
Scedosporium spp. from Clinical Setting in Argentina, with the Proposal of the New Pathogenic Species Scedosporium americanum

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains
2.2. DNA Extraction, Amplification and Sequencing
2.3. Molecular Identification and Phylogeny
2.4. Morphological Characterization
2.5. Antifungal Susceptibility Testing
3. Results
3.1. Molecular Presumptive Identification of the Strains
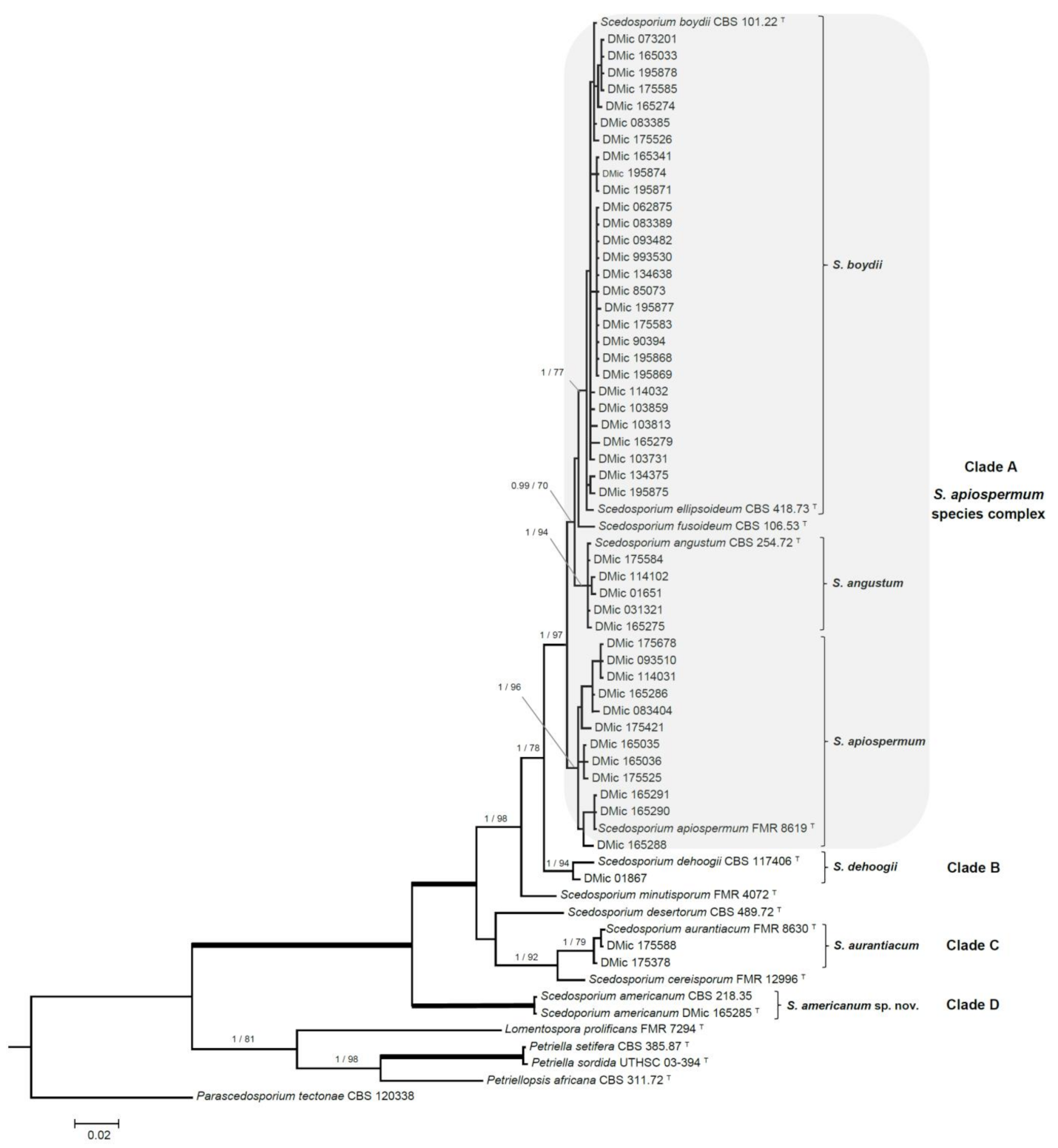
3.2. Phylogenetic Analyses
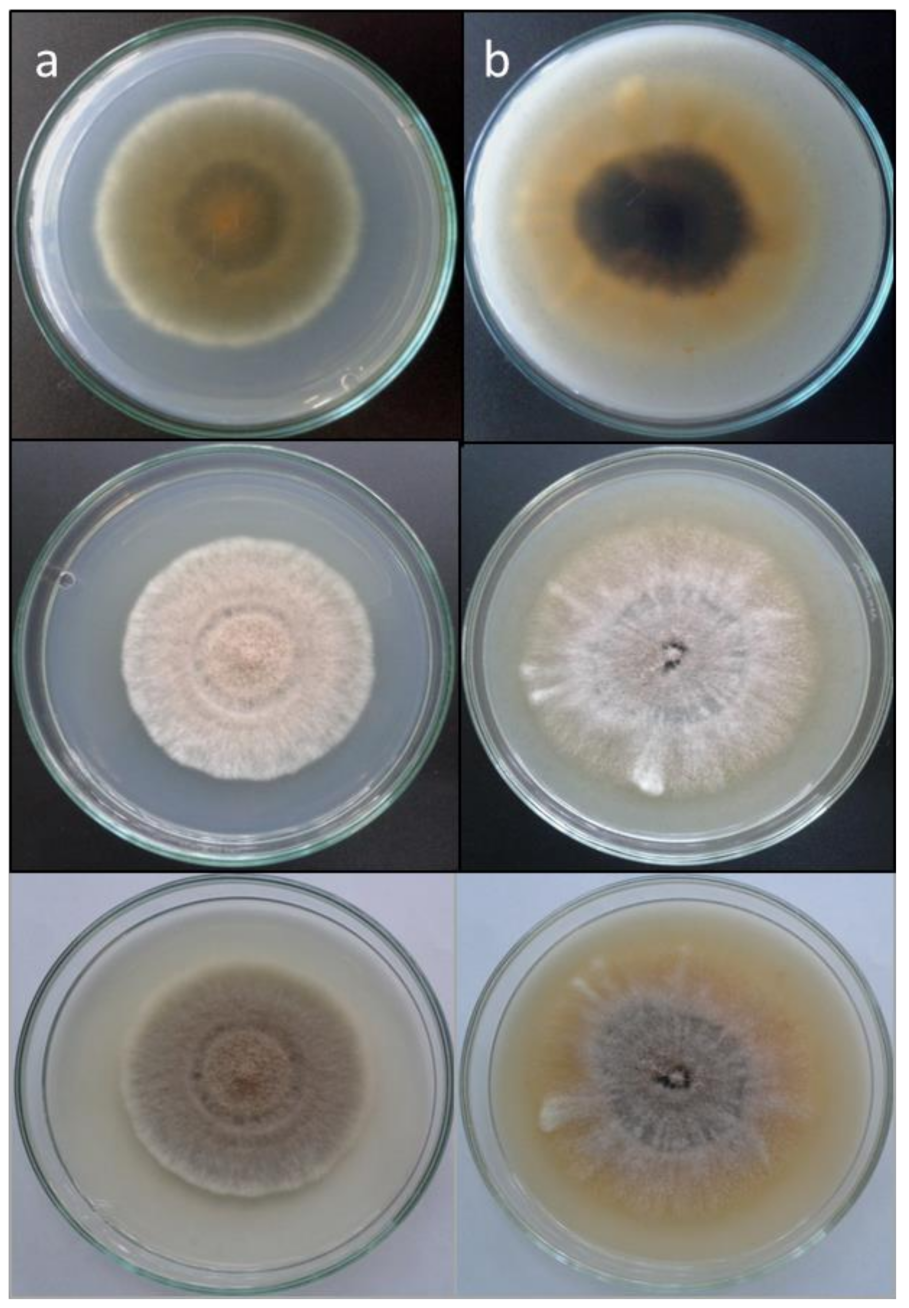
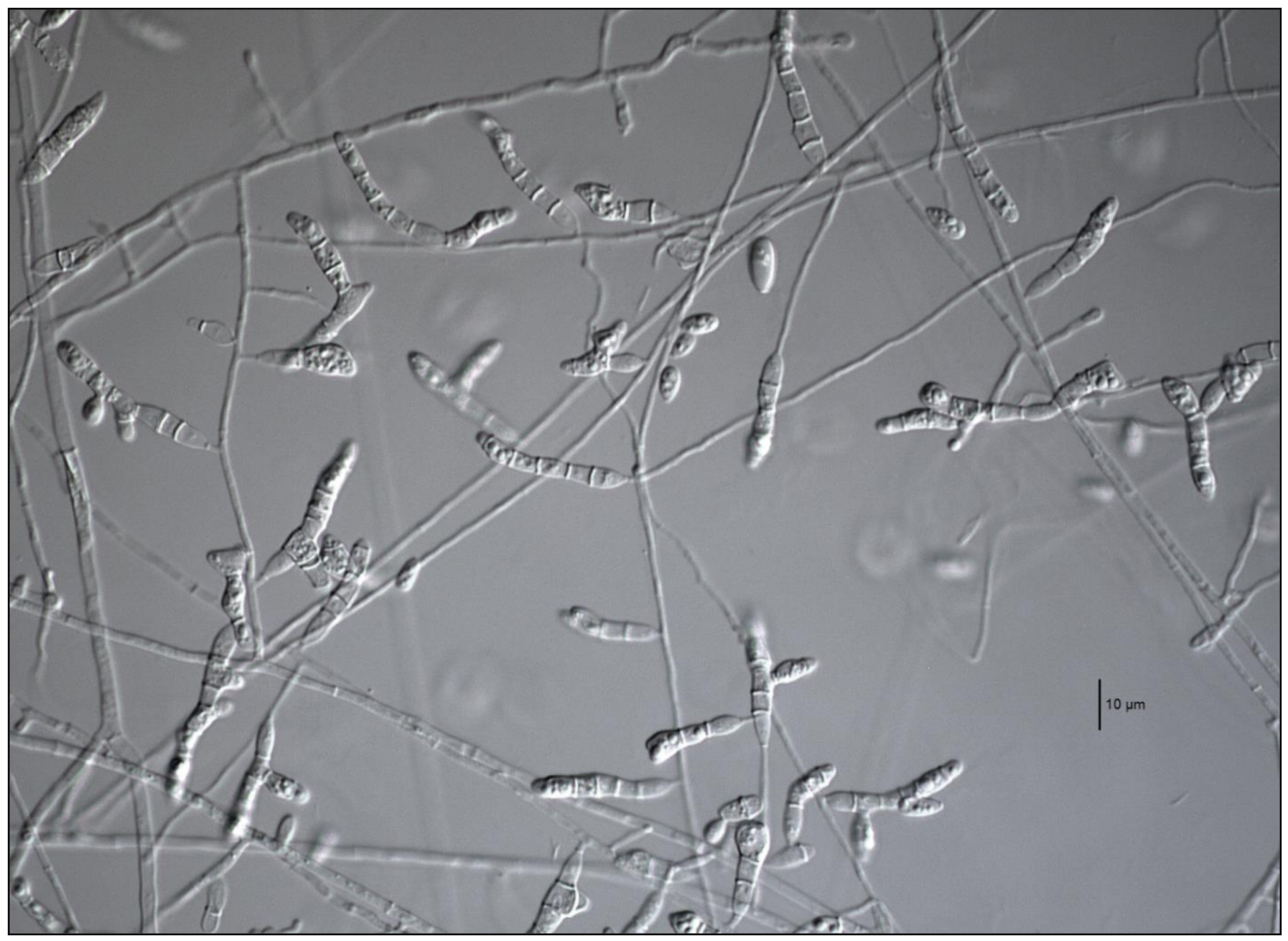
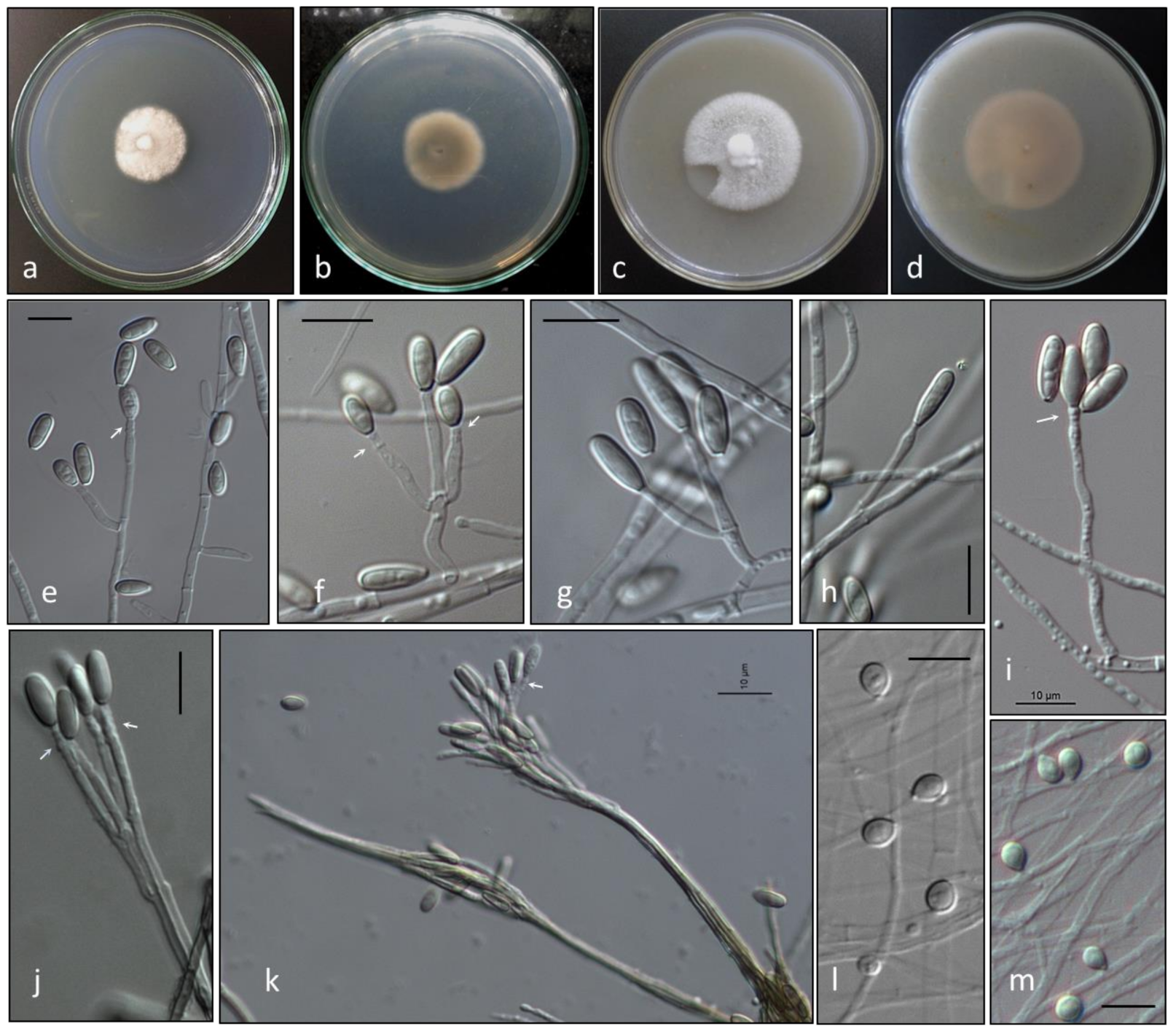
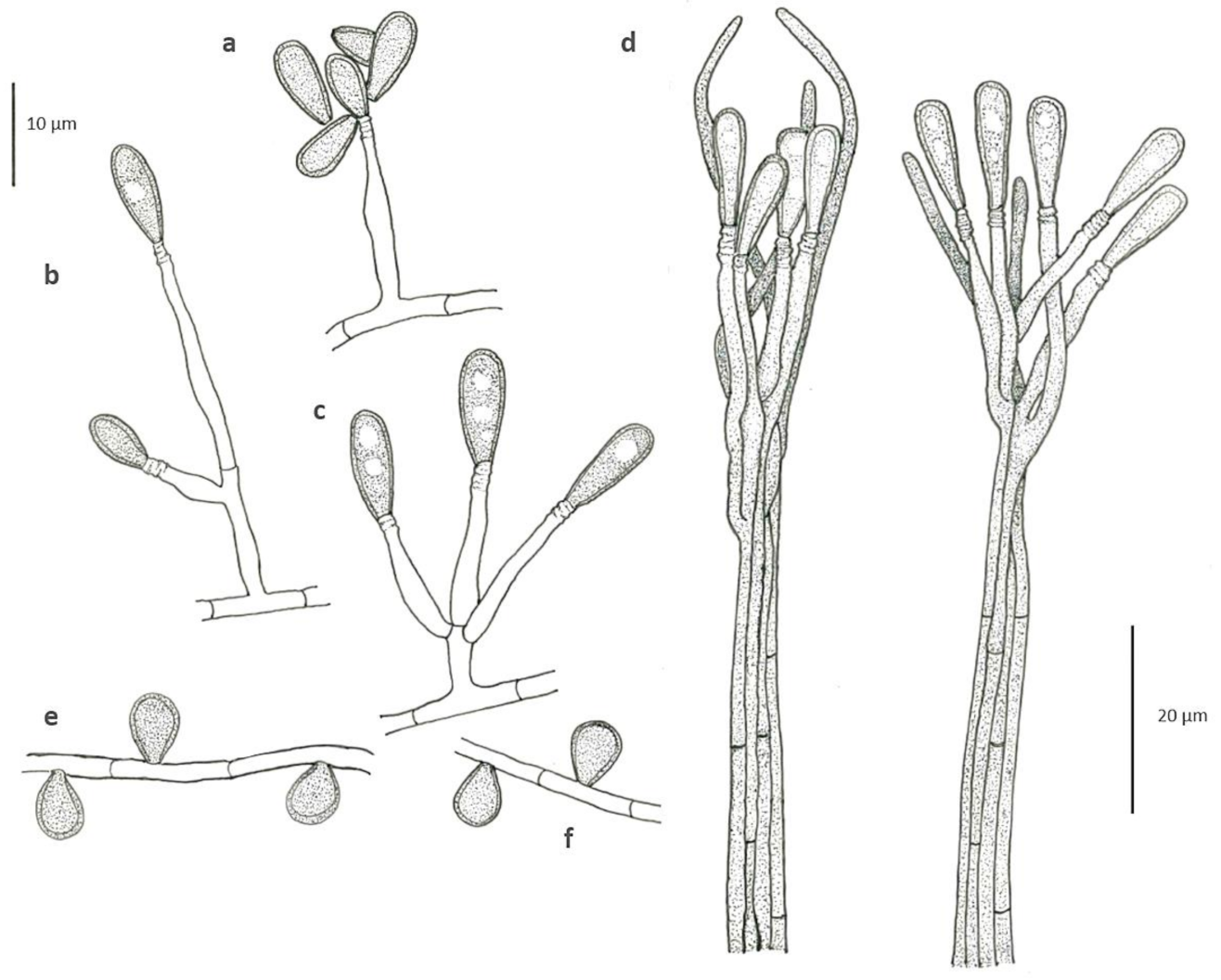
3.3. Morphology
3.4. Taxonomy
3.5. Antifungal Susceptibility Testing
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sandoval-Denis, M.; Guarro, J.; Cano-Lira, J.; Sutton, D.; Wiederhold, N.; De Hoog, G.; Abbott, S.; Decock, C.; Sigler, L.; Gené, J. Phylogeny and taxonomic revision of Microascaceae with emphasis on synnematous fungi. Stud. Mycol. 2016, 83, 193–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woudenberg, J.; Meijer, M.; Houbraken, J.; Samson, R. Scopulariopsis and scopulariopsis-like species from indoor environments. Stud. Mycol. 2017, 88, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Zhu, H.; Niu, Y.; Guo, Y.; Du, X.; Guo, J.; Zhang, L.; Qin, C. Phylogeny and taxonomic revision of Kernia and Acaulium. Sci. Rep. 2020, 10, 10302. [Google Scholar]
- Ramirez-Garcia, A.; Pellon, A.; Rementeria, A.; Buldain, I.; Barreto-Bergter, E.; Rollin-Pinheiro, R.; de Meirelles, J.V.; Xisto, M.I.D.S.; Ranque, S.; Havlicek, V.; et al. Scedosporium and Lomentospora: An updated over-view of underrated opportunists. Med. Mycol. 2018, 56, 102–125. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.K. Polycytella hominisgen. et sp. nov., a cause of human pale grain mycetoma. Med. Mycol. 1987, 25, 301–305. [Google Scholar] [CrossRef]
- Borman, A.M.; Campbell, C.K.; Linton, C.J.; Bridge, P.D.; Johnson, E.M. Polycytella Hominis is a mutated form of Scedosporium Apiospermum. Med. Mycol. 2006, 44, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Saccardo, P.A. Notae mycologicae. Series XIII. Ann. Mycol. 1911, 9, 249–257. [Google Scholar]
