
Aquatic Hyphomycete Taxonomic Relatedness Translates into Lower Genetic Divergence of the Nitrate Reductase Gene

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains, Growth Conditions and DNA Extraction
2.2. Fungal Identification
2.3. Nitrate Reductase Gene Amplification
2.4. Cloning and Sequence Alignments
2.5. In Silico Assays
2.6. Phylogenetic Analysis
3. Results
3.1. Evolutionary Taxonomic Divergence
3.2. Evolutionary Divergence of Nitrate Reductase Gene
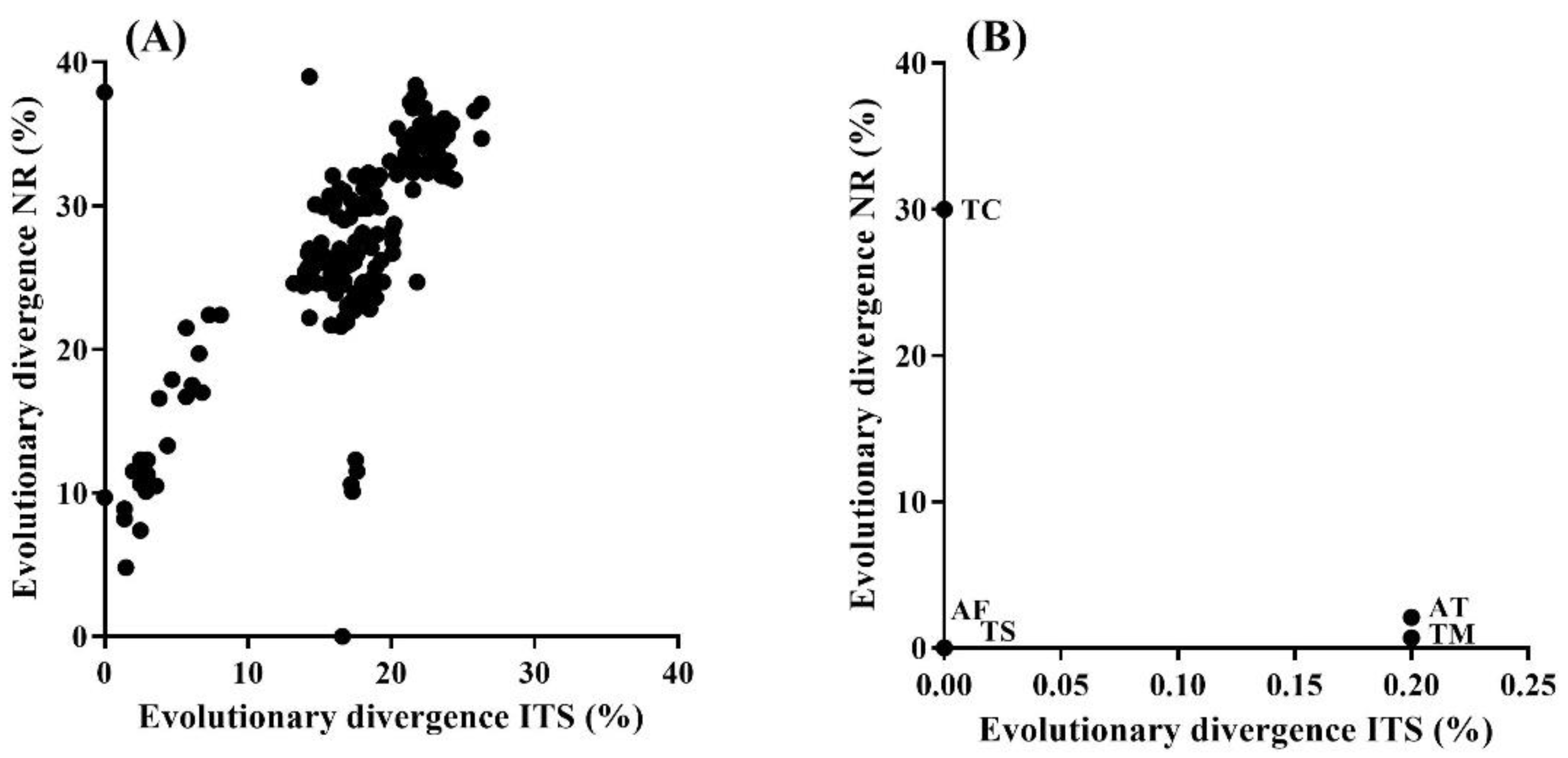
3.3. Relationship between Taxonomic and Functional Evolutionary Divergence
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Allan, J.D.; Castillo, M.M. Stream Ecology: Structure and Function of Running Waters, 2nd ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 1–9. [Google Scholar]
- Abelho, M. From litterfall to breakdown in streams: A review. Sci. World J. 2001, 1, 656–680. [Google Scholar] [CrossRef] [Green Version]
- Graça, M.A.S.; Ferreira, V.; Canhoto, C.; Encalada, A.C.; Guerrero-Bolaño, F.; Wantzen, K.M.; Boyero, L. A conceptual model of litter breakdown in low order streams. Int. Rev. Hydrobiol. 2015, 100, 1–12. [Google Scholar] [CrossRef]
- Marks, J.C. Revisiting the fates of dead leaves that fall into streams. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 547–568. [Google Scholar] [CrossRef]
- Gulis, V.; Suberkropp, K. Interactions between stream fungi and bacteria associated with decomposing leaf litter at different levels of nutrient availability. Aquat. Microb. Ecol. 2003, 30, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Pascoal, C.; Cássio, F. Contribution of fungi and bacteria to leaf litter decomposition in a polluted river. Appl. Environ. Microbiol. 2004, 70, 5266–5273. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Royer, T.V.; Leff, L.G. Diversity of fungi, bacteria, and actinomycetes on leaves decomposing in a stream. Appl. Environ. Microbiol. 2007, 73, 756–767. [Google Scholar] [CrossRef] [Green Version]
- Gessner, M.O.; Gulis, V.; Kuehn, K.A.; Chauvet, E.; Suberkropp, K. Fungal decomposers of plant litter in aquatic ecosystems. In Environmental and Microbial Relationships, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 301–324. [Google Scholar]
- Chamier, A.-C. Cell-wall-degrading enzymes of aquatic hyphomycetes: A review. Bot. J. Linn. Soc. 1985, 91, 67–81. [Google Scholar] [CrossRef]
- Zemek, J.; Marvanová, L.; Kuniak, L.; Kadleciková, B. Hydrolytic enzymes in aquatic hyphomycetes. Folia Microbiol. 1985, 30, 363–372. [Google Scholar] [CrossRef]
- Cross, W.F.; Benstead, J.P.; Rosemond, A.D.; Wallace, J.B. Consumer-resource stoichiometry in detritus-based streams. Ecol. Lett. 2003, 6, 721–732. [Google Scholar] [CrossRef] [Green Version]
- Gulis, V.; Suberkropp, K.; Rosemond, A.D. Comparison of fungal activities on wood and leaf litter in unaltered and nutrient-enriched headwater streams. Appl. Environ. Microbiol. 2008, 74, 1094–1101. [Google Scholar] [CrossRef] [Green Version]
