
Phylogeny and Systematics of the Genus Tolypocladium (Ophiocordycipitaceae, Hypocreales)

1
Yunnan Herbal Laboratory, College of Ecology and Environmental Sciences, Yunnan University, Kunming 650504, China
2
The International Joint Research Center for Sustainable Utilization of Cordyceps Bioresources in China and Southeast Asia, Yunnan University, Kunming 650504, China
*
Authors to whom correspondence should be addressed.
J. Fungi 2022, 8(11), 1158; https://doi.org/10.3390/jof8111158
Submission received: 30 August 2022
/
Revised: 29 October 2022
/
Accepted: 30 October 2022
/
Published: 1 November 2022
(This article belongs to the Section Fungal Evolution, Biodiversity and Systematics)
Abstract
:The taxonomy and phylogeny of the genus Tolypocladium are herein revised based on the most comprehensive dataset to date. Two species-level phylogenies of Tolypocladium were constructed: a single-gene phylogeny (ITS) of 35 accepted species and a multigene phylogeny (nrSSU, nrLSU, tef-1α, rpb1, and rpb2) of 27 accepted species. Three new species, Tolypocladium pseudoalbum sp. nov., Tolypocladium subparadoxum sp. nov., and Tolypocladium yunnanense sp. nov., are described in the present study. The genetic divergences of four markers (ITS, tef-1α, rpb1 and rpb2) among Tolypocladium species are also reported. The results indicated that species of Tolypocladium were best delimited by rpb1 sequence data, followed by the sequence data for the rpb2, tef-1α, and ITS provided regions. Finally, a key to the 48 accepted species of Tolypocladium worldwide is provided.
1. Introduction
Tolypocladium was originally described as an anamorph genus by Gams in 1971 to accommodate three species collected from soil: T. cylindrosporum W. Gams, T. geodes W. Gams, and T. inflatum W. Gams [1]. Subsequently, the species T. lignicola G.L. Barron, T. parasiticum G.L. Barron, and T. trigonosporum G.L. Barron, all of which were isolated from bdelloid rotifers, were added to this genus [2,3,4]. Bissett described T. nubicola and T. tundrense from soil in 1983 [5] and reassigned three species to Tolypocladium: T. balanoide (basionym: Cephalosporium balanoide), T. microsporum (basionym: Verticillium microsporum) and T. niveum (basionym: Pachybasium niveum). Additionally, Bissett [5] noted that the morphological characteristics of T. niveum were similar to those of T. inflatum. Because T. niveum precedes T. inflatum, Bissett proposed that T. inflatum be synonymized with T. niveum [5]. However, Dreyfuss observed that T. inflatum produces cyclosporine and is the type species of the genus Tolypocladium. The name T. inflatum is also commonly accepted [6]. Therefore, Dreyfuss rejected the synonymization of T. inflatum with T. niveum [6]. The genus Tolypocladium is morphologically characterized by sparingly branched conidiophores, swollen phialides, and one-celled conidia borne in slimy heads. Approximately 20 species have been included in the Tolypocladium based on morphological characteristics.
The taxonomy of Tolypocladium has been discussed extensively for decades. Cordyceps sensu lato was recently reclassified into three families (Clavicipitaceae sensu stricto, Cordycipitaceae, and Ophiocordycipitaceae) and four genera (Cordyceps s. str., Elaphocordyceps, Metacordyceps, and Ophiocordyceps) based on multigene phylogeny [7]. Molecular phylogenetic analyses suggested that Tolypocladium species fall within the Ophicordycipitaceae [7,8]. The genus Elaphocordyceps Sung and Spatafora 2007 was proposed for 23 species of the Cordyceps Fr. (1818: 316); these species parasitize the fungal genus Elaphomyces and some species of arthropods (e.g., cicada nymphs and beetle larvae) [7]. The Elaphocordyceps species within the Ophiocordycipitaceae form a clade sister to those of the genus Ophiocordyceps. Gams established the Chaunopycnis to accommodate C. alba, which morphologically resembles Tolypocladium species in its conidiogenesis [9]. With the end of dual nomenclature for fungi, the generic name Tolypocladium was chosen over Elaphocordyceps and Chaunopycnis as Tolypocladium is the oldest and most commonly used name [8]. Chaunopycnis was integrated into the genus Tolypocladium. Accordingly, C. alba, C. ovalispora, and C. pustulata were renamed T. album, T. ovalisporum, and T. pustulatum, respectively [8].
At present, 53 Tolypocladium records, including 5 varieties, are listed in the Index Fungorum (www.indexfungorum.org, accessed on 28 August 2022). Tolypocladium balanoides, which was reassigned to Drechmeria (as Drechmeria balanoides), and Tolypocladium parasiticum, which was reassigned to Metapochonia (as Tolypocladium parasiticum), should be excluded from the Tolypocladium. However, some of these records are doubtful, because the original identifications were presumptive based on host associations or based on the morphology of only one or two ascospore stages of the asexual or sexual morph. For 16 species, no molecular data are available in the GenBank database [10]. Tolypocladium species have a cosmopolitan distribution and a broad host range that includes bdelloid rotifers, mosquito larvae, nematodes, fireflies, beetles, cicada nymphs, batmoth larvae, macrocystic fungi, Ophiocordyceps sinensis, and even plants (as endophytes) [2,3,11,12,13,14,15,16,17,18,19].
Tolypocladium species have been widely studied due to their importance in the medicinal domain. These species can produce cyclosporine A, tolypoalbin, tolypin, cyclosporine D hydroperoxide, cylindromicin, and tolyprolinol [20,21], all of which have significant antitumoral, anti-inflammatory, antifungal, and/or antiparasitic properties [22]. Cyclosporine A, which is naturally isolated from T. inflatum, is widely used in autoimmune disease treatment and to prevent allograft rejection [23,24,25]. Tolypoalbin is a peptide mixture and a tetrameric acid produced by T. album [26]. Tolypin is also a peptide mixture [27]. Like kojic acid, cylindromicin is a significant bioactive inhibitor of tyrosinase [28]. Tolyprolinol, a dipeptide produced by Tolypocladium sp. FKI-7981, contains a rare moiety prolinol and was the first natural product isolated from Tolypocladium species. Tolyprolinol exhibits moderate antimalarial activity without cytotoxicity or any other antimicrobial properties [29].
Recent investigations and phylogenetic analyses have ascribed many new taxa to Tolypocladium. Therefore, the diversity of Tolypocladium may be underestimated. In the present study, we aimed first to investigate and document the worldwide diversity of Tolypocladium fungi using our current collection of specimens and data collected over the last several years. We used comprehensive morphological and molecular phylogenetic reconstructions to identify and reevaluate our specimens. Based on these reconstructions, we herein describe and illustrate three new taxa. We then clarify the phylogenetic affinities of these new taxa using rDNA sequence analyses.
2. Materials and Methods
2.1. Sampling
Tolypocladium species were collected in Kunming, Pu’er, Yunnan, China. Voucher specimens and the corresponding isolated strains were deposited in the Yunnan Herbal Herbarium (YHH) and the Yunnan Fungal Culture Collection (YFCC), respectively, of Yunnan University, Kunming, China.
Tolypocladium strains were isolated from soil samples, as described in our previous publication [30]. In brief, 2 g of soil was added to a flask containing 20 mL of sterilized water and glass beads. The suspension was then shaken for 10 min and diluted 100 times. Finally, 200 µL of diluted soil suspension was spread on petri dishes containing solidified onion garlic agar (OGA: 1 L of distilled water, 20 g of grated garlic, and 20 g of onion were boiled together for 1 h; the boiled biomass was filtered and 2% agar was added to the filtrate). Czapek yeast extract agar (CYA; Advanced Technology and Industrial Co., Ltd., Hong Kong, China) and potato dextrose agar (PDA; Difco, USA) were used. Rose bengal (50 mg/L) and kanamycin (100 mg/L) were added to all media. Conidia grown on insect cadavers were transferred to PDA plates and cultured at 22 °C. The filamentous fungal colonies isolated from the culture were transferred to fresh PDA media. The purified fungal strains were maintained at 22 °C in a culture room or transferred to PDA slants and stored at 4 °C.
2.2. Morphological Studies
Morphological studies were performed as described in our previous study [31]. Micromorphological characteristics, such as phialides and conidia, were studied by picking and mounting cultures on glass slides. The sizes and shapes of the microcharacteristics were determined using an Olympus CX40 and BX53 (Olympus Corporation, Tokyo, Japan). Individual length and width measurements were taken for 20–30 replicates, including the absolute minima and maxima. The morphological characteristics were described based on the digital images and the measurement dataset.