- Radaeli, F. Caso singolare di alterazione cutánea e profonda di natura probabilmente micotica in un piede (con pre-sentizioni di culture, di preparati microscopici e di microfotografie). Giornale Italiano Delle Malattie Veneree e Della Pelle 1911, 52, 109–116. [Google Scholar]
- Costantin, J. Les Mucédinées Simples; Paul Klincksieck: Paris, France, 1888. [Google Scholar]
- Negroni, P.; Fisher, I. Pseudallescheria sheari n. gen., n. sp. aislada de un paramicetoma de la rodilla. Rev. Inst. Bacteriológico “Dr Carlos G Malbrán” 1944, 12, 195–204. [Google Scholar]
- Shear, C. Life history of an undescribed ascomycete isolated from a granular mycetoma of man. Mycologia 1922, 14, 239–243. [Google Scholar] [CrossRef]
- Lackner, M.; de Hoog, G.S.; Yang, L.; Ferreira Moreno, L.; Ahmed, S.A.; Andreas, F.; Kaltseis, J.; Nagl, M.; Lass-Flörl, C.; Risslegger, B.; et al. Proposed nomenclature for Pseudallescheria, Scedosporium and related genera. Fungal Divers 2014, 67, 1–10. [Google Scholar] [CrossRef]
- Chen, M.; Zeng, J.; De Hoog, G.S.; Stielow, B.; Ende, A.H.G.G.V.D.; Liao, W.; Lackner, M. The ‘species complex’ issue in clinically relevant fungi: A case study in Scedosporium apiospermum. Fungal Biol. 2016, 120, 137–146. [Google Scholar] [CrossRef]
- Rougeron, A.; Giraud, S.; Alastruey-Izquierdo, A.; Cano-Lira, J.; Rainer, J.; Mouhajir, A.; Le Gal, S.; Nevez, G.; Meyer, W.; Bouchara, J.P. Ecology of Scedosporium Species: Present Knowledge and Future Research. Mycopathologia 2018, 183, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Rougeron, A.; Schuliar, G.; Leto, J.; Sitterl, E.; Landry, D.; Bougnoux, M.E.; Kobi, A.; Bouchara, J.P.; Giraud, S. Human-impacted areas of France are environmental reservoirs of the Pseudallescheria boydii/Scedosporium apiospermum species complex. Environ. Microbiol. 2015, 17, 1039–1048. [Google Scholar] [CrossRef]
- Guarro, J.; Kantarcioglu, A.S.; Horré, R.; Rodriguez-Tudela, J.L.; Estrella, M.C.; Berenguer, J.; De Hoog, G.S. Scedosporium apiospermum: Changing clinical spectrum of a therapy-refractory opportunist*. Med. Mycol. 2006, 44, 295–327. [Google Scholar] [CrossRef] [Green Version]
- Cortez, K.J.; Roilides, E.; Quiroz-Telles, F.; Meletiadis, J.; Antachopoulos, C.; Knudsen, T.; Buchanan, W.; Milanovich, J.; Sutton, D.A.; Fothergill, A.; et al. Infections Caused by Scedosporium spp. Clin. Microbiol. Rev. 2008, 21, 157–197. [Google Scholar] [CrossRef] [Green Version]