- Woodward, G.; Gessner, M.O.; Giller, P.S.; Gulis, V.; Hladyz, S.; Lecerf, A.; Malmqvist, B.; McKie, B.G.; Tiegs, S.D.; Cariss, H.; et al. Continental-scale effects of nutrient pollution on stream ecosystem functioning. Science (80-) 2012, 336, 1438–1440. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, I.; Seena, S.; Pascoal, C.; Cássio, F. Elevated temperature may intensify the positive effects of nutrients on microbial decomposition in streams. Freshw. Biol. 2014, 59, 2390–2399. [Google Scholar] [CrossRef]
- Ferreira, V.; Castagneyrol, B.; Koricheva, J.; Gulis, V.; Chauvet, E.; Graça, M.A.S. A meta-analysis of the effects of nutrient enrichment on litter decomposition in streams. Biol. Rev. 2014, 90, 669–688. [Google Scholar] [CrossRef]
- Pereira, A.; Geraldes, P.; Lima-Fernandes, E.; Fernandes, I.; Cássio, F.; Pascoal, C. Structural and functional measures of leaf-associated invertebrates and fungi as predictors of stream eutrophication. Ecol. Indic. 2016, 69, 648–656. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Caraco, N.F.; Correll, D.L.; Howarth, R.W.; Sharpley, A.N.; Smith, V.H. Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecol. Appl. 1998, 8, 559–568. [Google Scholar] [CrossRef]
- Melillo, J.M.; Naiman, R.J.; Aber, J.D.; Linkins, A.E. Factors controlling mass loss and nitrogen dynamics of plant litter decaying in northern streams. Bull. Mar. Sci. 1984, 35, 341–356. [Google Scholar]
- Daniel-Vedele, F.; Filleur, S.; Caboche, M. Nitrate transport: A key step in nitrate assimilation. Physiol. Metab. 1998, 1, 235–239. [Google Scholar] [CrossRef]
- Unkles, S.E.; Wang, R.; Wang, Y.; Glass, A.D.M.; Crawford, N.M.; Kinghorn, J.R. Nitrate reductase activity is required for nitrate uptake into fungal but not plant cells. J. Biol. Chem. 2004, 279, 28182–28186. [Google Scholar] [CrossRef] [Green Version]
- Marcos, A.T.; Ramos, M.S.; Marcos, J.F.; Carmona, L.; Strauss, J.; Cánovas, D. Nitric oxide synthesis by nitrate reductase is regulated during development in Aspergillus. Mol. Microbiol. 2016, 99, 15–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorfer, M.; Blumhoff, M.; Klaubauf, S.; Urban, A.; Inselsbacher, E.; Bandian, D.; Mitter, B.; Sessitsch, A.; Wanek, W.; Strauss, J. Community profiling and gene expression of fungal assimilatory nitrate reductases in agricultural soil. ISME J. 2011, 5, 1771–1783. [Google Scholar] [CrossRef] [Green Version]
- Bärlocher, F.; Corkum, M. Nutrient enrichment overwhelms diversity effects in leaf decomposition by stream fungi. Oikos 2003, 101, 247–252. [Google Scholar] [CrossRef]
- Duarte, S.; Pascoal, C.; Cássio, F.; Bärlocher, F. Aquatic hyphomycete diversity and identity affect leaf litter decomposition in microcosms. Oecologia 2006, 147, 658–666. [Google Scholar] [CrossRef]
- Fernandes, I.; Pascoal, C.; Cássio, F. Intraspecific traits change biodiversity effects on ecosystem functioning under metal stress. Oecologia 2011, 166, 1019–1028. [Google Scholar] [CrossRef] [Green Version]