2.3. Molecular Studies
2.3.1. DNA Extraction and PCR Amplification
Total DNA was extracted from the fungal mycelia on PDA plates or from herbarium materials using the modified CTAB procedure [32]. The primer pair nrSSU-CoF and nrSSU-CoR [33] was used to amplify nrSSU, the primer pair LR5 and LR0R [34,35] was used to amplify nrLSU, and the primer pair EF1α-EF and EF1α-ER [7,36] was used to amplify the translation elongation factor 1α (tef-1α). The primer pair RPB1-5′F and RPB1-5′R and the primer pair RPB2-5′F and RPB2-5′R [7,36] were used to amplify the largest and second-largest subunits of RNA polymerase II (rpb1 and rpb2), respectively. The ITS fragment was amplified using the primer pair ITS5 and ITS4 [37].
The matrix for the polymerase chain reaction (PCR) was comprised of 2.5 μL PCR 10× buffer (2 mmol/L Mg2+) (Transgen Biotech, Beijing, China), 1 μL forward primer (10 µmol/L), 1 μL reverse primer (10 µmol/L), 0.25 μL Taq DNA polymerase (Transgen Biotech, Beijing, China), 2 μL dNTP (2.5 mmol/L), 1 μL DNA template (500 ng/μL), and 17.25 μL sterile ddH2O. Amplification reactions were performed in a Bio-Rad T100 thermal cycler (Bio-Rad Laboratories, CA, USA). The PCR cycling conditions for the amplification of nrSSU were as follows: 95 °C for 4 min; eight cycles of 94 °C for 50 s, 56 °C for 50 s, and 72 °C for 2 min, with the annealing temperature decreasing 0.5 °C/cycle; 25 cycles of 94 °C for 50 s, 52 °C for 50 s, and 72 °C for 2 min; and 72 °C for 10 min. The nucleotide sequences of ITS, nrLSU, tef-1α, rpb1, and rpb2 were amplified using the following cycling conditions: 95 °C for 4 min; eight cycles of 94 °C for 50 s, 56 °C for 50 s, and 72 °C for 70 s, with the annealing temperature decreasing 0.5 °C/cycle; 25 cycles of 94 °C for 50 s, 52 °C for 50 s, and 72 °C for 70 s; and 72 °C for 10 min. PCR products were purified using a gel extraction and PCR purification combo kit (Beijing Genomics Institute, Shenzhen, China) and sequenced on an automatic sequence analyzer (BGI Co., Ltd., Shenzhen, China) using the amplification primers.
2.3.2. DNA Sequence Alignments
To investigate the placement of our samples within Tolypocladium, the nucleotide sequences of ITS, nrSSU, nrLSU, tef-1α, rpb1, and rpb2 were compared with sequences from representative Tolypocladium species downloaded from GenBank (Table 1, Figure 1 and Figure 2). Individual gene sequence datasets (ITS, nrSSU, nrLSU, tef-1α, rpb1, and rpb2) were aligned and manually checked using Bioedit v7.0.9 [38]. To identify possible phylogenetic conflicts among the datasets, the partition homogeneity (PH) test was performed with 1000 randomized replicates of heuristic searches with simple sequence addition in PAUP* 4.0a166 (http://paup.phylosolutions.com, accessed on 28 August 2022) [39]. The results showed that the phylogenetic signals from the five gene markers were in conflict.
2.3.3. Phylogenetic Analyses
Phylogenetic analyses were based on a concatenated five-gene dataset and the ITS sequences alone. nrSSU, nrLSU, tef-1α, rpb1, rpb2, and ITS sequences were retrieved from GenBank, and combined with those generated in this study. Taxon information and GenBank accession numbers are given in Table 1. Sequences were aligned using Clustal X2.0 and MEGA v6.06 [40,41]. Group I introns in the nrSSU sequences of some species were excluded from the phylogenetic analyses, and gaps were treated as missing data. After alignment of the five genes individually, the alignments were concatenated. A partition homogeneity test was conducted in PAUP* 4.0a166 [39], and the results indicated that there were no conflicts among the data partitions. PartitionFinder V1.1.1 identified eleven data partitions: nine corresponding to the three codon positions in each of the protein-coding genes (tef-1α, rpb1, and rpb2) and one each for nrLSU and nrSSU [42,43]. The results showed that the phylogenetic signals of the five genes were congruent (p = 0.02).
Maximum likelihood (ML) phylogenetic analyses were conducted using RaxML 7.0.3 [44] with the recommended partition parameters and 1000 rapid bootstrap replicates. Bayesian posterior probabilities (BP) were estimated with the same partition parameters using MrBayes v3.1.2 [45]. Bayesian inference (BI) analysis ran in MrBayes v3.1.2 for 5 million generations. Maximum parsimony (MP) analysis of the ITS dataset was performed using PAUP v. 4.0a166 [39], adopting the random addition of sequences model (10 replications), with gaps treated as missing data. A bootstrap (MPBS) analysis was performed using the maximum parsimony criterion in 1000 replications.
The following taxa were included in the five-gene concatenated dataset: Drechmeria W. Gams and H.-B. Jansson, Harposporium Lohde, Ophiocordyceps Petch, Purpureocillium Luangsa-Ard, Hywel-Jones, Houbraken and Samson, and Tolypocladium. Two species of Polycephalomyces Kobayasi were used as outgroups. ITS analysis was performed on Tolypocladium taxa only. Phylogenetic trees were visualized with FigTree v1.4.0 [46], edited in Microsoft PowerPoint, saved in PDF format, and converted to JPG format using Adobe Illustrator CS6 (Adobe Systems Inc., San Jose, USA). The finalized alignments and trees were submitted to TreeBASE (multigene submission ID 29808).
We calculated a phylogenetic distance matrix for the markers ITS, tef-1α, rpb1, and rpb2 to assess the species boundaries of the 10 Tolypocladium species (Supplementary Tables S1–S4), because the sequence data were complete for these four loci. The paired distances among the 10 Tolypocladium lineages were measured using the Kimura two-parameter model in MEGA v6.06 [41].
3. Results
3.1. Sequence Alignment and Phylogenetic Analyses
ITS, nrSSU, nrLSU, tef-1α, rpb1, and rpb2 sequences were generated from ten living cultures (accession numbers are given in Table 1). The concatenated five-gene alignment of 113 taxa contained 5371 base pairs in total: nrSSU, 1488 bp; nrLSU, 987 bp; tef-1α, 998 bp; rpb1, 756 bp; and rpb2, 1142 bp. Polycephalomyces formosus ARSEF 1424 and Polycephalomyces sinensis CN 80-2 were used as the outgroup sequences for the five-gene phylogenetic analyses. Both BI and ML analyses recovered six well-supported clades corresponding to the Ophiocordyceps (ML bootstrap, BS = 85% and bayesian posterior probability, BP = 1), Tolypocladium (BS = 99%, BP = 1), Purpureocillium (BS = 97%, BP = 1), Drechmeria (BS = 97%, BP = 1), Harposporium (BS = 88%, BP = 1), and Polycephalomyces (BS = 100%, BP = 1) (Figure 1) within Ophiocordycipitaceae. Phylogenetically, the Tolypocladium clade is the closest to the Ophiocordyceps clade, and it is well supported in this and other published analyses [7,8]. According to the current data, relationships for species in the Tolypocladium clade show strong statistical support for internal branches. Most sexual species are located at the top of the Tolypocladium clade, and asexual species are located at the bottom of the Tolypocladium clade, except T. subparadoxum and T. paradoxum. Three new species (i.e., Tolypocladium pseudoalbum sp. nov., Tolypocladium subparadoxum sp. nov., and Tolypocladium yunnanense sp. nov.) were recognized in Tolypocladium (shown in boldface in Figure 1). T. pseudoalbum sp. nov. formed a clade with T. pustulatum, T. tropicale, T. endophyticum, T. amazonense, and T. yunnanense sp. nov. (Figure 1), while T. subparadoxum sp. nov. formed a well-supported clade with Tolypocladium sp. and T. paradoxum (Figure 1). T. yunnanense sp. nov. was close to five other species: T. pustulatum, T. tropicale, T. endophyticum, T. amazonense, and T. pseudoalbum sp. nov. (Figure 1).
The ITS dataset used for phylogenetic analyses comprised 769 base pairs of sequence data for 61 taxa. Purpureocillium lilacinum CBS 284.36 and Purpureocillium lilacinum NHJ 3497 were chosen as outgroup sequences. The three phylogenetic algorithms (BI, ML, and MP) recovered trees with similar topologies (Figure 2). The three new species described herein (i.e., Tolypocladium pseudoalbum sp. nov., Tolypocladium subparadoxum sp. nov., and Tolypocladium yunnanense sp. nov.) formed an independent lineage with Tolypocladium (Figure 2).
3.2. Genetic Distance Analyses
Comparisons of genetic divergence showed that (1) the minimum thresholds (p-distances) required to distinguish species within the Tolypocladium lineages were 0.026, 0.017, 0.013, and 0.008 for tef-1α, rpb1, rpb2, and ITS, respectively (Supplementary Tables S1–S4); and (2) the phylogenetic relationships within Tolypocladium were best resolved by the rpb1 sequence data, followed by those of rpb2, tef-1α, and ITS (Supplementary Tables S1–S4).
