
Virulence Factors and in-Host Selection on Phenotypes in Infectious Probiotic Yeast Isolates (Saccharomyces ‘boulardii’)

 ,
,  ,
,  ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolates and Patient Data
2.2. Genotyping
2.3. Phenotyping
2.4. Immune Tests
2.5. Human Epithelium Model Interactions
2.6. Galleria Mellonella Larva Infection Model
2.7. Statistics and Data Visualization
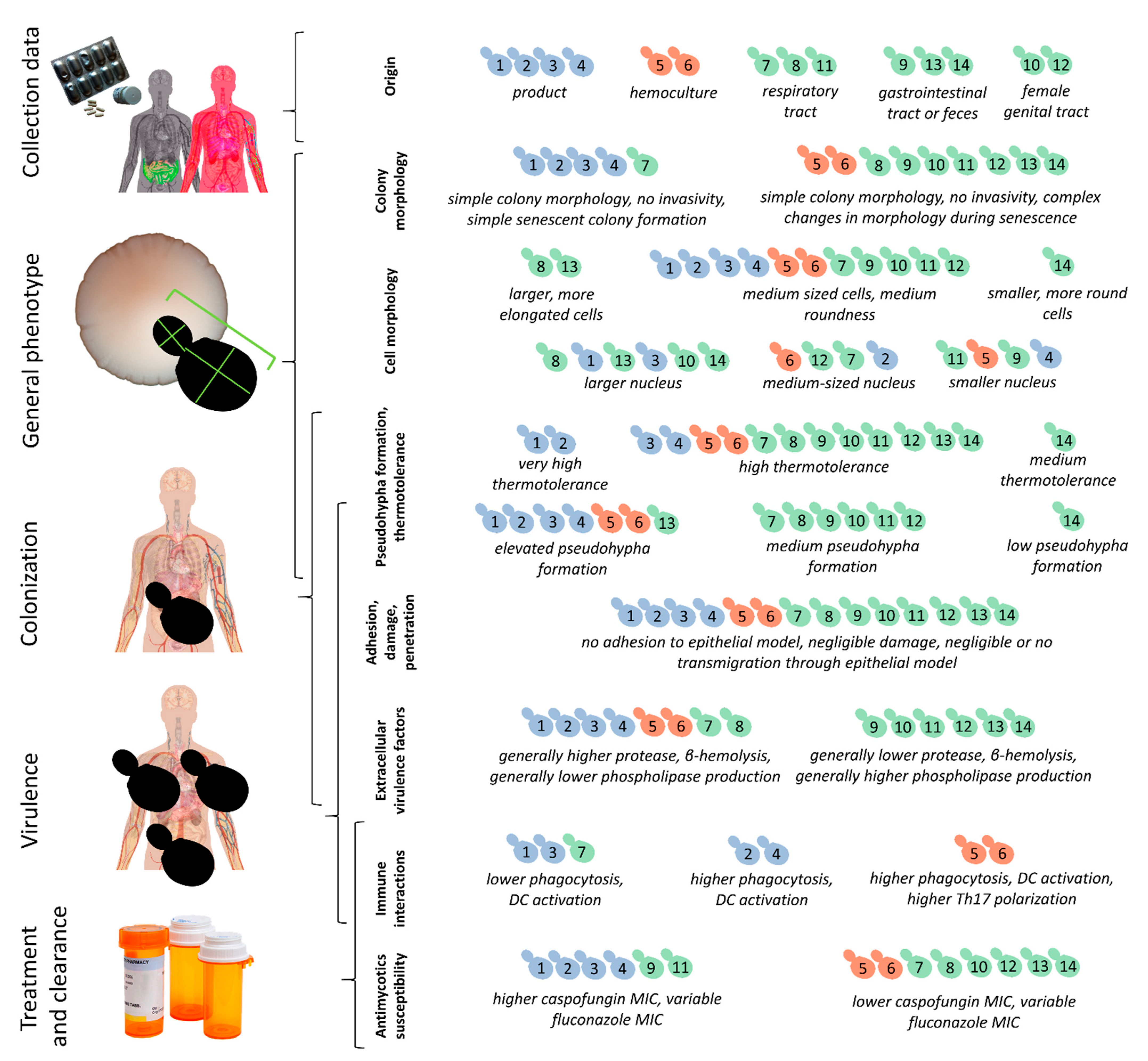
3. Results
3.1. S. ‘boulardii’ Isolates
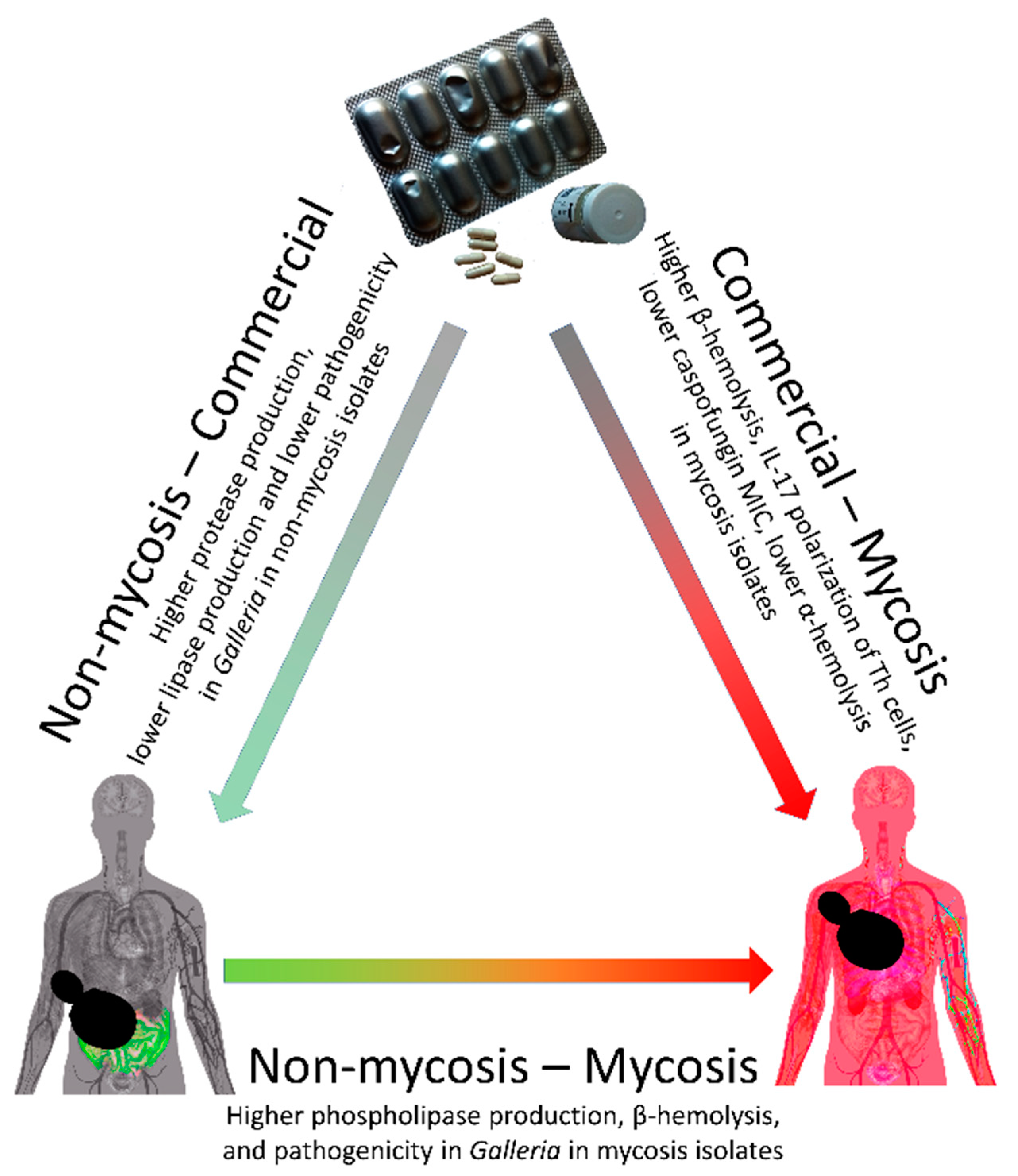
3.2. Phenotypic Differences among Commercial, Non-Mycosis, and Mycosis Isolates of S. ‘boulardii’
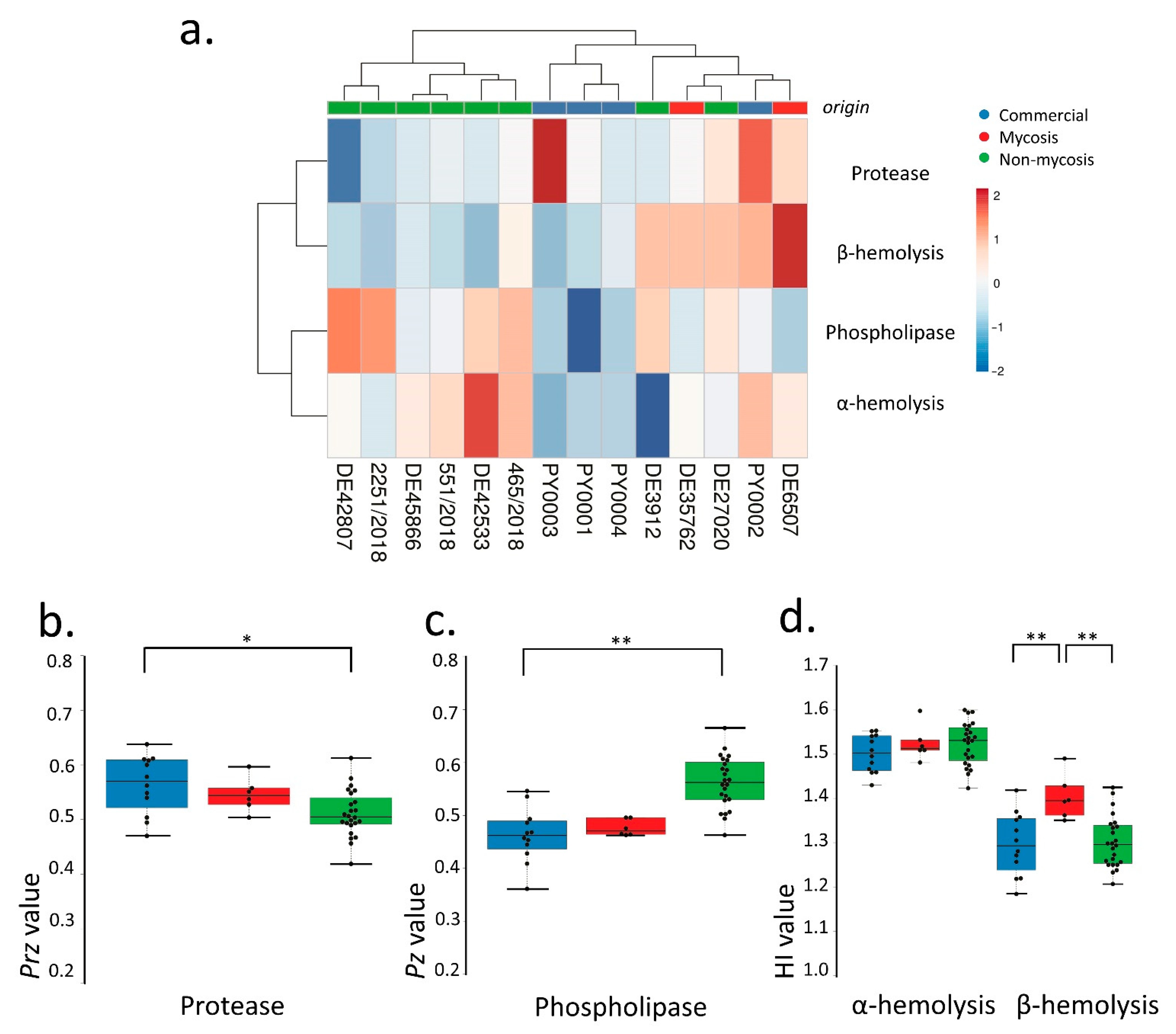
3.3. Virulence Factors and Pathogenicity of Commercial, Non-Mycosis, and Mycosis Isolates of S. ‘boulardii’
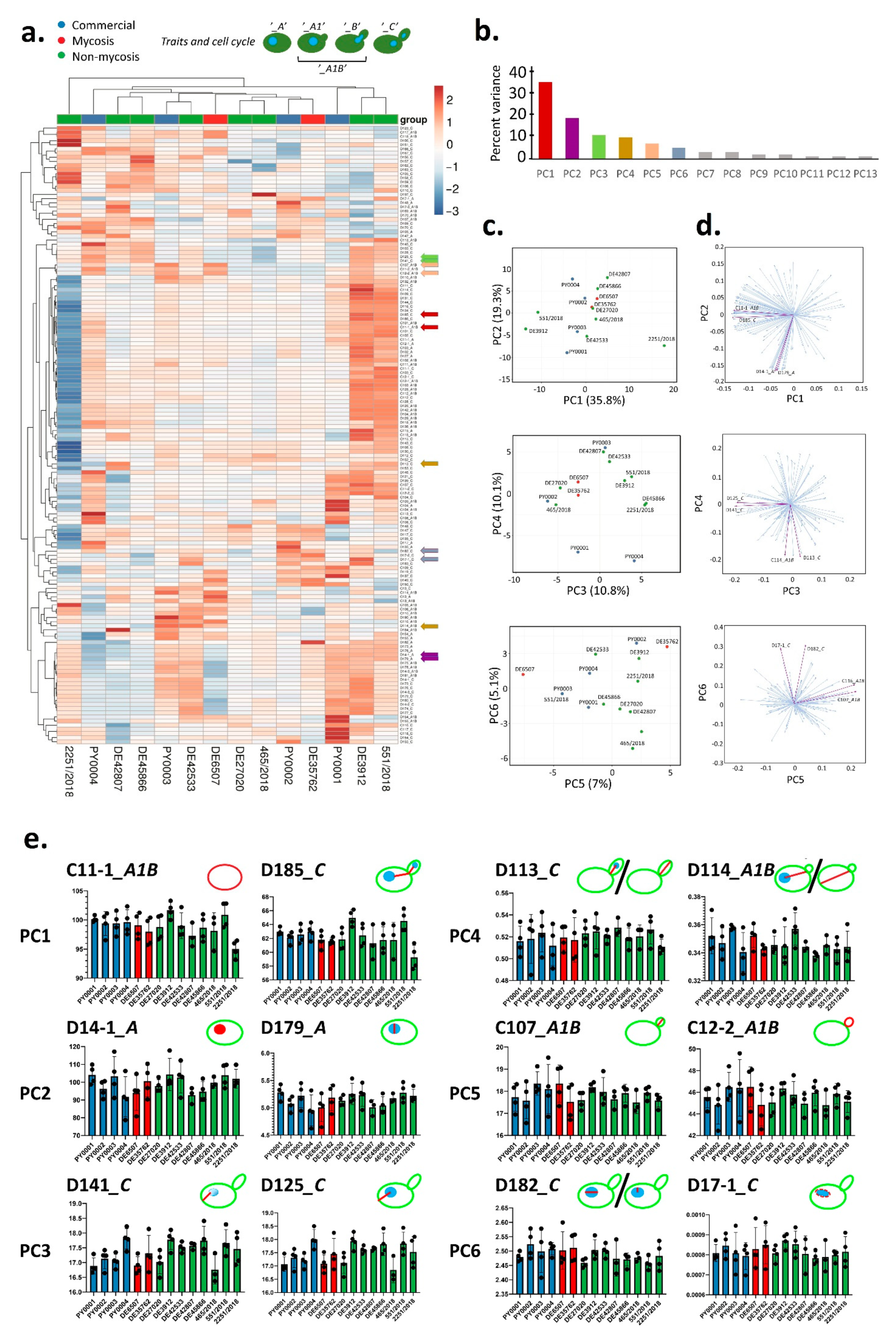
3.4. Cell Morphology and Pseudohypha Formation, Adhesion
3.5. Antifungal Susceptibility
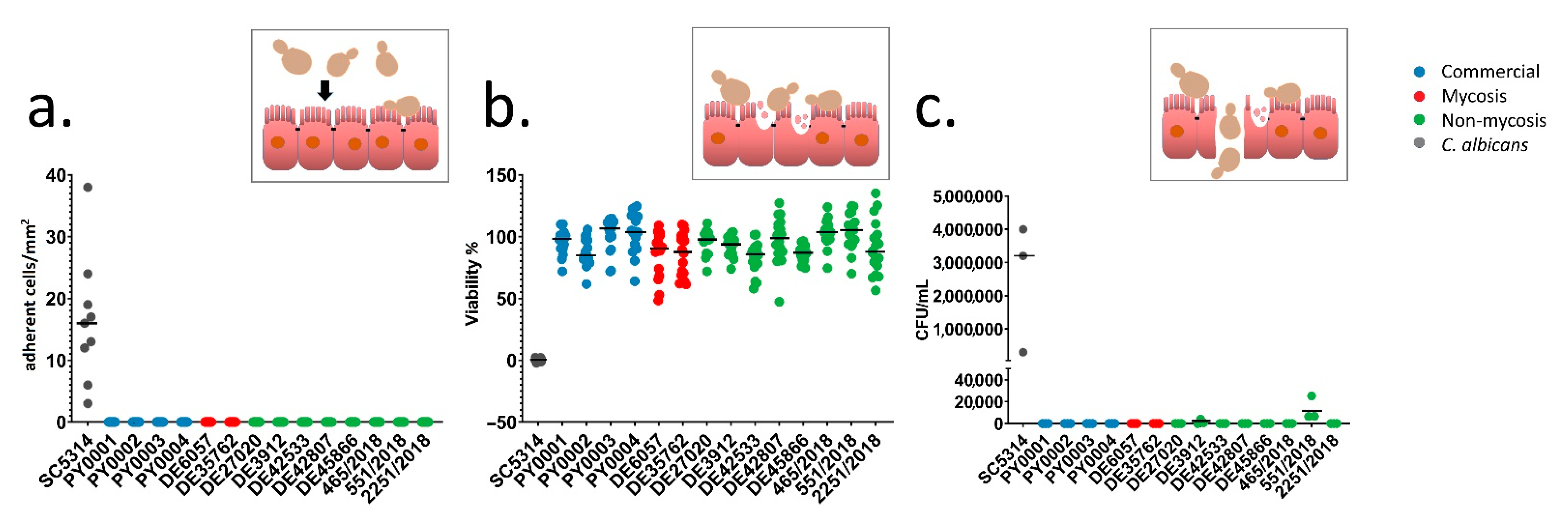
3.6. Epithelial Interactions (Adhesion, Damage, Transmigration)
3.7. Immunological Interactions
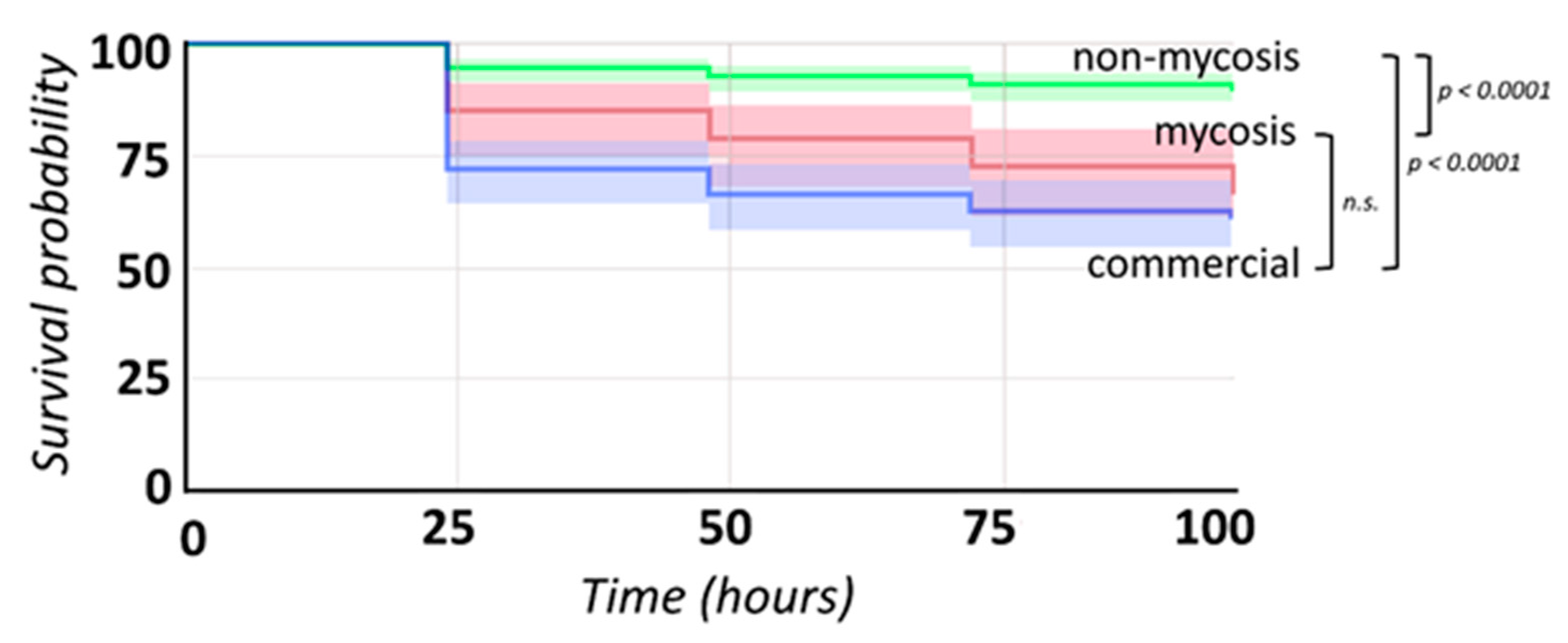
3.8. Galleria Infections
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parapouli, M.; Vasileiadis, A.; Afendra, A.S.; Hatziloukas, E. Saccharomyces cerevisiae and its industrial applications. AIMS Microbiol. 2020, 6, 1–31. [Google Scholar] [CrossRef]
- Nandy, S.K.; Srivastava, R.K. A review on sustainable yeast biotechnological processes and applications. Microbiol. Res. 2018, 207, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Labussière, E.; Achard, C.S.; Dubois, S.; Combes, S.; Castex, M.; Renaudeau, D. Saccharomyces cerevisiae boulardii CNCM I-1079 supplementation in finishing male pigs helps to cope with heat stress through feeding behavior and gut microbiota modulation. Br. J. Nutr. 2021, 1–35. [Google Scholar] [CrossRef]
- Santos, F.D.S.; Maubrigades, L.R.; Gonçalves, V.S.; Alves Ferreira, M.R.; Brasil, C.L.; Cunha, R.C.; Conceição, F.R.; Leite, F.P.L. Immunomodulatory effect of short-term supplementation with Bacillus toyonensis BCT-7112T and Saccharomyces boulardii CNCM I-745 in sheep vaccinated with Clostridium chauvoei. Vet. Immunol. Immunopathol. 2021, 237, 110272. [Google Scholar] [CrossRef]