- Mello, T.P.; Bittencourt, V.C.B.; Liporagi-Lopes, L.C.; Aor, A.C.; Branquinha, M.H.; Santos, A.L. Insights into the social life and obscure side of Scedosporium/Lomentospora species: Ubiquitous, emerging and multidrug-resistant opportunistic pathogens. Fungal Biol. Rev. 2019, 33, 16–46. [Google Scholar] [CrossRef]
- van de Sande, W.W.J. Global Burden of Human Mycetoma: A Systematic Review and Meta-analysis. PLoS Negl. Trop. Dis. 2013, 7, e2550. [Google Scholar] [CrossRef] [Green Version]
- Lackner, M.; de Hoog, G.S.; Verweij, P.E.; Najafzadeh, M.J.; Curfs-Breuker, I.; Klaassen, C.H.; Meis, J.F. Species-Specific Antifun-gal Susceptibility Patterns of Scedosporium and Pseudallescheria Species. Antimicrob. Agents Chemother. 2012, 56, 2635–2642. [Google Scholar] [CrossRef] [Green Version]
- Lackner, M.; Hagen, F.; Meis, J.F.; Gerrits van den Ende, A.H.G.; Vu, D.; Robert, V.; Fritz, J.; Moussa, T.A.A.; de Hoog, G.S. Susceptibility and Diversity in the Therapy-Refractory Genus Scedosporium. Antimicrob. Agents Chemother. 2014, 58, 5877–5885. [Google Scholar] [CrossRef] [Green Version]
- Delhaes, L.; Harun, A.; Chen, S.C.; Nguyen, Q.; Slavin, M.; Heath, C.H.; Maszewska, K.; Halliday, C.; Robert, V.; Sorrell, T.C.; et al. Molecular Typing of AustralianScedosporiumIsolates Showing Genetic Variability and NumerousS. aurantiacum. Emerg. Infect. Dis. 2008, 14, 282–290. [Google Scholar] [CrossRef]
- Kaur, J.; Kautto, L.; Penesyan, A.; Meyer, W.; Elbourne, L.D.H.; Paulsen, I.T.; Nevalainen, H.K.M. Interactions of an Emerging Fungal Pathogen Scedosporium aurantiacum with Human Lung Epithelial Cells. Sci. Rep. 2019, 9, 5035. [Google Scholar] [CrossRef] [PubMed]
- Heath, C.; Slavin, M.; Sorrell, T.; Handke, R.; Harun, A.; Phillips, M.; Nguyen, Q.; Delhaes, L.; Ellis, D.; Meyer, W.; et al. Population-based surveillance for scedosporiosis in Australia: Epidemiology, disease manifestations and emergence of Scedosporium aurantiacum infection. Clin. Microbiol. Infect. 2009, 15, 689–693. [Google Scholar] [CrossRef]
