
Biodiversity of Lignicolous Freshwater Hyphomycetes from China and Thailand and Description of Sixteen Species

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection, Isolation and Morphology
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
3. Results
3.1. Taxonomy
3.1.1. Dothideomycetes O.E. Erikss. and Winka, Myconet 1(1): 5 (1997)
- Pleosporales Luttr. ex M.E. Barr 1987
- Pseudoberkleasmiaceae Phukhams. and K.D. Hyde
- Pseudoberkleasmium Tibpromma and K.D. Hyde
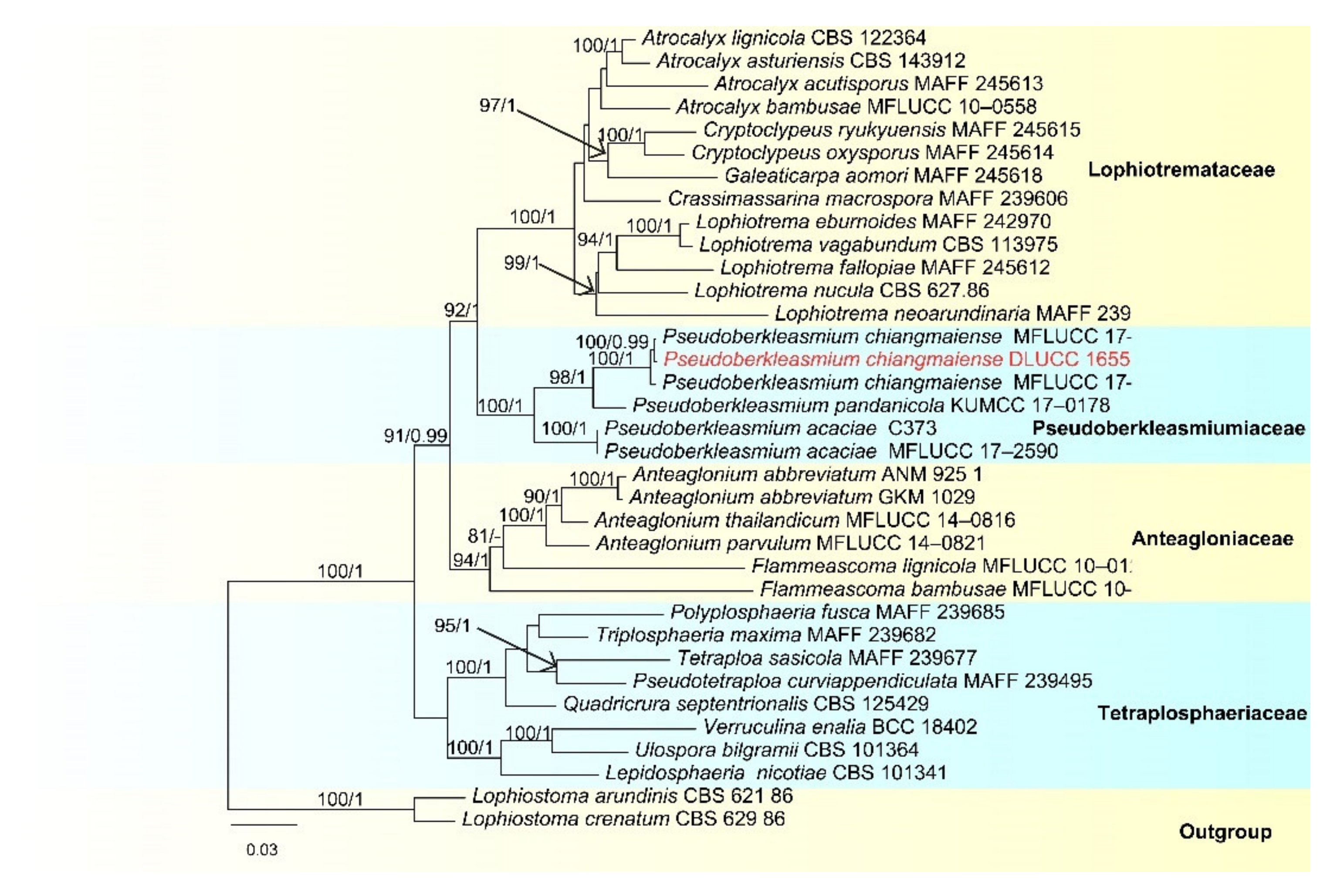
- Pseudoberkleasmium chiangmaiense Y.Z. Lu and K.D. Hyde, in Hyde et al., Fungal Diversity 96:38 (Figure 1).
- Tetraplosphaeriaceae Kaz. Tanaka and K. Hiray
- Tetraploa Berk. and Broome
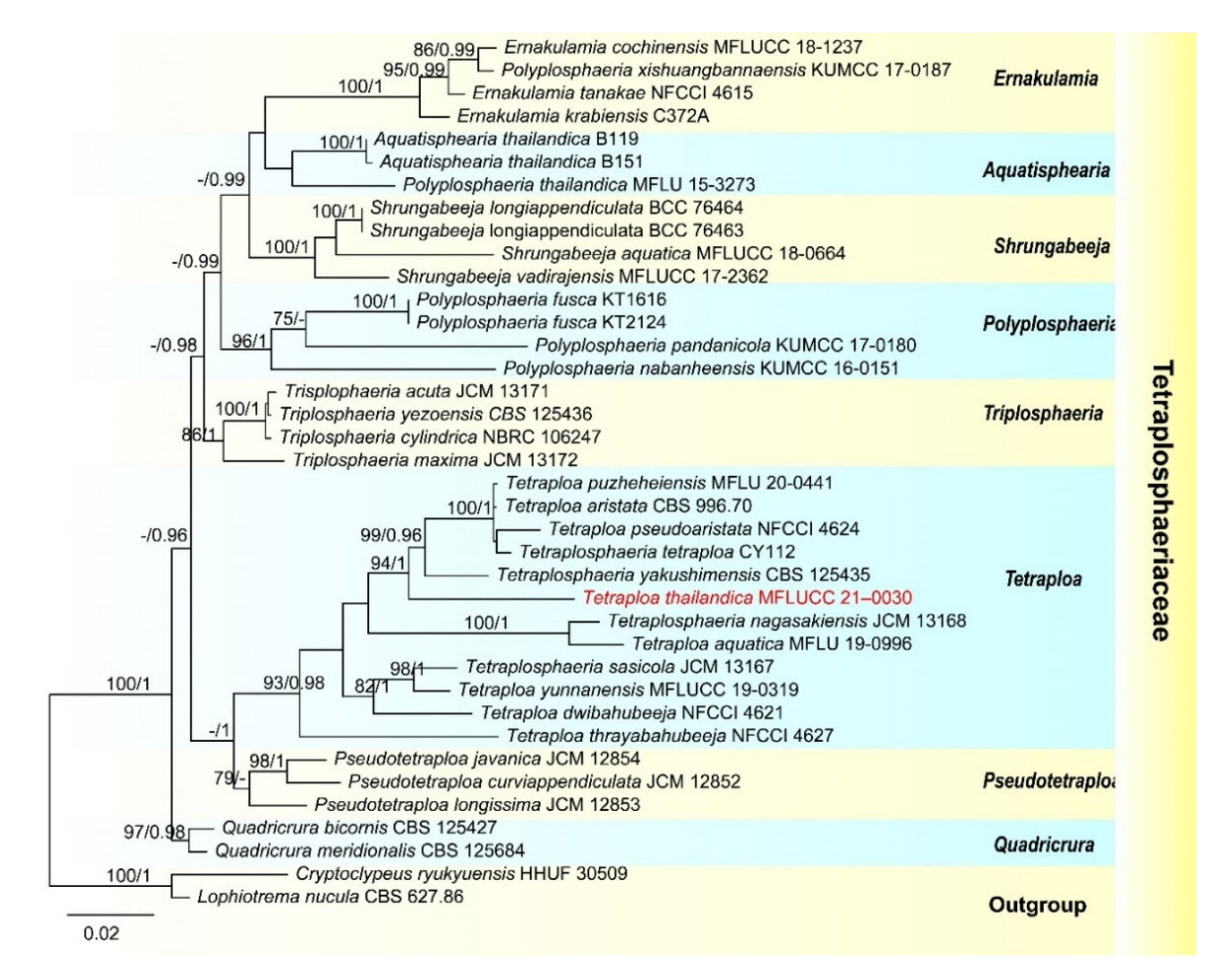
- Tetraploa thailandica D.F. Bao, H.Y. Su, K.D. Hyde and Z.L. Luo, sp. nov. (Figure 3).
- Index Fungorum number: IF558591; Facesoffungi number: FoF 09915
- Holotype: MFLU 21–0030
- Etymology—Referring to Thailand, where the fungus was collected.
- Pleomonodictydaceae Hern.-Restr., J. Mena and Gené
- Pleomonodictys Hern.-Restr., J. Mena and Gené
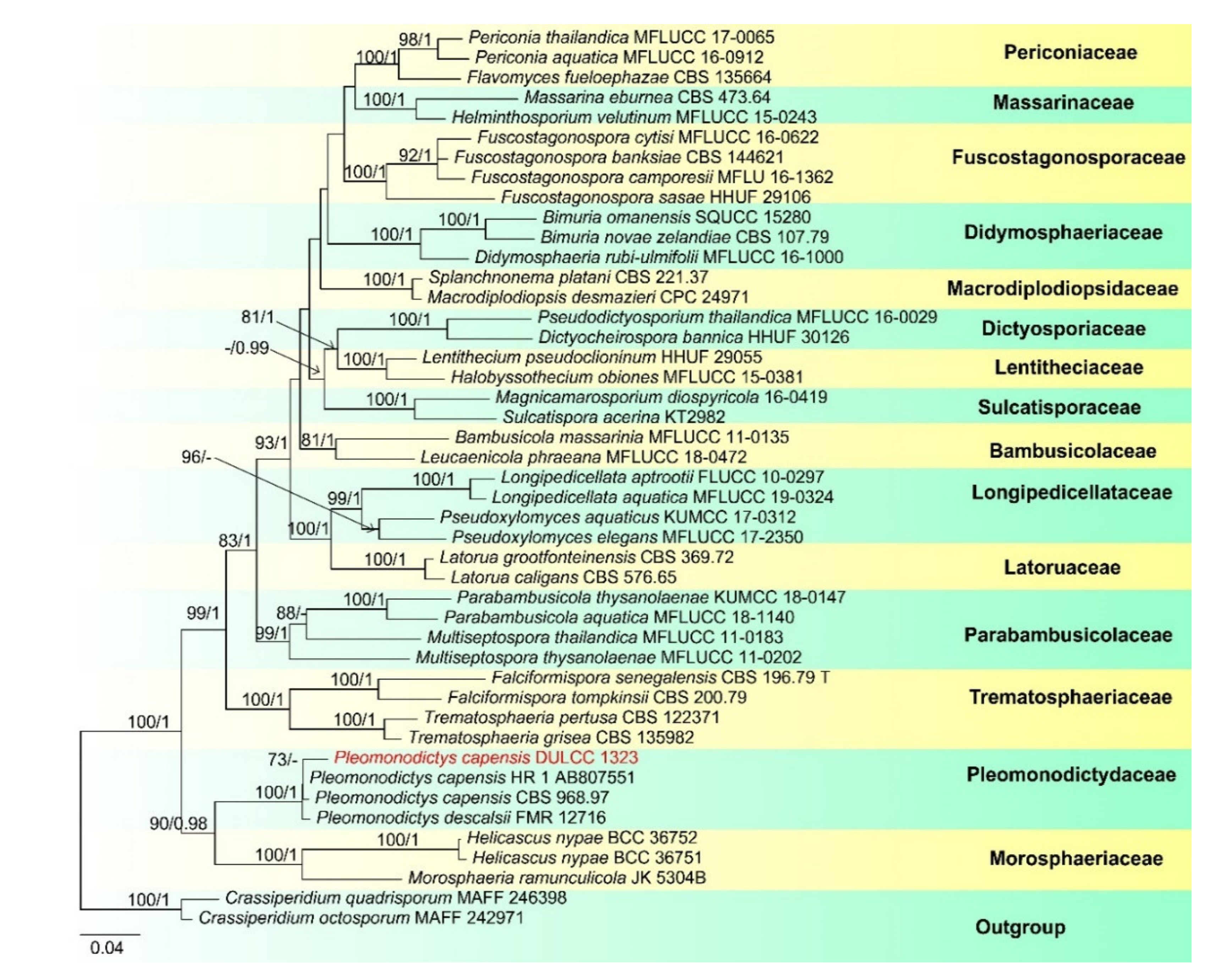
- Pleomonodictys capensis (R.C. Sinclair, Boshoff and Eicker) Hern.-Restr., J. Mena and Gené, Studies in Mycology 86:77 (2017) (Figure 5).