- Biswas, A.; Dev, K.; Tyagi, P.K.; Mandal, A. The effect of multi-strain probiotics as feed additives on performance, immunity, expression of nutrient transporter genes and gut morphometry in broiler chickens. Anim. Biosci. 2021. online ahead of print. [Google Scholar] [CrossRef]
- McFarland, L.V. Common Organisms and Probiotics: Saccharomyces boulardii. In The Microbiota in Gastrointestinal Pathophysiology: Implications for Human Health, Prebiotics, Probiotics, and Dysbiosis; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 145–164. ISBN 9780128040621. [Google Scholar]
- Pais, P.; Almeida, V.; Yılmaz, M.; Teixeira, M.C. Saccharomyces boulardii: What makes it tick as successful probiotic? J. Fungi 2020, 6, 78. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of action of probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [Green Version]
- Constante, M.; De Palma, G.; Lu, J.; Jury, J.; Rondeau, L.; Caminero, A.; Collins, S.M.; Verdu, E.F.; Bercik, P. Saccharomyces boulardii CNCM I-745 modulates the microbiota–gut–brain axis in a humanized mouse model of Irritable Bowel Syndrome. Neurogastroenterol. Motil. 2020, 33, e13985. [Google Scholar] [CrossRef]
- Leventogiannis, K.; Gkolfakis, P.; Spithakis, G.; Tsatali, A.; Pistiki, A.; Sioulas, A.; Giamarellos-Bourboulis, E.J.; Triantafyllou, K. Effect of a preparation of four probiotics on symptoms of patients with Irritable Bowel Syndrome: Association with intestinal bacterial overgrowth. Probiotics Antimicrob. Proteins 2019, 11, 627–634. [Google Scholar] [CrossRef] [Green Version]
- Khatri, I.; Tomar, R.; Ganesan, K.; Prasad, G.S.; Subramanian, S. Complete genome sequence and comparative genomics of the probiotic yeast Saccharomyces boulardii. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Peter, J.; De Chiara, M.; Friedrich, A.; Yue, J.-X.; Pflieger, D.; Bergstrom, A.; Sigwalt, A.; Barré, B.; Freel, K.; Llored, A.; et al. Genome evolution across 1011 Saccharomyces cerevisiae isolates. Nature 2018, 556, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Costa, R.L.; Moreira, J.; Lorenzo, A.; Lamas, C.C. Infectious complications following probiotic ingestion: A potentially underestimated problem? A systematic review of reports and case series. BMC Complement. Altern. Med. 2018, 18, 329. [Google Scholar] [CrossRef] [Green Version]