- Nakamura, Y.; Suzuki, N.; Nakajima, Y.; Utsumi, Y.; Murata, O.; Nagashima, H.; Saito, H.; Sasaki, N.; Fujimura, I.; Ogino, Y.; et al. Scedosporium auranti-acum brain abscess after near-drowning in a survivor of a tsunami in Japan. Respir. Investig. 2013, 51, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Refojo, N.; Minervini, P.; Hevia, A.I.; Abrantes, R.A.; Fernández, J.; Apestey, N.; Garnero, M.; Villada, M.; Davel, G. Keratitis caused by moulds in Santa Lucía Ophthalmology Hospital in Buenos Aires, Argentina. Rev. Iberoam. Micol. 2016, 33, 1–6. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.T.J. Amplification and direct sequencing offungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Academic Press, Inc.: Cambridge, MA, USA, 1990. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Secuence Alignament Editor and Analysis Program for Windows 95/98/NT; ScienceOpen, Inc.: Burlington, MA, USA, 1999. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program distributed by the author. Evol. Biol. Cent. Uppsala Univ. 2004, 2, 1–2. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Color, 3rd ed.; Eyre Methuen: London, UK, 1978. [Google Scholar]
- CLSI Standars M38. Reference Method for Brtoth Dlution Anti-fungal Susceptibility Testing of Filamentous Fungi, 3rd ed.; Clinical and Laboratory Standars Institute, CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Gilgado, F.; Cano, J.; Gené, J.; Guarro, J. Molecular Phylogeny of the Pseudallescheria boydii Species Complex: Proposal of Two New Species. J. Clin. Microbiol. 2005, 43, 4930–4942. [Google Scholar] [CrossRef] [Green Version]
- de Hoog, G.S.; Guarro, J.; Gené, J.; Ahmed, S.; Al-Hatmi, A.M.S.; Figueras, M.J.; Vitale, R.G. Atlas of Clinical Fungi, 3rd ed.; Reus: Utrecht, The Netherlands, 2019. [Google Scholar]
- Elizondo-Zertuche, M.; Treviño-Rangel, R.d.J.; Robledo-Leal, E.; Luna-Rodríguez, C.E.; Martínez-Fierro, M.L.; Rodríguez-Sánchez, I.P.; González, G.M. Molecular identification and in vitro antifungal susceptibility of Scedosporium complex isolates from high human activity sites in Mexico. Mycologia 2017, 109, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, E.; Sanhueza, C. New record of Scedosporium dehoogii from Chile: Phylogeny and susceptibility profiles to classic and novel putative antifungal agents. Rev. Iberoam. Micol. 2016, 33, 224–229. [Google Scholar] [CrossRef]
