Low Efficacy of Isaria fumosorosea against Box Tree Moth Cydalima perspectalis: Are Host Plant Phytochemicals Involved in Herbivore Defence against Fungal Pathogens?


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Entomopathogenic Fungus
2.3. Bioassays
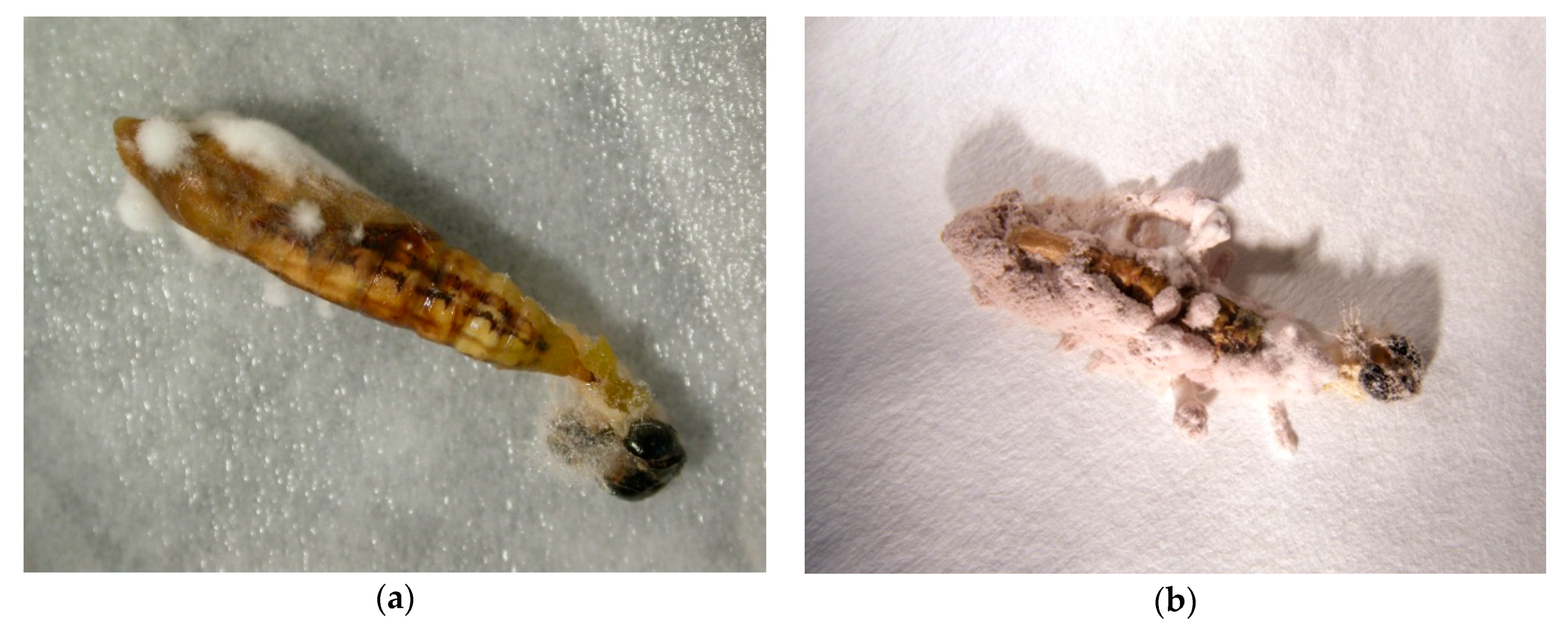
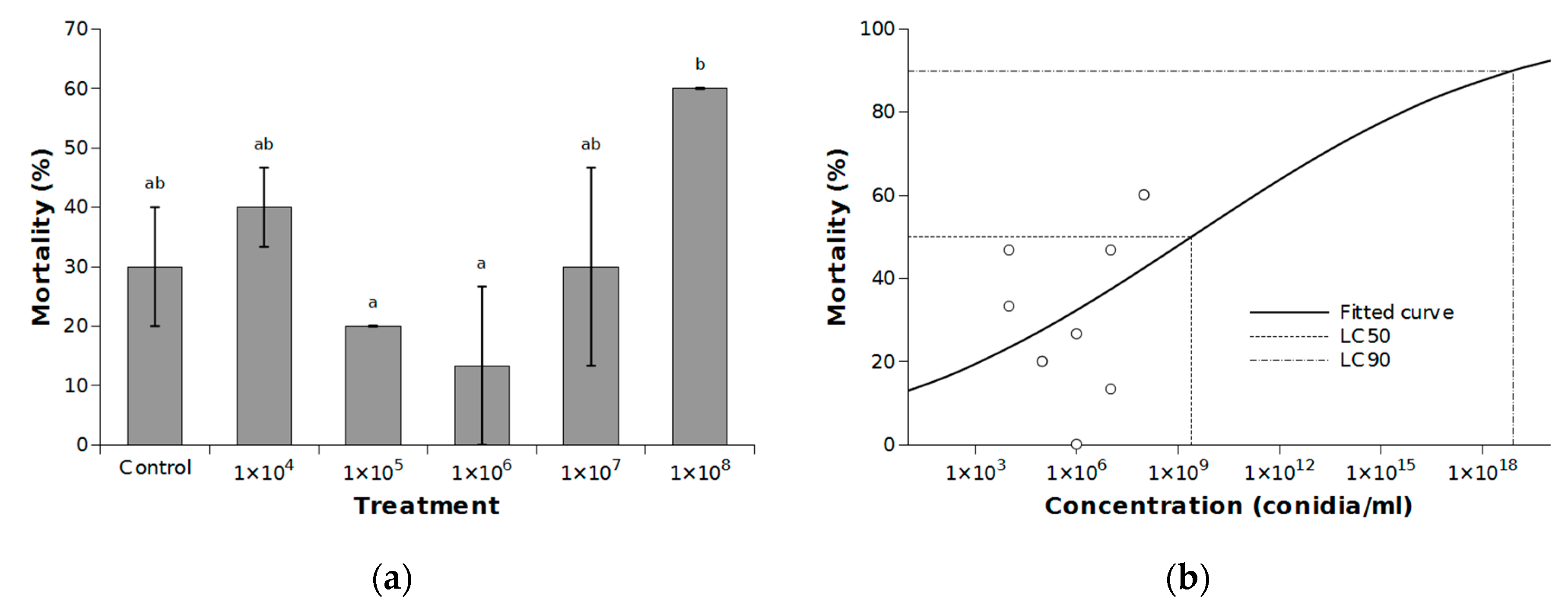
2.3.1. The Efficacy of I. fumosorosea against C. perspectalis
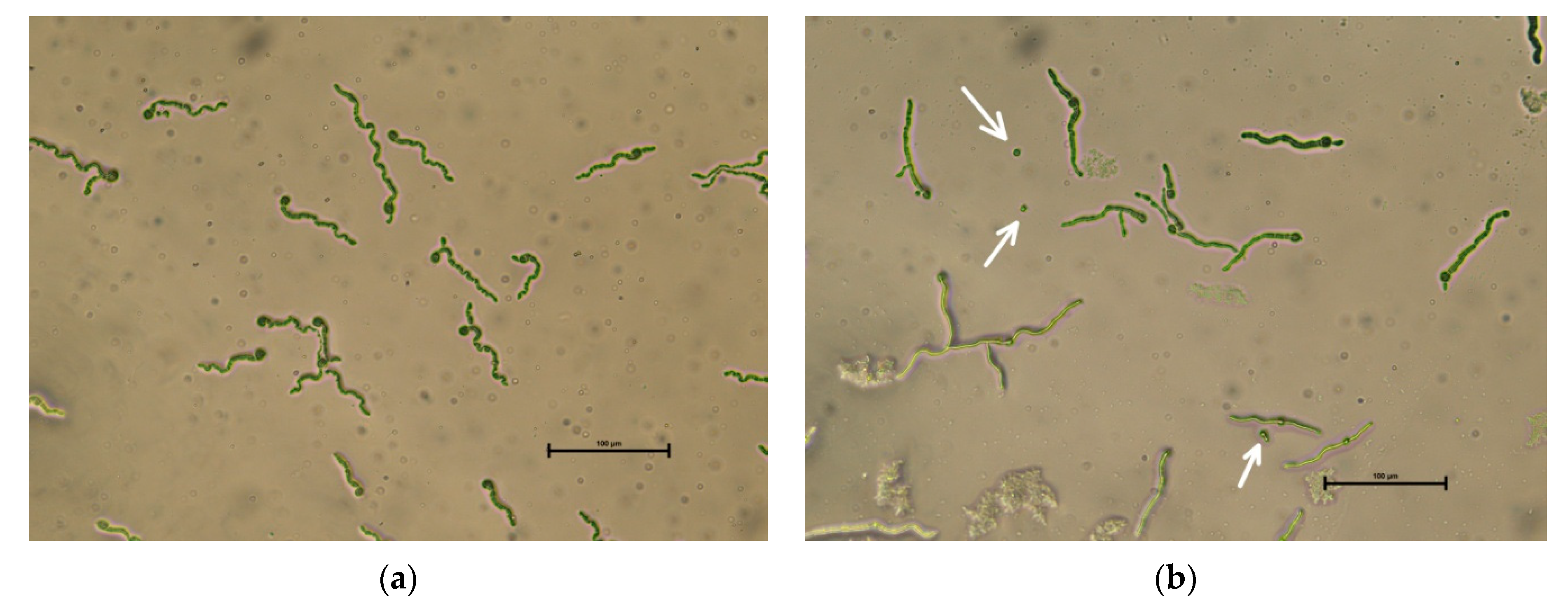
2.3.2. Scanning Electron Miscroscopy of I. fumosorosea Conidia Germination on Cuticle of C. perspectalis Larvae
2.3.3. The Effect of B. sempervirens Extract on I. fumosorosea Germination and Growth
2.4. Data Analysis
3. Results and Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Di Domenico, F.; Lucchese, F.; Magri, D. Buxus in Europe: Late Quaternary dynamics and modern vulnerability. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 354–362. [Google Scholar] [CrossRef]
- Mitchell, R.; Chitanava, S.; Dbar, R.; Kramarets, V.; Lehtijärvi, A.; Matchutadze, I.; Mamadashvili, G.; Matsiakh, I.; Nacambo, S.; Papazova-Anakieva, I.; et al. Identifying the ecological and societal consequences of a decline in Buxus forests in Europe and the Caucasus. Biol. Invasions 2018, 20, 3605–3620. [Google Scholar] [CrossRef]
- Bras, A.; Avtzis, D.N.; Kenis, M.; Li, H.; Vétek, G.; Bernard, A.; Courtin, C.; Rousselet, J.; Roques, A.; Auger-Rozenberg, M.-A. A complex invasion story underlies the fast spread of the invasive box tree moth (Cydalima perspectalis) across Europe. J. Pest Sci. 2019, 92, 1187–1202. [Google Scholar] [CrossRef]
- Wan, H.; Haye, T.; Kenis, M.; Nacambo, S.; Xu, H.; Zhang, F.; Li, H. Biology and natural enemies of Cydalima perspectalis in Asia: Is there biological control potential in Europe? J. Appl. Entomol. 2014, 138, 715–722. [Google Scholar] [CrossRef]
- Nacambo, S.; Leuthardt, F.L.G.; Wan, H.; Li, H.; Haye, T.; Baur, B.; Weiss, R.M.; Kenis, M. Development characteristics of the box-tree moth Cydalima perspectalis and its potential distribution in Europe. J. Appl. Entomol. 2014, 138, 14–26. [Google Scholar] [CrossRef]