- Ventoulis, I.; Sarmourli, T.; Amoiridou, P.; Mantzana, P.; Exindari, M.; Gioula, G.; Vyzantiadis, T.-A. Bloodstream infection by Saccharomyces cerevisiae in two COVID-19 patients after receiving supplementation of Saccharomyces in the ICU. J. Fungi 2020, 6, 98. [Google Scholar] [CrossRef]
- Chakravarty, S.; Parashar, A.; Acharyya, S. Saccharomyces cerevisiae sepsis following probiotic therapy in an infant. Indian Pediatrics 2019, 56, 971–972. [Google Scholar] [CrossRef] [PubMed]
- Imre, A.; Rácz, H.V.; Antunovics, Z.; Rádai, Z.; Kovács, R.; Lopandic, K.; Pócsi, I.; Pfliegler, W.P. A new, rapid multiplex PCR method identifies frequent probiotic origin among clinical Saccharomyces isolates. Microbiol. Res. 2019, 227, 126298. [Google Scholar] [CrossRef]
- Poncelet, A.; Ruelle, L.; Konopnicki, D.; Miendje Deyi, V.Y.; Dauby, N. Saccharomyces cerevisiae fungemia: Risk factors, outcome and links with S. boulardii-containing probiotic administration. Infect. Dis. Now 2021, 51, 293–295. [Google Scholar] [CrossRef]
- Edwards-Ingram, L.C.; Gent, M.E.; Hoyle, D.C.; Hayes, A.; Stateva, L.I.; Oliver, S.G. Comparative genomic hybridization provides new insights into the molecular taxonomy of the Saccharomyces sensu stricto complex. Genome Res. 2004, 14, 1043–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennequin, C.; Thierry, A.; Richard, G.F.; Lecointre, G.; Nguyen, H.V.; Gaillardin, C.; Dujon, B. Microsatellite typing as a new tool for identification of Saccharomyces cerevisiae strains. J. Clin. Microbiol. 2001, 39, 551–559. [Google Scholar] [CrossRef] [Green Version]
- McFarland, L.V. Systematic review and meta-analysis of Saccharomyces boulardii in adult patients. World J. Gastroenterol. 2010, 16, 2202–2222. [Google Scholar] [CrossRef] [PubMed]
- Anoop, V.; Rotaru, S.; Shwed, P.S.; Tayabali, A.F.; Arvanitakis, G. Review of current methods for characterizing virulence and pathogenicity potential of industrial Saccharomyces cerevisiae strains towards humans. FEMS Yeast Res. 2015, 15, fov057. [Google Scholar] [CrossRef] [Green Version]