- Dang, C.K.; Chauvet, E.; Gessner, M.O. Magnitude and variability of process rates in fungal diversity-litter decomposition relationships. Ecol. Lett. 2005, 8, 1129–1137. [Google Scholar] [CrossRef] [Green Version]
- Geraldes, P.; Cássio, F.; Pascoal, C. Effects of increased temperature and aquatic fungal diversity on litter decomposition. Fungal Ecol. 2012, 5, 734–740. [Google Scholar] [CrossRef] [Green Version]
- Andrade, R.; Pascoal, C.; Cássio, F. Effects of inter and intraspecific diversity and genetic divergence of aquatic fungal communities on leaf litter decomposition-a microcosm experiment. FEMS Microbiol. Ecol. 2016, 92, fiw102. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The microbial engines that drive earth’s biogeochemical cycles. Science (80-) 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolcheva, L.G.; Bärlocher, F. Taxon-specific fungal primers reveal unexpectedly high diversity during leaf decomposition in a stream. Mycol. Prog. 2004, 3, 41–49. [Google Scholar] [CrossRef]
- Seena, S.; Duarte, S.; Pascoal, C.; Cássio, F. Intraspecific variation of the aquatic fungus Articulospora tetracladia: An ubiquitous perspective. PLoS ONE 2012, 7, e35884. [Google Scholar]
- Baschien, C.; Tsui, C.K.; Gulis, V.; Szewzyk, U.; Marvanová, L. The molecular phylogeny of aquatic hyphomycetes with affinity to the Leotiomycetes. Fungal Biol. 2013, 117, 660–672. [Google Scholar] [CrossRef]
- Solé, M.; Müller, I.; Pecyna, M.J.; Fetzer, I.; Harms, H.; Schlossera, D. Differential regulation by organic compounds and heavy metals of multiple laccase genes in the aquatic hyphomycete Clavariopsis aquatica. Appl. Environ. Microbiol. 2012, 78, 4732–4739. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.L.; Marvanová, L. Broad geographical and ecological diversity from similar genomic toolkits in the ascomycete genus Tetracladium. bioRxiv 2020, 1–33. [Google Scholar]
- Bärlocher, F. Sporulation by aquatic hyphomycetes. In Methods to Study Litter Decomposition: A Practical Guide, 1st ed.; Graça, M.A.S., Bärlocher, F., Gessner, M.O., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 185–188. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Analysis of phylogenetic relationship by amplification and direct sequencing of ribosimal RNA genes. In PCR Protocols: A Guide to Methods and Applications, 1st ed.; Innis, M.A., Gefland, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: London, UK, 1990; pp. 315–322. [Google Scholar]
- Duarte, S.; Seena, S.; Bärlocher, F.; Cássio, F.; Pascoal, C. Preliminary insights into the phylogeography of six aquatic hyphomycete species. PLoS ONE 2012, 7, e45289. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, R.H.; Kristiansson, E.; Ryberg, M.; Hallenberg, N.; Larsson, K.-H. Intraspecific ITS variability in the kingdom fungi as expressed in the international sequence databases and its implications for molecular species identification. Evol. Bioinform. 