
Molecular Phylogenetic Diversity and Biological Characterization of Diaporthe Species Associated with Leaf Spots of Camellia sinensis in Taiwan

 ,
, 
 , , and
, , and
Abstract
:1. Introduction
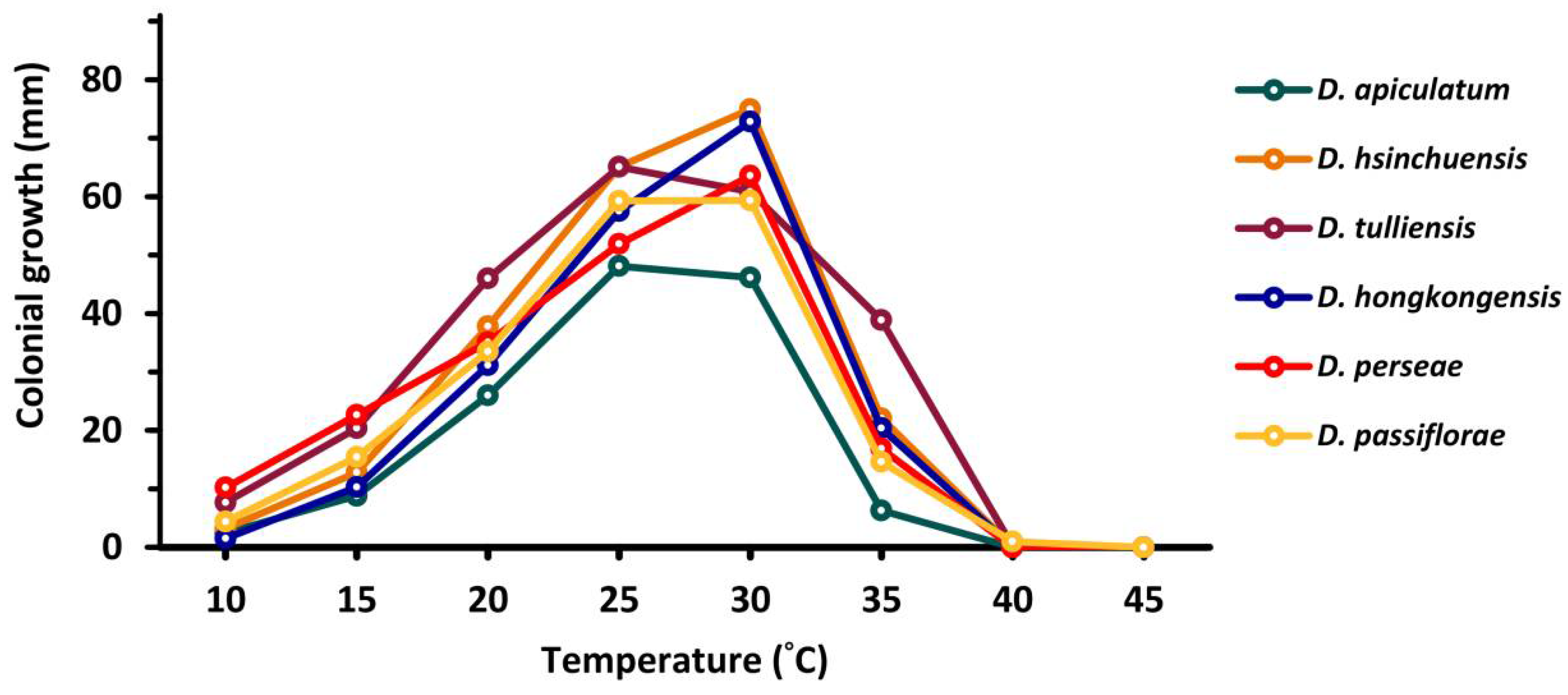
2. Results
2.1. Fungal Isolation
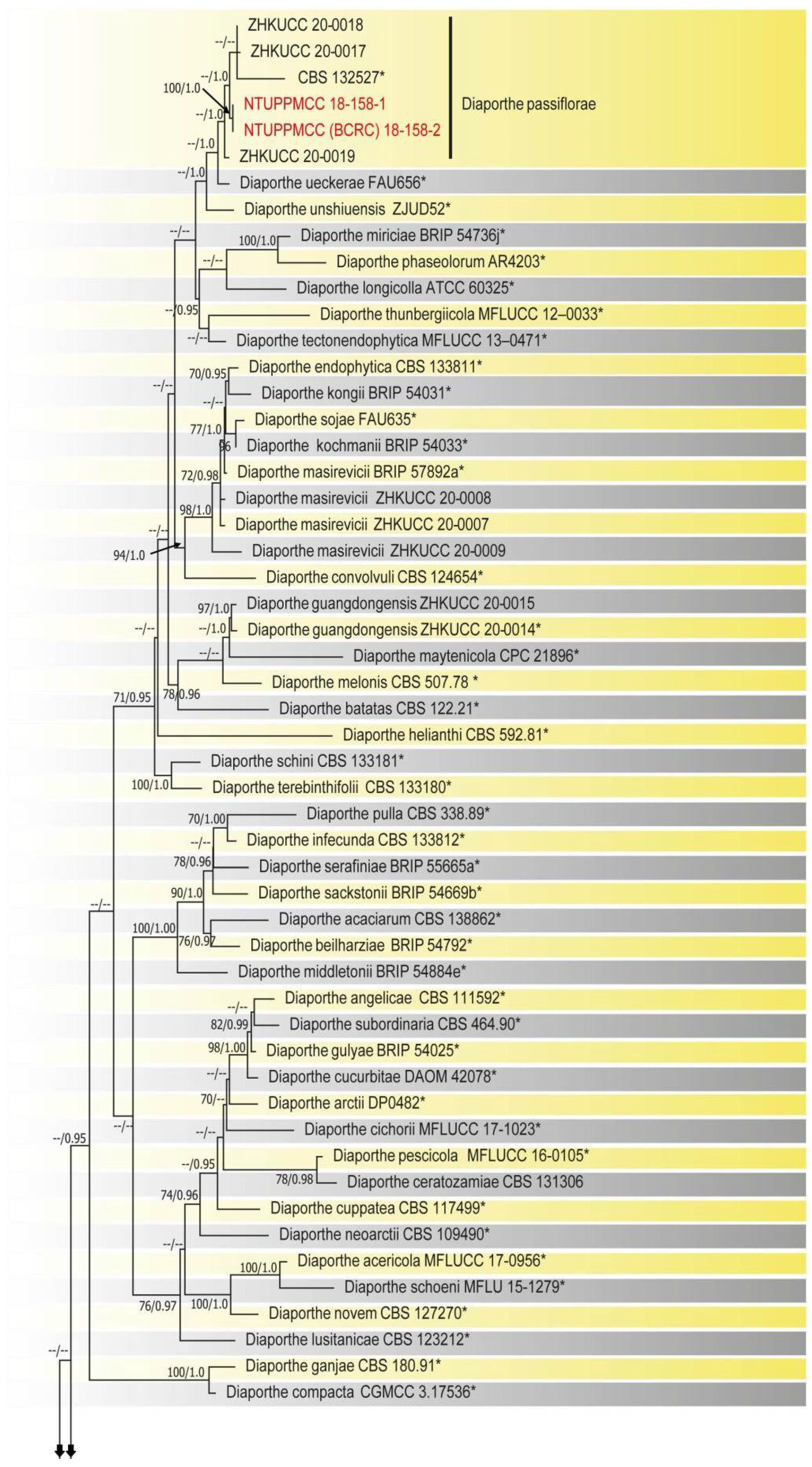
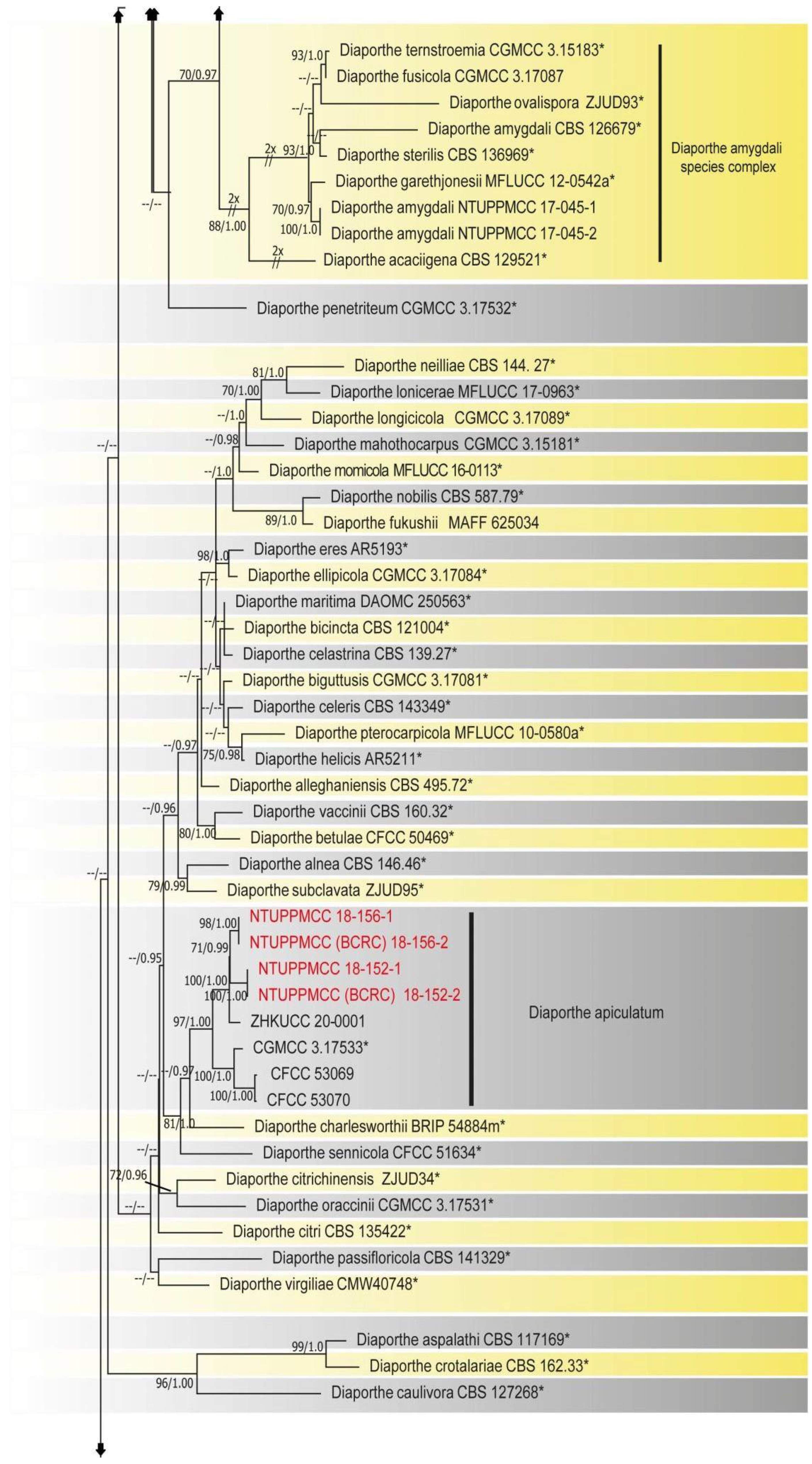
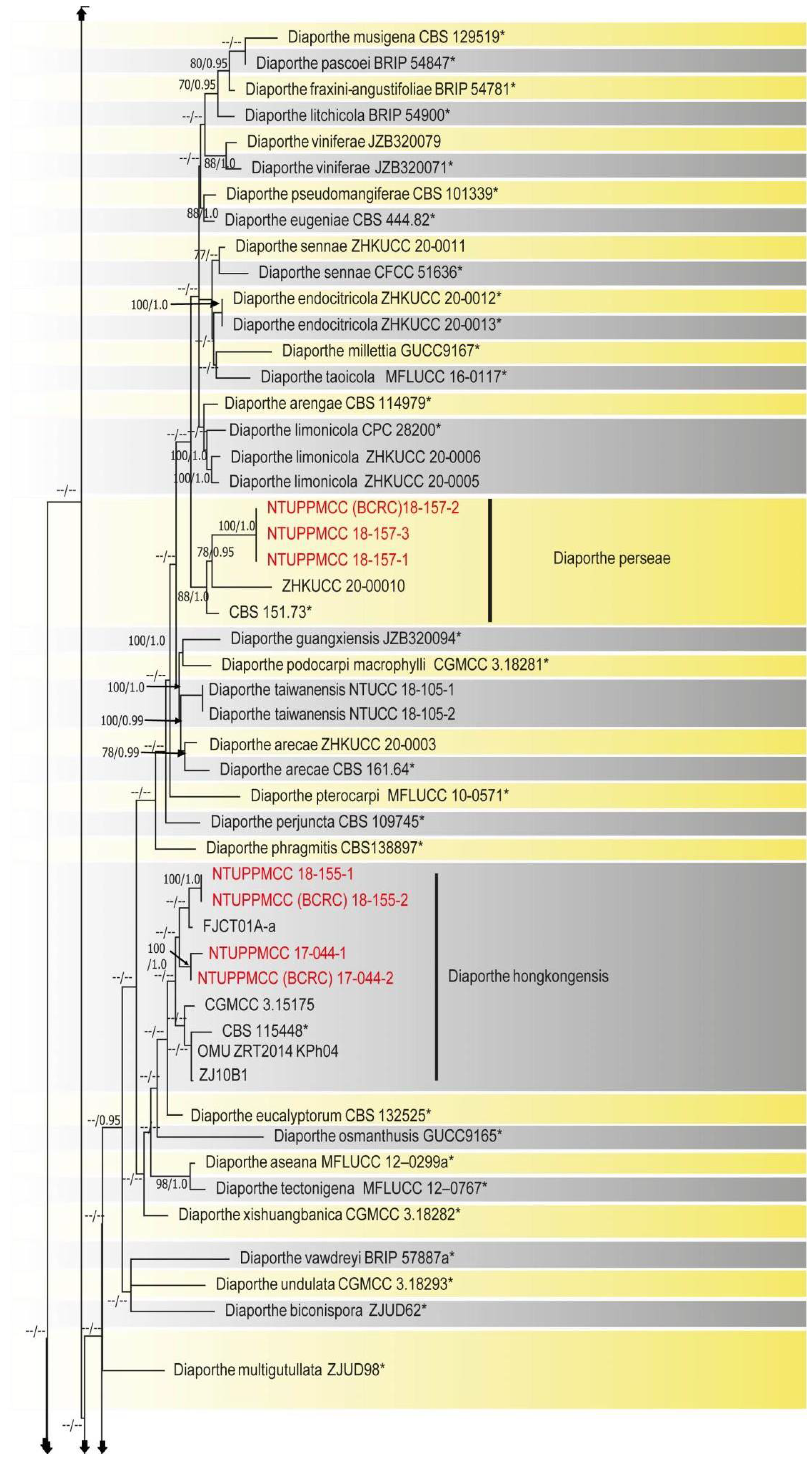
2.2. Phylogenetic Evaluation of the Concatenated Data Matrix of Diaporthe Species
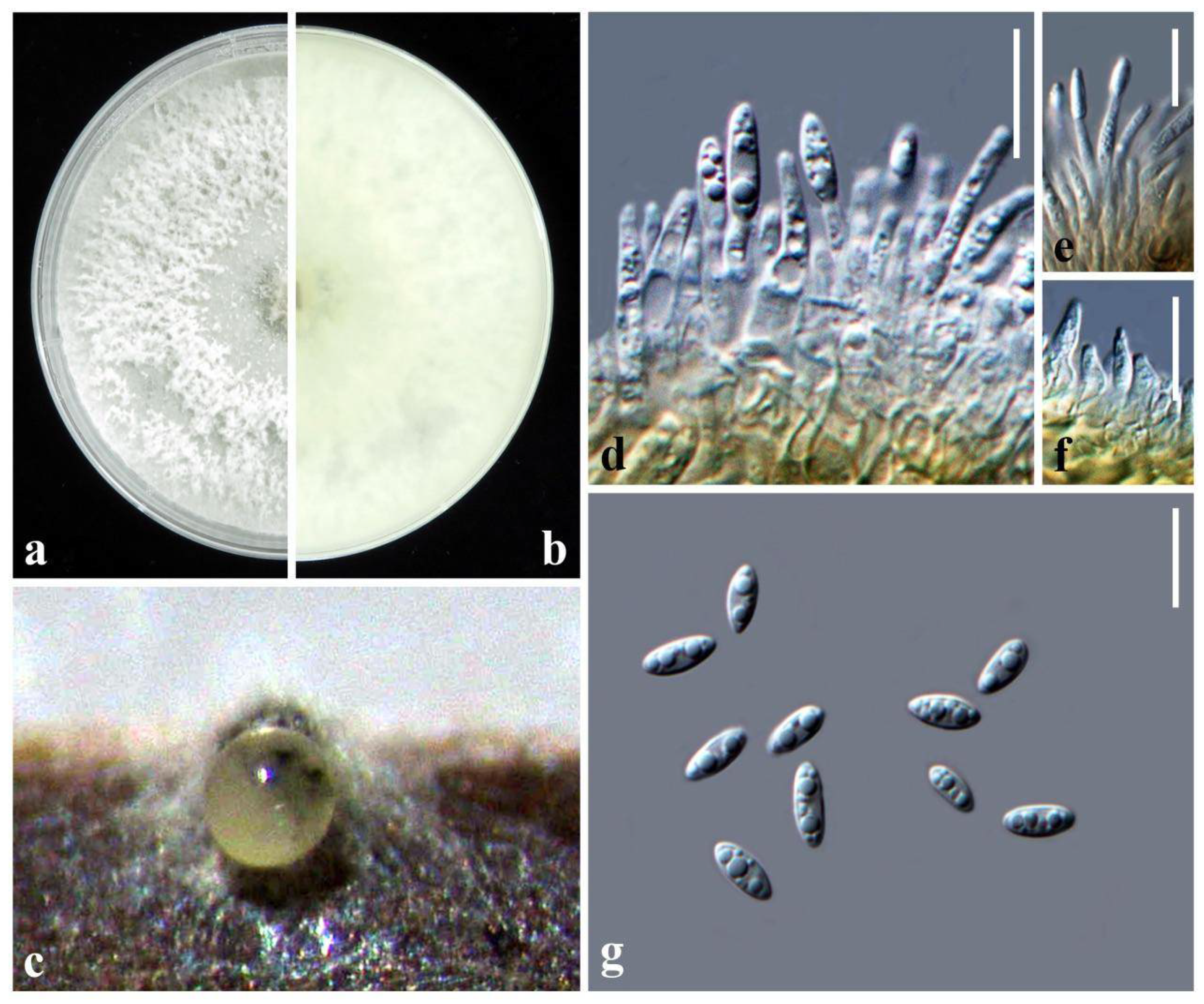
2.3. Taxonomy Diaporthe hsinchuensis Ariyawansa and I. Tsai, sp. nov.
2.4. Growth Rate
2.5. Temperature Effects
2.6. Optimal pH
2.7. Pathogenicity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sample Collection, Fungal Isolation, and Morphological Examination
5.2. DNA Extraction, PCR, and Sequencing
5.3. Strain Selection, Sequence Alignment, and Phylogenetic Analysis
5.4. Mycelial Growth Test
5.5. Temperature and pH Effects on Mycelial Growth
5.6. Pathogenicity Assessment
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. Insights into the genus Diaporthe: Phylogenetic species delimitation in the D. eres species complex. Fungal Divers. 2014, 67, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Fan, X.L.; Guarnaccia, V.; Tian, C.M. High diversity of Diaporthe species associated with dieback diseases in China, with twelve new species described. MycoKeys 2018, 39, 97–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Liu, F.; Cai, L. Unravelling Diaporthe species associated with Camellia. Syst. Biodivers. 2016, 14, 102–117. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Tsai, I.; Withee, P.; Tanjira, M.; Yen, C.Y.; Al-Rashed, S.; Elgorban, A.M.; Cheewangkoon, R. Diaporthe taiwanensis: A new taxon causing leaf spots and necrosis on Ixora chinensis in Taiwan. Phytotaxa 2020, 461, 155–165. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. The Diaporthe sojae species complex: Phylogenetic re-assessment of pathogens associated with soybean, cucurbits and other field crops. Fungal Biol. 2015, 119, 383–407. [Google Scholar] [CrossRef]