- Clemons, K.V.; McCusker, J.H.; Davis, R.W.; Stevens, D.A. Comparative pathogenesis of clinical and nonclinical isolates of Saccharomyces cerevisiae. J. Infect. Dis. 1994, 169, 859–867. [Google Scholar] [CrossRef]
- Klingberg, T.D.; Lesnik, U.; Arneborg, N.; Raspor, P.; Jespersen, L. Comparison of Saccharomyces cerevisiae strains of clinical and nonclinical origin by molecular typing and determination of putative virulence traits. FEMS Yeast Res. 2008, 8, 631–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCusker, J.H.; Clemons, K.V.; Stevens, D.A.; Davis, R.W. Genetic characterization of pathogenic Saccharomyces cerevisiae isolates. Genetics 1994, 136, 1261–1269. [Google Scholar] [CrossRef]
- de Llanos, R.; Fernández-Espinar, M.T.; Querol, A. A comparison of clinical and food Saccharomyces cerevisiae isolates on the basis of potential virulence factors. Antonie van Leeuwenhoek 2006, 90, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Zupan, J.; Raspor, P. Quantitative agar-invasion assay. J. Microbiol. Methods 2008, 73, 100–104. [Google Scholar] [CrossRef]
- Pérez-Torrado, R.; Llopis, S.; Jespersen, L.; Fernández-Espinar, T.; Querol, A. Clinical Saccharomyces cerevisiae isolates cannot cross the epithelial barrier in vitro. Int. J. Food Microbiol. 2012, 157, 59–64. [Google Scholar] [CrossRef]
- Yáñez, A.; Murciano, C.; Llopis, S.; Fernández-espinar, T.; Gil, M.L.; Gozalbo, D. In vivo and in vitro studies on virulence and host responses to Saccharomyces cerevisiae clinical and non-clinical isolates. Open Mycol. J. 2009, 3, 37–47. [Google Scholar] [CrossRef]
- van der Aa Kühle, A.; Skovgaard, K.; Jespersen, L. In vitro screening of probiotic properties of Saccharomyces cerevisiae var. boulardii and food-borne Saccharomyces cerevisiae strains. Int. J. Food Microbiol. 2005, 101, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Llopis, S.; Querol, A.; Heyken, A.; Hube, B.; Jespersen, L.; Fernández-Espinar, M.; Pérez-Torrado, R. Transcriptomics in human blood incubation reveals the importance of oxidative stress response in Saccharomyces cerevisiae clinical strains. BMC Genom. 2012, 13, 419. [Google Scholar] [CrossRef] [Green Version]