2008, 4, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Luo, X.; Wei, W.; Zheng, Y.; Dou, Y.; Cai, X. Calculation of evolutionary correlation between individual genes and full-length genome: A method useful for choosing phylogenetic markers for molecular epidemiology. PLoS ONE 2013, 8, e81106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, P.R.; Quijada, L.; Smith, C.A.; Baral, H.O.; Hosoya, T.; Baschien, C.; Pärtel, K.; Zhuang, W.Y.; Haelewaters, D.; Park, D.; et al. A multigene phylogeny toward a new phylogenetic classification of Leotiomycetes. IMA Fungus 2019, 10, 1–22. [Google Scholar] [CrossRef]
- Belliveau, M.J.R.; Bärlocher, F. Molecular evidence confirms multiple origins of aquatic hyphomycetes. Mycol. Res. 2005, 109, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- Baschien, C.; Marvanová, L.; Szewzyk, U. Phylogeny of selected aquatic hyphomycetes based on morphological and molecular data. Nov. Hedwig. 2006, 83, 311–352. [Google Scholar] [CrossRef]
- Campbell, J.; Marvanová, L.; Gulis, V. Evolutionary relationships between aquatic anamorphs and teleomorphs: Tricladium and Varicosporium. Mycol. Res. 2009, 113, 1322–1334. [Google Scholar] [CrossRef] [PubMed]
- Nikolcheva, L.G.; Bärlocher, F. Phylogeny of Tetracladium based on 18S rDNA. Czech Mycol. 2002, 53, 285–295. [Google Scholar] [CrossRef]
- Miersch, J.; Grancharov, K. Cadmium and heat response of the fungus Heliscus lugdunensis isolated from highly polluted and unpolluted areas. Amino Acids 2008, 34, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Quainoo, S.; Seena, S.; Graça, M.A.S. Copper tolerant ecotypes of Heliscus lugdunensis differ in their ecological function and growth. Sci. Total Environ. 2016, 544, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.L.; Carvalho, A.; Bärlocher, F.; Canhoto, C. Are fungal strains from salinized streams adapted to salt-rich conditions? Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seena, S.; Casotti, C.; Cornut, J. Inter- and intraspecific functional variability of aquatic fungal decomposers and freshwater ecosystem processes. Sci. Total Environ. 2020, 707, 135570. [Google Scholar] [CrossRef]
- Duarte, S.; Bärlocher, F.; Pascoal, C.; Cássio, F. Biogeography of aquatic hyphomycetes: Current knowledge and future perspectives. Fungal Ecol. 2016, 19, 169–181. [Google Scholar] [CrossRef]
- Grossart, H.P.; Massana, R.; McMahon, K.D.; Walsh, D.A. Linking metagenomics to aquatic microbial ecology and biogeochemical cycles. Limnol. Oceanogr. 2020, 65, S2–S20. [Google Scholar] [CrossRef] [Green Version]
- Green, S.J.; Venkatramanan, R.; Naqib, A. Deconstructing the polymerase chain reaction: Understanding and correcting bias associated with primer degeneracies and primer-template mismatches. PLoS ONE 2015, 10, e0128122. [Google Scholar] [CrossRef]
- Baral, H.-O. Inoperculate discomycetes. In Syllabus of Plant Families: A. Engler’s Syllabus der Pflanzenfamilien Part 1/2: Ascomycota; Jaklitsch, W., Baral, H.O., Lücking, R., Lumbsch, H.T., Frey, W., Eds.; Borntraeger: Stuttgar, Germany, 2016. [Google Scholar]