3.3. Taxonomy
Tolypocladium W. Gams, Persoonia 6(2): 185 (1971). emend. C. A. Quandt et al. IMA Fungus 5: 125 (2014).
Synonyms: Chaunopycnis W. Gams, Persoonia 11: 75 (1980).
Elaphocordyceps G. H. Sung and Spatafora, Stud. Mycol. 57: 36 (2007).
Sexual morph: Stromata are solitary or several, simple or branched. The stipe is tough, dark-brownish to greenish, cylindrical, and abruptly to enlarging in the fertile part. The fertile part is cylindrical to clavate. Perithecia are superficial, wholly or partially immersed, ordinal or oblique in arrangement. Asci are cylindrical with a thickened ascus apex. Ascospores are usually cylindrical, multiseptate, disarticulate into part spores, and are occasionally non-disarticulating. Part spores are cylindrical.
Asexual morph: Tolypocladium-like, Chaunopycnis-like, or Verticillium-like. Conidiophores typically are short and bear whorls of phialides. Phialides often have bent necks and are usually swollen at the base. Conidia are ellipsoidal, globose, or reniform, and aggregate in small heads at the tips of the phialides.
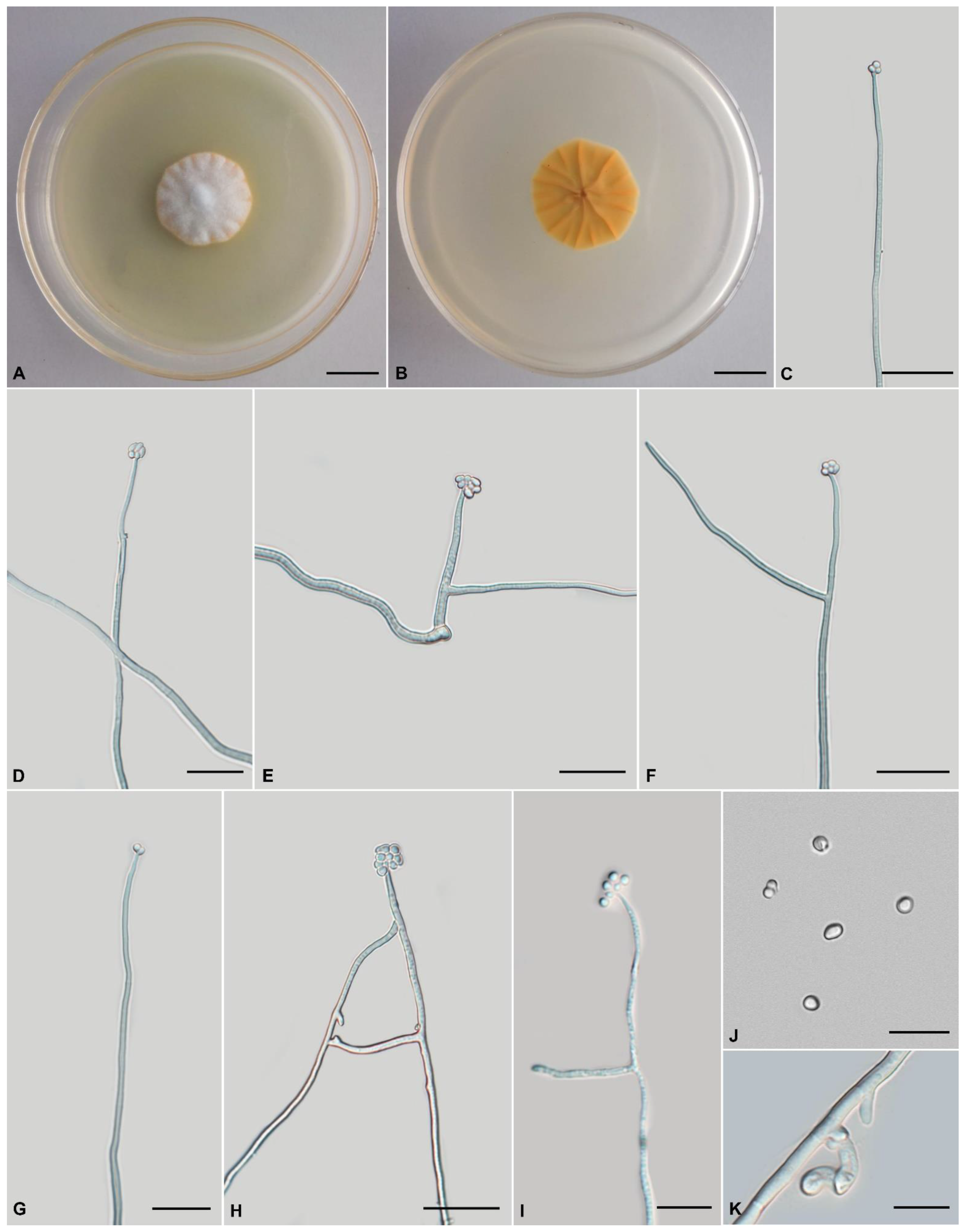
Tolypocladium pseudoalbum, H. Yu, Y. Wang and Q.Y. Dong, sp. nov., Figure 3.
MycoBank: MB 845430.
Etymology: Referring to the morphological resemblance of this species to Tolypocladium album, despite its phylogenetic dissimilarity.
Type: China, Yunnan Province, Kunming City, Wild Duck Forest Park (25°13′ N, 102°87′ E, 2100 m above sea level), from the soil on the forest floor, 10 August 2019, Yao Wang (holotype: YHH 875, dried specimen; ex-type living culture: YFCC 875).
Teleomorph: Unknown.
Anamorph: Colonies on PDA are moderately fast-growing, attaining a diameter of 42–44 mm in 21 days at 22 °C. Colonies pulvinate, with high mycelial density, white or pale yellow, reverse deep yellow. Hyphae branched, smooth-walled, septate, hyaline, 1.1–2.7 μm wide. Cultures readily produce phialides and conidia on PDA after two weeks at room temperature. Phialides arising from aerial hyphae, solitary, 12.3–48.5 × 1.0–2.0 μm, cylindrical, tapering gradually toward the apex, neck 1.4–4.6 × 0.8–1.8 µm. Conidia hyaline, one-celled, globose to broadly ellipsoidal 1.8–3.4 × 1.3–1.9 μm. Chlamydospores present.
Habitat: Soil.
Known distribution: China.
Additional specimens examined: China, Yunnan Province, Kunming City, Songming County, Dashao Village (25°23′ N, 102°33′ E, 2700 m above sea level), from the soil on the forest floor, 12 August 2018, Yao Wang (living culture: YFCC 876).
Comments: Five species are closely related to T. pseudoalbum sp. nov., i.e., T. pustulatum, T. tropicale, T. endophyticum, T. amazonense, and T. yunnanense sp. nov. This clade is characterized by cylindrical to lageniform phialides, globose to broadly ellipsoidal conidia, and primarily white colonies. The phialides of T. pseudoalbum sp. nov. (12.3–48.5 × 1.0–2.0 μm) are longer than those of T. album (3.5–10 × 1.0–1.5 µm).