- McCullough, M.J.; Clemons, K.V.; Mccusker, J.H.; Stevens, D.A. Species identification and virulence attributes of Saccharomyces boulardii (nom. inval.). J. Clin. Microbiol. 1998, 36, 2613–2617. [Google Scholar] [CrossRef] [Green Version]
- de Llanos, R.; Llopis, S.; Molero, G.; Querol, A.; Gil, C.; Fernández-Espinar, M.T. In vivo virulence of commercial Saccharomyces cerevisiae strains with pathogenicity-associated phenotypical traits. Int. J. Food Microbiol. 2011, 144, 393–399. [Google Scholar] [CrossRef]
- Llopis, S.; Hernández-Haro, C.; Monteoliva, L.; Querol, A.; Molina, M.; Fernández-Espinar, M.T. Pathogenic potential of Saccharomyces strains isolated from dietary supplements. PLoS ONE 2014, 9, e98094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfliegler, W.P.; Boros, E.; Pázmándi, K.; Jakab, Á.; Zsuga, I.; Kovács, R.; Urbán, E.; Antunovics, Z.; Bácsi, A.; Sipiczki, M.; et al. Commercial strain-derived clinical Saccharomyces cerevisiae can evolve new phenotypes without higher pathogenicity. Mol. Nutr. Food Res. 2017, 61, 1601099. [Google Scholar] [CrossRef] [Green Version]
- Rácz, H.V.; Mukhtar, F.; Imre, A.; Rádai, Z.; Gombert, A.K.; Rátonyi, T.; Nagy, J.; Pócsi, I.; Pfliegler, W.P. How to characterize a strain? Clonal heterogeneity in industrial Saccharomyces influences both phenotypes and heterogeneity in phenotypes. Yeast 2021, 38, 453–470. [Google Scholar] [CrossRef] [PubMed]
- Hanna, M.; Xiao, W. Isolation of nucleic acids. Methods Mol. Biol. 2006, 313, 15–20. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Csoma, H.; Zakany, N.; Capece, A.; Romano, P.; Sipiczki, M. Biological diversity of Saccharomyces yeasts of spontaneously fermenting wines in four wine regions: Comparative genotypic and phenotypic analysis. Int. J. Food Microbiol. 2010, 140, 239–248. [Google Scholar] [CrossRef]
- Huxley, C.; Green, E.D.; Dunham, I. Rapid assessment of S. cerevisiae mating type by PCR. Trends Genet. 1990, 6, 236. [Google Scholar] [CrossRef]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 4th ed.; CLSI standard M38; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Luo, G.; Samaranayake, L.P.; Yau, J.Y.Y. Candida species exhibit differential in vitro hemolytic activities. J. Clin. Microbiol. 2001, 39, 2971–2974. [Google Scholar] [CrossRef] [Green Version]