- Suppo, C.; Bras, A.; Robinet, C. A temperature- and photoperiod-driven model reveals complex temporal population dynamics of the invasive box tree moth in Europe. Ecol. Model. 2020, 432, 109229. [Google Scholar] [CrossRef]
- Göttig, S.; Herz, A. Are egg parasitoids of the genus Trichogramma (Hymenoptera: Trichogrammatidae) promising biological control agents for regulating the invasive Box tree pyralid, Cydalima perspectalis (Lepidoptera: Crambidae)? Biocontrol Sci. Technol. 2016, 26, 1471–1488. [Google Scholar] [CrossRef]
- Martini, A.; Di Vitantonio, C.; Dindo, M.L. Acceptance and suitability of the box tree moth Cydalima perspectalis as host for the tachinid parasitoid Exorista larvarum. Bull. Insectol. 2019, 72, 150–160. [Google Scholar]
- Alkan Akncı, H.; Kurdoğlu, O. Damage level of Cydalima perspectalis (Lepidoptera: Crambidae) on naturally growing and ornamental box populations in Artvin, Turkey. Kastamonu Üniversitesi Orman Fakültesi Derg. 2019. [Google Scholar] [CrossRef] [Green Version]
- Fora, C.G.; Sasu, L.; Poşta, D.; Berac, C. Chemical possibilities of Cydalima perspectalis Walk. (Lepidoptera: Crambidae) control. J. Hortic. For. Biotechnol. 2016, 20, 31–34. [Google Scholar]
- Göttig, S.; Herz, A. Susceptibility of the Box tree pyralid Cydalima perspectalis Walker (Lepidoptera: Crambidae) to potential biological control agents Neem (NeemAzal®-T/S) and entomopathogenic nematodes (Nemastar®) assessed in laboratory bioassays and field trials. J. Plant Dis. Prot. 2018, 125, 365–375. [Google Scholar] [CrossRef]
- Shahid, A.; Rao, Q.; Bakhsh, A.; Husnain, T. Entomopathogenic fungi as biological controllers: New insights into their virulence and pathogenicity. Arch. Biol. Sci. 2012, 64, 21–42. [Google Scholar] [CrossRef]