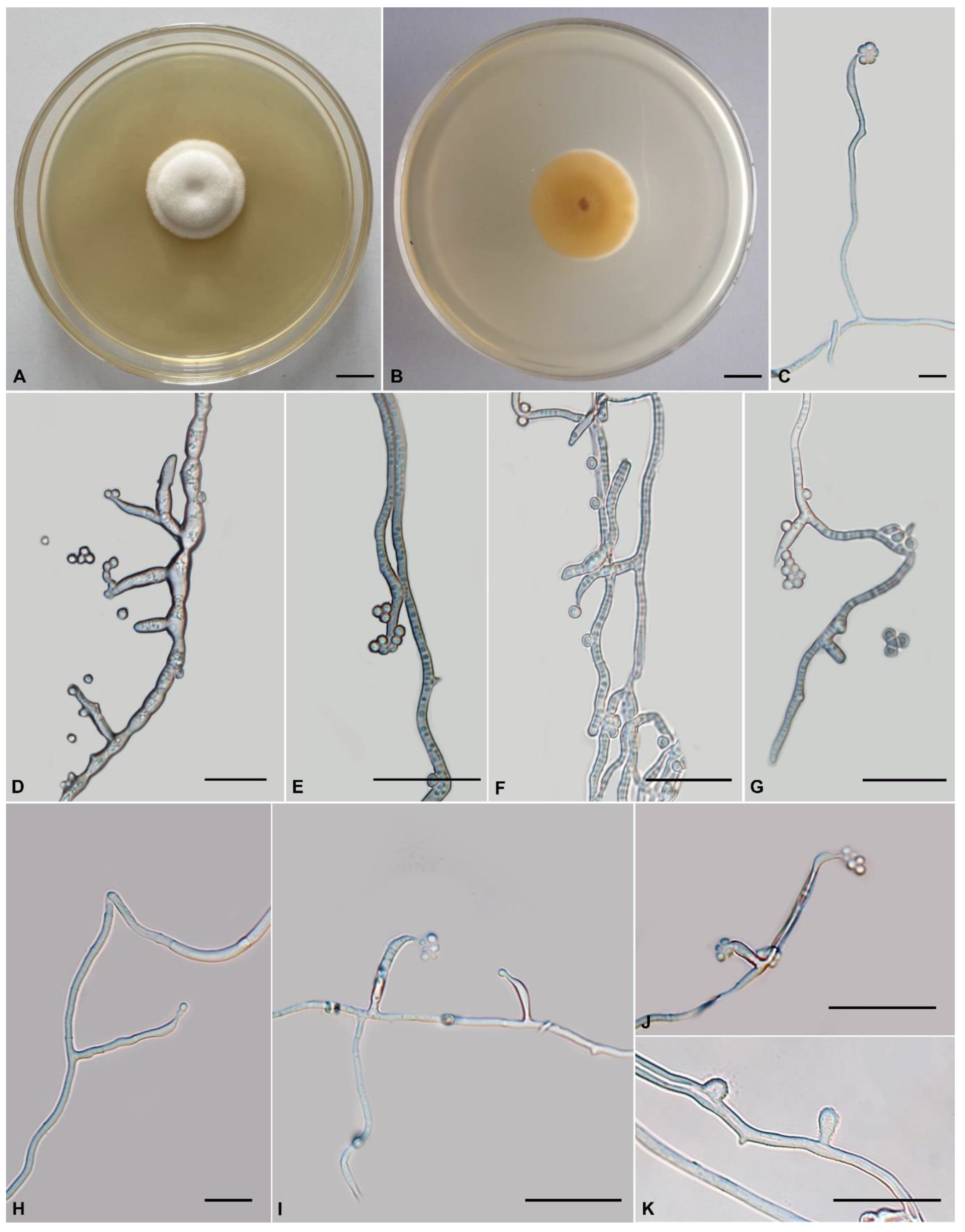
Tolypocladium subparadoxum H. Yu, Y. Wang and Q.Y. Dong, sp. nov., Figure 4.
MycoBank: MB 845431.
Etymology: Referring to the phylogenetic placement is closely related to T. paradoxum.
Holotype: China, Yunnan Province, Pu’er City, Simao District (22°43′ N, 100°58′ E, 1360 m above sea level), from soil on the forest floor, 27 August 2021, Yao Wang (holotype: YHH 879, dried specimen; ex-type living culture: YFCC 879).
Teleomorph: Not observed.
Anamorph: Colonies on PDA are moderately fast-growing, attaining a diameter of 36–38 mm in 21 days at 22 °C. Colonies flocculent, fluffy, with low mycelial density, white or pale yellow, reverse deep yellow. Hyphae smooth-walled, branched, septate, hyaline, 0.8–2.2 μm wide. Cultures produce phialides and conidia on PDA after two weeks at room temperature. Phialides arising from aerial hyphae, solitary, or in verticils of two to four, 5.4–40.1 × 0.9–1.8 μm, cylindrical, tapering gradually toward the apex, neck 3.2–5 × 0.7–1.2 µm. Conidia hyaline, one-celled, ellipsoidal or globose, single or aggregating in heads at the apex of phialides, 2.6–6.5 × 1.0–2.9 μm. Chlamydospores not observed.
Habitat: Soil, larvae of cicada.
Known distribution: China, Japan.
Additional specimens examined: NBRC 106958, Niryo, Takatsuki-shi, Osaka Prefecture.
Comments: Our phylogenetic analysis indicates that Tolypocladium subparadoxum sp. nov. is closely related to Tolypocladium sp. and T. paradoxum. The two strains (YFCC 879 and NBRC 106958) formed a distinct lineage. NBRC 106958 was firstly isolated from cicada in Japan by S. Ban (https://www.nite.go.jp/nbrc/catalogue/NBRCCatalogueDetailServlet?ID=NBRCandCAT=00106958, accessed on 28 August 2022) and subsequently isolated from soil in China (YFCC 879). Since no significant morphological differences were found between the Chinese collections and that of Japan (Supplementary Figure S1), we treated YFCC 879 and NBRC 106958 as Tolypocladium subparadoxum. Tolypocladium paradoxum was originally described as Cordyceps paradoxa by Kobayasi, which was a cicada pathogen that produces solitary, pale ochraceous to dark olivaceous, fleshy stromata with cylindrical asci, breaking into cylindrical part spores [76]. Morphologically, T. subparadoxum differs from T. paradoxum in the following aspects. Relatively, T. paradoxum has longer phialides measured 5.8–58.3 × 1.8–4.3 µm, broader neck (0.9–1.9 µm vs 0.7–1.2 µm), and minor conidia (2.3–4.8 × 1.9–5.2 µm vs 2.6–6.5 × 1.0–2.9 μm) (Supplementary Figure S1).
Tolypocladium subparadoxum similar to T. dujiaolongae and sharing cicada host, solitary, or verticillate, cylindrical or conical phialides, globose to ovoid conidia, and conidia aggregating mostly in small heads, but the latter differs by its relatively shorter phialides (11–35 × 1.0–2.7 μm vs 5.4–40.1 × 0.9–1.8 μm) [19]. Our phylogenetic analysis inferred from ITS data (Figure 2) suggests that they represent two distinct species.
Tolypocladium geodes is also similar to T. subparadoxum in their soil habitats and ellipsoidal or globose conidia. However, T. geodes has relatively shorter phialides (5.6–12.4 × 1.4–2.4 µm) and somewhat minor conidia (1.9–2.4 × 1.6–2.0 µm) [5]. Molecular phylogenetic analyses (Figure 1 and Figure 2) indicate that they are distinct species.
Tolypocladium yunnanense H. Yu, Y. Wang and Q.Y. Dong, sp. nov., Figure 5
MycoBank: MB 845432.
Etymology:Yunnanense (Lat.) refers to the type locality (Yunnan, China).
Holotype: China, Yunnan Province, Kunming City, Wild Duck Forest Park (25°14′ N, 102°87′ E, 2080 m above sea level), from soil on the forest floor, 12 August 2018, Yao Wang (holotype: YHH 877, dried specimen; ex-type living culture: YFCC 877).
Teleomorph: Unknown.
Anamorph: Colonies on PDA are moderately fast-growing, attaining a diameter of 44–46 mm in 21 days at 22 °C. Colonies pulvinate, with high mycelial density, whitish to orange-yellow, reverse deep yellow. Hyphae smooth-walled, branched, septate, hyaline, 1.0–2.4 μm wide. Cultures produce phialides and conidia on PDA after two weeks at room temperature. Phialides are usually curved, solitary, 7.6–62.6 × 0.9–2.3 μm, cylindrical, narrowing slightly or abruptly into a neck, 3–4.2 × 0.5–1 µm. Conidia hyaline, one-celled, elliptical to subglobose, 1.2–2.4 × 0.9–1.9 μm. Chlamydospores present.
Habitat: Soil.
Known distribution: China.
Additional specimens examined: China, Yunnan Province, Pu’er City, Simao District (22°42′ N, 100°57′ E, 1348 m above sea level), from soil on the forest floor, 7 October 2019, Yao Wang (living culture: YFCC 878).
Comments:Tolypocladium yunnanense sp. nov. is characterized by its solitary cylindrical phialides (7.6–62.6 × 0.9–2.3 μm), elliptical to subglobose conidia (1.2–2.4 × 0.9–1.9 μm), and white colonies. The five-gene phylogenetic analysis suggested that T. yunnanense sp. nov. was closely related to five other species (T. pustulatum, T. tropicale, T. endophyticum, T. amazonense and T. pseudoalbum sp. nov.). Phylogenetic analyses of this clade using ITS sequences, for which more complete data were available, showed that T. yunnanense sp. nov. formed clade with T. album, T. pseudoalbum sp. nov., T. tropicale, T. amazonense, and T. endophyticum. Morphologically, Tolypocladium yunnanense sp. nov. has longer phialides than other species in this clade: Tolypocladium yunnanense sp. nov., 7.6–62.6 × 0.9–2.3 μm; T. pustulatum, 4–10 × 2–4 µm, T. tropicale, 4.6 × 1.5 µm; T. endophyticum, 4.1 × 1.6 µm; T. amazonense, 4.1 × 1.6 µm; T. pseudoalbum sp. nov., 12.3–48.5 × 1.0–2.0 μm, and T. album, 3.5–10 × 1.0–1.5 µm.