- Shuster, A.; Osherov, N.; Rosenberg, M. Alcohol-mediated haemolysis in yeast. Yeast 2004, 21, 1335–1342. [Google Scholar] [CrossRef] [Green Version]
- Ohya, Y.; Sese, J.; Yukawa, M.; Sano, F.; Nakatani, Y.; Saito, T.L.; Saka, A.; Fukuda, T.; Ishihara, S.; Oka, S.; et al. High-dimensional and large-scale phenotyping of yeast mutants. Proc. Natl. Acad. Sci. USA 2005, 102, 19015–19020. [Google Scholar] [CrossRef] [Green Version]
- Okada, H.; Ohnuki, S.; Ohya, Y. Quantification of cell, actin, and nuclear DNA morphology with high-throughput microscopy and CalMorph. Cold Spring Harb. Protoc. 2015. [Google Scholar] [CrossRef] [PubMed]
- Bauer, C.R.; Li, S.; Siegal, M.L. Essential gene disruptions reveal complex relationships between phenotypic robustness, pleiotropy, and fitness. Mol. Syst. Biol. 2015, 11, 773. [Google Scholar] [CrossRef]
- Jakab, Á.; Mogavero, S.; Förster, T.M.; Pekmezovic, M.; Jablonowski, N.; Dombrádi, V.; Pócsi, I.; Hube, B. Effects of the glucocorticoid betamethasone on the interaction of Candida albicans with human epithelial cells. Microbiology 2016, 162, 2116–2125. [Google Scholar] [CrossRef]
- Nemes, D.; Kovács, R.; Nagy, F.; Mező, M.; Poczok, N.; Ujhelyi, Z.; Pető, Á.; Fehér, P.; Fenyvesi, F.; Váradi, J.; et al. Interaction between different pharmaceutical excipients in liquid dosage forms—Assessment of cytotoxicity and antimicrobial activity. Molecules 2018, 23, 1827. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, B.B.; O’Brien, E.; Khoury, J.B.E.; Mylonakis, E. Methods for using Galleria mellonella as a model host to study fungal pathogenesis. Virulence 2010, 1, 475–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, R. VassarStats. Available online: http://vassarstats.net/index.html (accessed on 11 August 2021).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 3 September 2021).
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Statistics Kingdom: Kaplan-Meier Survival Analysis. Available online: https://www.statskingdom.com/350kaplan_meier.html (accessed on 23 July 2021).