- Mondal, S.N.; Vicent, A.; Reis, R.F.; Timmer, L.W. Efficacy of pre-and postinoculation application of fungicides to expanding young citrus leaves for control of melanose, scab, and Alternaria brown spot. Plant Dis. 2007, 91, 1600–1606. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Hyde, K.D. Species limits in Diaporthe: Molecular re-assessment of D. citri, D. cytosporella, D. foeniculina and D. rudis. Persoonia 2014, 32, 83–101. [Google Scholar] [CrossRef] [Green Version]
- Guarnaccia, V.; Crous, P.W. Emerging citrus diseases in Europe caused by species of Diaporthe. IMA Fungus 2017, 8, 317–334. [Google Scholar] [CrossRef] [Green Version]
- Guarnaccia, V.; Crous, P.W. Species of Diaporthe on Camellia and Citrus in the Azores Islands. Phytopathol. Mediterr. 2018, 57, 307–319. [Google Scholar] [CrossRef]
- Chaisiri, C.; Liu, X.Y.; Lin, Y.; Li, J.B.; Xiong, B.; Luo, C.X. Phylogenetic analysis and development of molecular tool for detection of Diaporthe citri causing melanose disease of citrus. Plants 2020, 9, 329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allan-Perkins, E.; Li, D.W.; Schultes, N.; Yavuz, S.; LaMondia, J. The identification of a new species, Diaporthe humulicola, a pathogen causing Diaporthe leaf spot on common hop. Plant Dis. 2020, 104, 2377–2390. [Google Scholar] [CrossRef]
- Hilário, S.; Amaral, I.A.; Gonçalves, M.F.; Lopes, A.; Santos, L.; Alves, A. Diaporthe species associated with twig blight and dieback of Vaccinium corymbosum in Portugal, with description of four new species. Mycologia 2020, 112, 293–308. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Dissanayake, A.J.; Li, X.; Liu, M.; Wanasinghe, D.N.; Xu, J.; Zhao, W.; Zhang, W.; Zhou, Y.; Hyde, K.D.; et al. High genetic diversity and species complexity of Diaporthe associated with grapevine dieback in China. Front. Microbiol. 2019, 10, 1936. [Google Scholar] [CrossRef]
- Li, D.; Zhang, H.; Song, Q.; Liu, J.; Yang, Q.; Luan, F.; Li, D. First report of Diaporthe eres causing branch canker on Cinnamomum camphora (camphor tree) in Jiangxi Province, China. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Guo, Y.S.; Crous, P.W.; Bai, Q.; Fu, M.; Yang, M.M.; Wang, X.H.; Du, Y.M.; Hong, N.; Xu, W.X.; Wang, G.P. High diversity of Diaporthe species associated with pear shoot canker in China. Persoonia 2020, 45, 132–162. [Google Scholar] [CrossRef]