- Gleason, F.H.; Kagami, M.; Lefevre, E.; Sime-Ngando, T. The ecology of chytrids in aquatic ecosystems: Roles in food web dynamics. Fungal Biol. Rev. 2008, 22, 17–25. [Google Scholar] [CrossRef]
- Chamberlain, S.A.; Bronstein, J.L.; Rudgers, J.A. How context dependent are species interactions? Ecol. Lett. 2014, 17, 881–890. [Google Scholar] [CrossRef]
- Jabiol, J.; Bruder, A.; Gessner, M.O.; Makkonen, M.; McKie, B.G.; Peeters, E.T.H.M.; Vos, V.C.A.; Chauvet, E. Diversity patterns of leaf-associated aquatic hyphomycetes along a broad latitudinal gradient. Fungal Ecol. 2013, 6, 439–448. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, I.; Pereira, A.; Trabulo, J.; Pascoal, C.; Cássio, F.; Duarte, S. Microscopy- or DNA-based analyses: Which methodology gives a truer picture of stream-dwelling decomposer fungal diversity? Fungal Ecol. 2015, 18, 130–134. [Google Scholar] [CrossRef]
- Hughes, A.R.; Inouye, B.D.; Johnson, M.T.J.; Underwood, N.; Vellend, M. Ecological consequences of genetic diversity. Ecol. Lett. 2008, 11, 609–623. [Google Scholar] [CrossRef]
- Graham, E.B.; Knelman, J.E.; Schindlbacher, A.; Siciliano, S.; Breulmann, M.; Yannarell, A.; Beman, J.M.; Abell, G.; Philippot, L.; Prosser, J.; et al. Microbes as engines of ecosystem function: When does community structure enhance predictions of ecosystem processes? Front. Microbiol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linz, A.M.; Aylward, F.O.; Bertilsson, S.; McMahon, K.D. Time-series metatranscriptomes reveal conserved patterns between phototrophic and heterotrophic microbes in diverse freshwater systems. Limnol. Oceanogr. 2020, 65, S101–S112. [Google Scholar] [CrossRef] [Green Version]
- Kamp, A.; Høgslund, S.; Risgaard-Petersen, N.; Stief, P. Nitrate storage and dissimilatory nitrate reduction by eukaryotic microbes. Front. Microbiol. 2015, 6, 1492. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Species | Culture Collection | Codes | Class | Order | Genbank Accession Number | ||
|---|---|---|---|---|---|---|---|
| ITS | Nitrate Reductase | Genome | |||||
| Alatospora acuminata Ingold | UMB collection, Portugal | UMB-741.11 | Leotiomycetes | Helotiales | MZ773535 | MZ812105 | n/a |
| Alatospora pulchella Marvanová | UMB collection, Portugal | UMB-1115 | Leotiomycetes | Helotiales | MZ773536 | MZ812106 | n/a |
| Anguillospora crassa Ingold | UMB collection, Portugal | UMB-1150 | Dothideomycetes | Pleosporales | MZ77353539 | MZ812109 | n/a |
| Anguillospora filiformis Greath | UMB collection, Portugal | UMB-225.02 | Dothideomycetes | Pleosporales | MZ773533 | MZ812102 | n/a |
| UMB-232.02 | OK037621 | MZ812103 | n/a | ||||
| Aquanectria penicillioides (Ingold) L. Lombard and Crous | UMB collection, Portugal | UMB-304.05 | Sordariomycetes | Hypocreales | GQ411325.1 | n/a | n/a |
| Nakdonggang National Institute of Biological Resources, South Korea | NNIBRFG19 | s/m | s/m | PYIU00000000.1 | |||
