Morphological Characterization and Genetic Diversity of Rice Blast Fungus, Pyricularia oryzae, from Thailand Using ISSR and SRAP Markers


Abstract
:1. Introduction
2. Materials and Methods
2.1. Rice Blast Materials
2.2. Fungal Morphology Characterization
2.3. ITS Amplification and Sequencing
2.4. ISSR Analysis
2.5. SRAP Analysis
2.6. Data Analysis
3. Results
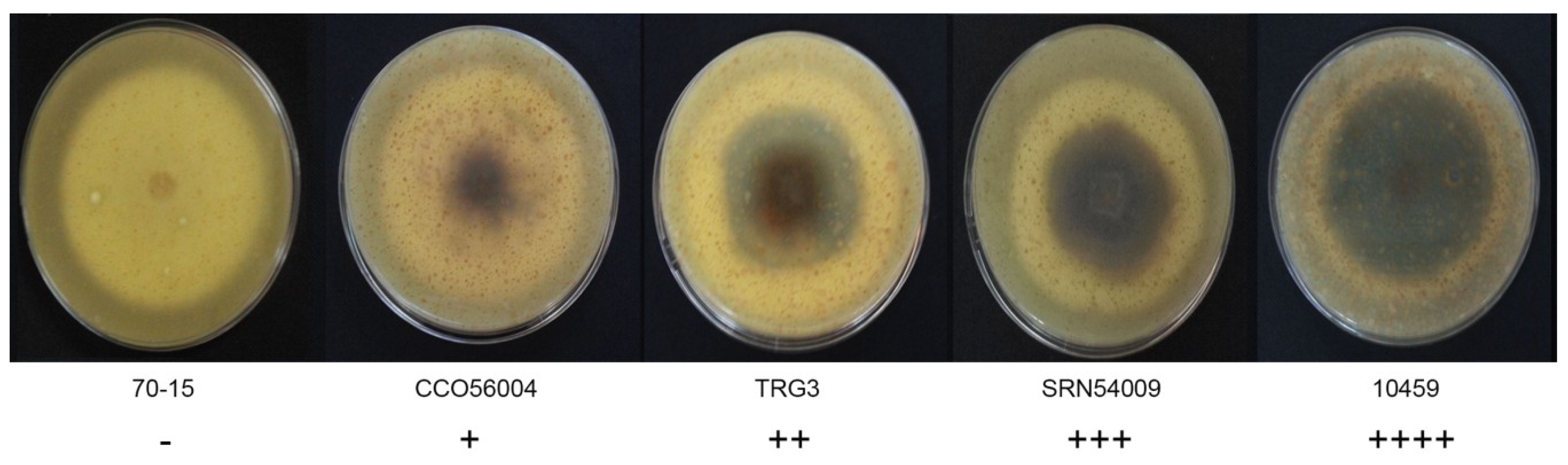
3.1. Fungal Morphology Variation
3.2. ITS Sequence Analysis
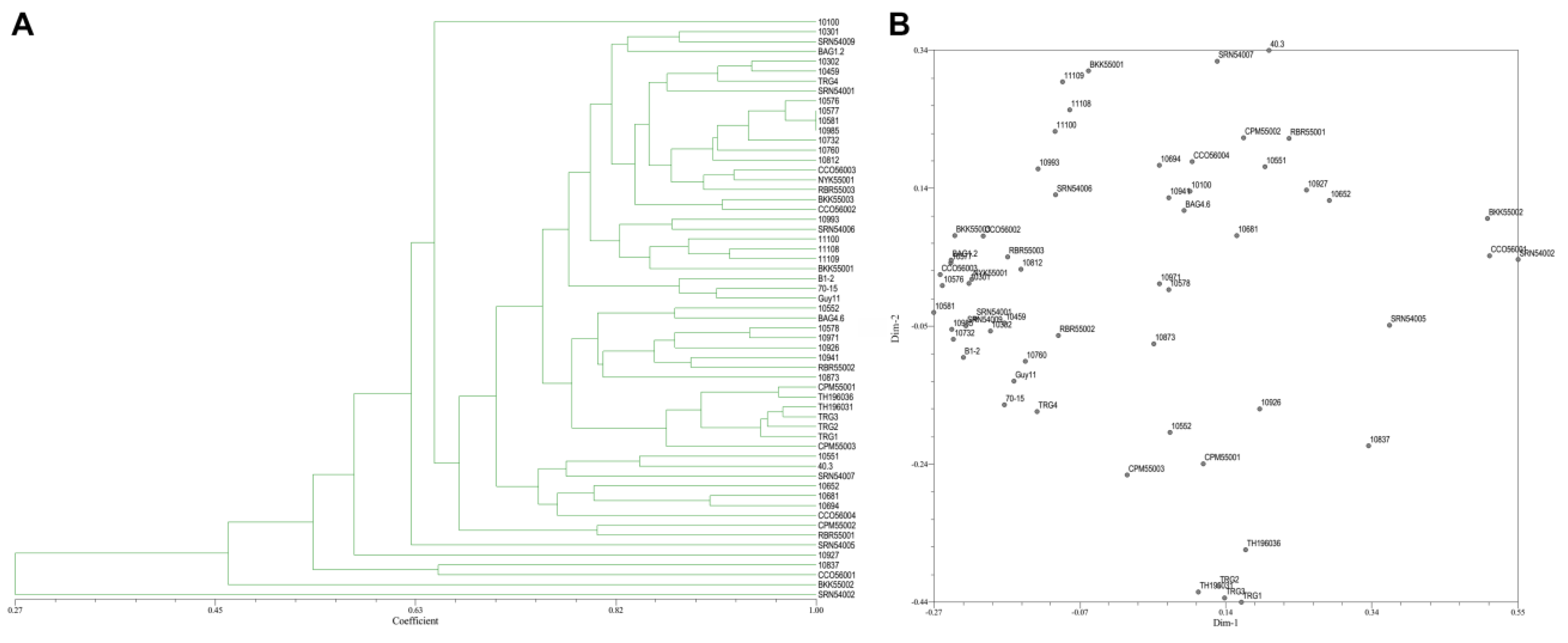
3.3. Genetic Polymorphism from ISSR Marker
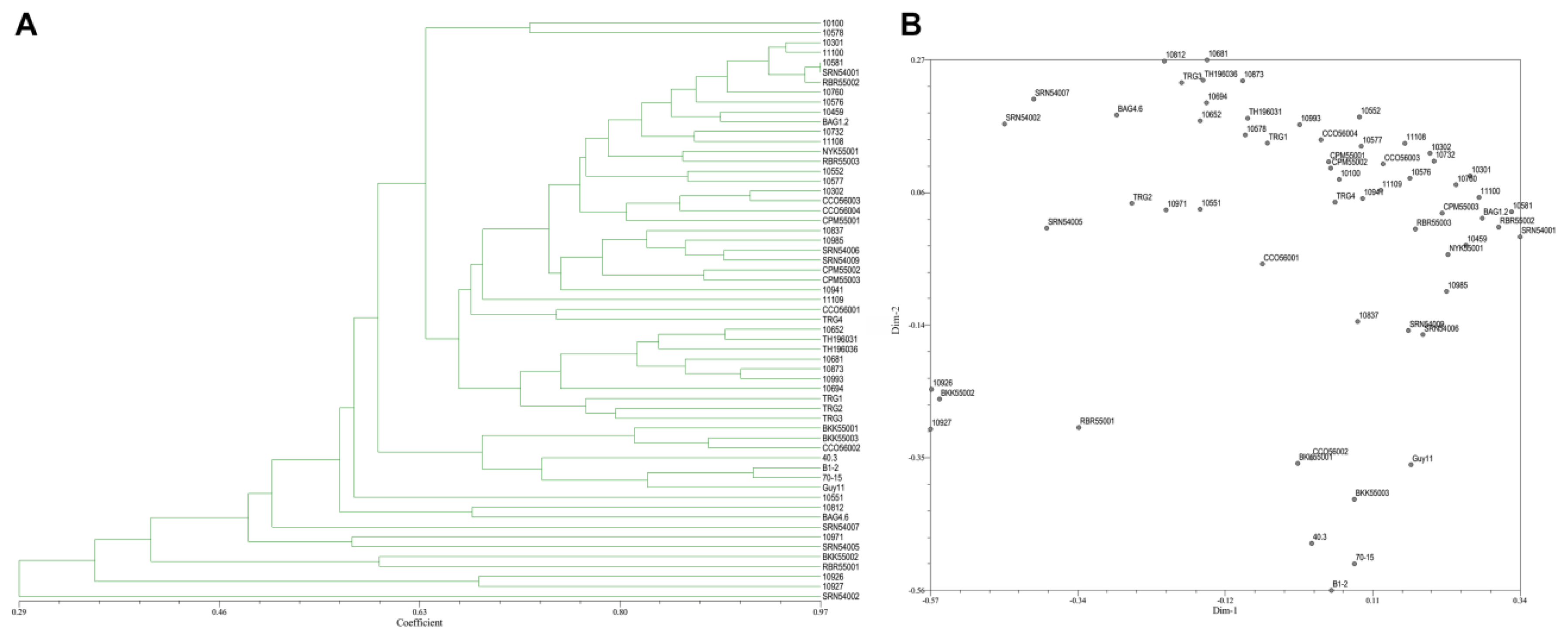
3.4. Genetic Polymorphism from SRAP Marker
3.5. Genetic Polymorphism from ISSR and SRAP Markers Combination
4. Discussion
4.1. Morphological Characteristics of Rice Blast Fungus Strains in Thailand are Diverse
4.2. Genetic Diversity of Pyricularia oryzae Using SRAP and ISSR Markers
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Langner, T.; Białas, A.; Kamoun, S. The blast fungus decoded: Genomes in flux. MBio 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, R.; Pereira, G.; Garrido, I.; Tavares-de-Sousa, M.M.; Espinosa, F. Comparison of RAPD, ISSR, and AFLP molecular markers to reveal and classify Orchardgrass (Dactylis glomerata L.) germplasm variations. PLoS ONE 2016, 11, e0152972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robarts, D.W.H.; Wolfe, A.D. Sequence-related amplified polymorphism (SRAP) markers: A potential resource for studies in plant molecular biology. Appl. Plant Sci. 2014, 2. [Google Scholar] [CrossRef]
- Li, G.; Quiros, C. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: Its application to mapping and gene tagging in Brassica. Theor. Appl. Genet. 2001, 103, 455–461. [Google Scholar] [CrossRef]
- Agarwal, M.; Shrivastava, N.; Padh, H. Advances in molecular marker techniques and their applications in plant sciences. Plant Cell Rep. 2008, 27, 617–631. [Google Scholar] [CrossRef]