- Muntanola-Cvetkovic, M.; Mihaljcevic, M.; Petrov, M. On the identity of the causative agent of a serious Phomopsis-Diaporthe disease in sunflower plant. Nova Hedwig. 1981, 34, 417–435. [Google Scholar]
- Thompson, S.M.; Tan, Y.P.; Young, A.J.; Neate, S.M.; Aitken, E.A.B.; Shivas, R.G. Stem cankers on sunflower (Helianthus annuus) in Australia reveal a complex of pathogenic Diaporthe (Phomopsis) species. Persoonia 2011, 27, 80–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz, G.A.; Latorre, B.A.; Lolas, M.; Ferrada, E.; Naranjo, P.; Zoffoli, J.P. Identification and characterization of Diaporthe ambigua, D. australafricana, D. novem, and D. rudis causing a postharvest fruit rot in kiwifruit. Plant Dis. 2017, 101, 1402–1410. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Reyne, A.; López-Medrano, F.; Morales, J.M.; Garcia-Esteban, C.; Martin, I.; Eraña, I.; Meije, Y.; Lalueza, A.; Alastruey-Izquierdo, A.; Rodríguez-Tudela, J.L.; et al. Cutaneous infection by Phomopsis longicolla in a renal transplant recipient from Guinea: First report of human infection by this fungus. Transpl. Infect. Dis. 2011, 13, 204–207. [Google Scholar] [CrossRef]
- Mostert, L.; Crous, P.W.; Kang, J.C.; Phillips, A.J. Species of Phomopsis and a Libertella sp. occurring on grapevines with specific reference to South Africa: Morphological, cultural, molecular and pathological characterization. Mycologia 2001, 93, 146–167. [Google Scholar] [CrossRef]
- Mostert, L.; Kang, J.C.; Crous, P.W.; Denman, S. Phomopsis saccharata sp. nov., causing a canker and die-back disease of Protea repens in South Africa. Sydowia 2001, 53, 227–235. [Google Scholar]
- Muralli, T.S.; Suryanarayanan, T.S.; Geeta, R. Endophytic Phomopsis species: Host range and implications for diversity estimates. Can. J. Microbiol. 2006, 52, 673–680. [Google Scholar] [CrossRef]