3.1.2. Sordariomycetes
- Diaporthomycetidae
- Distoseptisporales Z.L. Luo, K.D. Hyde and H.Y. Su
- Aquapteridosporaceae K.D. Hyde and Hongsanan
- Aquapteridospora J. Yang, K.D. Hyde and Maharachch
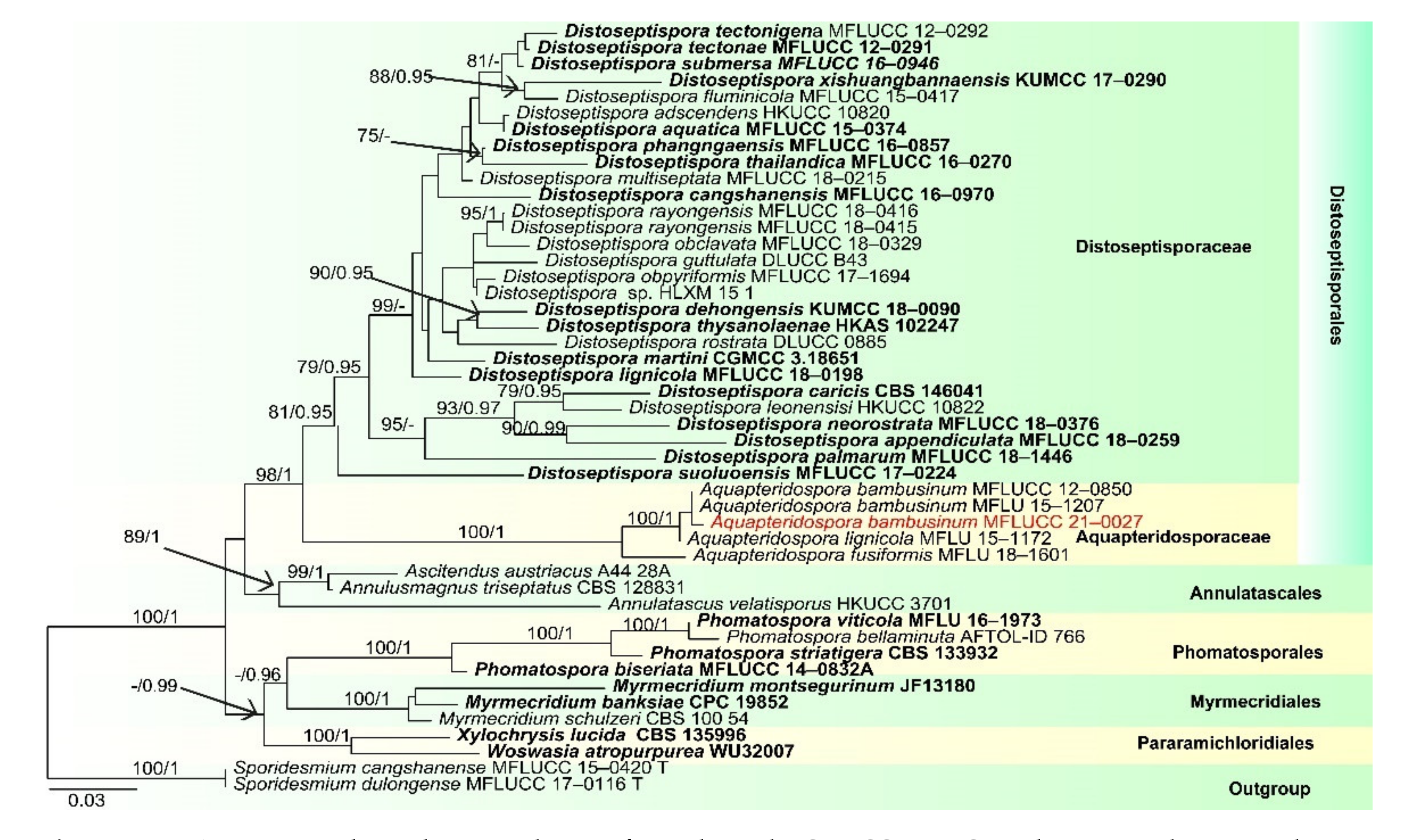
- Aquapteridospora bambusinum (D.Q. Dai and K.D. Hyde) D.F. Bao, com. nov. (Figure 7).
- ≡Pleurophragmium bambusinum D.Q. Dai and K.D. Hyde, in Dai et al., Fungal Diversity 82:92
- Index Fungorum number: IF 558592
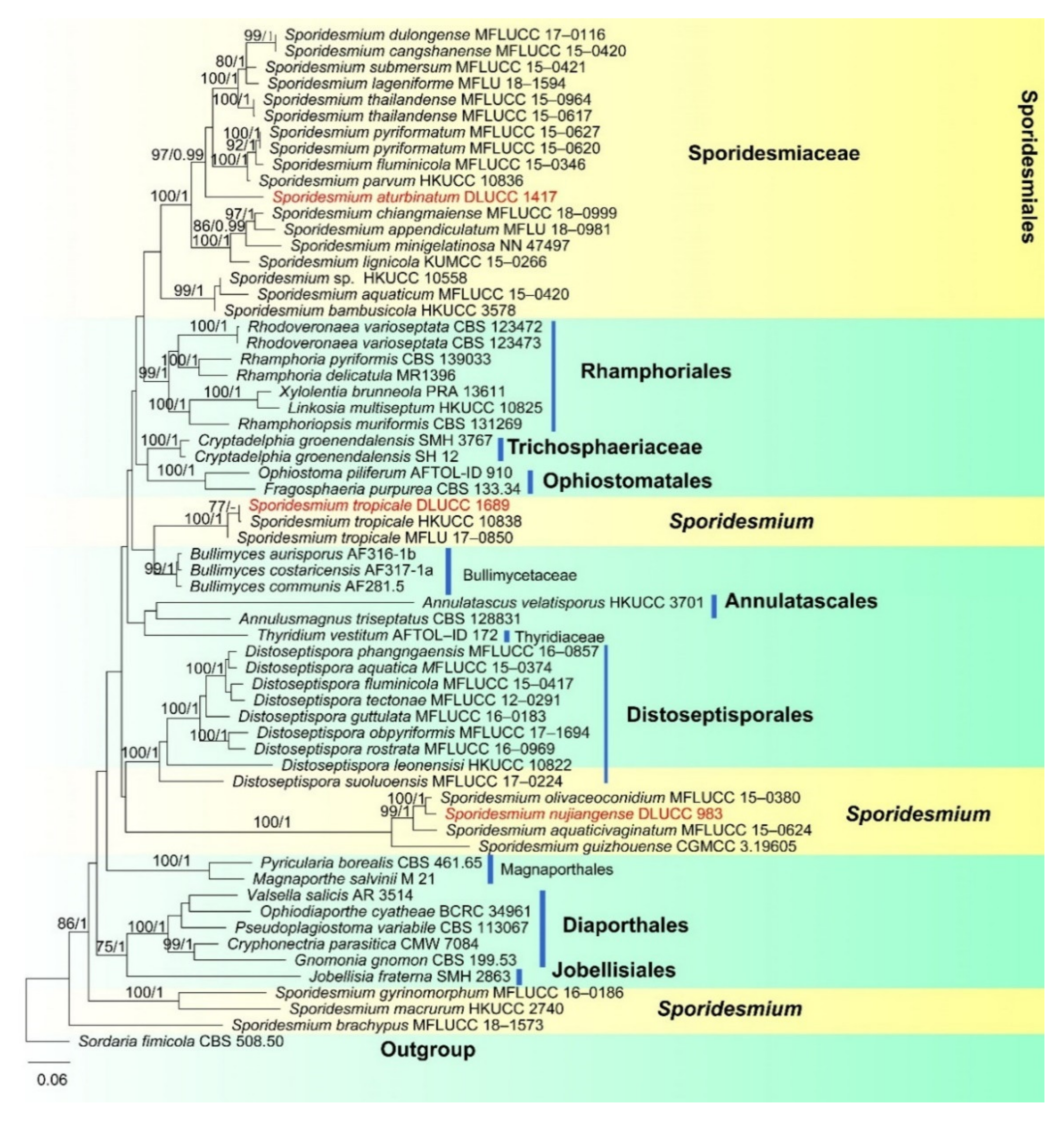
- Sporidesmiales Crous
- Sporidesmiaceae Fr., Summa veg
- Sporidesmium Link, Mag. Gesell. naturf.
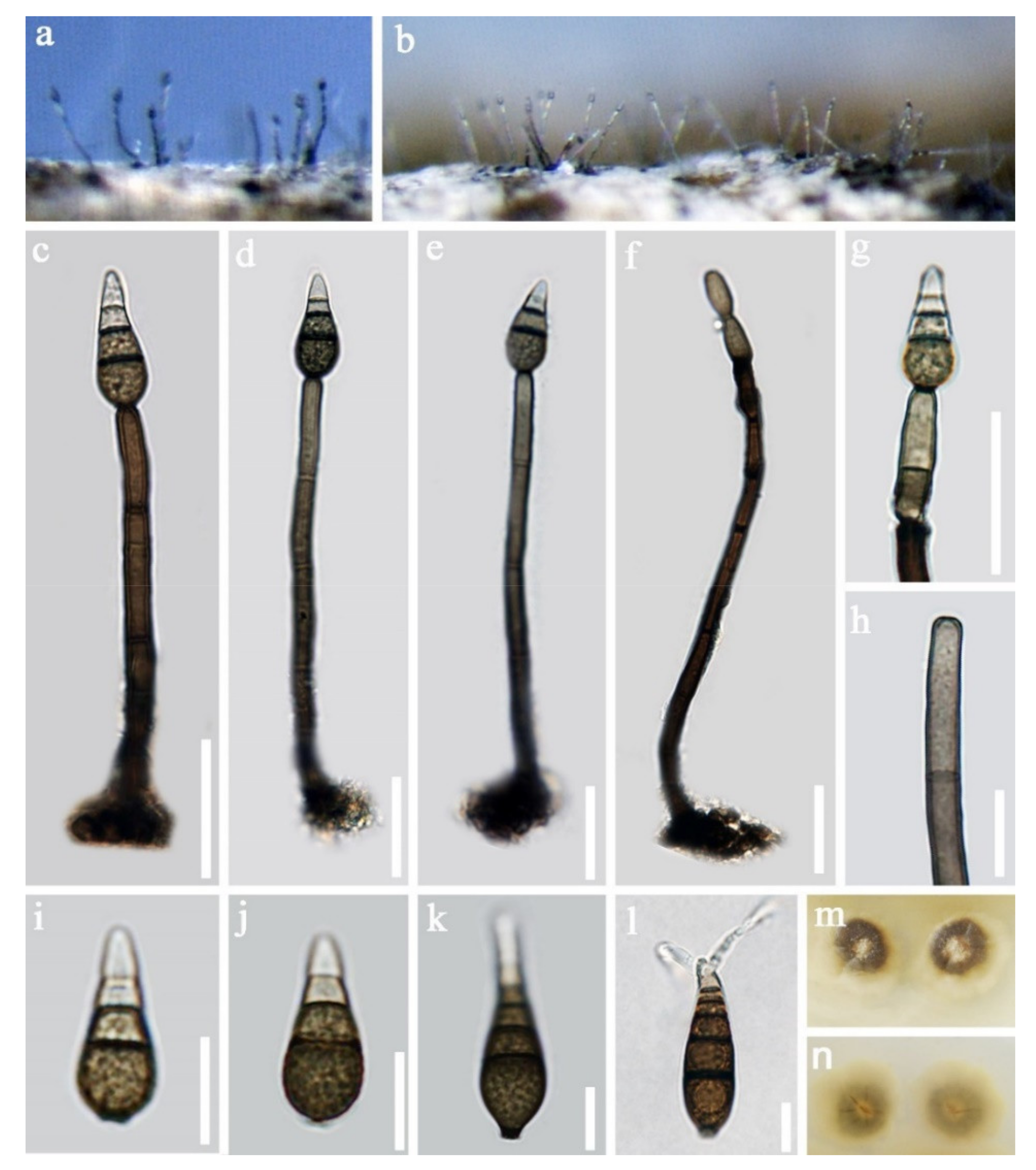
- Sporidesmium aturbinatum (S. Hughes) M.B. Ellis, Mycol. Pap. 70: 49 (1958). (Figure 9).