- de Faria, M.R.; Wraight, S.P. Mycoinsecticides and mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- SangMyeong, L.; DongWoon, L.; HoYul, C.; JiWoong, P. Pathogenicities of Beauveria bassiana GY1-17 against some agro-forest insect pests. Korean J. Appl. Entomol. 1997, 36, 351–356. [Google Scholar]
- Smith, P. Control of Bemisia tabaci and the potential of Paecilomyces fumosoroseus as a biopesticide. Biocontrol News Inf. 1993, 14, 71N–78N. [Google Scholar]
- Dunlap, C.A.; Jackson, M.A.; Wright, M.S. A foam formulation of Palecilomyces fumosoroseus, an entomopathogenic biocontrol agent. Biocontrol Sci. Technol. 2007, 17, 513–523. [Google Scholar] [CrossRef]
- Hoy, M.A.; Singh, R.; Rogers, M.E. Evaluations of a novel isolate of Isaria fumosorosea for control of the asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Fla. Entomol. 2010, 93, 24–32. [Google Scholar] [CrossRef]
- Kim, J.S.; Je, Y.H.; Roh, J.Y. Production of thermotolerant entomopathogenic Isaria fumosorosea SFP-198 conidia in corn-corn oil mixture. J. Ind. Microbiol. Biotechnol. 2010, 37, 419–423. [Google Scholar] [CrossRef]
- Zemek, R.; Prenerova, E.; Weyda, F. The first record of entomopathogenic fungus Paecilomyces fumosoroseus (Deuteromycota: Hyphomycetes) on the hibernating pupae of Cameraria ohridella (Lepidoptera: Gracillariidae). Entomol. Res. 2007, 37, A135–A136. [Google Scholar]
- Prenerova, E.; Zemek, R.; Volter, L.; Weyda, F. Strain of Entomopathogenic Fungus Isaria fumosorosea CCM 8367 (CCEFO.011.PFR) and the Method for Controlling Insect and Mite Pests. U.S. Patent No. US 08574566, 5 November 2013. [Google Scholar]
- Prenerova, E.; Zemek, R.; Volter, L.; Weyda, F. Strain of Entomopathogenic Fungus Isaria fumosorosea CCM 8367 (CCEFO.011.PFR) and the Method for Controlling Insect and Mite Pests. EPO Patent No. EP2313488, 29 April 2015. [Google Scholar]
- Skalický, A.; Bohatá, A.; Šimková, J.; Osborne, L.S.; Landa, Z. Selection of indigenous isolates of entomopathogenic soil fungus Metarhizium anisopliae under laboratory conditions. Folia Microbiol. 2014, 59, 269–276. [Google Scholar] [CrossRef]