- Longya, A.; Chaipanya, C.; Franceschetti, M.; Maidment, H.R.J.; Banfield, J.M.; Jantasuriyarat, C. Gene duplication and mutation in the emergence of a novel aggressive allele of the AVR-Pik effector in the rice blast fungus. Mol. Plant Microbe Interact. 2019, 32, 740–749. [Google Scholar] [CrossRef]
- Roldán-Ruiz, I.; Dendauw, J.; Van-Bockstaele, E.; Depicker, A.; De-Loose, M. AFLP markers reveal high polymorphic rates in ryegrasses (Lolium spp.). Mol. Breed. 2000, 6, 125–134. [Google Scholar] [CrossRef]
- Sonah, H.; Deshmukh, R.K.; Parida, S.K.; Chand, S.; Kotasthane, A.S. Morphological and genetic variation among different isolates of Magnaporthe grisea collected from Chhattisgarh. Indian Phytopathol. 2009, 62, 469–477. [Google Scholar]
- Suzuki, H. Studies on the behaviour of the rice blast fungus spore and the application for forecasting method of the rice blast disease. Bull 1969, 10, 114–118. [Google Scholar]
- Gaddeyya, G.; Niharika, P.S.; Bharathi, P.; Kumar, P.K.R. Isolation and identification of soil mycoflora in different crop fields at Salur Mandal. Adv. Appl. Sci. Res. 2012, 3, 2020–2026. [Google Scholar]
- Moriyama, H.; Urayama, S.I.; Higashiura, T.; Le, T.M.; Komatsu, K. Chrysoviruses in Magnaporthe oryzae. Viruses 2018, 10, 697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirata, K.; Kusaba, M.; Chuma, I.; Osue, J.; Nakayashiki, H.; Mayama, S.H.; Tosa, Y. Speciation in Pyricularia inferred from multilocus phylogenetic analysis. Mycol. Res. 2007, 111 Pt 7, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Abed-Ashtiani, F.; Kadir, J.; Nasehi, A.; Hashemian-Rahaghi, S.; Vadamalai, G.; Rambe, S. Characterisation of Magnaporthe oryzae isolates from rice in peninsular Malaysia. Czech J. Genet. Plant Breed. 2016, 52, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Luana, B.; Antonella, A.; Deborah, A.; Emanuela, P.; Lucia, P.; Chiara, G.; Vilberto, S. A new pair of primers designed for amplification of the ITS region in Tuber species. FEMS Microbiol. Lett. 1999, 173, 239–245. [Google Scholar] [CrossRef] [Green Version]