| Key to Tolypocladium species worldwide | |
| Sexual state observed…………………………………………………………………………………………………………………………..1 | |
| Sexual state not observed…………………………………………………………………………………………………………………..27 | |
| 1a. Perithecia superficial or half-immersed ……………………………………………………………………………………………..2 | |
| 1b. Perithecia completely immersed………………………………………………………………………………………3 | |
| 2a. Perithecia pyriform, relatively larger, 520–550 × 260–280 µm, asci relatively larger, 400–450 × 7–7.5 µm, part spores 2.5–3.0 × 3.0 µm, on cicada nymphs, stromata relatively longer, 14 cm long……………………………………………………………………………………………………………T. inegoense | |
| 2b. Perithecia ovoid, relatively smaller, 320–380 × 220–280 µm, asci cylindrical, smaller, 240–250 × 6 µm, not dissociate into part spores, on Elaphomyces, stromata shorter, 3.5–4.5 cm long……………………………T. ramosum | |
| 3a. Perithecia ellipsoid, subglobose to ovoid…………………………………………………………………………………….4 | |
| 3b. Perithecia ampullaceous……………………………………………………………………………………………………………………………..26 | |
| 4a. From multiple substrate/host (beetle or moth larvae, Larvae of Scarabaeidae (sexual morph); soil, humus, Picea glauca, roots of Picea mariana, the surface of Mycobates sp. (Acari, Mycobatidae), the sclerotium of Ophiocordyceps gracilis (asexual morph)………………………………………………………………….………..T. inflatum | |
| 4b. From simple substrate/host…………………………………………………………………………………………….5 | |
| 5a. On beetle or unidentified host……………………………………………………………………………………………6 | |
| 5b. On Elaphomyces…………………………………………………………………………………………………………..9 | |
| 6a. On the unidentified host, asci relatively wider, 10–15 µm…………………………………………………T. cucullae | |
| 6b. On beetle, asci narrower than 10 µm………………………………………………………………………………….7 | |
| 7a. Stromata was connected to the host through a yellowish rhizomorph-like structure………………..T. fumosum | |
| 7b. Stromata arising directly from the host, never rhizomorphic……………………………………………………….8 | |
| 8a. Part spores short cylindrical, truncate at both ends, 3–5 × 1.5–2 µm……………………………….…T. paradoxum | |
| 8b. Part spores very short, almost cuboid in side view, without flattened ends, 1.5–2.5 × 1.5–1.7 µm…………………………………………………………………………………………………T. toriharamontanum | |
| 9a. Stromata clavate, the fertile part not abruptly enlarged from the stipe………………………………………….10 | |
| 9b. Stromata capitate, the fertile part spherical, oval or cylindrical abruptly enlarged from the stipe………………17 | |
| 10a. Stromata size relatively larger, 10–12 cm long………………………..…………………………………T. jezoense | |
| 10b. Stromata size < 10 cm………………………………………………………………………………………………….11 | |
| 11a. Part spores articulate, moniliform……………………………………………………………………T. szemaoense | |
| 11b. Part spores cylindrical……………………………………………………………………………………………….12 | |
| 12a. Stromata was connected to the host through a rhizomorph-like structure……………………………………………..13 | |
| 12b. Stromata arising directly from the host, never rhizomorphic…………………………………………………….14 | |
| 13a. Fertile part yellowish-green when young, turning olive-green as it matures, perithecia relatively smaller, 480–590 × 195–235 μm…………………………………………………………………………………T. bacillisporum | |
| 13b. Fertile part reddish brown to olivaceous brown, perithecia larger, 600–800 × 250–500 µm…T. ophioglossoides | |
| 14a. Fertile part black, yellow black, dark chestnut brown when dried……………………………..………………15 | |
| 14b. Fertile part pale bluish to grayish blue………………………………………………………T. valvatistipitatum | |
| 15a. Perithecia ≤ 700 µm long…………………………………………..…………………………………………………16 | |
| 15b. Perithecia > 700 µm long (750–1000 × 250–300 µm)……………..…………………………………T. tenuisporum | |
| 16a. Perithecia relatively narrower, 567–697 × 206–248 µm, part spores smaller, 2–5 × 1.5–2 µm, stromata 1.5–3 cm long…………………………………………………………………………………………………..……T. flavonigrum | |
| 16b. Perithecia relatively wider, 500–700 × 250–350 µm, part spores larger, 10–18 × 2.5–4 µm, stromata 2.5–7 cm long………………………………………………………………………………………………………..…T. japonicum | |
| 17a. Perithecia larger………………………………………………………………………………………………………18 | |
| 17b. Perithecia smaller, 400 × 250 µm…………………………………………………………………….………T. virens | |
| 18a. Stromata 12 cm long, part spores very long, 40–65 µm long.………………………………..T. longisegmentatum | |
| 18b. Stromata shorter than 12 cm, part spores < 40 µm long……………………………………………………………19 | |
| 19a. Part spores ≤ 8 µm long……………………………………………………………………………………………….20 | |
| 19b. Part spores > 8 µm long…………………………………………………………………………………………….…22 | |
| 20a. Asci shorter than 300 µm (240–300 × 7–8 µm), perithecia relatively smaller (450–540 × 230–260 µm).…………………………………………………………………………………………………….…T. intermedium | |
| 20b. Asci longer than 300 µm, perithecia larger……………………………………………………………………..…21 | |
| 21a. Stipe slender, 0.5–1.0 mm thick, yellowish green to olivaceous, stromata shorter, 1.5–2.5 cm long, part spores, 2–5 × 1.5–2 µm…………………………………………………………………………………………………T. fractum | |
| 21b. Stipe 1–5 mm thick, dark brown, smooth or furfuraceous, stromata 5–7 cm long, part spores longer, 3–8 × 2 µm……………………………………………………………………………………………………………T. valliforme | |
| 22a. Perithecia < 550 µm long (480–540 µm)……………………………………………………….T. delicatistipitatum | |
| 22b. Perithecia > 550 µm long……………………………………………………………………………………………..23 | |
| 23a. Part spores < 15 µm long (8–11 µm)………………………………………………………………..T. miomoteanum | |
| 23b. Part spores ≥ 15 µm long…………………………………………………………………………….……………….24 | |
| 24a. Part spores < 3 µm wide……………………………………………………………………………………………..25 | |
| 24b. Part spores ≥ 3 µm wide (3.0–4.5 µm)…………………………………………………………T. inusitaticapitatum | |
| 25a. Fertile part olive-brown to olive-black, perithecia relatively larger, 650–950 × 250–420 µm, asci wider, 350–540 × 10–12 μm, part spores cylindrical or somewhat fusoid, 8–25 × 2.5–3 µm……………..……………T. capitatum | |
| 25b. Fertile part purple-brown, blacker when older, perithecia smaller, 600–750 × 200–300 µm, asci slender, 350–500 × 8–10 µm, part spores filiform, spindle-shaped, 15–20 × 2–3 µm……………………………….T. rouxii | |
| 26a. Perithecia relatively shorter, 520–740 × 300–330 μm, part spores cylindrical, 3–7 × 2–3 μm, on cicada nymphs……………………………………………………………………………………………………T. dujiaolongae | |
| 26b. Perithecia relatively longer, 900–930 × 220–250 µm, part spores fusoid, 16–18 × 3 µm, on Elaphomyces…………………………………………………………………………………………………………T. minazukiense | |
| 27a. From multiple substrate/host…………………………………………………………………………………..……28 (T. album, T. cylindrosporum, T. inflatum, T. pustulatum, T. subparadoxum) | |
| 27b. From only a type of substrate/host………………………………………………………………………………….31 | |
| 28a. Phialides cylindrical…….……………………………………………………………………………………………29 | |
| 28b. Phialides ellipsoidal to subglobos…………………………………………………………….…T. cylindrosporum | |
| 29a. Colonies white, conidia globose to ovoid (phialides 3.5–10 × 1–1.5 µm, conidia 3.5 × 1.5–2.0 µm)……………………………………………………………………………………………………….…………T. album | |
| 29b. Colonies white to pale yellow, conidia ellipsoidal, globose or broadly ellipsoidal……………………………30 | |
| 30a. Phialides 4–10 × 2–4 µm, conidia 2–3 × 1.5–2.5 µm…………………………………………………T. pustulatum | |
| 30b. Phialides 5.4–40.1 × 0.9–1.8 μm, conidia larger, 2.6–6.5 × 1–2.9 μm……………………………T. subparadoxum | |
| 31a. From substrate…………………………………………………………………………………………………….….32 | |
| 31b. On insects……………………………………………………………………………………………………………..45 | |
| 32a. Substrate is not fungus………………………………………………………………………………………………33 | |
| 32b. Substrate is fungus……………………………………………………………………………………………………43 | |
| 33a. From plant tissue……………………………………………………………………………………..………………34 (T. amazonense, T. endophyticum, T. ovalisporum, T. tropicale) | |
| 33b. From soil…………………………………………………………………………………………………………………………37 (T. geodes, T. microsporum, T. nubicola, T. pseudoalbum, T. terricola, T. tundrense, T. yunnanense) | |
| 34a. Conidia relatively more minor (globose,1.3 µm diam)……………………………………………………….T. endophyticum | |
| 34b. Conidia larger, diam > 1.3 µm……………………………………………………………………………….………35 | |
| 35a. Conidia > 4 µm long (4.5–9.0 × 2.5–3.5 µm)…………..………………………………………………T. ovalisporum | |
| 35b. Conidia < 4 µm long……………………………………………………………………………………………….…36 | |
| 36a. Phialides 4.6 ± 1.2 × 1.5 ± 0.3µm, conidia spherical, larger, 2.1–2.2 µm diam……………………….T. amazonense | |
| 36b. Phialide 4.6 × 1.5 µm, conidia spherical, relatively smaller, 1.5 ± 0.1 µm diam………………………………T. tropicale | |
| 37a. Phialides cylindrical………………………………………………………………………………………………….38 | |