- Sitterlé, E.; Maufrais, C.; Sertour, N.; Palayret, M.; d’Enfert, C.; Bougnoux, M.-E. Within-host genomic diversity of Candida albicans in healthy carriers. Sci. Rep. 2019, 9, 2563. [Google Scholar] [CrossRef]
- Forche, A.; Cromie, G.; Gerstein, A.C.; Solis, N.V.; Pisithkul, T.; Srifa, W.; Jeffery, E.; Abbey, D.; Filler, S.G.; Dudley, A.M.; et al. Rapid phenotypic and genotypic diversification after exposure to the oral host niche in Candida albicans. Genetics 2018, 209, 725–741. [Google Scholar] [CrossRef] [Green Version]
- Phadke, S.S.; Maclean, C.J.; Zhao, S.Y.; Mueller, E.A.; Michelotti, L.A.; Norman, K.L.; Kumar, A.; James, T.Y. Genome-wide screen for Saccharomyces cerevisiae genes contributing to opportunistic pathogenicity in an invertebrate model host. G3 2018, 8, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Grimberg, B.; Zeyl, C. The effects of sex and mutation rate on adaptation in test tubes and to mouse hosts by Saccharomyces cerevisiae. Evolution 2005, 59, 431–438. [Google Scholar] [CrossRef]
- Mroczyńska, M.; Brillowska-Dąbrowska, A. Virulence of clinical Candida isolates. Pathogens 2021, 10, 466. [Google Scholar] [CrossRef]
- Pérez-Torrado, R.; Querol, A. Saccharomyces cerevisiae show low levels of traversal across the human blood brain barrier in vitro. F1000Research 2017, 6, 944. [Google Scholar] [CrossRef]
- Czerucka, D.; Rampal, P. Diversity of Saccharomyces boulardii CNCM I-745 mechanisms of action against intestinal infections. World J. Gastroenterol. 2019, 25, 2188–2203. [Google Scholar] [CrossRef]
- Stier, H.; Bischoff, S.C. Influence of Saccharomyces boulardii CNCM I-745 on the gut-associated immune system. Clin. Exp. Gastroenterol. 2016, 9, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Smith, I.M.; Christensen, J.E.; Arneborg, N.; Jespersen, L. Yeast modulation of human dendritic cell cytokine secretion: An in vitro study. PLoS ONE 2014, 9, 12–14. [Google Scholar]
- Di Paola, M.; Rizzetto, L.; Stefanini, I.; Vitali, F.; Massi-Benedetti, C.; Tocci, N.; Romani, L.; Ramazzotti, M.; Lionetti, P.; De Filippo, C.; et al. Comparative immunophenotyping of Saccharomyces cerevisiae and Candida spp. strains from Crohn’s disease patients and their interactions with the gut microbiome. J. Transl. Autoimmun. 2020, 3, 100036. [Google Scholar] [CrossRef] [PubMed]
- Rizetto, L.; Kuka, M.; De Filippo, C.; Cambi, A.; Netea, M.G.; Beltrame, L.; Napolitani, G.; Torcia, M.G.; D’Oro, U.; Cavalieri, D. Differential IL-17 production and mannan recognition contribute to fungal pathogenicity and commensalism. J. Immunol. 2010, 184, 4258–4268. [Google Scholar] [CrossRef] [Green Version]
- Kelesidis, T.; Pothoulakis, C. Efficacy and safety of the probiotic Saccharomyces boulardii for the prevention and therapy of gastrointestinal disorders. Therap. Adv. Gastroenterol. 2012, 5, 111–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennequin, C.; Kauffmann-Lacroix, C.; Jobert, A.; Viard, J.P.; Ricour, C.; Jacquemin, J.L.; Berche, P. Possible role of catheters in Saccharomyces boulardii fungemia. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 16–20. [Google Scholar] [CrossRef]
- Cassone, M.; Serra, P.; Mondello, F.; Girolamo, A.; Scafetti, S.; Pistella, E.; Venditti, M. Outbreak of Saccharomyces cerevisiae Subtype boulardii fungemia in patients neighboring those treated with a probiotic preparation of the organism. J. Clin. Microbiol. 2003, 41, 5340–5343. [Google Scholar] [CrossRef] [Green Version]
- Cohen, L.; Ranque, S.; Raoult, D. Saccharomyces cerevisiae boulardii transient fungemia after intravenous self-inoculation. Med. Mycol. Case Rep. 2013, 2, 63–64. [Google Scholar] [CrossRef] [PubMed]
- Fadhel, M.; Patel, S.; Liu, E.; Levitt, M.; Asif, A. Saccharomyces cerevisiae fungemia in a critically ill patient with acute cholangitis and long term probiotic use. Med. Mycol. Case Rep. 2019, 23, 23–25. [Google Scholar] [CrossRef]
- Dauby, N. Risks of Saccharomyces boulardii-containing probiotics for the prevention of Clostridium difficile Infection in the elderly. Gastroenterology 2017, 153, 1450–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragonnaud, E.; Biragyn, A. Gut microbiota as the key controllers of “healthy” aging of elderly people. Immun. Ageing. 2021, 18, 2. [Google Scholar] [CrossRef] [PubMed]
- Santino, I.; Alari, A.; Bono, S.; Teti, E.; Marangi, M.; Bernardini, A.; Magrini, L.; Di Somma, S.; Teggi, A. Saccharomyces cerevisiae fungemia, a possible consequence of the treatment of Clostridium difficile colitis with a probioticum. Int. J. Immunopathol. Pharmacol. 2014, 27, 143–146. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Isolate | Type | Formulation | Component | Place of Acquisition | Date of Acquisition | Country of Manufacturing | Reference | ||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | PY0001 | probiotic supplement | active dry | single component | Debrecen, Hungary | March 2015 | France | [16,34] | ||
| 2 | PY0002 | probiotic supplement | active dry | single component | Debrecen, Hungary | November 2017 | France | [16] | ||
| 3 | PY0003 | probiotic supplement | active dry | multicomponent | Debrecen, Hungary | September 2017 | Czechia | [16] | ||
| 4 | PY0004 | probiotic supplement | active dry | multicomponent | Debrecen, Hungary | November 2017 | Czechia | [16] | ||
| ID | Isolate | Type | Age (yr) at Sampling | Sex | Prevailing Medical Condition during Isolation | Mycosis Case | Anatomical Origin/Sample Type | Date of Sampling | Geographic Origin | Reference |