- McDonald, B.A.; McDermott, J.M.; Goodwin, S.B. The population biology of host pathogen interactions. Annu. Rev. Phytopathol. 1989, 27, 77–94. [Google Scholar] [CrossRef]
- Ngernmuen, A.; Suktrakul, W.; Damchuay, K.; Longya, A.; Katengam, S.; Jantasuriyarat, C. Substantial enhancement of high polymorphic SSR marker development using in silico method from 18 available rice blast fungus genome sequences and its application in genetic diversity assessment. Biologia 2019, 74, 1181–1189. [Google Scholar] [CrossRef]
- Silva, G.; Prabhu, A.; Filippi, M.; Trindade, M.; Araújo, L.; Zambolim, L. Genetic and phenotypic diversity of Magnaporthe oryzae from leaves and panicles of rice in commercial fields in the State of Goiás, Brazil. Trop. Plant Pathol. 2009, 34, 77–86. [Google Scholar] [CrossRef]
- Xu, X.; Yang, W.; Tian, K.; Zheng, J.; Liu, X.; Li, K.; Lu, W.; Tan, Y.; Qin, Y.; Wang, C. Genetic diversity and pathogenicity dynamics of Magnaporthe oryzae in the Wuling Mountain area of China. Eur. J. Plant Pathol. 2018, 153, 731–742. [Google Scholar] [CrossRef]
- Noguchi, M.T. Parasexual recombination in Magnaporthe oryzae. Jpn. Agric. Res. Q. 2011, 45, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Chuma, I.; Isobe, C.; Hotta, Y.; Ibaragi, K.; Futamata, N.; Kusaba, M.; Yoshida, K.; Terauchi, R.; Fujita, Y.; Nakayashiki, H.; et al. Multiple translocation of the AVR-Pita effector gene among chromosomes of the rice blast fungus Magnaporthe oryzae and related species. PLoS Pathog. 2011, 7, e1002147. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Origin (a) | Year | Mycelium Features (b) | Melanin Pigment (c) | Conidia Shape (d) | ITS (e) |