| Articulospora tetracladia Ingold | UMB collection, Portugal | UMB-72.01 | Leotiomycetes | Helotiales | OK605571 | n/a | n/a |
| UMB-712.10 | OK605572 | n/a | n/a | ||||
| UMB-719.10 | OK037616 | MZ812096 | n/a | ||||
| UMB-1144 | OK605573 | n/a | n/a | ||||
| Nakdonggang National Institute of Biological Resources, South Korea | NNIBRFG329 | s/m | s/m | GCA_003415645.1 | |||
| Clavariopsis aquatica De Wild. | Leibniz-Institute of Freshwater Ecology and Inland Fisheries, Germany | WD(A)-00-1 | Dothideomycetes | Pleosporales | s/m | s/m | GCA_013620735.1 |
| Collembolispora barbata Marvanová, Pascoal and Cássio | UMB collection, Portugal | UMB-88.01 | n/a | n/a | MZ773532 | MZ812101 | n/a |
| Dactylella cylindrospora (R.C. Cooke) A. Rubner | Laboratory for Conservation and Utilization of Bio-Resources and Key Laboratory for Microbial Diversity, Southwest China | CBS325.70 | Orbiliomycetes | Orbiliales | s/m | s/m | GCA_012184295.1 |
| Dimorphospora foliicola Tubaki | UMB collection, Portugal | UMB-1119 | Leotiomycetes | Helotiales | MZ773538 | MZ812108 | n/a |
| Neonectria lugdunensis (Sacc. and Therry) L. Lombard and Crous | UMB collection, Portugal | UMB-3.00 | Sordariomycetes | Hypocreales | OK605574 | n/a | n/a |
| UMB-160.01 | OK605575 | n/a | n/a | ||||
| UMB-161.01 | OK605576 | n/a | n/a | ||||
| UMB-311.06 | OK605577 | n/a | n/a | ||||
| Lemonniera aquatica De Wild. | UMB collection, Portugal | UMB-594.10 | Leotiomycetes | Helotiales | MZ773530 | MZ812094 | n/a |
| Lunulospora curvula Ingold | UMB collection, Portugal | UMB-108.01 | n/a | n/a | OK605578 | n/a | n/a |
| UMB-498.09 | OK605579 | n/a | n/a | ||||
| Margaritispora aquatica Ingold | Nakdonggang National Institute of Biological Resources, South Korea | NNIBRFG339 | Leotiomycetes | Helotiales | s/m | s/m | GCA_007644065.1 |
| Tetracladium apiense R.C. Sinclair and Eicker | UMB collection, Portugal | UMB-535.10 | Leotiomycetes | Helotiales | OK037615 | MZ812093 | n/a |
| Tetracladium furcatum Descals | Biology Dept., Mount Allison University, Sackville, NB, Canada | F-11883 | Leotiomycetes | Helotiales | AF411026.1 | HQ234857.1 | n/a |
| Tetracladium marchalianum De Wild. | UMB collection, Portugal | UMB-1028.13 | Leotiomycetes | Helotiales | OK037619 | MZ812099 | n/a |
| UMB-1079.13 | OK037620 | MZ812100 | n/a | ||||
| Biology Dept., Mount Allison University, Sackville, NB, Canada | F-312 | AF411023.1 | HQ234858.1 | n/a | |||
| Tetracladium maxilliforme (Rostr.) Ingold | Biology Dept., Mount Allison University, Sackville, NB, Canada | F-14286 | Leotiomycetes | Helotiales | AF411027.1 | HQ234859.1 | n/a |
| Tetracladium setigerum (Grove) Ingold | Institute for Environmental Sciences, University of Koblenz-Landau, Germany | CCM F-20987 | Leotiomycetes | Helotiales | KU519120.1 | HQ234860.1 | n/a |
| Thelonectria rubi (Osterw.) C. Salgado & P. Chaverri | Agricultural Research Service, United States Department of Agriculture, USA | CBS 177.27 | Sordariomycetes | Hypocreales | s/m | s/m | GCA_013420875.1 |