- Rehner, S.A.; Uecker, F.A. Nuclear ribosomal internal transcribed spacer phylogeny and host diversity in the coelomycete Phomopsis. Can. J. Bot. 1994, 72, 1666–1674. [Google Scholar] [CrossRef]
- Santos, J.M.; Vrandečić, K.; Ćosić, J.; Duvnjak, T.; Phillips, A.J.L. Resolving the Diaporthe species occurring on soybean in Croatia. Persoonia 2011, 27, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udayanga, D.; Liu, X.; McKenzie, E.H.; Chukeatirote, E.; Bahkali, A.H.; Hyde, K.D. The genus Phomopsis: Biology, applications, species concepts and names of common phytopathogens. Fungal Divers. 2011, 50, 189. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.; Crous, P.W.; McKenzie, E.H.; Chukeatirote, E.; Hyde, K.D. A multi-locus phylogenetic evaluation of Diaporthe (Phomopsis). Fungal Divers. 2012, 56, 157–171. [Google Scholar] [CrossRef]
- Van Rensburg, J.C.J.; Lamprecht, S.C.; Groenewald, J.Z.; Castlebury, L.A.; Crous, P.W. Characterisation of Phomopsis spp. associated with die-back of rooibos (Aspalathus linearis) in South Africa. Stud. Mycol. 2006, 55, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossman, A.Y.; Adams, G.C.; Cannon, P.F.; Castlebury, L.A.; Crous, P.W.; Gryzenhout, M.; Jaklitsch, W.M.; Mejia, L.C.; Stoykov, D.; Udayanga, D.; et al. Recommendations of generic names in Diaporthales competing for protection or use. IMA Fungus 2015, 6, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Manawasinghe, I.S.; Huang, Y.; Shu, Y.; Phillips, A.J.L.; Dissanayake, A.J.; Hyde, K.D.; Xiang, M.; Luo, M. Endophytic Diaporthe associated with Citrus grandis cv Tomentosa in China. Front. Microbiol. 2021, 11, 3621. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Camporesi, E.; Hyde, K.D.; Wei, Z.; Yan, J.Y.; Li, X.H. Molecular phylogenetic analysis reveals seven new Diaporthe species from Italy. Mycosphere 2017, 8, 853–877. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Tsai, I.; Thambugala, K.M.; Chuang, W.Y.; Lin, S.R.; Hozzein, W.N.; Cheewangkoon, R. Species diversity of Pleosporalean taxa associated with Camellia sinensis (L.) Kuntze in Taiwan. Sci. Rep. 2020, 10, 12762. [Google Scholar] [CrossRef]