| 5 | DE6507 | clinical isolate (probiotic-derived) | 63 | ♂ | pneumonia | yes | haemoculture | 18 February 2017 | Debrecen, University Clinic | [16] |
| 6 | DE35762 | clinical isolate (probiotic-derived) | 66 | ♀ | respiratory failure | yes | haemoculture | 5 November 2015 | Debrecen, University Clinic | [16] |
| 7 | DE27020 | clinical isolate (probiotic-derived) | 40 | ♀ | sepsis (bacterial) | no | bronchus (sampling during intubation) | 23 August 2015 | Debrecen, University Clinic | [16,34] |
| 8 | DE3912 | clinical isolate (probiotic-derived) | 85 | ♂ | pneumonia | no | trachea (sampling from tracheal cannula) | 31 January 2018 | Debrecen, University Clinic | [16] |
| 9 | DE42533 | clinical isolate (probiotic-derived) | 2 | ♂ | fluid homeostasis disorder | no | throat | 15 December 2017 | Debrecen, University Clinic | [16] |
| 10 | DE42807 | clinical isolate (probiotic-derived) | 1 | ♀ | diarrhea | no | vagina | 4 December 2017 | Debrecen, University Clinic | [16] |
| 11 | DE45866 | clinical isolate (probiotic-derived) | 64 | ♂ | cerebral infarction | no | bronchus (sampling during intubation) | 29 December 2017 | Debrecen, University Clinic | [16] |
| 12 | 465/2018 | clinical isolate (probiotic-derived) | 41 | ♀ | amenorrhea | no | vagina | 3 January 2018 | Szeged, University Clinic | - |
| 13 | 551/2018 | clinical isolate (probiotic-derived) | 81 | ♂ | paralytic ileus | no | feces | 3 January 2018 | Szeged, University Clinic | - |
| 14 | 2251/2018 | clinical isolate (probiotic-derived) | 17 | ♂ | ulcerative colitis | no | feces | 8 January 2018 | Szeged, University Clinic | - |
| Biofilm Formation on Plastic Surface | Secreted Enzymatic Virulence Factors | Hemolytic Index, 37 °C | Galleria Larva Survival | ||||||
|---|---|---|---|---|---|---|---|---|---|
| YPD Liquid, 37 °C, 3 d | Phospholipase Secretion, Pz Value, 37 °C | Aspartate Protease Secretion, Prz Value, 37 °C | α-Hemolysis, 1 d | α-Hemolysis, 2 d | β-Hemolysis, 2 d | β-Hemolysis, 3 d | p Value Log-Rank Test | ||
| ANOVA p Value | 0.53792 | <0.0001 | 0.00577 | 0.03899 | 0.36651 | 0.91412 | 0.00182 | ||
| Commercial vs. Mycosis (Tukey HSD) | n.s. (p ≥ 0.05) | n.s. (p ≥ 0.05) | n.s. (p ≥ 0.05) | C > M (p < 0.05) | n.s. (p ≥ 0.05) | n.s. (p ≥ 0.05) | C < M (p < 0.01) | Commercial vs. Mycosis | 0.30283 |
| Commercial vs. Non-mycosis (Tukey HSD) | n.s. (p ≥ 0.05) | C < NM (p < 0.01) | C > NM (p < 0.05) | n.s. (p ≥ 0.05) | n.s. (p ≥ 0.05) | n.s. (p ≥ 0.05) | n.s. (p ≥ 0.05) | Commercial vs. Non-mycosis | <0.0001 |
| Mycosis vs. Non-mycosis (Tukey HSD) | n.s. (p ≥ 0.05) | M < NM (p < 0.01) | n.s. (p ≥ 0.05) | n.s. (p ≥ 0.05) | n.s. (p ≥ 0.05) | n.s. (p ≥ 0.05) | M > NM (p < 0.01) | Mycosis vs. Non-mycosis | <0.0001 |
| Trait | Phagocytosis | CD40 (Costimulatory) | CD80 (Costimulatory) | CD86 (Costimulatory) | CD83 (Maturation Marker) | HLA- DQ (Antigen-Presenting) | IL-6 (Proinflammatory) | IL-8 (Chemokine) | TNFα (Proinflammatory) | IL-12 (Proinflammatory) | IL-1β (Proinflammatory) | IL-17 (Proinflammatory) | IFN-γ (Proinflammatory) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Donor-dependence | ANOV A/T-test p-value | 0.10721 | <0.00010 | <0.0001 | <0.0001 | <0.0001 | 0.54060 | <0.0001 | <0.0001 | <0.0001 | 0.07714 (T-test) | <0.0001 | <0.0001 | 0.00026 (T-test) |
| Tukey HSD/T-test | none | D1 < D3 (p < 0.01) | D1 < D3 (p < 0.01) | D1 < D2 (p < 0.01) | D1 > D2 | none | D1 > D3 | D1 > D2 (p < 0.05) | D1 > D3 (p < 0.01) | none (T-test) | D1 > D2 (p < 0.01) | D1 < D2 (p < 0.01) | D3 > D4 (T-test) | |
| D2 < D3 (p < 0.01) | D1 < D4 (p < 0.01) | D1 < D3 (p < 0.01) | D1 < D3 (p < 0.0) | D2 > D3 (p < 0.0) | D1 > D3 (p < 0.01) | D1 > D4 (p < 0.01) | D1 > D4 (p < 0.01) | D1 > D4 (p < 0.01) | ||||||
| D3 > D4 (p < 0.01) | D2 < D3 (p < 0.01) | D1 < D4 (p < 0.01) | D1 > D4 | D3 < D 4 | D1 > D4 (p < 0.01) | D2 > D3 (p < 0.01) | D2 > D4 (p < 0.01) | D2 > D4 (p < 0.01) | ||||||
| D2 < D4 (p < 0.01) | D2 < D3 (p < 0.01) | D2 < D3 | D2 > D3 (p < 0.01) | D2 > D4 (p < 0.01) | ||||||||||
| D3 > D4 (p < 0.01) | D2 < D4 (p < 0.01) | D2 > D4 (p < 0.0) | D3 < D4 (p < 0.01) | |||||||||||
| D3 > D4 (p < 0.01) | D3 > D4 | |||||||||||||
| Commercial vs. Mycosis | T-test | 0.64298 | 0.87434 | 1.0000 | 0.98422 | 0.70758 | 0.56780 | 0.88993 | 0.65003 | 0.54811 | 0.31408 | 0.89588 | 0.03792 (C < M) | 0.11320 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imre, A.; Kovács, R.; Pázmándi, K.; Nemes, D.; Jakab, Á.; Fekete, T.; Rácz, H.V.; Dóczi, I.; Bácskay, I.; Gácser, A.; et al. Virulence Factors and in-Host Selection on Phenotypes in Infectious Probiotic Yeast Isolates (Saccharomyces ‘boulardii’). J. Fungi 2021, 7, 746. https://doi.org/10.3390/jof7090746
Imre A, Kovács R, Pázmándi K, Nemes D, Jakab Á, Fekete T, Rácz HV, Dóczi I, Bácskay I, Gácser A, et al. Virulence Factors and in-Host Selection on Phenotypes in Infectious Probiotic Yeast Isolates (Saccharomyces ‘boulardii’). Journal of Fungi. 2021; 7(9):746. https://doi.org/10.3390/jof7090746
Chicago/Turabian StyleImre, Alexandra, Renátó Kovács, Kitti Pázmándi, Dániel Nemes, Ágnes Jakab, Tünde Fekete, Hanna Viktória Rácz, Ilona Dóczi, Ildikó Bácskay, Attila Gácser, and et al. 2021. "Virulence Factors and in-Host Selection on Phenotypes in Infectious Probiotic Yeast Isolates (Saccharomyces ‘boulardii’)" Journal of Fungi 7, no. 9: 746. https://doi.org/10.3390/jof7090746