- Sporidesmium nujiangense D.F. Bao, H.Y. Su, K.D. Hyde and Z.L. Luo, sp. nov. (Figure 12).
- Index Fungorum number: IF558594; Faces of fungi number: FoF 09916
- Holotype—HKAS 115795
- Etymology—Referring to the place “Nujiang River” where this species was collected.
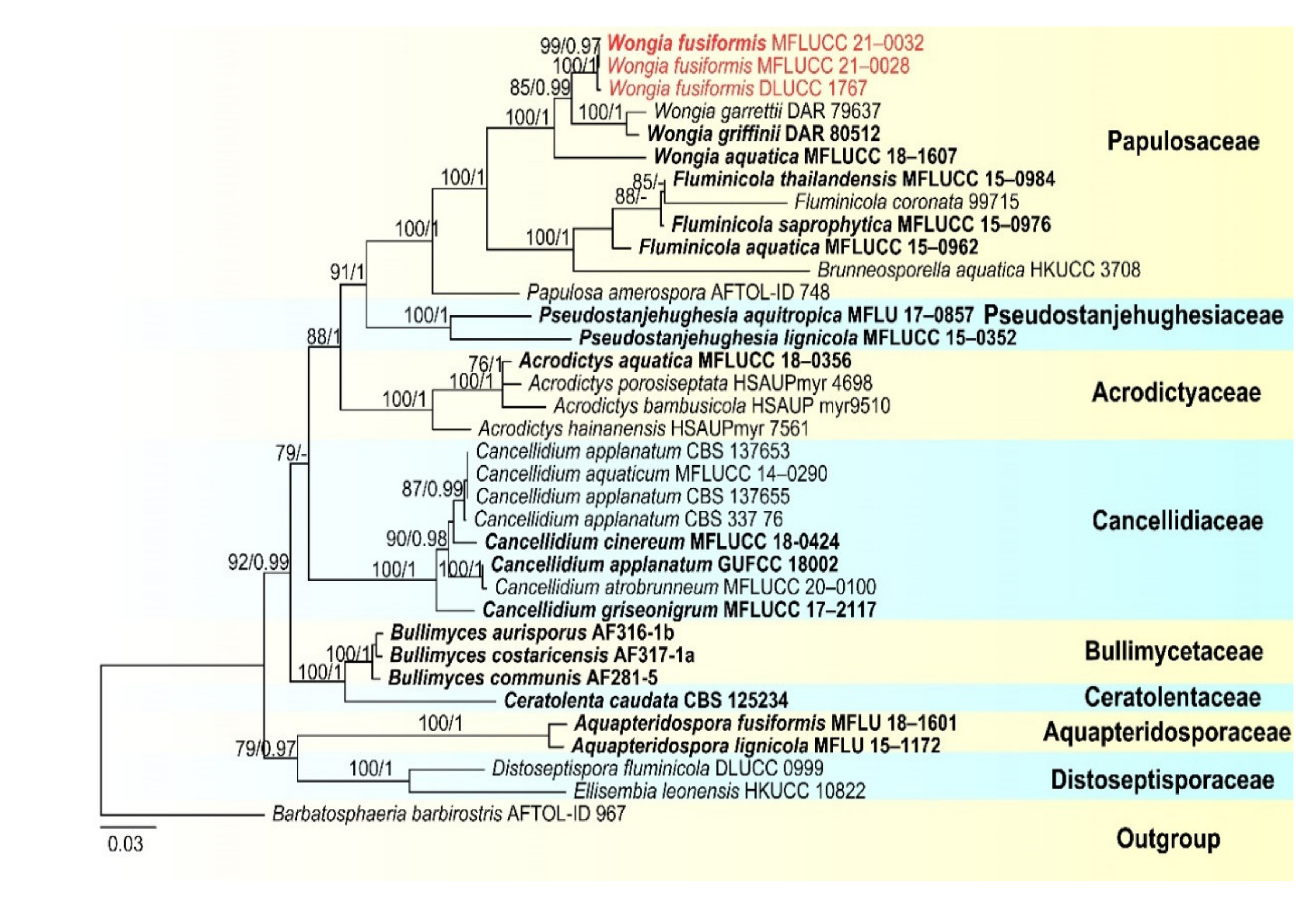
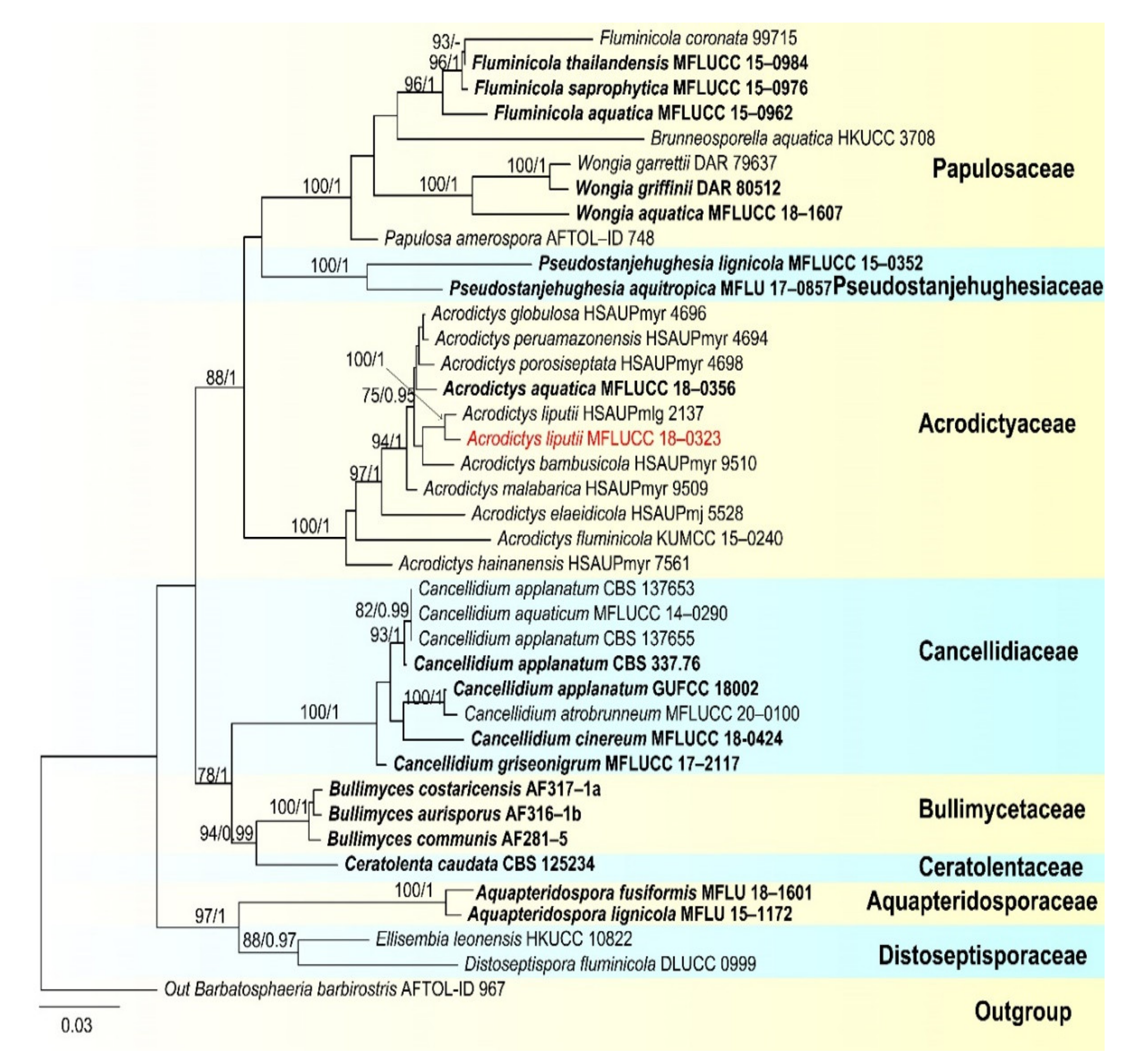
- Papulosaceae Winka and O.E. Erikss., Mycoscience 41 (2):102 (2000)
- Wongia Khemmuk, Geering and R.G. Shivas
- Wongia fusiformis D.F. Bao, H.Y. Su, K.D. Hyde and Z.L. Luo, sp. nov. (Figure 13).