- Tsai, I.; Chung, C.L.; Lin, S.R.; Hung, T.H.; Shen, T.L.; Hu, C.Y.; Hozzein, W.N.; Ariyawansa, H.A. Cryptic diversity, molecular systematics, and pathogenicity of genus Pestalotiopsis and allied genera causing gray blight disease of tea in Taiwan, with a description of a new Pseudopestalotiopsis species. Plant Dis. 2021, 105, 425–443. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Chen, Y.Y.; Liu, J.K.J. Unravelling Diaporthe species associated with woody hosts from karst formations (Guizhou) in China. J. Fungi 2020, 6, 251. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Groenewald, J.Z.; Woodhall, J.; Armengol, J.; Cinelli, T.; Eichmeier, A.; Ezra, D.; Fontaine, F.; Gramaje, D.; Gutierrez-Aguirregabiria, A.; et al. Diaporthe diversity and pathogenicity revealed from a broad survey of grapevine diseases in Europe. Persoonia 2018, 40, 135–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, H.; Zhang, Q.; Hao, Y.Y.; Shao, X.Q.; Wei, X.X.; Hyde, K.D.; Wang, Y.; Zhao, D.G. Diaporthe species in south-western China. MycoKeys 2019, 57, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Summerell, B.A.; Shivas, R.G.; Burgess, T.I.; Decock, C.A.; Dreyer, L.L.; Granke, L.L.; Guest, D.I.; Hardy, G.S.J.; Hausbeck, M.K.; et al. Fungal planet description sheets: 107–127. Persoonia 2012, 28, 138–182. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, I.C.; Crous, P.W.; Groenewald, J.Z.; Maharachchikumbura, S.S.; Jeewon, R.; Phillips, A.J.; Bhat, J.D.; Perera, R.H.; Li, Q.R.; Li, W.J.; et al. Families of Diaporthales based on morphological and phylogenetic evidence. Stud. Mycol. 2017, 86, 217–296. [Google Scholar] [CrossRef]
- Tzean, S.S.; Tzeng, K.C.; Chang, C.A.; Tsay, T.T.; Yen, S.F. List of Plant Diseases in Taiwan; Bureau of Animal and Plant Health Inspection and Quarantine, Council of Agriculture, Executive Yuan: Taipei, Taiwan, 2019. [Google Scholar]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 13 June 2021).