- Gilgado, F.; Cano, J.; Gené, J.; Sutton, D.A.; Guarro, J. Molecular and Phenotypic Data Supporting Distinct Species Statuses for Scedosporium apiospermum and Pseudallescheria boydii and the Proposed New Species Scedosporium dehoogii. J. Clin. Microbiol. 2007, 46, 766–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alastruey-Izquierdo, A.; Cuenca-Estrella, M.; Monzón, A.; Tudela, J.L.R. Prevalence and Susceptibility Testing of New Species of Pseudallescheria and Scedosporium in a Collection of Clinical Mold Isolates. Antimicrob. Agents Chemother. 2006, 51, 748–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivero-Menendez, O.; Cuenca-Estrella, M.; Alastruey-Izquierdo, A. In vitro activity of APX001A against rare moulds us-ing EUCAST and CLSI methodologies. J. Antimicrob. Chemother. 2019, 74, 1295–1299. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Collection Accession Number | Source | GenBank Accession Number | |
|---|---|---|---|---|
| BT2 | ITS | |||
| Scedosporium boydii | DMic 85073 | No data, 1985 | MT813143 | MT803004 |
| Scedosporium boydii | DMic 90394 | No data, 1990 | MT813144 | MT803005 |
| Scedosporium boydii | DMic 993530 | Lung tissue, 1999 | MT813145 | MT803006 |
| Scedosporium angustum | DMic 01651 | No data, 2001 | MT813146 | MT803007 |
| Scedosporium dehoogii | DMic 01867 | Corneal abscess, 2001 * | MT813147 | MT803008 |
| Scedosporium angustum | DMic 031321 | No data, 2003 | MT813148 | MT803009 |
| Scedosporium boydii | DMic 062875 | Lung tissue, 2006 | MT813149 | MT803010 |
| Scedosporium boydii | DMic 073201 | Sputum, 2007 | MT813150 | MT803011 |
| Scedosporium boydii | DMic 083385 | Sputum, 2008 | MT813151 | MT803012 |
| Scedosporium boydii | DMic 083389 | Lung tissue, 2008 | MT813152 | MT803013 |
| Scedosporium apiospermum | DMic 083404 | Corneal abscess, 2008 | MT813153 | MT803014 |
| Scedosporium boydii | DMic 093482 | Lung tissue, 2009 | MT813154 | MT803015 |
| Scedosporium apiospermum | DMic 093510 | Corneal abscess, 2009 | MT813155 | MT803016 |
| Scedosporium boydii | DMic 103859 | Sputum, 2010 | MT813156 | MT803017 |
| Scedosporium boydii | DMic 103731 | Corneal abscess, 2010 | MT813157 | MT803018 |
| Scedosporium boydii | DMic 103813 | Corneal abscess, 2010 | MT813158 | MT803019 |
| Scedosporium apiospermum | DMic 114031 | Corneal abscess, 2011 | MT813159 | MT803020 |