- Index Fungorum number: IF558595; Facesoffungi number: FoF 09917
- Holotype—MFLU 21–0028
- Etymology—Referring to the fusiform conidia of this fungus
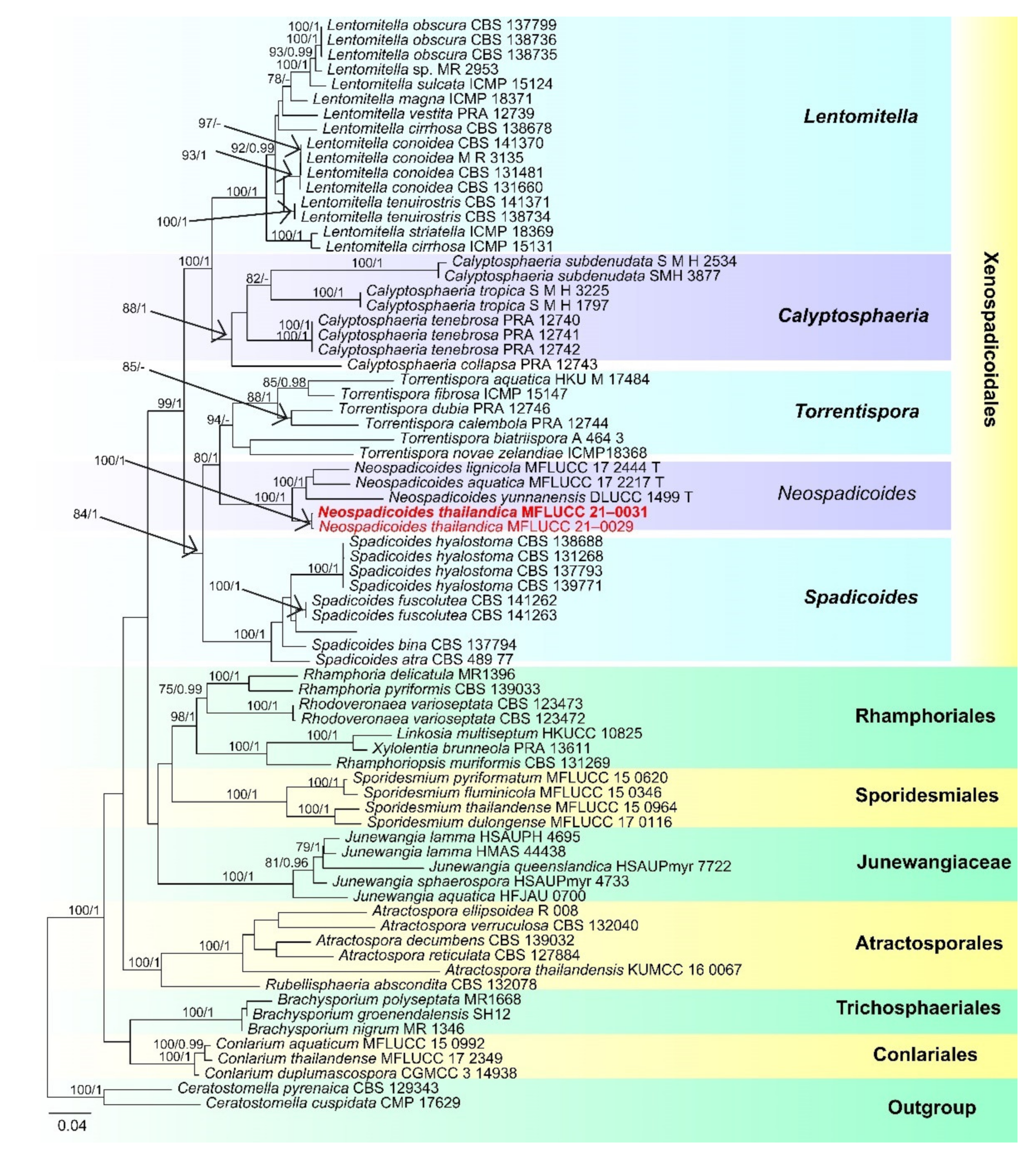
- Xenospadicoidales Hern.-Restr., J. Mena and Gené
- Xenospadicoidaceae Hern.-Restr., J. Mena and Gené
- Neospadicoides thailandica D.F. Bao, H.Y. Su, K.D. Hyde and Z.L. Luo, sp. nov. (Figure 15).
- Index Fungorum number: IF 558596; Faces of fungi number: FoF 09918
- Holotype: MFLU 21–0032
- Etymology: Referring to the species was collected from Thailand.
- Acrodictyaceae J.W. Xia and X.G. Zhang
- Acrodictys M.B. Ellis
- Acrodictys liputii L. Cai, K.Q. Zhang, McKenzie, W.H. Ho and K.D. Hyde, Nova Hedwigia 75 (3–4): 526, 2002. (Figure 17).
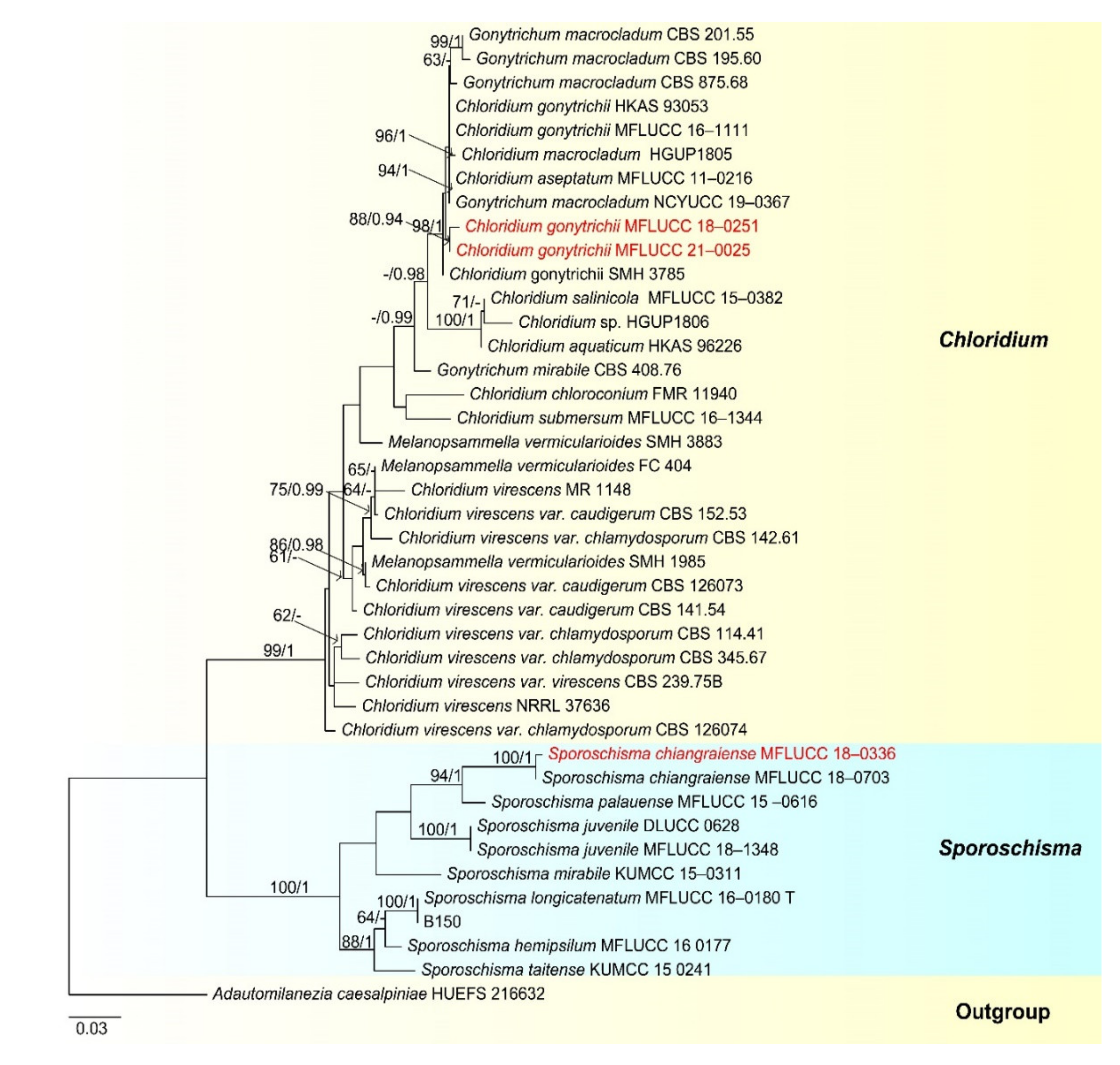
- Chaetosphaeriaceae Réblová, M.E. Barr and Samuels, Sydowia 51(1): 56 (1999)
- Chloridium Link, Mag. Gesell. naturf. Freunde, Berlin 3(1–2): 13 (1809)
- Chloridium gonytrichii (F.A. Fernández and Huhndorf) Réblová & Seifert, in Réblová et al., IMA Fungus 7(1): 134 (2016) (Figure 19).
- ≡ Gonytrichum macrocladum (Sacc.) S. Hughes, Trans. Br. Mycol. Soc. 34(4): 565 (1952) (1951)
- ≡ Chloridium aseptatum M.J. Wei and H. Zhang, in Wei, Zhang, Dong, Boonmee & Zhang, Phytotaxa 362(2): 191 (2018)
- ≡ Chloridium macrocladum (Sacc.) Karun, Maharachch., C.H.Kuo and K.D.Hyde, in Yuan et al., Fungal Diversity [74] (2020)
- Sporoschisma Berk. & Broome
- Sporoschisma chiangraiense N.G. Liu and K.D. Hyde, Fungal Diversity 96: 160 (2019) (Figure 21).
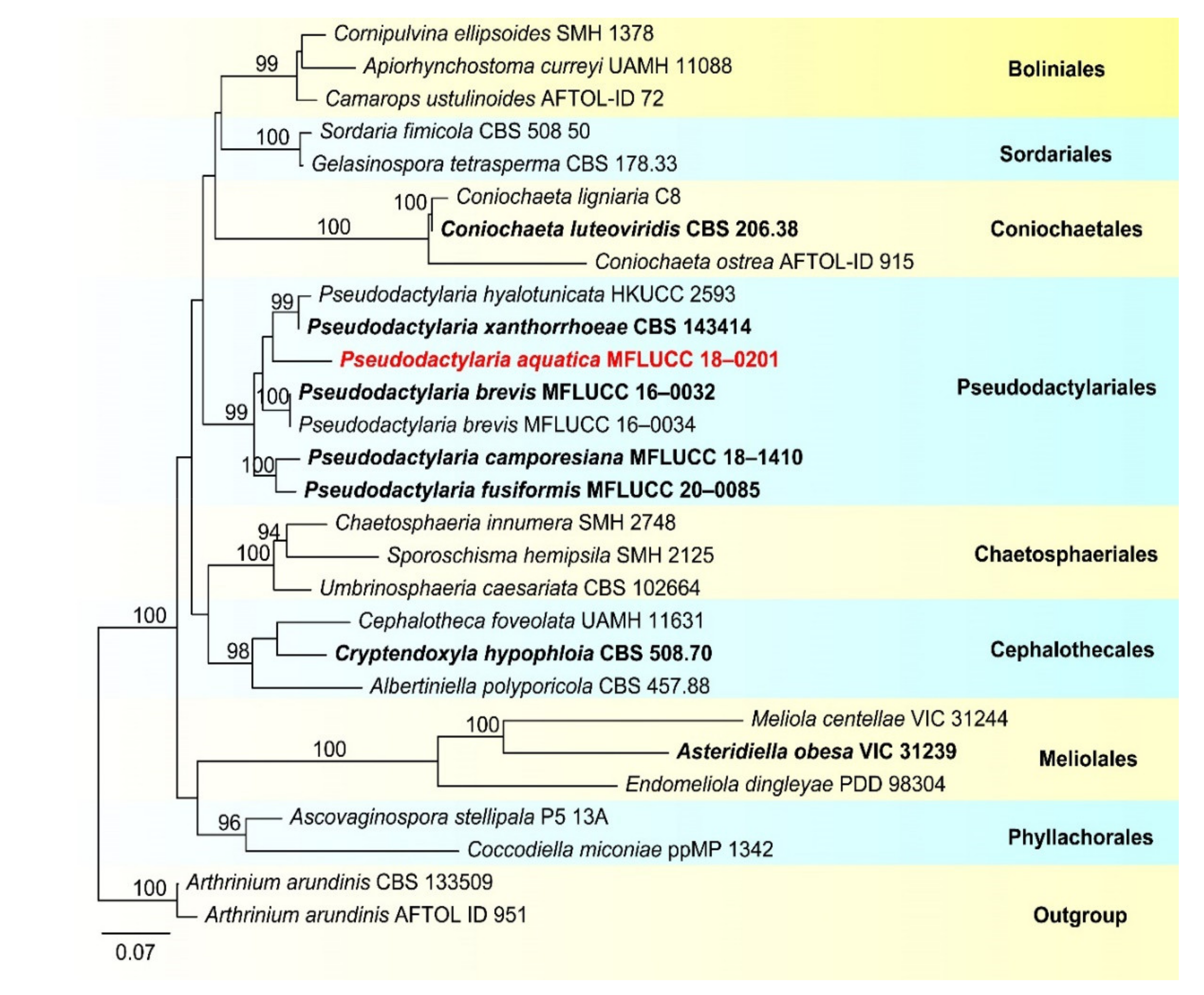
- Pseudodactylariales Crous,
- Pseudodactylariaceae Crous
- Pseudodactylaria Crous
- Pseudodactylaria aquatica D.F. Bao, H.Y. Su, K.D. Hyde and Z.L. Luo, sp. nov. (Figure 23).