- Dissanayake, A.J.; Liu, M.; Zhang, W.; Chen, Z.; Udayanga, D.; Chukeatirote, E.; Li, X.; Yan, J.; Hyde, K.D. Morphological and molecular characterisation of Diaporthe species associated with grapevine trunk disease in China. Fungal Biol. 2015, 119, 283–294. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Z.B.; Huang, Y.T.; Wang, F.; Hu, W.H.; Dai, L.Y.; Zhong, J.; Liu, Y.; Zhu, J.Z. First report of Diaporthe hongkongensis causing fruit rot on peach (Prunus persica) in China. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Erper, I.; Turkkan, M.; Ozean, M.; Luongo, L.; Belisario, A. Characterization of Diaporthe hongkongensis species causing stem-end rot on kiwifruit in Turkey. J. Plant Pathol. 2017, 99, 779–782. [Google Scholar]
- Kee, Y.J.; Wong, K.L.; Zakaria, L.; Mohd, M.H. Diaporthe species causing stem gray blight of red-fleshed dragon fruit (Hylocereus polyrhizus) in Malaysia. Sci. Rep. 2021, 11, 3907. [Google Scholar] [CrossRef]
- Li, H.; Yu, S.; Tang, W.; Miao, M.; Liu, Y. First report of Diaporthe passiflorae and Diaporthe nobilis causing a postharvest kiwifruit rot in Sichuan province, China. Plant Dis. 2019, 103, 771. [Google Scholar] [CrossRef]
- Lim, L.; Mohd, M.H.; Zakaria, L. Identification and pathogenicity of Diaporthe species associated with stem-end rot of mango (Mangifera indica L.). Eur. J. Plant Pathol. 2019, 155, 687–696. [Google Scholar] [CrossRef]
- Bai, Q.; Wang, G.P.; Hong, N.; Guo, Y.S.; Fu, M. First report of Diaporthe tulliensis and Diaporthe actinidiae causing kiwifruit stem canker in Hubei and Anhui provinces, China. Plant Dis. 2017, 101, 508–509. [Google Scholar] [CrossRef]
- Gong, J.L.; Lu, Y.; Wu, W.H.; He, C.P.; Liang, Y.Q.; Huang, X.; Zheng, J.L.; Xi, J.G.; Tang, S.B.; Yi, K.X. First report of Phomopsis heveicola (anamorph of Diaporthe tulliensis) causing leaf blight of coffee (Coffea arabica) in China. Plant Dis. 2020, 104, 570. [Google Scholar] [CrossRef]
- Huang, C.C.; Liu, H.H.; Wu, P.H.; Chang, H.X. First report of leaf spot caused by Diaporthe tulliensis on Boston ivy (Parthenocissus tricuspidata) in Taiwan. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Chen, K.L.; Kirschner, R. Fungi from leaves of lotus (Nelumbo nucifera). Mycol. Prog. 2018, 17, 275–293. [Google Scholar] [CrossRef]