|---|---|---|---|---|---|---|
| 10100 | Srakaew | 2006 | Cream, Fluffy, Densely | + | C | MT126183 |
| 10301 | Sisaket | 2006 | White, Flat, Densely | ++++ | C | MT126184 |
| 10302 | Sisaket | 2006 | White, Flat, Densely | + | B | MT126185 |
| 10459 | Lampang | 2006 | Grey, Flat, Densely | ++++ | C | MT126186 |
| 10551 | Nakhon Ratchasima | 2006 | White, Flat, Densely | ++++ | C | MT126187 |
| 10552 | Nakhon Ratchasima | 2006 | White, Fluffy, Densely | + | C | MT126188 |
| 10576 | Kalasin | 2006 | Cream, Fluffy, Densely | ++++ | C | MT126189 |
| 10577 | Nakhon Ratchasima | 2006 | White, Flat, Densely | - | D | MT126190 |
| 10578 | Nakhon Ratchasima | 2006 | Cream, Flat, Densely | + | C | MT126191 |
| 10581 | Nakhon Ratchasima | 2006 | White, Fluffy, Densely | +++ | C | MT126192 |
| 10652 | Maha Sarakham | 2006 | Cream, Fluffy, Densely | +++ | C | MT126193 |
| 10681 | Roi Et | 2006 | White, Flat, Densely | ++ | B | MT126194 |
| 10694 | Nong Khai | 2006 | Cream, Flat, Densely | + | B | MT126195 |
| 10732 | Kamphaeng Phet | 2006 | White, Flat, Densely | - | C | MT126196 |
| 10760 | Kamphaeng Phet | 2006 | White, Fluffy, Densely | - | D | MT126197 |
| 10812 | Chiang Rai | 2006 | White, Flat, Densely | - | D | MT126198 |
| 10837 | Surin | 2006 | White, Flat, Densely | ++ | B | MT126199 |
| 10873 | Mae Hong Son | 2006 | Cream, Fluffy, Highly dense | ++ | C | MT126200 |
| 10926 | Phayao | 2006 | White, Flat, Densely | - | C | MT126201 |
| 10927 | Phayao | 2006 | White, Flat, Densely | - | C | MT126202 |
| 10941 | Lampang | 2006 | White, Flat, Densely | - | C | MT126203 |
| 10971 | Nan | 2006 | White, Flat, Densely | - | C | MT126204 |
| 10985 | Sisaket | 2006 | Cream, Flat, Densely | + | C | MT126205 |
| 10993 | Buriram | 2006 | White, Fluffy, Densely | + | C | MT126206 |
| 11100 | Udon Thani | 2006 | White, Fluffy, Densely | + | C | MT126207 |