| Tricladium chaetocladium Ingold | UMB collection, Portugal | UMB-904.12 | Leotiomycetes | Helotiales | OK037617 | MZ812097 | n/a |
| UMB-1116 | MZ773531 | MZ812095 | n/a | ||||
| Tricladium splendens Ingold | UMB collection, Portugal | UMB-100.01 | Leotiomycetes | Helotiales | OK037618 | MZ812098 | n/a |
| UMB-414.09 | OK605580 | n/a | n/a | ||||
| UMB-1117 | MZ773537 | MZ812107 | n/a | ||||
| Varicosporium elodeae W. Kegel | UMB collection, Portugal | UMB-310.06 | Leotiomycetes | Helotiales | OK605581 | n/a | n/a |
| UMB-713.10 | MZ773534 | MZ812104 | n/a | ||||
| UMB-878.12 | OK605582 | n/a | n/a | ||||
| Species | Strain Code | Nitrate Reductase Amplification Success |
|---|---|---|
| Alatospora acuminata | UMB-741.11 | Yes |
| Alatospora pulchella | UMB-1115 | Yes |
| Anguillospora crassa | UMB-217.02 | No |
| Anguillospora crassa | UMB-1150 | Yes |
| Anguillospora filiformis | UMB-225.02 | Yes |
| Anguillospora filiformis | UMB-232.02 | Yes |
| Aquanectria penicillioides | UMB-304.05 | No |
| Articulospora tetracladia | UMB-72.01 | No |
| Articulospora tetracladia | UMB-712.10 | Yes |
| Articulospora tetracladia | UMB-719.10 | Yes |
| Articulospora tetracladia | UMB-1144 | No |
| Collembolispora barbata | UMB-88.01 | Yes |
| Dimorphospora foliicola | UMB-1119 | Yes |
| Lemonniera aquatica | UMB-594.10 | Yes |
| Lunulospora curvula | UMB-108.01 | No |
| Lunulospora curvula | UMB-498.09 | No |
| Neonectria lugdunensis | UMB-3.00 | No |
| Neonectria lugdunensis | UMB-160.01 | No |
| Neonectria lugdunensis | UMB-161.01 | No |
| Neonectria lugdunensis | UMB-311.06 | No |
| Tetracladium apiense | UMB-535.10 | Yes |
| Tetracladium marchalianum | UMB-1028.13 | Yes |
| Tetracladium marchalianum | UMB-1079.13 | Yes |
| Tricladium chaetocladium | UMB-904.12 | Yes |
| Tricladium chaetocladium | UMB-1116 | Yes |
| Tricladium splendens | UMB-100.01 | Yes |
| Tricladium splendens | UMB-414.09 | No |
| Tricladium splendens | UMB-1117 | Yes |
| Varicosporium elodeae | UMB-310.06 | No |
| Varicosporium elodeae | UMB-713.10 | Yes |
| Varicosporium elodeae | UMB-878.12 | No |
| Amplification success rate (%) | 58.1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariz, J.; Franco-Duarte, R.; Cássio, F.; Pascoal, C.; Fernandes, I. Aquatic Hyphomycete Taxonomic Relatedness Translates into Lower Genetic Divergence of the Nitrate Reductase Gene. J. Fungi 2021, 7, 1066. https://doi.org/10.3390/jof7121066
Mariz J, Franco-Duarte R, Cássio F, Pascoal C, Fernandes I. Aquatic Hyphomycete Taxonomic Relatedness Translates into Lower Genetic Divergence of the Nitrate Reductase Gene. Journal of Fungi. 2021; 7(12):1066. https://doi.org/10.3390/jof7121066
Chicago/Turabian StyleMariz, Joana, Ricardo Franco-Duarte, Fernanda Cássio, Cláudia Pascoal, and Isabel Fernandes. 2021. "Aquatic Hyphomycete Taxonomic Relatedness Translates into Lower Genetic Divergence of the Nitrate Reductase Gene" Journal of Fungi 7, no. 12: 1066. https://doi.org/10.3390/jof7121066