- Arciuolo, R.; Camardo Leggieri, M.; Chiusa, G.; Castello, G.; Genova, G.; Spigolon, N.; Battilani, P. Ecology of Diaporthe eres, the causal agent of hazelnut defects. PLoS ONE 2021, 16, e0247563. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Jiang, N.; Tian, C.M. Three new Diaporthe species from Shaanxi Province, China. MycoKeys 2020, 67, 1–18. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Phillips, A.J.L.; Hyde, K.D.; Yan, J.Y.; Li, X.H. The current status of species in Diaporthe. Mycosphere 2017, 8, 1106–1156. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, F.; Duan, W.; Crous, P.W.; Cai, L. Diaporthe is paraphyletic. IMA Fungus 2017, 8, 153–187. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J. MrModeltest v2. Program; Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (and Other Methods), Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Silvestro, D.; Michalak, I. RaxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A. FigTree: Tree Figure Drawing Tool, Version 1.2.2; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, Scotland, 2008. [Google Scholar]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Dettman, J.R.; Jacobson, D.J.; Turner, E.; Pringle, A.; Taylor, J.W. Reproductive isolation and phylogenetic divergence in Neurospora: Comparing methods of species recognition in a model eukaryote. Evolution 2003, 57, 2721–2741. [Google Scholar] [CrossRef]
- Huang, F.; Hou, X.; Dewdney, M.M.; Fu, Y.; Chen, G.; Hyde, K.D.; Li, H. Diaporthe species occurring on citrus in China. Fungal Divers. 2013, 61, 237–250. [Google Scholar] [CrossRef]
- Keith, L.M.; Velasquez, M.E.; Zee, F.T. Identification and characterization of Pestalotiopsis spp. causing scab disease of guava, Psidium guajava, in Hawaii. Plant Dis. 2006, 90, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abubakar, A.; Suberu, H.A.; Bello, I.M.; Abdulkadir, R.; Daudu, O.A.; Lateef, A.A. Effect of pH on mycelial growth and sporulation of Aspergillus parasiticus. J. Plant Sci. 2013, 1, 64–67. [Google Scholar] [CrossRef]