| 11108 | Ubon Ratchathani | 2006 | White, Flat, Densely | ++++ | C | MT126208 |
| 11109 | Ubon Ratchathani | 2006 | Cream, Fluffy, Densely | + | C | MT126209 |
| BAG1.2 | Phitsanulok | 2006 | White, Fluffy, Densely | + | C | MT126210 |
| BAG4.6 | Phitsanulok | 2006 | White, Flat, Densely | +++ | D | MT126211 |
| 40.3 | n/a | n/a | Grey, Fluffy, Densely | ++ | C | MT126212 |
| SRN54001 | Surin | 2011 | White, Flat, Densely | +++ | D | MT126213 |
| SRN54002 | Surin | 2011 | White, Fluffy, Densely | ++ | D | MT126214 |
| SRN54005 | Surin | 2011 | White, Flat, Densely | ++ | D | MT126215 |
| SRN54006 | Surin | 2011 | White, Fluffy, Densely | ++ | C | MT126216 |
| SRN54007 | Surin | 2011 | Cream, Flat, Densely | ++++ | C | MT126217 |
| SRN54009 | Surin | 2011 | White, Flat, Densely | +++ | C | MT126218 |
| CCO56001 | Chachoengsao | 2013 | Cream, Fluffy, Densely | ++ | C | MT126219 |
| CCO56002 | Chachoengsao | 2013 | White, Flat, Densely | ++ | C | MT126220 |
| CCO56003 | Chachoengsao | 2013 | Cream, Fluffy, Densely | ++++ | C | MT126221 |
| CCO56004 | Chachoengsao | 2013 | White, Fluffy, Densely | + | C | MT126222 |
| CPM55001 | Chaiyaphum | 2012 | Cream, Flat, Densely | + | C | MT126223 |
| CPM55002 | Chaiyaphum | 2012 | Grey, Flat, Densely | ++++ | C | MT126224 |
| CPM55003 | Chaiyaphum | 2012 | White, Fluffy, Densely | ++++ | C | MT126225 |
| NYK55001 | Nakhon Nayok | 2011 | Grey, Flat, Densely | ++ | C | MT126226 |
| BKK55001 | Bangkok | 2011 | White, Fluffy, Densely | +++ | C | MT126227 |
| BKK55002 | Bangkok | 2011 | White, Flat, Densely | + | D | MT126228 |
| BKK55003 | Bangkok | 2011 | Grey, Fluffy, Densely | +++ | C | MT126229 |
| RBR55001 | Ratchaburi | 2011 | Cream, Fluffy, Highly dense | ++++ | A | MT126230 |
| RBR55002 | Ratchaburi | 2011 | White, Flat, Densely | - | C | MT126231 |
| RBR55003 | Ratchaburi | 2011 | White, Fluffy, Densely | ++ | C | MT126232 |