| 37b. Phialides subglobose or ellipsoidal…………………………………………………………………………………41 | |
| 38a. Conidia ellipsoidal, globose or broadly ellipsoidal………………………………………………….……………39 | |
| 38b. Conidia asymmetrically flattened, with a minute apical……………………………………….…T. microsporum | |
| 39a. Colonies white………………………………………………………………………………………………………..40 | |
| 39b. Colonies white or pale yellow (Phialides 12.3–48.5 × 1.0–2.0 μm, conidia smaller, 1.8–3.4 × 1.3–1.9 μm)………………………………………………………….…………………………………………….T. pseudoalbum | |
| 40a. Phialides shorter, 5.6–12.4 × 1.4–2.4 µm, conidia 1.9–2.4 × 1.6–2.0 µm………………………………………………..T. geodes | |
| 40b. Phialides longer, 7.6–62.6 × 0.9–2.3 μm, conidia 1.2–2.4 × 0.9–1.9 μm……………………………………..T. yunnanense | |
| 41a. Conidia only one type…………………………………………………………………………..……………………42 | |
| 41b. Conidia two types (microconidia ellipsoidal or reniform, 2.3–4.2 × 1.3–2.3 µm, macroconidia: cylindrical, 10 × 2.4 µm) …………………………………………………………..……………………………………………T. tundrense | |
| 42a. Phialides relatively longer, 4.4–7.8 × 1.5–2.7 µm, conidia cylindrical, 2.6–4.1 × 0.8–1.3 µm, colonies white to pale cream……………………………………………………………………………………………………..T. nubicola | |
| 42b. Phialides shorter, 2.8–3.5 × 2.0–3.0 µm, conidia broadly oval, 2.5–3 × 2.0–2.5 µm, colonies white….T. terricola | |
| 43a. On Elaphomyces………………………………………………………………………………………T. guangdongense | |
| 43b. From Ophiocordyceps sinensis……………………………………………………………………………………………………44 | |
| 44a. Conidia reniform, 1.0–3.2 × 0.7–1.6 µm, phialides 3.4–10.6 × 1.1–3.8 µm……………………….T. reniformisporum | |
| 44b. Conidia spherical, 1.4–3.6 µm diam, phialides 7.6–19.4 × 2.9–3.6 µm.……………………………………T. sinense | |
| 45a. On mosquito larvae, conidia two types (ellipsoidal: 2–2.5 × 1.5–2 µm, subglobose to ellipsoidal, or kidney-shaped: 3.5–4 × 3–3.5 µm)…………………………………………………………………………….. T. extinguens | |
| 45b. On bdelloid rotifers, conidia only one type……………………………………………………..…………………46 | |
| 46a. Phialides thicker, 4–8 × 3–4.5 µm, conidia circular, 2.5–3.2 × 1.5–2.0 µm, colonies pure white………T. lignicola | |
| 46b. Phialides slender, 4.8–9.8 × 1.4–3.5 µm, conidia like an equilateral triangle or less ellipsoidal, 2–3 × 1.3–1.7 µm, colonies white or pale yellow……………………………………………………………………………………T. trigonosporum | |
4. Discussion
Tolypocladium is one of the most diverse fungal groups in terms of shape, substrate or host, and habitat range. Many new species have recently been added to Tolypocladium [11,12,13,14,73]. The present study described three new species (T. pseudoalbum sp. nov., T. subparadoxum sp. nov., and T. yunnanense sp. nov.) based on phylogenetic analyses and morphological characteristics. Phylogenetically, these three species fell within the Tolypocladium clade, while morphologically all three species possessed cylindrical phialides and ellipsoidal or globose conidia. It is challenging to distinguish species of Tolypocladium based only on morphological characteristics, because several species in this genus are morphologically cryptic [7,8,11]. Sexual morphological features are diverse: the ovoid perithecia may be superficial or completely immersed and part spores size varies [7,10]. However, the asexual morphological features are relatively simple.
Species of Tolypocladium play a significant role in a variety of artificial and wild ecosystems and may participate in antifungal, host–fungi, and insecticidal interactions [10,77]. Many species have been described in Tolypocladium based on host associations or morphology [11,12]. Over the past several decades, the increasing number of new fungal species being discovered globally has dramatically changed the classification of early-diverging fungi [78]. In most previous studies, the classification of Tolypocladium was developed based on morphological characteristics. However, the advent of molecular biology, which was an important scientific milestone, revolutionized the taxonomic characterization of this genus. Over the last few decades, the number of accepted species in Tolypocladium has doubled.
All 48 of the currently accepted species of Tolypocladium were included in the key developed in this study. However, because the sequence loci for many of these taxa were incomplete, only 27 species were included in the multigene phylogenetic analyses (Figure 1). The multilocus phylogenetic approach used in this study of the genus Tolypocladium shed considerable light on this influential group of fungi.
The ITS region is the most commonly used molecular marker for species delimitation in fungi. Schoch et al. proposed ITS as the standard barcode for fungi. That proposal will satisfy most fungal biologists, but not all [57,79,80]. Species-level identification of fungi has long been considered challenging. Carlson et al. reported that ITS has a low molecular variation in Trametes leading to poorly resolved phylogenies and unclear species boundaries, especially in the T. versicolor species complex [80]. The results of this study indicated that the ITS sequences did not help substantially to separate Tolypocladium species. However, the ITS sequences did help to resolve the phylogenetic relationships between Tolypocladium and related genera. The analyses of molecular phylogeny based on ITS sequences used in the current classification of the genus fungus are congruent with the higher genus clades inferred from these analyses. However, ITS sequence data are not likely to resolve species-level relationships or to delimitate closely related species and species complexes. Using the ITS phylogeny, it was still not possible to identify some species of Tolypocladium with confidence in the new classification system; the ITS region alone could not accurately identify species in Tolypocladium. For example, in the ITS phylogeny, T. varium CBS 429.94 was inseparable from T. inflatum OSC 71235 and T. inflatum NBRC 31669, while T. tundrense CBS 569.84 was inseparable from T. cylindrosporum ARSEF 2920 and T. cylindrosporum YFCC 1805001 (Figure 2). In contrast, relationships among Tolypocladium species were highly resolved in the phylogeny based on the protein-coding gene rpb1. Multilocus sequence analyses provide additional information to better characterize species boundaries [81]. Therefore, we used both morphological and multilocus phylogenetic evidence to support the novelty of the new species described in this study and to ensure accurate species identifications.
Tolypocladium extinguens was first reported from New Zealand by Samson et al. The original description was based on only a single isolate [82]. Tolypocladium extinguens is characterized by its prolonged growth in pure culture and its subglobose to ellipsoidal, sometimes kidney-shaped, conidia [82]. Our phylogenetic analysis did not support the placement of this species in Tolypocladium due to long branch attraction in the phylogenetic tree. More taxa must be added to this analysis in future to clarify the phylogenetic position of this species.
Tolypocladium species are well-known medicinal fungi that are also plant endophytes, soil inhabitants, and insect pathogens [10,12]. Because many of species of fungi are present in the soil environment at some stage of their life cycle, this substrate is preferred by researchers for the isolation of Tolypocladium. At least eight species have been reported from the soil: T. geodes, T. microsporum, T. nubicola, T. pseudoalbum sp. nov., T. subparadoxum sp. nov., T. terricola, T. tundrense, and T. yunnanense sp. nov. In Asia (China, Japan, and Thailand), Tolypocladium species are mainly known from insects [19], and few studies have focused on Tolypocladium species in the soil and in plant roots. Recently, Tolypocladium species in Chinese soils were surveyed, but no new species were identified.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/jof8111158/s1. Table S1: Pairwise genetic distance matrix of Tolypocladium species for tef-1α sequences. Table S2: Pairwise genetic distance matrix of Tolypocladium species for partial ITS sequences. Table S3: Pairwise genetic distance matrix of Tolypocladium species for partial rpb1 sequences. Table S4: Pairwise genetic distance matrix of Tolypocladium species for partial rpb2 sequences. Figure S1. Morphology of Tolypocladium subparadoxum NBRC 106958 and Tolypocladium paradoxum NBRC 100945.