- Saha, A.; Mandal, P.; Dasgupta, S.; Saha, D. Influence of culture media and environmental factors on mycelial growth and sporulation of Lasiodiplodia theobromae (Pat.) Griffon and Maubl. J. Environ. Biol. 2008, 29, 407–410. [Google Scholar]
- Wang, Y.; Xiong, F.; Lu, Q.; Hao, X.; Zheng, M.; Wang, L.; Li, N.; Ding, C.; Wang, X.; Yang, Y. Diversity of Pestalotiopsis-like species causing gray blight disease of tea plants (Camellia sinensis) in China, including two novel Pestalotiopsis species, and analysis of their pathogenicity. Plant Dis. 2019, 103, 2548–2558. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Primers | Sequence (5′→3′) | Reference |
|---|---|---|---|
| ITS | ITS5 ITS4 | GGA AGT AAA AGT CGT AAC AAG G TCC GCT TAT TGA TAT GC | [52] |
| tef1-α | EF1-728F EF1-986R | CAT CGA GAA GTT CGA GAA GG TAC TTG AAG GAA CCC TTA CC | [53] |
| tub2 | Bt2a Bt2b | GGT AAC CAA ATC GGT GCT TTC ACC CTC AGT GTA GTG ACC CTT GGC | [54] |
| cal | CAL-228F CAL-737R | GAG TTC AAG GAG GCC TTC TCC C CAT CTT TCT GGC CAT GG | [53] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ariyawansa, H.A.; Tsai, I.; Wang, J.-Y.; Withee, P.; Tanjira, M.; Lin, S.-R.; Suwannarach, N.; Kumla, J.; Elgorban, A.M.; Cheewangkoon, R. Molecular Phylogenetic Diversity and Biological Characterization of Diaporthe Species Associated with Leaf Spots of Camellia sinensis in Taiwan. Plants 2021, 10, 1434. https://doi.org/10.3390/plants10071434
Ariyawansa HA, Tsai I, Wang J-Y, Withee P, Tanjira M, Lin S-R, Suwannarach N, Kumla J, Elgorban AM, Cheewangkoon R. Molecular Phylogenetic Diversity and Biological Characterization of Diaporthe Species Associated with Leaf Spots of Camellia sinensis in Taiwan. Plants. 2021; 10(7):1434. https://doi.org/10.3390/plants10071434
Chicago/Turabian StyleAriyawansa, Hiran A., Ichen Tsai, Jian-Yuan Wang, Patchareeya Withee, Medsaii Tanjira, Shiou-Ruei Lin, Nakarin Suwannarach, Jaturong Kumla, Abdallah M. Elgorban, and Ratchadawan Cheewangkoon. 2021. "Molecular Phylogenetic Diversity and Biological Characterization of Diaporthe Species Associated with Leaf Spots of Camellia sinensis in Taiwan" Plants 10, no. 7: 1434. https://doi.org/10.3390/plants10071434