L. Recent progress in the research of Angelica sinensis (Oliv.) Diels polysaccharides: Extraction, purification, structure and bioactivities. Chem. Biol. Technol. Agric. 2021, 8, 1–14. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, Y.G.; Shi, L.P.; Liu, J.X.; Yu, Y. Research progress on pharmacological effects of Angelica sinensis prepared products and their chemical constituents. Mod. Chin. Med. 2023, 48, 6003–6010. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Bi, W.G.; Yu, Y.; Liao, W.B. Angelica sinensis (Oliv.) Diels in China: Distribution, cultivation, utilization and variation. Genet. Resour. Crop Evol. 2012, 59, 607–613. [Google Scholar] [CrossRef]
- Wu, W.G.; Cai, Z.P.; Wang, G.X.; Wang, B. Morphological change characteristics and material accumulative process of Angelica sinensis. Mod. Chin. Med. 2022, 24, 1945–1951. [Google Scholar] [CrossRef]
- Xu, Z.W.; Du, W.F.; Ma, X.H.; Bi, Y.H.; Wang, B.C.; Bian, N.; Li, J.W. Analysis of the rationality of clinical use of Angelica sinensis based on the difference of effective components before and after compatibility of different medicinal parts. Lishizhen Med. Mater. Med. Res. 2022, 33, 1513–1515. [Google Scholar] [CrossRef]
- Xue, W.X.; Hua, Y.L.; Guo, Y.S.; Ji, P.; Wu, H.Y.; Wei, Y.M. Changes of composition in different parts of Angelicae sinensis based on geoherbs. J. Gansu Agric. Univ. 2012, 47, 149–154. [Google Scholar] [CrossRef]
- Li, J.; Wei, Y.H.; Qin, X.M.; Li, Z.Y. Chemical Comparison constituents on different parts of Angelica sinensis Radix based on NMR metabonomics. Chin. Tradit. Herb. Drugs 2017, 48, 1409–1415. [Google Scholar]
- Lin, W.J.; Sun, F.L.; Zhang, Y.M.; Xu, X.M.; Lu, X.H.; Li, L.S.; Xu, R.Q. Comparative transcriptome and metabolite profiling of four tissues from Alisma orientale (Sam.) Juzep reveals its inflorescence developmental and medicinal characteristics. Sci. Rep. 2019, 9, 12310. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Xu, X.D.; Chen, Y.Z.; Gao, J.; Shi, Q.Y.; Tian, L.; Cao, L. Combined Metabolome and Transcriptome Analyses Reveal the Flavonoids Changes and Biosynthesis Mechanisms in Different Organs of Hibiseu manihot L. Front. Plant Sci. 2022, 13, 817378. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Huang, S.W.; Duan, H.J.; Tang, H.; Liu, B.Y.; Li, W.L. Isolation, identification and antimicrobial activity of endophytic fungi from Polygala fallax Hemsl. J. South. Agric. 2023, 54, 1136–1145. [Google Scholar] [CrossRef]
- Zhu, J.; Li, X.J.; Sun, L.; Guo, S.X.; Chen, J. Ecological distribution and diversity of medical Ferula species produced in Xinjiang. J. Tradit. Chin. Med. 2015, 40, 356–361. [Google Scholar] [CrossRef]
- Wu, W.; Chen, W.H.; Liu, S.Y.; Wu, J.J.; Zhu, Y.T.; Qin, L.P.; Zhu, B. Benefcial relationships between endophytic bacteria and medicinal plants. Front. Plant Sci. 2021, 12, 646146. [Google Scholar] [CrossRef] [PubMed]
- Katoch, M.; Pull, S. Endophytic fungi associated with Monarda citriodora, an aromatic and medicinal plant and their biocontrol potential. Pharm. Biol. 2017, 55, 1528–1535. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.Y.; Liu, X.; Cui, J.L.; Wang, J.H.; Wang, M.L.; Zhang, G. Endophytic fungi and their bioactive secondary metabolites in medicinal leguminosae plants: Nearly untapped medical resources. FEMS Microbiol. Lett. 2022, 369, fnac052. [Google Scholar] [CrossRef]