| B1-2 | Ubon Ratchathani | 2001 | White, Fluffy, Densely | ++++ | C | MT126233 |
| TH196031 | Ubon Ratchathani | 2001 | White, Fluffy, Densely | ++ | C | MT126234 |
| TH196036 | Ubon Ratchathani | 2001 | White, Fluffy, Densely | ++ | C | MT126235 |
| TRG1 | Nong Khai | 2013 | Grey, Fluffy, Densely | + | C | MT126236 |
| TRG2 | Nong Khai | 2013 | White, Flat, Densely | - | C | MT126237 |
| TRG3 | Nong Khai | 2013 | Grey, Fluffy, Densely | ++ | C | MT126238 |
| TRG4 | Nong Khai | 2013 | Cream, Flat, Densely | - | C | MT126239 |
| 70-15 | Laboratory strain | 1988 | Grey, Flat, Densely | - | C | MT126240 |
| Guy11 | French Guyana | 1988 | Grey, Fluffy, Densely | - | C | MT126241 |
| Primer ID | Primer Name (a) | Total Bands | Polymorphic Bands | % Polymorphic | PIC (b) |
|---|---|---|---|---|---|
| ISSR1 | UBC-807 | 12 | 4 | 33.33 | 0.39 |
| ISSR2 | UBC-808 | 14 | 8 | 57.14 | 0.15 |
| ISSR3 | UBC-809 | 8 | 2 | 25.00 | 0.49 |
| ISSR4 | UBC-811 | 8 | 3 | 37.50 | 0.16 |
| ISSR5 | UBC-812 | 11 | 4 | 36.36 | 0.49 |
| ISSR6 | UBC-813 | 3 | 3 | 100.0 | 0.49 |
| ISSR7 | UBC-814 | 0 | |||
| ISSR8 | UBC-815 | 0 | |||
| ISSR9 | UBC-817 | 6 | 2 | 33.33 | 0.26 |
| ISSR10 | UBC-818 | 5 | 1 | 20.00 | 0.38 |
| ISSR11 | UBC-820 | 0 | |||
| ISSR12 | UBC-823 | 2 | 2 | 100.0 | 0.32 |
| ISSR13 | UBC-824 | 0 | |||
| ISSR14 | UBC-825 | 8 | 4 | 50.00 | 0.50 |
| ISSR15 | UBC-826 | 10 | 9 | 90.00 | 0.50 |
| ISSR16 | UBC-827 | 0 | |||
| ISSR17 | UBC-841 | 16 | 14 | 87.50 | 0.50 |
| ISSR18 | UBC-847 | 4 | 1 | 25.00 | 0.07 |
| ISSR19 | UBC-848 | 11 | 5 | 45.45 | 0.33 |
| ISSR20 | UBC-849 | 0 | |||
| Total | 118 | 62 | |||
| Average | 8.43 | 4.43 | 52.90 | 0.36 |
| Primer ID | Primer Names (a) | Total Bands | Polymorphic Bands | % Polymorphic | PIC (b) |
|---|---|---|---|---|---|
| SRAP1 | me1/em1 | 5 | 4 | 80.00 | 0.50 |
| SRAP2 | me1/em2 | 0 | |||
| SRAP3 | me1/em3 | 5 | 5 | 100.0 | 0.49 |
| SRAP4 | me1/em4 | 0 | |||