Author Contributions
Conceptualization, Y.W.; methodology, Y.W.; software, Q.-Y.D.; validation, Z.-Q.W. and Y.-F.L.; formal analysis, Q.-Y.D.; investigation, Y.W.; resources, H.Y.; data curation, Z.-Q.W.; writing—original draft preparation, Q.-Y.D.; writing—review and editing, Y.W. and H.Y.; visualization, Y.W.; funding acquisition, Y.W. and H.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (31870017 and 32160005).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets presented in this study can be found in GenBank. The accession numbers can be found in the article (Table 1).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gams, W. Tolypocladium, eine Hyphomycetengattung mit geschwollenen Phialiden. Persoonia 1971, 6, 185–191. [Google Scholar]
- Barron, G.L. Fungal parasites of rotifers a new Tolypocladium with underwater conidiation. Can. J. Bot. 1980, 58, 439–442. [Google Scholar] [CrossRef]
- Barron, G.L. Two new fungal parasites of bdelloid rotifers. Can. J. Bot. 1981, 59, 1449–1455. [Google Scholar] [CrossRef]
- Barron, G.L. Structure and biology of a new Tolypocladium attacking bdelloid rotifers. Can. J. Bot. 1983, 61, 2566–2569. [Google Scholar] [CrossRef]
- Bisset, J. Notes on Tolypocladium and related genera. Can. J. Bot. 1983, 61, 1311–1329. [Google Scholar] [CrossRef]
- Dreyfuss, M.E.; Gams, W. Proposal to reject Pachybasium niveum Roster. in order to retain the name Tolypocladium inflatum W. Gams for the fungus that produces cyclosporin. Taxon 1994, 43, 660–661. [Google Scholar] [CrossRef]
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [Green Version]
- Quandt, C.A.; Kepler, R.M.; Gams, W.; Araujo, J.P.; Ban, S.; Evans, H.C.; Hughes, D.; Humber, R.; Hywel-Jones, N.; Li, Z.; et al. Phylogenetic-based nomenclatural proposals for Ophiocordycipitaceae (Hypocreales) with new combinations in Tolypocladium. IMA Fungus 2014, 5, 121–134. [Google Scholar] [CrossRef]
- Gams, W. Chaunopycnis alba, gen. et sp. nov., A soil fungus intermediete between Moniliales and Sphaeropsidales. Persoonia 1980, 11, 75–79. [Google Scholar]
- Liu, Y.F. Studies on Phylogeny and Mitogenomes of the Genus Tolypocladium; Yunnan University: Kunming, China, 2020. [Google Scholar]
- Wang, Y.; Liu, Y.F.; Tang, D.X.; Wang, Y.B.; Adams, A.E.M.; Yu, H. Tolypocladium reniformisporum sp. nov. and Tolypocladium cylindrosporum (Ophiocordycipitaceae, Hypocreales) co-occurring on Ophiocordyceps sinensis. Mycol. Prog. 2022, 21, 199–214. [Google Scholar] [CrossRef]
- Yu, F.M.; Thilini Chethana, K.W.; Wei, D.P.; Liu, J.W.; Zhao, Q.; Tang, S.M.; Li, L.; Hyde, K.D. Comprehensive Review of Tolypocladium and Description of a Novel Lineage from Southwest China. Pathogens 2021, 10, 1389. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Sugawa, G.; Takeda, K.; Degawa, Y. Tolypocladium bacillisporum (Ophiocordycipitaceae) A new parasite of Elaphomyces from Japan. Truffology 2022, 5, 15–21. [Google Scholar]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Larsson, E.; Angelini, C.; Brandrud, T.E.; Dearnaley, J.D.W.; Dima, B.; Dovana, F.; et al. Fungal Planet description sheets: 1112–1181. Persoonia 2020, 45, 251–409. [Google Scholar] [CrossRef] [PubMed]
- Montalva, C.; Silva, J.J.; Rocha, L.F.N.; Luz, C.; Humber, R.A. Characterization of Tolypocladium cylindrosporum (Hypocreales, Ophiocordycipitaceae) isolates from Brazil and their efficacy against Aedes aegypti (Diptera, Culicidae). J. Appl. Microbiol. 2019, 126, 266–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quandt, C.A.; Patterson, W.; Spatafora, J.W. Harnessing the power of phylogenomics to disentangle the directionality and signatures of interkingdom host jumping in the parasitic fungal genus Tolypocladium. Mycologia 2018, 110, 104–117. [Google Scholar] [CrossRef]
- Weiser, J.; Matha, V.; Jegorov, A. Tolypocladium terricola sp. n., a new mosquito-killing species of the genus Tolypocladium Gams (Hyphomycetes). Folia Parasit. 1991, 38, 363–369. [Google Scholar]
- Bills, G.F.; Polishook, J.D.; Goetz, M.A.; Sullivan, R.F.; White, J.F. Chaunopycnis pustulata sp. nov., a new clavicipitalean anamorph producing metabolites that modulate potassium ion channels. Mycol. Prog. 2002, 1, 3–17. [Google Scholar] [CrossRef]
- Li, C.; Hywel-Jones, N.; Cao, Y.; Nam, S.; Li, Z. Tolypocladium dujiaolongae sp. nov. and its allies. Mycotaxon 2018, 133, 229–241. [Google Scholar] [CrossRef]
- Robert, M.; Graham, M.D. Cyclosporine mechanisms of action and toxicity. Clev. Clin. J. Med. 1994, 61, 308–313. [Google Scholar]
- Aouali, N.; Eddabra, L.; Macadre, J.; Morjani, H. Immunosuppressors and reversion of multidrug-resistance. Crit. Rev. Oncol. Hematol. 2005, 56, 61–70. [Google Scholar] [CrossRef]
- Leung, P.H.; Zhang, Q.X.; Wu, J.Y. Mycelium cultivation, chemical composition and antitumour activity of a Tolypocladium sp. fungus isolated from wild Cordyceps sinensis. J. App. Microbiol. 2006, 101, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Thell, K.; Hellinger, R.; Schabbauer, G.; Gruber, C.W. Immunosuppressive peptides and their therapeutic applications. Drug Discov. Today 2014, 19, 645–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Survase, S.A.; Kagliwal, L.D.; Annapure, U.S.; Singhal, R.S. Cyclosporin A–a review on fermentative production, downstream processing and pharmacological applications. Biotechnol. Adv. 2011, 29, 418–435. [Google Scholar] [CrossRef]
- Agarwal, P.; Rupenthal, I.D. Modern approaches to the ocular delivery of cyclosporine A. Drug Discov. Today 2016, 21, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Sudoh, Y.; Tsuchiya, Y.; Okuda, T.; Matsuura, N.; Motojima, A.; Oikawa, T.; Igarashi, Y. Tolypoalbin, a new tetramic acid from Tolypocladium album TAMA 479. J. Antibiot. 2015, 68, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Bruckner, H.; Degenkolb, T. Sequences of Tolypins, Insecticidal Efrapeptin-Type Peptaibiotics from Species of the Fungal Genus Tolypocladium. Chem. Biodivers. 2020, 17, e2000276. [Google Scholar] [CrossRef]
- Khan, I.; Peng, J.; Fang, Z.; Liu, W.; Zhang, W.; Zhang, Q.; Ma, L.; Zhang, G.; Zhang, C.; Zhang, H. Cylindromicin from Arctic-Derived Fungus Tolypocladium sp. SCSIO 40433. Molecules 2021, 26, 1080. [Google Scholar] [CrossRef]
- Fukasawa, W.; Mori, N.; Iwatsuki, M.; Hokari, R.; Ishiyama, A.; Nakajima, M.; Ouchi, T.; Nonaka, K.; Kojima, H.; Matsuo, H.; et al. Tolyprolinol, a new dipeptide from Tolypocladium sp. FKI-7981. J. Antibiot. 2018, 71, 682–684. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.R.; Han, Y.F.; Liang, Z.Q. A new thermotolerant species of Taifanglania. Mycosystema 2015, 34, 345–349. [Google Scholar]