- Chen, H.M. Deciphering Salvia miltiorrhiza associated microbiome and regulating mechanism of tanshinone biosynthesis by their dominant species. Northwest AF Univ. 2021. [Google Scholar] [CrossRef]
- Qin, D.; You, C.; Lan, W.Y.; Wang, Y.M.; Yu, B.H.; Peng, Y.J.; Xu, J.R.; Dong, J.Y. Microbial assemblages of Schisandraceae plants and the correlations between endophytic species and the accumulation of secondary metabolites. Plant Soil 2023, 483, 85–107. [Google Scholar] [CrossRef]
- Fu, L.F.; Shao, S.; Feng, Y.; Ye, F.; Sun, X.; Wang, Q.L.; Yu, F.; Wang, Q.S.; Huang, B.Y.; Niu, P.H.; et al. Mechanism of Microbial Metabolite Leupeptin in the Treatment of COVID-19 by Traditional Chinese Medicine Herbs. mBio 2021, 12, e0222021. [Google Scholar] [CrossRef]
- Yuan, M.Y.; Li, Y.; Tan, X.H.; Liu, Y.; Fang, X.Y.; Ran, Y.L.; He, M.J. Active components of Fritillaria hupehensis and secondary metabolites of its endophytes. J. Huazhong Agric. Univ. 2021, 40, 27–34. [Google Scholar] [CrossRef]
- Wu, Z.J.; Wang, X.W.; Chen, M.; Hu, H.Y.; Cao, J.; Chai, T.Y.; Wang, H. A Study on Tissue-Specific Metabolite Variations in Polygonum cuspidatum by High-Resolution Mass Spectrometry-Based Metabolic Profiling. Molecules 2019, 24, 1058. [Google Scholar] [CrossRef]
- Chen, X.; Li, Q.; Cao, Q.Q.; Huang, C.P. Community structure of cultivable endophytes in different organs of Gentiana straminea maxim and its correlation with the content of gentiopicrin. Food Sci. Biotechnol. 2019, 38, 21–29. [Google Scholar] [CrossRef]
- Zhou, H.L.; Chen, H.Y.; Bao, D.P.; Shin, T.Y.; Zhong, Y.J.; Zhang, X.; Wu, Y.Y. Recent advances of gamma-aminobutyric acid: Physiological and immunity function, enrichment, and metabolic pathway. Front. Nutr. 2022, 9, 1076223. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, H.P.; Cheng, H.Y.; Wan, Y.; Yang, L. Diversity and biomarker exploration of endophytic fungi communities in different varieties of Paris Rhizoma. Chin. Tradit. Herb. Drugs 2020, 51, 4531–4537. [Google Scholar] [CrossRef]
- White, R.A.; Borkum, M.I.; Rivas-Ubach, A.; Bilbao, A.; Wendler, J.P.; Colby, S.M.; Koberl, M.; Jansson, C. From data to knowledge: The future of multi-omics data analysis for the rhizosphere. Rhizosphere 2017, 3, 222–229. [Google Scholar] [CrossRef]
- Mannaa, M.; Han, G.; Seo, Y.S.; Park, I. Evolution of Food Fermentation Processes and the Use of Multi-Omics in Deciphering the Roles of the Microbiota. Foods 2021, 10, 2861. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Duan, W.Y.; Ran, Z.F.; Chen, X.L.; Yu, H.X.; Fang, L.; Guo, L.P.; Zhou, J. Diversity and correlation analysis of endophytes and metabolites of Panax quinquefolius L. in various tissues. BMC Plant Biol. 2023, 23, 275. [Google Scholar] [CrossRef]
- Marsola, S.J.; Jorge, L.F.; Meniqueti, A.B.; Berteli, M.B.D.; de Lima, T.E.F.; Bezerra, J.L.; Lopes, A.D.; Gazim, Z.C.; do Valle, J.S.; Colauto, N.B.; et al. Endophytic fungi of Brunfelsia uniflora: Isolation, cryopreservation, and determination of enzymatic and antioxidant activity. World J. Microbiol. Biotechnol. 2022, 38, 94. [Google Scholar] [CrossRef]