- Ruther, J.; Podsiadlowski, L.; Hilker, M. Quinones in cockchafers: Additional function of a sex attractant as an antimicrobial agent. Chemoecology 2001, 11, 225–229. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- SAS Institute. SAS Stat Studio 3.8: User’s Guide; SAS Institute: Cary, NC, USA, 2018. [Google Scholar]
- SAS Institute. SAS/STAT 14.3: User’s Guide; SAS Institute: Cary, NC, USA, 2017. [Google Scholar]
- Hussein, H.M.; Zemek, R.; Habuštová, S.O.; Prenerová, E.; Adel, M.M. Laboratory evaluation of a new strain CCM 8367 of Isaria fumosorosea (syn. Paecilomyces fumosoroseus) on Spodoptera littoralis (Boisd.). Arch. Phytopathol. Plant Prot. 2013, 46, 1307–1319. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Šefrová, H.; Laštůvka, Z. Dispersal of the horse-chestnut leafminer, Cameraria ohridella Deschka & Dimic, 1986, in Europe: Its course, ways and causes (Lepidoptera: Gracillariidae). Entomol. Zeitschrift 2001, 111, 194–198. [Google Scholar]
- Hussein, H.M.; Skoková Habuštová, O.; Půža, V.; Zemek, R. Laboratory evaluation of Isaria fumosorosea CCM 8367 and Steinernema feltiae Ustinov against immature stages of the Colorado potato beetle. PLoS ONE 2016, 11, e0152399. [Google Scholar] [CrossRef] [PubMed]
- Zemek, R.; Prenerová, E.; Volter, L.; Awad, M.; Weyda, F.; Hussein, H.M.; Skoková Habuštová, O.; Půža, V. Non-target impacts of Isaria fumosorosea (Hypocreales: Cordycipitaceae) on natural enemies of arthropod pests. In Proceedings of the 5th International Symposium on Biological Control of Arthropods, Langkawi, Malaysia, 11–15 September 2017; Mason, P.G., Gillespie, D.R., Vincent, C., Eds.; CABI: Wallingford, UK, 2017; pp. 294–298, ISBN 978-1-78639-411-8. [Google Scholar]
- Pekrul, S.; Grula, E.A. Mode of infection of the corn earworm (Heliothis zea) by Beauveria bassiana as revealed by scanning electron microscopy. J. Invertebr. Pathol. 1979, 34, 238–247. [Google Scholar] [CrossRef]