- Index Fungorum number: IF 558597; Faces of fungi number: FoF 09919
- Holotype—MFLU 21–0037
- Etymology—Referring the fungus was collected form aquatic habitat.
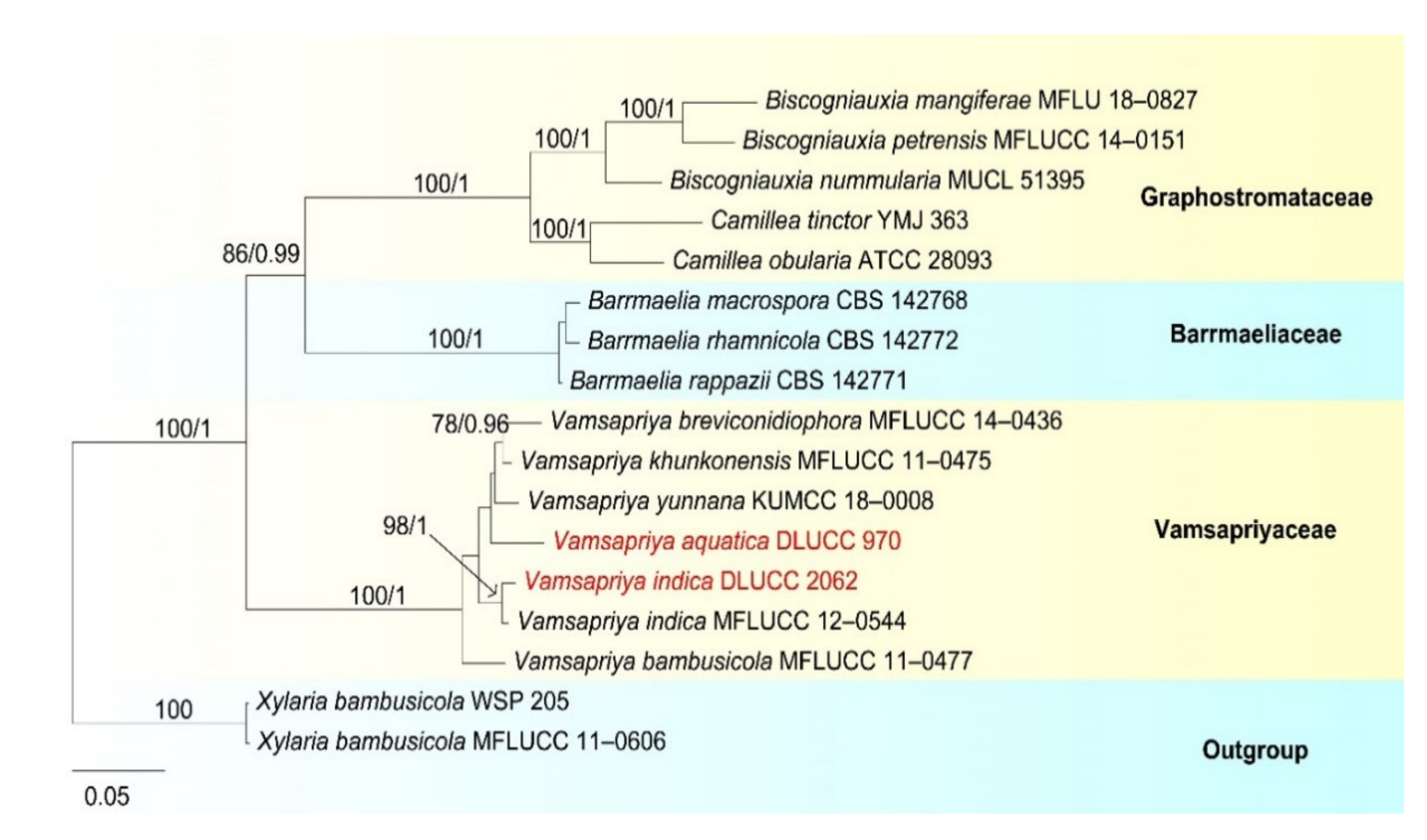
- Xylariales Nannf., Nova Acta R. Soc
- Xylariaceae Tul. and C. Tul
- Vamsapriya Gawas and Bhat, Mycotaxon 94: 150 (2006)
- Vamsapriya aquatica D.F. Bao, H.Y. Su, K.D. Hyde and Z.L. Luo, sp. nov. (Figure 25).
- Index Fungorum number: IF 558598; Faces of fungi number: FoF 09920
- Holotype—HKAS 115791
- Etymology—Referring the fungus was collected form aquatic habitat.
- Vamsapriya indica Gawas & Bhat, Mycotaxon 94: 150 (Figure 27).
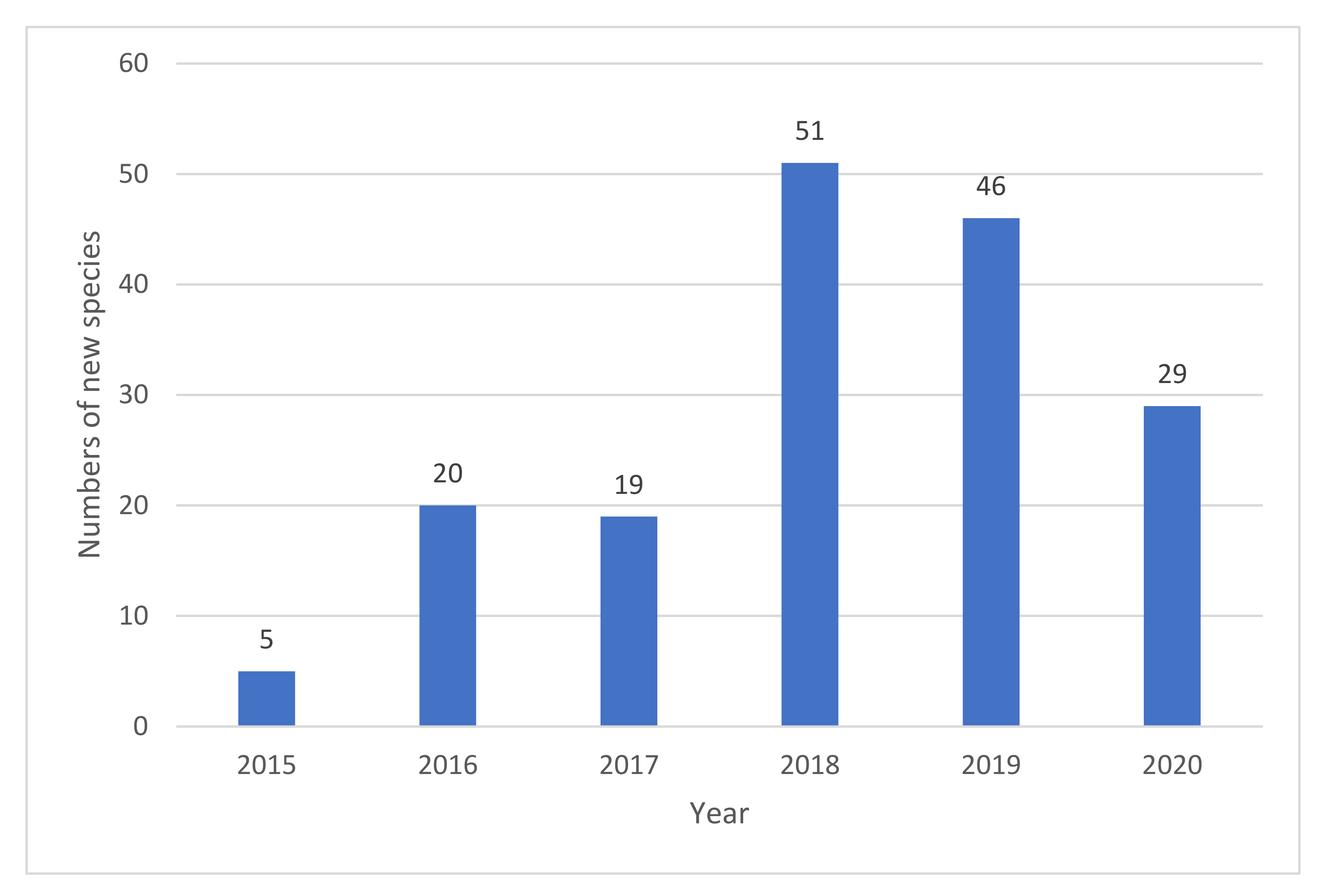
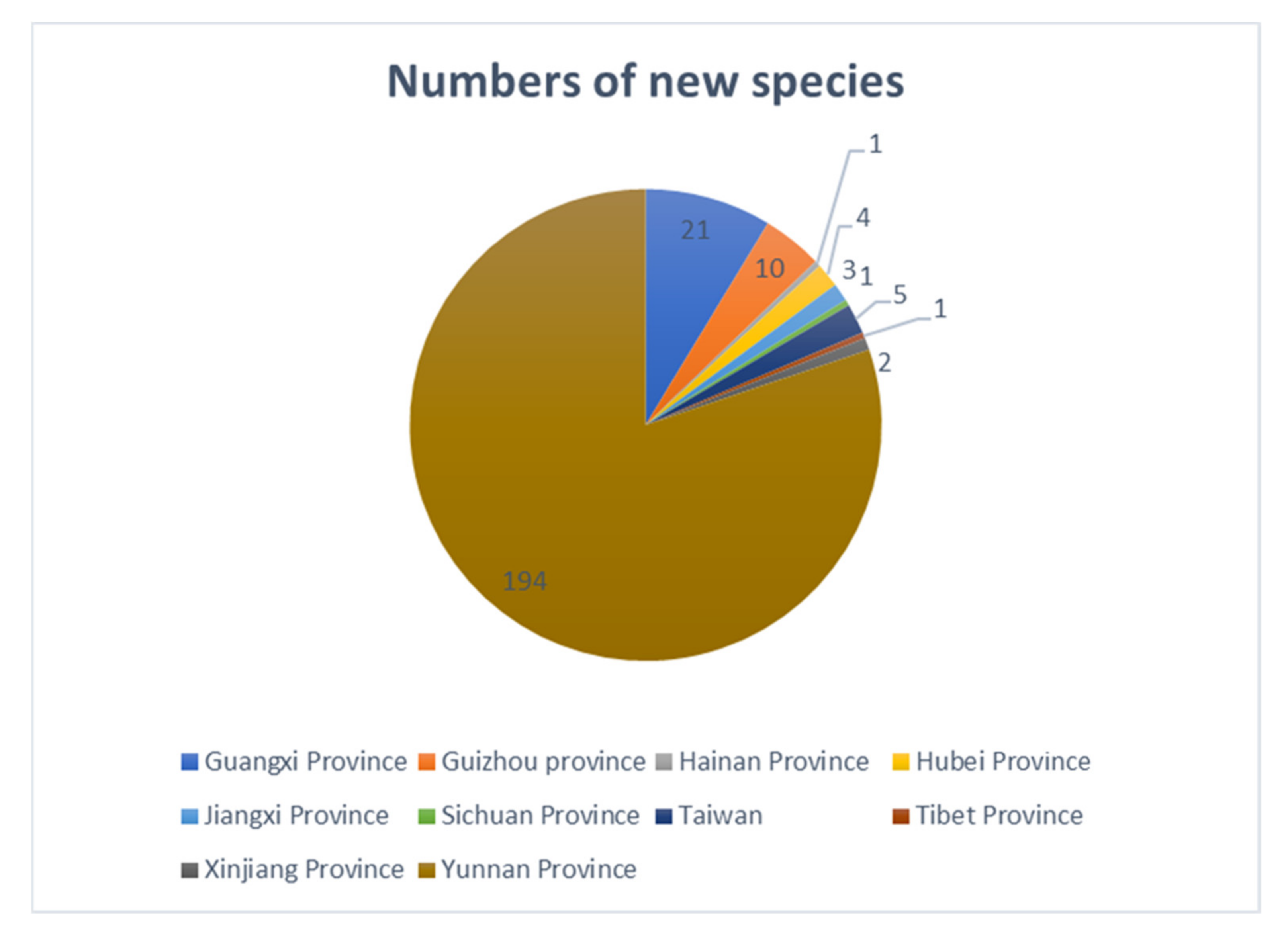
3.2. Biodiversity of Freshwater Fungi in China
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shearer, C.A.; Descals, E.; Kohlmeyer, B.; Kohlmeyer, J.; Marvanová, L.; Padgett, D.; Porter, D.; Raja, H.A.; Schmit, J.P.; Thorton, H.A.; et al. Fungal biodiversity in aquatic habitats. Biodivers. Conserv. 2007, 16, 49–67. [Google Scholar] [CrossRef]
- Wong, K.M.K.; Goh, T.K.; Hodgkiss, I.J.; Hyde, K.D.; Ranghoo, V.M.; Tsui, C.K.M.; Ho, W.H.; Wong, W.S.W.; Yuen, T.K. Role of fungi in freshwater ecosystems. Biodivers. Conserv. 1998, 7, 1187–1206. [Google Scholar] [CrossRef]
- Hyde, K.D.; Fryar, S.; Tian, Q.; Bahkali, A.H.; Xu, J.C. Lignicolous freshwater fungi along a north–south latitudinal gradient in the Asian/Australian region; can we predict the impact of global warming on biodiversity and function? Fungal Ecol. 2016, 19, 190–200. [Google Scholar] [CrossRef]
- Raja, H.A.; Shearer, C.A.; Tsui, C.K.M. Freshwater Fungi. eLS 2018, 1–13. [Google Scholar] [CrossRef]
- Goh, T.K.; Hyde, K.D. Biodiversity of freshwater fungi. J. Ind. Microbiol. 1996, 17, 328–334. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Hyde, K.D.; Pang, K.L. Freshwater Fungi and Fungal-Like Organisms; De Gruyter: Berlin, Germany, 2014. [Google Scholar]
- Da Silva, G.V.R.; Castañeda-Ruiz, R.F.; Malosso, E. Comparison of aquatic hyphomycetes communities between lotic and lentic environments in the Atlantic rain forest of Pernambuco, Northeast Brazil. Fungal Biol. 2019, 123, 660–668. [Google Scholar] [CrossRef]
- Ingold, C.T. Aquatic hyphomycetes of decaying alder leaves. Trans. Brit. Mycol. Soc. 1942, 25, 339–417. [Google Scholar] [CrossRef]
- Ingold, C.T. Aquatic ascomycetes: Ceriospora caudaesuis n. sp. and Ophiobolus tyhae. Trans. Brit. Mycol. Soc. 1951, 34, 210–215. [Google Scholar] [CrossRef]
- Ingold, C.T.; Chapman, B. Aquatic ascomycetes: Loramyces juncicola Weston and L. macrospora n. sp. Trans. Brit. Mycol. Soc. 1952, 35, 268–272. [Google Scholar] [CrossRef]
- Hyde, K.D.; Goh, T.K. Fungi on submerged wood in Lake Barrine, north Queensland, Australia. Mycol. Res. 1998, 102, 739–749. [Google Scholar] [CrossRef]
- Cai, L.; Tsui, C.; Zhang, K.; Hyde, K.D. Aquatic fungi from Lake Fuxian, Yunnan, China. Fungal Divers. 2002, 9, 57–70. [Google Scholar]
- Voglmayr, H.; Delgado-Rodriguez, G. Dendroclathra caeruleofusca gen. nov. et sp. nov., an aeroaquatic hyphomycete from Cuba. Can. J. Bot. 2001, 79, 995–1000. [Google Scholar] [CrossRef]
- Sridhar, K.R.; Kaveriappa, K.M. Colonization of leaves by water-borne hyphomycetes in a tropical stream. Mycol. Res. 1989, 92, 392–396. [Google Scholar] [CrossRef]
- Minoura, K.; Muroi, T. Some freshwater ascomycetes from Japan. Trans. Mycol. Soc. 1978, 19, 129–134. [Google Scholar]
- Nawawi, A. Some interesting hyphomycetes from water. Mycotaxon 1985, 24, 217–226. [Google Scholar]
- Kuthubutheen, A.J.; Nawawi, A. Dictyochaeta guadalcanalensis comb. nov. and several new records of the genus in Malaysia. Mycol. Res. 1991, 95, 1220–1223. [Google Scholar] [CrossRef]
- Sivichai, S.; Hywel-Jones, N. Biflagellospora (aero-aquatic hyphomycetes) from submerged wood in Thailand. Mycol. Res. 1999, 103, 908–914. [Google Scholar] [CrossRef]
- Sivichai, S.; Jones, E.B.G.; Hywel-Jones, N. Fungal colonisation of wood in a freshwater stream at Tad Ta Phu, Khao Yai National Park, Thailand. In: Fungal Succession (eds. KD Hyde, EB GarethJones). Fungal Divers. 2002, 10, 113–129. [Google Scholar]
- Shearer, C.A.; Raja, H.A. Freshwater Ascomycetes Database. Available online: http://fungi.life.illinois.Edu/ (accessed on 11 May 2013).
- Vijaykrishna, D.; Jeewon, R.; Hyde, K.D. Fusoidispora aquatica: New freshwater ascomycetes from Hong Kong based on morphology and molecules. Sydowia 2005, 57, 267–280. [Google Scholar]
- Hirayama, K.; Tanaka, K.; Raja, H.A.; Miller, A.N.; Shearer, C.A. A molecular phylogenetic assessment of Massarina ingoldiana sensu lato. Mycologia 2010, 102, 729–746. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, A.; Miller, A.N.; Shearer, C.A. Minutisphaera and Natipusilla: Two new genera of freshwater Dothideomycetes. Mycologia 2011, 103, 411–423. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, F.R.; Gusmao, L.F.P.; Raja, H.A.; Shearer, C.A. New species and new records of freshwater ascomycetes from Brazil and Costa Rica. Mycologia 2013, 105, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Raja, H.A.; Oberlies, N.H.; Figueroa, M.; Tanaka, K.; Hirayama, K.; Hashimoto, A.; Miller, A.N.; Zelski, S.E. Freshwater ascomycetes: Minutisphaera (Dothideomycetes) revisited, including one new species from Japan. Mycologia 2013, 105, 959–976. [Google Scholar] [CrossRef] [Green Version]
- Raja, H.A.; El-Elimat, T.; Oberlies, N.H.; Shearer, C.A.; Miller, A.N.; Tanaka, K.; Hashimoto, A.; Fournier, J. Minutisphaerales (Dothideomycetes, Ascomycota): A new order of freshwater ascomycetes including a new family, Minutisphaeraceae, and two new species from North Carolina, USA. Mycologia 2015, 107, 845–862. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Bhat, D.J.; McKenzie, E.H.C.; Bahkali, A.H.; Jones, E.B.G.; Liu, Z.Y. Aquapteridospora lignicola gen. et sp. nov., a new hyphomycetous taxon (Sordariomycetes) from wood submerged in a freshwater stream. Cryptogam. Mycol. 2015, 36, 469–479. [Google Scholar] [CrossRef]