- Zheng, Z.Y.; Chai, S.T.; Chen, J.; Yang, H.; Chang, J.Y.; Yang, G.E. Isolation and identification of flavonoid-producing endophytic fungi from Loranthus tanakae Franch. & Sav that exhibit antioxidant and antibacterial activities. J. Appl. Microbiol. 2022, 133, 1892–1904. [Google Scholar] [CrossRef]
- Sanches, P.H.G.; de Oliveira, D.C.; dos Reis, I.G.M.; Fernandes, A.; Silva, A.A.R.; Eberlin, M.N.; Carvalho, P.O.; Duarte, G.H.B.; Porcari, A.M. Fitting Structure-Data Files (.SDF) Libraries to Progenesis QI Identification Searches. J. Braz. Chem. Soc. 2023, 34, 1013–1019. [Google Scholar] [CrossRef]
- Huang, G.R.; Wu, Y.B.; Liu, M.; Sun, X.J.; Lu, W.Y.; Gao, B.Y.; Wang, T.T.Y.; Yu, L.L. Potential Biomarkers for Early Detection of 3-MCPD Dipalmitate Exposure in Sprague-Dawley Rats. J. Agric. Food Chem. 2020, 68, 9594–9602. [Google Scholar] [CrossRef]
- Wu, M.X.; Wang, Y.H.; Wang, Y.J.; Wang, X.F.; Yu, M.; Liu, G.X.; Tang, H. Study on the diversity of epiphytic bacteria on corn and alfalfa using Illumina MiSeq/NovaSeq high-throughput sequencing system. Ann. Microbiol. 2021, 71, 1–11. [Google Scholar] [CrossRef]
- Enjalbert, F.; Zened, A.; Cauquil, L.; Meynadier, A. Integrating data from spontaneous and induced trans-10 shift of ruminal biohydrogenation reveals discriminant bacterial community changes at the OTU level. Front. Microbiol. 2023, 13, 1012341. [Google Scholar] [CrossRef]
- Banerjee, S.; Schlaeppi, K.; van der Heijden, M.G.A. Keystone taxa as drivers of microbiome structure and functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef]
- Lefcheck, J.S. PIECEWISESEM: Piecewise structural equation modelling in R for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- McKenna, S.; Meyer, M.; Gregg, C.; Gerber, S. s-CorrPlot: An Interactive Scatterplot for Exploring Correlation. J. Comput. Graph. Stat. 2016, 25, 445–463. [Google Scholar] [CrossRef]
- You, Q.D. Pharmaceutical Chemistry; People’s Medical Publishing House: Beijing, China, 2016. [Google Scholar]
- Zhu, D.Y. Biopharmaceutical Analysis and Testing, 2nd ed.; Chemical Industry Press: Beijing, China, 2016. [Google Scholar]
- Kong, L.Y.; Feng, W.S. Chemistry of Traditional Chinese Medicine; People’s Medical Publishing House: Beijing, China, 2021. [Google Scholar]
- Ispirli, H.; Colquhoun, I.J.; Sahin, E.; Sagdic, O.; Dertli, E. Preparation of gentiobiose-derived oligosaccharides by glucansucrase E81 and determination of prebiotic and immune-modulatory functions. Carbohydr. Res. 2019, 486, 107837. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.J.; Huang, Z.W.; Zhu, W.J.; Liu, Y.Y.; Bai, X.L.; Zhang, H.W. Fusarium-Derived Secondary Metabolites with Antimicrobial Effects. Molecules 2023, 28, 3424. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Chen, L.; Xin, H.L.; Zheng, C.J.; Rahman, K.; Han, T.; Qin, L.P. A friendly relationship between endophytic fungi and medicinal plants: A systematic review. Front. Microbiol. 2016, 7, 906. [Google Scholar] [CrossRef]
- Liang, Y.; Li, S.W.; Song, X.D.; Zhou, D.S.; Zhi, D.J.; Hao, B.C.; Liu, Y.; Liang, J.P.; Wang, Z. Swainsonine producing performance of Alternaria oxytropis was improved by heavy-ion mutagenesis technology. FEMS Microbiol. Lett. 2021, 368, fnab047. [Google Scholar] [CrossRef]