- Hassan, A.E.M.; Dillon, R.J.; Charnley, A.K. Influence of accelerated germination of conidia on the pathogenicity of Metarhizium anisopliae for Manduca sexta. J. Invertebr. Pathol. 1989, 54, 277–279. [Google Scholar] [CrossRef]
- Wang, C.; St. Leger, R.J. Developmental and transcriptional responses to host and nonhost cuticles by the specific locust pathogen Metarhizium anisopliae var. acridum. Eukaryot. Cell 2005, 4, 937–947. [Google Scholar] [CrossRef] [Green Version]
- Ment, D.; Churchill, A.C.L.; Gindin, G.; Belausov, E.; Glazer, I.; Rehner, S.A.; Rot, A.; Donzelli, B.G.G.; Samish, M. Resistant ticks inhibit Metarhizium infection prior to haemocoel invasion by reducing fungal viability on the cuticle surface: Metarhizium-tick interactions and host resistance. Environ. Microbiol. 2012, 14, 1570–1583. [Google Scholar] [CrossRef]
- Sawada, M.; Sano, T.; Hanakawa, K.; Sirasoonthorn, P.; Oi, T.; Miura, K. Benzoquinone synthesis-related genes of Tribolium castaneum confer the robust antifungal host defense to the adult beetles through the inhibition of conidial germination on the body surface. J. Invertebr. Pathol. 2020, 169, 107298. [Google Scholar] [CrossRef] [PubMed]
- Grizanova, E.V.; Coates, C.J.; Dubovskiy, I.M.; Butt, T.M. Metarhizium brunneum infection dynamics differ at the cuticle interface of susceptible and tolerant morphs of Galleria mellonella. Virulence 2019, 10, 999–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, F.E.; Dowd, P.F.; McGuire, M.R.; Jackson, M.A.; Nelsen, T.C. In-vitro effects of secondary plant compounds on germination of blastospores of the entomopathogenic fungus Paecilomyces fumosoroseus (Deuteromycotina: Hyphomycetes). J. Invertebr. Pathol. 1997, 70, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Leuthardt, F.L.G.; Glauser, G.; Baur, B. Composition of alkaloids in different box tree varieties and their uptake by the box tree moth Cydalima perspectalis. Chemoecology 2013, 23, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Kiran, B.; Olgun, C.; Verep, D.; Gur, M.; Guney, K.; Altuner, E.M.; Ates, S. Determination of flavonoids and antimicrobial Behavior of non-wood forest product extracts. Fresenius Environ. Bull. 2018, 27, 2499–2504. [Google Scholar]