| Scedosporium boydii | DMic 114032 | Bronchoalveolar lavage, 2011 | MT813160 | MT803021 |
| Scedosporium angustum | DMic 114102 | Subcutaneous mycosis (foot), 2011 | MT813161 | MT803022 |
| Scedosporium boydii | DMic 134638 | Sputum, 2013 | MT813162 | MT803023 |
| Scedosporium boydii | DMic 165274 | Subcutaneous mycosis (leg), 2013 | MT813163 | MT803024 |
| Scedosporium boydii | DMic 134375 | Sputum, 2014 | MT813164 | MT803025 |
| Scedosporium angustum | DMic 165275 | Corneal abscess, 2014 | MT813165 | MT803026 |
| Scedosporium boydii | DMic 165279 | Tracheal aspirate, 2014 | MT813166 | MT803027 |
| Scedosporium boydii | DMic 165033 | Bronchoalveolar lavage, 2015 | MT813167 | MT803028 |
| Scedosporium apiospermum | DMic 165035 | Subcutaneous mycosis (arm), 2015 | MT813168 | MT803029 |
| Scedosporium apiospermum | DMic 165036 | Corneal abscess, 2015 | MT813169 | MT803030 |
| Scedosporium apiospermum | DMic 165286 | Subcutaneous mycosis (hand), 2015 | MT813170 | MT803031 |
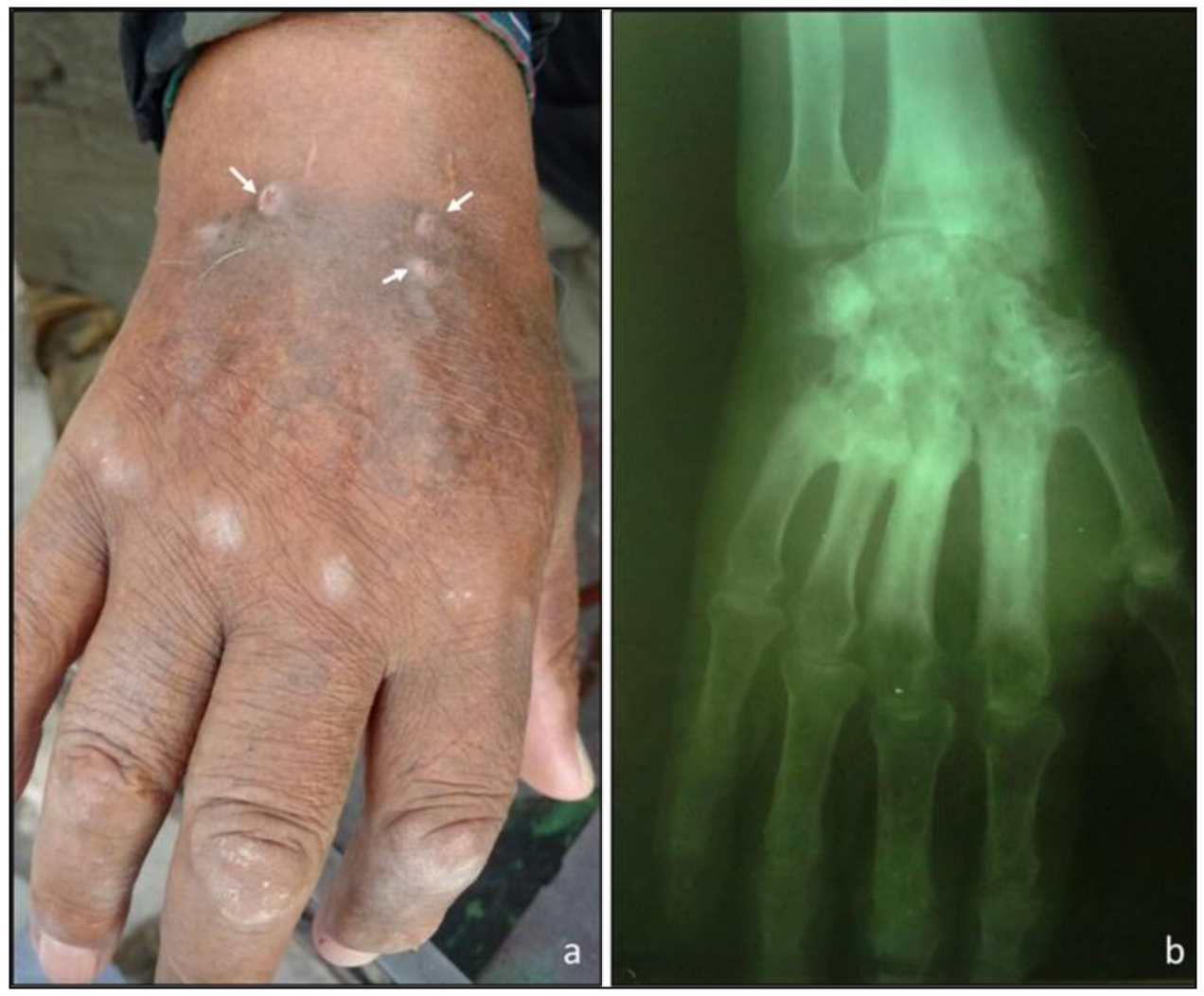
| Scedosporium americanum sp. nov. | DMic 165285 | Subcutaneous mycosis (hand), 2016 | MT813171 | MT803032 |
| Scedosporium apiospermum | DMic 165288 | Bronchoalveolar lavage, 2016 | MT813172 | MT803033 |
| Scedosporium apiospermum | DMic 165290 | Bronchoalveolar lavage, 2016 | MT813173 | MT803034 |
| Scedosporium apiospermum | DMic 165291 | Brain biopsy, 2016 | MT813174 | MT803035 |
| Scedosporium boydii | DMic 165341 | Sputum, 2016 | MT813175 | MT803036 |
| Scedosporium aurantiacum | DMic 175378 | Sputum, 2016 | MT813176 | MT803037 |
| Scedosporium apiospermum | DMic 175421 | Subcutaneous mycosis, turtle shell **, 2016 | MT813177 | MT803038 |
| Scedosporium apiospermum | DMic 175525 | Sputum, 2017 | MT813178 | MT803039 |
| Scedosporium boydii | DMic 175526 | Sputum, 2017 | MT813179 | MT803040 |
| Scedosporium boydii | DMic 175583 | Sputum, 2017 | MT813180 | MT803041 |
| Scedosporium angustum | DMic 175584 | Sputum, 2017 | MT813181 | MT803042 |
| Scedosporium boydii | DMic 175585 | Brain biopsy, 2017 | MT813182 | MT803043 |
| Scedosporium aurantiacum | DMic 175588 | Sputum, 2017 | MT813183 | MT803044 |
| Scedosporium boydii | DMic 195868 | Sputum, 2017 | MT813184 | MT803045 |
| Scedosporium apiospermum | DMic 175678 | Tracheal aspirate, 2017 | MT813185 | MT803046 |