- Yang, J.; Liu, J.K.; Hyde, K.D.; Bhat, D.J.; Jones, E.B.G.; Liu, Z.Y. New species of Sporoschisma (Chaetosphaeriaceae) from aquatic habitats in Thailand. Phytotaxa 2016, 289, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.F.; Bhat, D.J.; Lin, C.G.; Li, W.L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Raja, H.A.; Tanaka, K.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Doilom, M.; et al. Freshwater Dothideomycetes. Fungal Divers. 2020, 105, 319–575. [Google Scholar] [CrossRef]
- Chapela, I.H.; Petrini, O.; Hagann, L. Monolignol glucosides as specific recognition messengers in fungus-plant symbioses. Physiol. Mol. Plant. Pathol. 1991, 39, 289–298. [Google Scholar] [CrossRef]
- Feau, N.; Vialle, A.; Allaire, M.; Tanguay, P.; Joly, D.L.; Frey, P.; Callan, B.E.; Hamelin, R.C. Fungal pathogen (mis-) identifications: A case study with DNA barcodes on Melampsora rusts of aspen and white poplar. Mycol. Res. 2009, 113, 713–724. [Google Scholar] [CrossRef] [Green Version]
- Mortimer, P.E.; Jeewon, R.; Xu, J.K.; Lumyong, S.; Wanasinghe, D.N. Morpho-Phylo taxonomy of novel Dothideomycetous fungi Associated with dead woody twigs in Yunnan Province, China. Front. Microbiol. 2021, 12, 654683. [Google Scholar] [CrossRef]
- Guarro, J.; Gene, J.; Stchigel, A.M. Developments in Fungal Taxonomy. Clin. Microbiol. Rev. 1999, 12, 454–500. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Bao, D.F.; Hongsanan, S.; Chethana, K.W.T.; Yang, J.; Suwanarach, N. Evolution of freshwater Diaporthomycetidae (Sordariomycetes) provides evidence for five new orders and six new families. Fungal Divers. 2021, 107, 71–105. [Google Scholar] [CrossRef]
- Calabon, M.; Hyde, K.D.; Jones, E.B.G.; Chandrasiri, S.; Dong, W.; Fryar, S.C.; Yang, J.; Luo, Z.L.; Lu, Y.Z.; Bao, D.F. www. freshwaterfungi. org, an online platform for the taxonomic classification of freshwater fungi. Asian J. Mycol. 2020, 3, 419–445. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Rajeshkumar, K.C.; Hawksworth, D.L.; Madrid, H.; Kirk, P.M.; Braun, U.; Singh, R.V.; Crous, W.; Kukwa, M.; et al. Notes for genera: Ascomycota. Fungal Divers. 2017, 86, 1–594. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Y.; Udayanga, D.; Luo, Z.L.; Chen, L.J.; Zhou, D.Q.; Su, H.Y.; Hyde, K.D. Backbone tree for Chaetothyriales with four new species of Minimelanolocus from aquatic habitats. Fungal Biol. 2015, 119, 1046–1062. [Google Scholar] [CrossRef]
- Wang, G.N.; Yu, X.D.; Dong, W.; Bhat, D.J.; Boonmee, S.; Zhang, D.; Zhang, H. Freshwater hyphomycetes in Eurotiomycetes: A new species of Minimelanolocus and a new collection of Thysanorea papuana (Herpotrichiellaceae). Mycol. Prog. 2019, 18, 511–522. [Google Scholar] [CrossRef]
- Dong, W.; Hyde, K.D.; Doilom, M.; Yu, X.D.; Bhat, D.J.; Jeewon, R.; Boonmee, S.; Wang, G.N.; Nalumpang, S.; Zhang, H. Pseudobactrodesmium (Dactylosporaceae, Eurotiomycetes, Fungi) a novel lignicolous genus. Front. Microbiol. 2020, 11, 456. [Google Scholar] [CrossRef] [Green Version]
- Sivichai, S.; Jones, E.B.G.; Hywel-Jones, N.L. Lignicolous freshwater Ascomycota from Thailand: Hymenoscyphus varicosporoides and its Tricladium anamorph. Mycologia 2003, 95, 340–346. [Google Scholar] [CrossRef]
- Hyde, K.D.; McKenzie, E.H.C.; KoKo, T.W. Towards incorporating anamorphic fungi in a natural classification-checklist and notes for 2010. Mycosphere 2011, 2, 1–98. [Google Scholar]
- Sati, S.C.; Pathak, R. Anamorph (asexual stage) teleomorph (sexual stage) connections in aquatic hyphomycetes. Int. J. Plant Biol. 2016, 8, 128–135. [Google Scholar]
- Tanney, J.; Miller, A.N. Asexual-sexual morph connection in the type species of Berkleasmium. IMA Fungus 2017, 8, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.L.; Hyde, K.D.; Bhat, D.J.; Jeewon, R.; Maharachchikumbura, S.S.N.; Bao, D.F.; Li, W.L.; Su, X.J.; Yang, X.Y.; Su, H.Y. Morphological and molecular taxonomy of novel species Pleurotheciaceae from freshwater habitats in Yunnan, China. Mycol. Prog. 2018, 17, 511–530. [Google Scholar] [CrossRef]
- Tubaki, K.; Watanabe, K.; Manoch, L. Aquatic hyphomycetes from Thailand. Trans. Brit. Mycol. Soc. 1983, 24, 451–457. [Google Scholar]
- Sivichai, S.; Jones, N.H.; Jones, E.B.G. Lignicolous freshwater Ascomycota from Thailand: 1. Ascotaiwania sawada and its anamorph state Monotosporella. Mycoscience 1998, 39, 307–311. [Google Scholar] [CrossRef]
- Sivichai, S.; Jones, N.H.; Somrithipol, S. Lignicolous freshwater Ascomycota from Thailand: Melanochaeta and Sporoschisma anamorphs. Mycol. Res. 2000, 104, 478–485. [Google Scholar] [CrossRef] [Green Version]
- Sivichai, S.; Jones, E.B.G. Teleomorphic-anamorphic connections of freshwater fungi. In: Tsui CKM, Hyde KD, eds. Freshwater Mycology. Fungal Divers. 2003, 10, 259–272. [Google Scholar]
- Jones, E.B.G.; Wong, S.; Sivichai, S.; Au, D.T.W.; Hywel-Jones, N.L. Lignicolous freshwater Ascomycota from Thailand: Micropeltopsis quinquecladiopsis sp. nov. Mycol. Res. 1999, 103, 729–735. [Google Scholar] [CrossRef]
- Xu, L.; Bao, D.F.; Luo, Z.L.; Su, X.J.; Shen, H.W.; Su, H.Y. Lignicolous freshwater ascomycota from Thailand: Phylogenetic and morphological characterization of two new freshwater fungi: Tingoldiago hydei sp. nov. and T. clavata sp. nov. from eastern Thailand. MycoKeys 2020, 65, 119–138. [Google Scholar] [CrossRef] [Green Version]
- Sivichai, S.; Boonyene, N. Freshwater fungi. In Thai Fungal Diversity; Jones, E.B.G., Tanticharoen, M., Hyde, K.D., Eds.; BIOTEC: Pathum Thani, Thailand, 2004; pp. 95–106. [Google Scholar]