- Zazharskyi, V.V.; Davydenko, P.О.; Kulishenko, O.М.; Borovik, I.V.; Brygadyrenko, V.V. Antimicrobial activity of 50 plant extracts. Biosyst. Divers. 2019, 27, 163–169. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment 1 | Last-Instar Larvae | Pupae | Malformed Adults | ||||
|---|---|---|---|---|---|---|---|
| Duration | Died/Mycosed | Duration | Died/Mycosed | ||||
| Mean ± SE | n | % | Mean ± SE | n | % | % | |
| Control | 4.24 ± 0.41 | 29 | 3.3/0 | 10.10 ± 0.15 | 21 | 27.6/0 | 0 |
| 1 × 104 | 5.04 ± 0.58 | 28 | 6.7/0 | 10.33 ± 0.11 | 18 | 35.7/0 | 0 |
| 1 × 105 | 5.53 ± 0.54 | 30 | 0/0 | 10.17 ± 0.13 | 24 | 20.0/0 | 0 |
| 1 × 106 | 4.87 ± 0.43 | 30 | 0/0 | 10.46 ± 0.10 | 26 | 13.3/0 | 0 |
| 1 × 107 | 5.00 ± 0.31 | 30 | 0/0 | 10.33 ± 0.11 | 21 | 30.0/10.0 | 0 |
| 1 × 108 | 3.96 ± 0.32 | 28 | 6.7/3.3 | 10.27 ± 0.25 | 15 | 46.4/28.6 | 20.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zemek, R.; Konopická, J.; Ul Abdin, Z. Low Efficacy of Isaria fumosorosea against Box Tree Moth Cydalima perspectalis: Are Host Plant Phytochemicals Involved in Herbivore Defence against Fungal Pathogens? J. Fungi 2020, 6, 342. https://doi.org/10.3390/jof6040342
Zemek R, Konopická J, Ul Abdin Z. Low Efficacy of Isaria fumosorosea against Box Tree Moth Cydalima perspectalis: Are Host Plant Phytochemicals Involved in Herbivore Defence against Fungal Pathogens? Journal of Fungi. 2020; 6(4):342. https://doi.org/10.3390/jof6040342
Chicago/Turabian StyleZemek, Rostislav, Jana Konopická, and Zain Ul Abdin. 2020. "Low Efficacy of Isaria fumosorosea against Box Tree Moth Cydalima perspectalis: Are Host Plant Phytochemicals Involved in Herbivore Defence against Fungal Pathogens?" Journal of Fungi 6, no. 4: 342. https://doi.org/10.3390/jof6040342