| Scedosporium boydii | DMic 195875 | Bronchoalveolar lavage, 2018 | MT813186 | MT803047 |
| Scedosporium boydii | DMic 195871 | Bx cutanea, 2018 | MT813187 | MT803048 |
| Scedosporium boydii | DMic 195874 | Sputum, 2018 | MT813188 | MT803049 |
| Scedosporium boydii | DMic 195869 | Sputum, 2018 | MT813189 | MT803050 |
| Scedosporium boydii | DMic 195877 | Sputum, 2018 | MT813190 | MT803051 |
| Scedosporium boydii | DMic 195878 | Cerebrospinal fluid, 2018 | MT813191 | MT803052 |
| Scedosporium sp. | CBS 218.35 | Human mycetoma pedis, Paraguay | MT813192 | AM712309 |
| Parascedosporium tectonae | CBS 120.338 | Garden soil Buenos Aires, Argentina | AM409105 | AM409113 |
| Petriellopsis africana | CBS 311.72 | Brown sandy soil, Namibia | AJ889603 | AJ888425 |
| Scedosporium angustum | CBS 254.72 | Half-digested-sewage tank Ohio, USA | AJ889604 | AJ888414 |
| Scedosporium boydii | CBS 101.22 | Mycetoma Texas, USA | AJ889590 | AJ888435 |
| Scedosporium desertorum | CBS 489.72 | Salt-marsh soil, Kuwait | AM409106 | AM409101 |
| Scedosporium ellipsoideum | CBS 418.73 | Soil, Tayikistán | AJ889595 | AJ888426 |
| Scedosporium fusoideum | CBS 106.53 | Goat dung Aligarh, India | AJ889601 | AJ888428 |
| Scedosporium apiospermum | FMR 8619 | Keratitis, Brazil | AJ889584 | NR130664 |
| Scedosporium minutisporum | FMR 4072 | River sediment Tordera river, Spain | AJ889592 | AJ888384 |
| Scedosporium aurantiacum | FMR 8630 | Ankle ulcer S. Compostela, Spain | AJ889597 | AJ888440 |
| Scedosporium dehoogii | CBS 117406 | Garden soil, Barcelona, Spain | KT163401 | KT163400 |
| Scedosporium cereisporum | FMR 12996 | Wastewater sludge, Mûrs-Erignés, France | KJ599659 | KJ599660 |
| Lomentospora prolificans | FMR 7294 | Clinical California, USA | AJ889591 | AJ888444 |
| Petriella setifera | CBS 385.87 | Human toe nail Helsinki, Finland | EU977491 | AY882345 |
| Petriella sordida | UTHSC 03-394 | Nasal infection California, USA | AM409104 | AM409114 |
| Strains | MIC Parameters | Antifungal Drugs (mg/L) | ||||||
|---|---|---|---|---|---|---|---|---|
| AMB | ITR | VOR | POS | ANI * | CAS * | MICA * | ||
| S. boydii (n = 28) | GM | 1.52 | 0.34 | 0.15 | 0.16 | 0.53 | 0.44 | 0.19 |
| MIC 50 | 2 | 0.5 | 0.125 | 0.25 | 1 | 1 | 0.25 | |
| MIC 90 | 8 | 4 | 0.25 | 1 | 4 | 4 | 2 | |