- Zhang, H.; Jones, E.B.G.; Zhou, D.; Bahkali, A.H.; Hyde, K.D. Checklist of freshwater fungi in Thailand. Cryptogam. Mycol. 2011, 32, 199–217. [Google Scholar] [CrossRef]
- He, S.C. A new record genus and species of aquatic cup fungi from China. Acta Mycol. Sinica. 1988, 7, 120–121. [Google Scholar]
- Tsui, K.M.; Goh, T.K.; Hyde, K.D. A new species of Dactylaria from Hong Kong. Sydowia 1997, 49, 182–186. [Google Scholar]
- Tsui, C.K.M.; Hyde, K.D.; Hodgkiss, I.J. Biodiversity of fungi on submerged wood in Hong Kong streams. Aquat. Microb. Ecol. 2000, 21, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Tsui, C.K.M.; Goh, T.K.; Hyde, K.D.; Hodgkiss, I.J. New records or species of Dictyochaeta, Endophragmiella and Ramichloridium from submerged wood in Hong Kong freshwater streams. Cryptogam. Mycol. 2001, 22, 139–145. [Google Scholar] [CrossRef]
- Tsui, C.K.M.; Goh, T.K.; Hyde, K.D.; Hodgkiss, I.J. New species or records of Cacumisporium, Helicosporium, Monotosporella and Bahusutrabeeja on submerged wood in Hong Kong streams. Mycologia 2001, 93, 389–397. [Google Scholar] [CrossRef]
- Tsui, C.K.M.; Hodgkiss, I.J.; Hyde, K.D. Three new species of Aquaticola (Ascomycetes) from tropical freshwater habitats. Nova Hedwigia 2003, 77, 161–168. [Google Scholar] [CrossRef]
- Goh, T.K.; Tsui, K.M.; Hyde, K.D. Elegantimyces sporidesmiopsis gen. et sp. nov. on submerged wood from Hong Kong. Mycol. Res. 1998, 102, 239–242. [Google Scholar] [CrossRef]
- Ho, W.H.; Hyde, K.D.; Hodgkiss, I.J. Digitodesmium recurvum, a new species of chirosporous hyphomycete from Hong Kong. Mycologia 1999, 91, 900–904. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Yin, L.; Cai, L.; Zhang, K.; Hyde, K.D. Freshwater fungi in Lake Dianchi, a heavily polluted lake in Yunnan, China. Fungal Divers. 2004, 16, 93–112. [Google Scholar]
- Hu, D.M.; Liu, F.; Cai, L. Biodiversity of aquatic fungi in China. Mycol. Int. J. Fungal Biol. 2013, 4, 125–168. [Google Scholar] [CrossRef]
- Li, W.L.; Luo, Z.L.; Liu, J.K.; Bhat, D.J.; Bao, D.F.; Su, H.Y.; Hyde, K.D. Lignicolous freshwater fungi from China I: Aquadictyospora lignicola gen. et sp. nov. and new record of Pseudodictyosporium wauense from northwestern Yunnan Province. Mycosphere 2017, 8, 1587–1597. [Google Scholar] [CrossRef]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Bhat, D.J.; Bao, D.F.; Li, W.L.; Su, H.Y. Lignicolous freshwater fungi from China II: Novel Distoseptispora (Distoseptisporaceae) species from northwestern Yunnan Province and a suggested unified method for studying lignicolous freshwater fungi. Mycosphere 2018, 9, 444–461. [Google Scholar] [CrossRef]
- Bao, D.F.; Luo, Z.L.; Liu, J.K.; Bhat, D.J.; Sarunya, N.; Li, W.L.; Su, H.Y.; Hyde, K.D. Lignicolous freshwater fungi in China III: New species and record of Kirschsteiniothelia from northwestern Yunnan Province. Mycosphere 2018, 9, 755–768. [Google Scholar] [CrossRef]
- Liu, N.G.; Hyde, K.D.; Bhat, D.J.; Jumpathong, J.; Liu, J.K. Morphological and phylogenetic studies of Pleopunctum gen. nov. (Phaeoseptaceae, Pleosporales) from China. Mycosphere 2019, 10, 757–775. [Google Scholar] [CrossRef]
- Bao, D.F.; Su, H.Y.; Maharachchikumbura, S.S.N.; Liu, J.K.; Nalumpang, S.; Luo, Z.L.; Hyde, K.D. Lignicolous freshwater fungi from China and Thailand: Multi-gene phylogeny reveals new species and new records in Lophiostomataceae. Mycosphere 2019, 10, 1080–1099. [Google Scholar] [CrossRef]
- Lu, Y.Z.; Zhang, J.Y.; Lin, C.G.; Luo, Z.L.; Liu, J.K. Pseudodactylaria fusiformis sp. nov. from freshwater habitat in China. Phytotaxa 2020, 446, 95–102. [Google Scholar] [CrossRef]
- Su, H.Y.; Udayanga, D.; Luo, Z.L.; Manamgoda, D.S.; Zhao, Y.C.; Yang, J.; Liu, X.Y.; McKenzie, E.H.C.; Zhou, D.Q.; Hyde, K.D. Hyphomycetes from aquatic habitats in Southern China: Species of Curvularia (Pleosporaceae) and Phragmocephala (Melannomataceae). Phytotaxa 2015, 226, 201–4216. [Google Scholar] [CrossRef]
- Su, H.Y.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Ariyawansa, H.A.; Luo, Z.L.; Promputtha, I.; Tian, Q.; Lin, C.G.; Shang, Q.J.; Zhao, Y.C.; et al. The families Distoseptisporaceae fam. nov., Kirschsteiniotheliaceae, Sporormiaceae and Torulaceae, with new species from freshwater in Yunnan Province, China. Fungal Divers. 2016, 80, 375–409. [Google Scholar] [CrossRef]
- Song, H.Y.; Zhong, P.A.; Liao, J.L.; Wang, Z.H.; Hu, D.M.; Huang, Y.J. Junewangia aquatica (Junewangiaceae), a new species from freshwater habitats in China. Phytotaxa 2018, 336, 272–278. [Google Scholar] [CrossRef]
- Song, H.Y.; Huo, G.H.; Hu, D.M. Dictyosporella hydei sp. nov., an asexual species from freshwater habitats in China. Phytotaxa 2018, 358, 181–188. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, D.J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The faces of fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. Res. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, G.M., Shinsky, D., White, T., Eds.; Academic: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Glez-Peña, D.; Gómez-Blanco, D.; Reboiro-Jato, M.; Fdez-Riverola, F. ALTER: Programoriented conversion of DNA and protein alignments. Nucleic Acids Res. 2010, 38, 14–18. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web-servers. Syst. Biol. 2008, 75, 758–771. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2 Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef]
- Zhaxybayeva, O.; Gogarten, J.P. Bootstrap, Bayesian probability and maximum likelihood mapping: Exploring new tools for comparative genome analyses. BMC Genom. 2002, 3, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree v1.4: Tree Figure Drawing Tool. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 12 August 2021).