- Moreno-Gavira, A.; Dianez, F.; Sanchez-Montesinos, B.; Santos, M. Paecilomyces variotii as A Plant-Growth Promoter in Horticulture. Agronomy 2020, 10, 597. [Google Scholar] [CrossRef]
- Raut, I.; Calin, M.; Capra, L.; Gurban, A.M.; Doni, M.; Radu, N.; Jecu, L. Cladosporium sp. Isolate as Fungal Plant Growth Promoting Agent. Agronomy 2021, 11, 392. [Google Scholar] [CrossRef]
- Zhang, P. Secondary metabolites of four algal/mangrove-derived endophytic fungi and their bioactivities. Univ. Chin. Acad. Sci. 2015. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | log2FC | p | VIP | Efficacies |
|---|---|---|---|---|
| Up-regulated in ART | ||||
| pos_8029 (potential structural analogue of U-73122) | 29.619735 | 0.000009 | 1.548186 | A&J |
| Corchorusoside E | 28.693819 | 0.001097 | 1.429212 | A |
| Ile Val Val Phe | 28.417091 | 0.006163 | 1.290369 | B&F |
| His His Arg Lys | 27.949976 | 0.000712 | 1.446415 | B&F |
| 11-Deoxy-17-hydroxycorticosterone | 26.934995 | 0.003347 | 1.341104 | H |
| 6-pentadecyl salicylic acid | 26.593057 | 0.011570 | 1.223086 | A&D |
| pos_7956 (potential structural analogues of corticosterone) | 26.034194 | 0.000350 | 1.465804 | H |
| Thapsigargin | 23.260326 | 0.002925 | 1.351772 | L |
| Ononin | 18.885931 | 0.041072 | 1.101388 | A&D&E&K&J |
| Hexadecyl ferulate | 18.372359 | 0.024347 | 1.037334 | A |
| Up-regulated in ARH | ||||
| Virginiamycin S1 | 24.388477 | 0.013138 | 1.186106 | D |
| 2’-O-methylcytidine | 18.113741 | 0.026446 | 1.086260 | O/U |
| beta-Nicotinamide D-ribonucleotide | 15.582520 | 0.000222 | 1.485885 | O/U |
| 6,7-Dimethoxyisoquinoline | 12.977668 | 0.018082 | 1.170311 | O/U |
| L-alpha-Glutamyl-L-tyrosine | 12.822422 | 0.001699 | 1.410215 | B&F&I |
| Anethole | 12.567253 | 0.012978 | 1.204438 | B |
| 7-(4-Hydroxyphenyl)-1-phenyl-4-hepten-3-one | 10.474889 | 0.000054 | 1.525504 | D&J |
| Viburtinal | 9.307067 | 0.029676 | 1.093222 | C&E&I&J |
| Tryptamine | 7.205199 | 0.000005 | 1.524918 | C |
| Nalpha-Methylhistidine | 6.841870 | 0.019508 | 1.133064 | O/U |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Guo, R.; Geng, Y.; Shang, H.; Guo, X.; Wu, Y.; Wang, Y.; Li, L.; Li, X.; Zhang, S.; et al. Longitudinal Distribution Map of the Active Components and Endophytic Fungi in Angelica sinensis (Oliv.) Diels Root and Their Potential Correlations. Metabolites 2024, 14, 48. https://doi.org/10.3390/metabo14010048
Sun Y, Guo R, Geng Y, Shang H, Guo X, Wu Y, Wang Y, Li L, Li X, Zhang S, et al. Longitudinal Distribution Map of the Active Components and Endophytic Fungi in Angelica sinensis (Oliv.) Diels Root and Their Potential Correlations. Metabolites. 2024; 14(1):48. https://doi.org/10.3390/metabo14010048
Chicago/Turabian StyleSun, Ying, Rong Guo, Yuting Geng, Hushan Shang, Xiaopeng Guo, Yue Wu, Yonggang Wang, Li Li, Xuee Li, Shengli Zhang, and et al. 2024. "Longitudinal Distribution Map of the Active Components and Endophytic Fungi in Angelica sinensis (Oliv.) Diels Root and Their Potential Correlations" Metabolites 14, no. 1: 48. https://doi.org/10.3390/metabo14010048