| Range | 0.032–8 | 0.008–8 | 0.015–0.5 | 0.008–1 | 0.008–8 | 0.008–8 | 0.008–8 | |
| n (%) | 17 (60.7) | 5 (17.8) | 1 (3.5) | 0 (0) | 11(39.2) | 12(42.8) | 3 (10.7) | |
| S. apiospermum (n = 12) | GM | 2.52 | 1.41 | 0.35 | 0.44 | 1.69 | 1.20 | 0.66 |
| MIC 50 | 4 | 1 | 0.5 | 0.5 | 4 | 1 | 0.5 | |
| MIC 90 | 8 | 8 | 0.5 | 1 | 8 | 16 | 8 | |
| Range | 0.5–16 | 0.125–16 | 0.125–4 | 0.125–16 | 0.032–16 | 0.032–16 | 0.064–16 | |
| n (%) | 8 (66.6) | 5 (41.6) | 1 (8.3) | 1 (8.3) | 8 (66.6) | 5 (41.6) | 3 (25) | |
| S. angustum (n = 5) | GM | 2.30 | 1.15 | 0.28 | 0.43 | 2.30 | 2 | 0.32 |
| Range | 1–8 | 0.25–8 | 0.064–1 | 0.064–2 | 1–4 | 0.5–4 | 0.125–0.5 | |
| n (%) | 3 (60) | 2 (40) | 0 (0) | 1 (20) | 4 (80) | 4 (80) | 0 (0) | |
| S. aurantiacum (n = 2) | DMic 175378 | 16 | 2 | 0.5 | 1 | 8 | 16 | 8 |
| DMic 175588 | 8 | 4 | 0.5 | 1 | 4 | 8 | 8 | |
| n (%) | 2 (100) | 2 (100) | 0 (0) | 0 (0) | 2 (100) | 2 (100) | 2 (100) | |
| S. americanum sp. nov. (n = 2) | DMic 165285 | 8 | 1 | 0.5 | 1 | 1 | 2 | 0.5 |
| CBS 128.35 | 4 | 0.5 | 0.25 | 0.25 | 2 | 1 | 0.25 | |
| n (%) | 2 (100) | 0 (0) | 0 (0) | 0 (0) | 1 (50) | 1 (50) | 0 (0) | |
| S. dehoogii (n = 1) | DMic 01867 | 8 | 0.25 | 0.25 | 0.25 | 4 | 4 | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abrantes, R.A.; Refojo, N.; Hevia, A.I.; Fernández, J.; Isla, G.; Córdoba, S.; Dávalos, M.F.; Lubovich, S.; Maldonado, I.; Davel, G.O.; et al. Scedosporium spp. from Clinical Setting in Argentina, with the Proposal of the New Pathogenic Species Scedosporium americanum. J. Fungi 2021, 7, 160. https://doi.org/10.3390/jof7030160
Abrantes RA, Refojo N, Hevia AI, Fernández J, Isla G, Córdoba S, Dávalos MF, Lubovich S, Maldonado I, Davel GO, et al. Scedosporium spp. from Clinical Setting in Argentina, with the Proposal of the New Pathogenic Species Scedosporium americanum. Journal of Fungi. 2021; 7(3):160. https://doi.org/10.3390/jof7030160
Chicago/Turabian StyleAbrantes, Ruben A., Nicolás Refojo, Alejandra I. Hevia, Julián Fernández, Guillermina Isla, Susana Córdoba, María F. Dávalos, Silvina Lubovich, Ivana Maldonado, Graciela O. Davel, and et al. 2021. "Scedosporium spp. from Clinical Setting in Argentina, with the Proposal of the New Pathogenic Species Scedosporium americanum" Journal of Fungi 7, no. 3: 160. https://doi.org/10.3390/jof7030160