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.M.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Hatakeyama, S.; Harada, Y.; Sano, T.; Shirouzu, T.; Hosoya, T. Molecular taxonomy of bambusicolous fungi: Tetraplosphaeriaceae, a new pleosporalean family with Tetraploa-like anamorphs. Stud. Mycol. 2009, 64, 175–209. [Google Scholar] [CrossRef] [Green Version]
- Li, W.L.; Bao, D.F.; Bhat, D.J.; Su, H.Y. Tetraploa aquatica (Tetraplosphaeriaceae), a new freshwater fungal species from Yunnan Province, China. Phytotaxa 2020, 459, 181–189. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Hernandez-Restrepo, M.J.; Gené, R.; Castañeda-Ruiz, J.; Mena-Portales, J.; Crous, P.W.; Guarro, J. Phylogeny of saprobic microfungi from Southern Europe. Stud. Mycol. 2017, 86, 53–97. [Google Scholar] [CrossRef]
- Sinclair, R.C.; Boshoff, S.; Eicker, A. A new species of Monodictys from South Africa. Mycotaxon 1996, 59, 359–366. [Google Scholar]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Ellis, M.B. Clasterosporium and some allied Dematiaceae-Phragmosporae. I. Mycol. Pap. 1958, 70, 1–89. [Google Scholar]
- Ellis, M.B. More Dematiaceous Hyphomycetes; Commonwealth Mycological Institute; Kew Publisher: London, UK, 1976; p. 507. [Google Scholar]
- Ellis, M.B. Dematiaceous Hyphomycetes; Kew; Commonwealth Mycological Institute; Kew Publisher: London, UK, 1971; p. 608. [Google Scholar]
- Wu, W.P.; Zhuang, W.Y. Sporidesmium, Endophragmiella and related genera from China. Fungal Divers. 2005, 15, 1–531. [Google Scholar]
- Yang, J.; Maharachchikumbura, S.S.N.; Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Al-Sadi, A.M.; Liu, Z.Y. Pseudostanjehughesia aquitropica gen. et sp. nov. and Sporidesmium sensu lato species from freshwater habitats. Mycol. Prog. 2018, 17, 591–616. [Google Scholar] [CrossRef]
- Liu, L.L.; Yang, J.; Liu, N.G.; Chen, Y.Y.; Gui, X.X.; Liu, Z.Y. Sporidesmium guizhouense sp. nov. (Sordariomycetes incertae sedis), a new species from a freshwater habitat in Guizhou Province, China. Phytotaxa 2019, 422, 144–156. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Khemmuk, W.; Geering, A.D.; Shivas, R.G. Wongia gen. nov.(Papulosaceae, Sordariomycetes), a new generic name for two root-infecting fungi from Australia. IMA Fungus 2016, 7, 247–252. [Google Scholar] [CrossRef]
- Cai, L.; Zhang, K.; McKenzie, E.H.C.; Ho, W.H.; Hyde, K.D. Acrodictys liputii sp. nov. and Digitodesmium bambusicola sp. nov. from bamboo submerged in Liput River in the Philippines. Nova Hedwigia. 2002, 75, 525–532. [Google Scholar] [CrossRef]
- Xia, J.W.; Ma, Y.R.; Li, Z.; Zhang, X.G. Acrodictys-like wood decay fungi from southern China, with two new families Acrodictyaceae and Junewangiaceae. Sci. Rep. 2017, 7, 1–21. [Google Scholar] [CrossRef]
- Fernández, F.A.; Huhndorf, S.M. New species of Chaetosphaeria, Melanopsammella and Tainosphaeria gen. nov. from the Americas. Fungal Divers. 2005, 18, 15–57. [Google Scholar]
- Crous, P.W.; Verkley, G.J.M.; Christensen, M.; Castaneda-Ruiz, R.F.; Groenewald, J.Z. How important are conidial appendages? Persoonia 2012, 28, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Réblová, M.; Miller, A.N.; Rossman, A.Y.; Seifert, K.A.; Crous, P.W.; Hawksworth, D.L.; Abdel-Wahab, M.A.; Cannon, P.F.; Daranagama, D.A.; De Beer, Z.W.; et al. Recommendations for competing sexual-asexually typified generic names in Sordariomycetes (except Diaporthales, Hypocreales, and Magnaporthales). IMA Fungus 2016, 7, 131–153. [Google Scholar] [CrossRef]
- Lin, C.G.; McKenzie, E.H.C.; Liu, J.K.; Jones, E.B.G.; Hyde, K.D. Hyaline-spored chaetosphaeriaceous hyphomycetes from Thailand and China, with a review of the family Chaetosphaeriaceae. Mycosphere 2019, 10, 655–700. [Google Scholar] [CrossRef]
- Yuan, H.S.; Lu, X.; Dai, Y.C.; Hyde, K.D.; Kan, Y.H.; Kušan, Y.; He, S.H.; Liu, N.G.; Sarma, V.V.; Zhao, C.L.; et al. Fungal diversity notes 1277–1386: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2020, 104, 1–266. [Google Scholar] [CrossRef]
- Wei, M.J.; Zhang, H.; Dong, W.; Boonmee, S.; Zhang, D. Introducing Dictyochaeta aquatica sp. nov. and two new species of Chloridium (Chaetosphaeriaceae, Sordariomycetes) from aquatic habitats. Phytotaxa 2018, 362, 187–199. [Google Scholar] [CrossRef]
- Lin, C.G.; McKenzie, E.H.C.; Bhat, D.J.; Liu, J.K.; Hyde, K.D.; Lumyong, S. Pseudodactylaria brevis sp. nov. from Thailand confirms the status of Pseudodactylariaceae. Phytotaxa 2018, 369, 241–250. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.J.; Hardy, G.; Smith, D. Fungal Planet description sheets: 625–715. Persoonia 2017, 39, 270–467. [Google Scholar] [CrossRef]
- Dai, D.Q.; Bahkali, A.H.; Li, Q.R.; Bhat, D.J.; Wijayawardene, N.N.; Li, W.J.; Chukeatirote, E.; Zhao, R.L.; Xu, J.C.; Hyde, K.D. Vamsapriya (Xylariaceae) re-described, with two new species and molecular sequence data. Cryptogam. Mycol. 2014, 35, 339–357. [Google Scholar] [CrossRef]
- Jiang, H.B.; Phookamsak, R.; Bhat, D.J.; Khan, S.; Bahkali, A.H.; Elgorban, A.M.; Hyde, K.D. Vamsapriya yunnana, a new species of Vamsapriya (Xylariaceae, Xylariales) associated with bamboo from Yunnan, China. Phytotaxa 2018, 356, 61–70. [Google Scholar] [CrossRef]
- Gawas, P.; Bhat, D.J. Vamsapriya indica gen. et sp. nov., a bambusicolous, synnematous fungus from India. Mycotaxon 2005, 94, 149–154. [Google Scholar]
- Ling, Y.; Li, H.H.; Xia, J.W.; Zhang, X.G.; Zhuang, L. Vamsapriya jinniuensis sp. nov., and a first record of Garnaudia elegans from southern China. Mycotaxon 2018, 133, 367–372. [Google Scholar] [CrossRef]
- Castañeda-Ruiz, R.F.; Zhang, X.G.; Li, D.W.; Gusmão, L.F.P.; Pérez-Martínez, S.; Sosa, D. Notes on Vamsapriya and V. camagueyensis comb. nov. Mycotaxon 2017, 132, 553–557. [Google Scholar] [CrossRef]
- Jones, E.B.G. Form and function of fungal spore appendages. Mycoscience 2006, 47, 167–183. [Google Scholar] [CrossRef]
- Chan, S.; Goh, T.; Hyde, K.D. Ingoldian fungi in Hong Kong. Fungal Divers. 2000, 5, 89–107. [Google Scholar]
- Sridhar, K. Freshwater hyphomycetes. Available online: https://www.researchgate.net/publication/267452702_Freshwater_hyphomycetes (accessed on 12 August 2021).
- Read, S.J.; Moss, S.T.; Jones, E.B.G. Attachment and germination of conidia. In The Ecology of Aquatic Hyphomycetes; Baerlocher, F., Ed.; Springer: Berlin, Germany, 1992. [Google Scholar]
- Baerlocher, F.; Kendrick, B. Dynamics of the fungal population on leaves in a stream. J. Ecol. 1974, 62, 761–791. [Google Scholar] [CrossRef]
- Godfrey, B.E.S. Growth of two terrestrial microfungi on submerged alder leaves. Trans. Brit. Mycol. Soc. 1983, 79, 418–421. [Google Scholar] [CrossRef]
- Graça, M.A.S.; Ferreira, C.F. The ability of selected aquatic hyphomycetes and terrestrial fungi to decompose leaves in freshwater. Sydowia 1995, 47, 167–179. [Google Scholar]
- Dubey, M.K.; Upadhyay, R.S.; Kamil, D.; Gupta, R.C. Sporidesmium binserum sp. nov. from Binser forest of Almora, Himalaya, India. Indian Phytopathol. 2018, 71, 457–462. [Google Scholar] [CrossRef]
- Subramanian, C. A reassessment of Sporidesmium (Hyphomycetes) and some related taxa. Proc. Indian Natn. Sci. Acad. 1992, 58, 179–190. [Google Scholar]
- Hernandez-Gutierrez, A.; Sutton, B.C. Imimyces and Linkosia, two new genera segregated from Sporidesmium sensu lato, and redescription of Polydesmus. Mycol. Res. 1997, 101, 201–209. [Google Scholar] [CrossRef]
- Shenoy, B.D.; Jeewon, R.; Wu, W.P.; Bhat, D.J.; Hyde, K.D. Ribosomal and RPB2 DNA sequence analyses suggest that Sporidesmium and morphologically similar genera are polyphyletic. Mycol. Res. 2006, 110, 916–928. [Google Scholar] [CrossRef]
- Sri-indrasutdhi, V.; Boonyuen, N.; Suetrong, S.; Chuaseeharonnachai, C.; Sivichai, S.; Jones, E.B.G. Wood-inhabiting freshwater fungi from Thailand: Ascothailandia grenadoidia gen. et sp. nov., Canalisporium grenadoidia sp. nov. with a key to Canalisporium species (Sordariomycetes, Ascomycota). Mycoscience 2010, 51, 411–420. [Google Scholar] [CrossRef]
- Calabon, M.S.; Jones, E.B.G.; Boonmee, S.; Doilom, M.; Lumyong, S.; Hyde, K.D. Five Novel freshwater ascomycetes indicate high undiscovered diversity in lotic habitats in Thailand. J. Fungi. 2021, 7, 117. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jeewon, R.; Chen, Y.J.; Bhunjun, C.S.; Calabon, M.S.; Jiang, H.B.; Lin, C.G.; Norphanphoun, C.; Sysouphanthong, P.; Pem, D. The numbers of fungi: Is the descriptive curve flattening? Fungal Divers. 2020, 103, 219–271. [Google Scholar] [CrossRef]
- Nagy, L.G.; Petkovits, T.; Kovács, G.M.; Voigt, K.; Vágvölgyi, C.; Papp, T. Where is the unseen fungal diversity hidden? A study of Mortierella reveals a large contribution of reference collections to the identification of fungal environmental sequences. New Phytol. 2011, 191, 789–794. [Google Scholar] [CrossRef]
- Sota, T.; Kagata, H.; Ando, Y.; Utsumi, S.; Osono, T. Metagenomic Approach Yields Insights into Fungal Diversity and Functioning. Species Divers. Community Struct. 2013, 1–23. [Google Scholar] [CrossRef]
- Fouquier, J.; Schwartz, T.; Kelley, S.T. Rapid assemblage of diverse environmental fungal communities on public restroom floors. Indoor. Air. 2016, 26, 869–879. [Google Scholar] [CrossRef]
- Aline, B.M.V.; Paula, L.C.F.; Laura, R.L.; Fernanda, B.; Anna, C.M.S.; Flavio, M.G.A.; Sara, C.O.; Ângelo, A.D.; Carlos, A.R.; Guilherme, O.; et al. Using next-generation sequencing (NGS) to uncover diversity of wood-decaying fungi in neotropical atlantic forests. Phytotaxa 2017, 295, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Purahong, W.; Wubet, T.; Hyde, K.D.; Zhang, W.; Xu, H.; Zhang, G.J.; Fu, C.H.; Liu, M.; Xin, Q.K.; et al. Direct comparison of culture-dependent and culture-independent molecular approaches reveal the diversity of fungal endophytic communities in stems of grapevine (Vitis vinifera). Fungal Divers. 2018, 90, 85–107. [Google Scholar] [CrossRef]





























Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, D.-F.; Hyde, K.D.; McKenzie, E.H.C.; Jeewon, R.; Su, H.-Y.; Nalumpang, S.; Luo, Z.-L. Biodiversity of Lignicolous Freshwater Hyphomycetes from China and Thailand and Description of Sixteen Species. J. Fungi 2021, 7, 669. https://doi.org/10.3390/jof7080669
Bao D-F, Hyde KD, McKenzie EHC, Jeewon R, Su H-Y, Nalumpang S, Luo Z-L. Biodiversity of Lignicolous Freshwater Hyphomycetes from China and Thailand and Description of Sixteen Species. Journal of Fungi. 2021; 7(8):669. https://doi.org/10.3390/jof7080669
Chicago/Turabian StyleBao, Dan-Feng, Kevin D. Hyde, Eric H. C. McKenzie, Rajesh Jeewon, Hong-Yan Su, Sarunya Nalumpang, and Zong-Long Luo. 2021. "Biodiversity of Lignicolous Freshwater Hyphomycetes from China and Thailand and Description of Sixteen Species" Journal of Fungi 7, no. 8: 669. https://doi.org/10.3390/jof7080669