| SRAP5 | me1/em5 | 9 | 6 | 66.67 | 0.47 |
| SRAP6 | me1/em6 | 10 | 5 | 50.00 | 0.50 |
| SRAP7 | me2/em1 | 0 | |||
| SRAP8 | me2/em2 | 11 | 4 | 36.36 | 0.50 |
| SRAP9 | me2/em3 | 0 | |||
| SRAP10 | me2/em4 | 14 | 6 | 42.86 | 0.48 |
| SRAP11 | me2/em5 | 1 | 1 | 100.0 | 0.32 |
| SRAP12 | me2/em6 | 9 | 3 | 33.33 | 0.40 |
| SRAP13 | me3/em1 | 6 | 5 | 83.33 | 0.37 |
| SRAP14 | me3/em2 | 6 | 4 | 66.67 | 0.49 |
| SRAP15 | me3/em3 | 6 | 5 | 83.33 | 0.44 |
| SRAP16 | me3/em4 | 3 | 2 | 66.67 | 0.41 |
| SRAP17 | me3/em5 | 0 | |||
| SRAP18 | me3/em6 | 6 | 4 | 66.67 | 0.38 |
| SRAP19 | me4/em1 | 2 | 2 | 100.0 | 0.39 |
| SRAP20 | me4/em2 | 5 | 1 | 20.00 | 0.03 |
| SRAP21 | me4/em3 | 0 | |||
| SRAP22 | me4/em4 | 0 | |||
| SRAP23 | me4/em5 | 0 | |||
| SRAP24 | me4/em6 | 2 | 2 | 100.0 | 0.50 |
| SRAP25 | me5/em1 | 0 | |||
| SRAP26 | me5/em2 | 0 | |||
| SRAP27 | me5/em3 | 0 | |||
| SRAP28 | me5/em4 | 1 | 1 | 100.0 | 0.34 |
| SRAP29 | me5/em5 | 2 | 1 | 50.00 | 0.03 |
| SRAP30 | me5/em6 | 0 | |||
| Total | 103 | 61 | |||
| Average | 5.72 | 3.39 | 69.22 | 0.39 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Longya, A.; Talumphai, S.; Jantasuriyarat, C. Morphological Characterization and Genetic Diversity of Rice Blast Fungus, Pyricularia oryzae, from Thailand Using ISSR and SRAP Markers. J. Fungi 2020, 6, 38. https://doi.org/10.3390/jof6010038
Longya A, Talumphai S, Jantasuriyarat C. Morphological Characterization and Genetic Diversity of Rice Blast Fungus, Pyricularia oryzae, from Thailand Using ISSR and SRAP Markers. Journal of Fungi. 2020; 6(1):38. https://doi.org/10.3390/jof6010038
Chicago/Turabian StyleLongya, Apinya, Sucheela Talumphai, and Chatchawan Jantasuriyarat. 2020. "Morphological Characterization and Genetic Diversity of Rice Blast Fungus, Pyricularia oryzae, from Thailand Using ISSR and SRAP Markers" Journal of Fungi 6, no. 1: 38. https://doi.org/10.3390/jof6010038