- Dong, Q.Y.; Wang, Y.; Wang, Z.Q.; Tang, D.X.; Zhao, Z.Y.; Wu, H.J.; Yu, H. Morphology and Phylogeny Reveal Five Novel Species in the Genus Cordyceps (Cordycipitaceae, Hypocreales) From Yunnan, China. Front. Microbiol. 2022, 13, 846909. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Wang, Y.B.; Yu, H.; Dai, Y.D.; Wu, C.K.; Zeng, W.B.; Yuan, F.; Liang, Z.Q. Polycephalomyces agaricus, a new hyperparasite of Ophiocordyceps sp. infecting melolonthid larvae in southwestern China. Micological Prog. 2015, 14, 70. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Bischoff, J.F.; Rehner, S.A.; Humber, R.A. Metarhizium frigidum sp. nov.: A cryptic species of M. anisopliae and a member of the M. flavoviride Complex. Mycologia 2006, 98, 737–745. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes forphylogenetics. In PCR Protocols, a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0a165; Sinauer Associates: Sunderland, MA, USA, 2019. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. Partitionfinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Kepler, R.; Ban, S.; Nakagir, A.; Bischoff, J.; Hywel-Jones, N.; Owensby, C.A.; Spatafora, J.W. The phylogenetic placement of hypocrealean insect pathogens in the genus Polycephalomyces: An application of One Fungus One Name. Fungal. Fungal Biol.-UK 2013, 117, 611–622. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML Web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree. Tree Figure Drawing Tool Version 1.3.1; University of Edinburgh: Edinburgh, UK, 2006. [Google Scholar]
- Sung, G.H.; Spatafora, J.W.; Zare, R.; Gams, W. A revision of Verticillium sect. Prostrata. II. Phylogenetic analyses of SSU and LSU nuclear rDNA sequences from anamorphs and teleomorphs of the Clavicipitaceae. Nova Hedwigia 2001, 72, 311–328. [Google Scholar]
- Spatafora, J.W.; Sung, G.-H.; Sung, G.H.; Hywel-Jones, N.; White, J.F. Phylogenetic evidence for an animal pathogen origin of ergot and the grass endophytes. Mol. Ecol. 2007, 16, 1701–1711. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, Z.; Guo, Q.; Fokkens, L.; Miskei, M.; Pócsi, I.; Zhang, W.; Chen, M.; Wang, L.; Sun, Y.; et al. Insights into Adaptations to a NearObligate Nematode Endoparasitic Lifestyle from the Finished Genome of Drechmeria coniospora. Sci. Rep. 2016, 6, 23122. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Hou, S.Y.; Zhang, M.Y.; Zhang, T.Y.; Zhang, Y.X. Drechmeria panacis sp. nov., an endophyte isolated from Panax notoginseng. Int. J. Syst. Evol. Micr. 2018, 68, 3255–3259. [Google Scholar] [CrossRef]
- Chaverri, P.; Samuels, G.J.; Hodge, K.T. The genus Podocrella and its nematode-killing anamorph Harposporium. Mycologia 2005, 97, 433–443. [Google Scholar] [CrossRef]
- Simmons, D.R.; Kepler, R.M.; Rehner, S.A.; Groden, E. Phylogeny of Hirsutella species (Ophiocordycipitaceae) from the USA: Remedying the paucity of Hirsutella sequence data. IMA Fungus 2015, 6, 345–356. [Google Scholar] [CrossRef]
- Lai, Y.; Liu, K.; Zhang, X.; Zhang, X.; Li, K.; Wang, N.; Shu, C.; Wu, Y.; Wang, C.; Bushley, K.E.; et al. Comparative genomics and transcriptomics analyses reveal divergent lifestyle features of nematode endoparasitic fungus Hirsutella minnesotensis. Genome Biol. Evol. 2014, 6, 3077–3093. [Google Scholar] [CrossRef]
- Agrawal, Y.; Khatri, I.; Subramanian, S.; Shenoy, B.D. Genome sequence, comparative analysis, and evolutionary insights into chitinases of entomopathogenic fungus Hirsutella thompsonii. Genome Biol. Evol. 2015, 7, 916–930. [Google Scholar] [CrossRef]
- Sanjuan, T.I.; Franco-Molano, A.E.; Kepler, R.M.; Spatafora, J.W.; Tabima, J.; Vasco-Palacios, A.M.; Restrepo, S. Five new species of entomopathogenic fungi from the Amazon and evolution of neotropical Ophiocordyceps. Fungal Bio. 2015, 119, 901–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ban, S.; Sakane, T.; Nakagiri, A. Three new species of Ophiocordyceps and overview of anamorph types in the genus and the family Ophiocordyceptaceae. Mycol. Prog. 2015, 14, 1017. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding, C.; Fungal Barcoding Consortium Author, L. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, B.H.; Mitchell, J.; de Beer, Z.W.; Caroe, C.; Gilbert, M.T.; Eilenberg, J.; Poulsen, M.; de Fine Licht, H.H. Draft genome of the fungus-growing termite pathogenic fungus Ophiocordyceps bispora (Ophiocordycipitaceae, Hypocreales, Ascomycota). Data Brief 2017, 11, 537–542. [Google Scholar] [CrossRef]
- Luangsa-Ard, J.; Tasanathai, K.; Thanakitpipattana, D.; Khonsanit, A.; Stadler, M. Novel and interesting Ophiocordyceps spp. (Ophiocordycipitaceae, Hypocreales) with superficial perithecia from Thailand. Stud. Mycol. 2018, 89, 125–142. [Google Scholar] [CrossRef]
- Wang, L.; Li, H.-H.; Chen, Y.-Q.; Zhang, W.-M.; Qu, L.-H. Polycephalomyces lianzhouensis sp. nov., a new species, co-occurs with Ophiocordyceps crinalis. Mycol. Prog. 2014, 13, 996. [Google Scholar] [CrossRef]
- Chen, Z.H.; Dai, Y.D.; Yu, H.; Yang, K.; Yang, Z.L.; Yuan, F.; Zeng, W.B. Systematic analyses of Ophiocordyceps lanpingensis sp. nov., a new species of Ophiocordyceps in China. Microbiol. Res. 2013, 168, 525–532. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.; Crane, C.; Barrett, S.; Cano-Lira, J.F.; Le Roux, J.J.; Thangavel, R.; Guarro, J.; et al. Fungal Planet description sheets: 469–557. Persoonia 2016, 37, 218–403. [Google Scholar] [CrossRef]
- Johnson, D.; Sung, G.; Hywel-Jones, N.L.; Luangsa-Ard, J.J.; Bischoff, J.F.; Kepler, R.M.; Spatafora, J.W. Systematics and evolution of the genus Torrubiella (Hypocreales, Ascomycota). Mycol. Res. 2009, 113, 279–289. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Y.J.; Liu, X.Z.; Zhang, H.; Liu, D.S. On the reliability of DNA sequences of Ophiocordyceps sinensis in public databases. J. Ind. Microbiol. Biotechnol. 2013, 40, 365–378. [Google Scholar] [CrossRef]
- Wang, Y.B.; Nguyen, T.T.; Dai, Y.D.; Yu, H.; Zeng, W.B.; Wu, C.K. Molecular phylogeny and morphology of Ophiocordyceps unituberculata sp. nov. (Ophiocordycipitaceae), a pathogen of caterpillars (Noctuidae, Lepidoptera) from Yunnan, China. Mycol. Prog. 2018, 17, 745–753. [Google Scholar] [CrossRef]
- Wen, T.C.; Zhu, R.C.; Kang, J.C.; Huang, M.H.; Tan, D.B.; Ariyawansha, H.; Hyde, K.D.; Liu, H.A.O. Ophiocordyceps xuefengensis sp. nov. from larvae of Phassus nodus (Hepialidae) in Hunan Province, southern China. Phytotaxa 2013, 123, 41–50. [Google Scholar] [CrossRef]
- Bischoff, J.F.; Sullivan, R.F.; Struwe, L.; Hywel-Jones, N.L.; White, J.F.J. Resurrection of Blistum tomentosum and its exclusion from Polycephalomyces (Hyphomycetes, Deuteromycota) based on 28S rDNA sequence data. Mycotaxon 2003, 86, 433–444. [Google Scholar]
- Wang, Y.B. Studies on Phylogeny of Polycephalomycetaceae fam. nov., with Microbial Diversities of Polycephalomyces multiramosus and Its Host. Ph.D. Thesis, Yunnan University, Kunming, China, 2015. [Google Scholar]
- Wang, W.J.; Wang, X.L.; Li, Y.; Xiao, S.R.; Kepler, R.M.; Yao, Y.J. Molecular and morphological studies of Paecilomyces sinensis reveal a new clade in clavicipitaceous fungi and its new systematic position. Syst. Biodivers. 2012, 10, 221–232. [Google Scholar] [CrossRef]
- Perdomo, H.; Cano, J.; Gene, J.; Garcia, D.; Hernandez, M.; Guarro, J. Polyphasic analysis of Purpureocillium lilacinum isolates from different origins and proposal of the new species Purpureocillium lavendulum. Mycologia 2013, 105, 151–161. [Google Scholar] [CrossRef]
- Luangsa-ard, J.J.; Hywel-Jones, N.L.; Samson, R.A. The polyphyletic nature of Paecilomyces sensu lato based on 18S-generated rDNA phylogeny. Mycologia 2004, 96, 773–780. [Google Scholar] [CrossRef]
- Gazis, R.; Skaltsas, D.; Chaverri, P. Novel endophytic lineages of Tolypocladium provide new insights into the ecology and evolution of Cordyceps-like fungi. Mycologia 2014, 106, 1090–1105. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Dissanayake, L.S.; Li, Q.R.; Dai, D.Q.; Xiao, Y.P.; Wen, T.C.; Karunarathna, S.C.; Wu, X.X.; Zhang, H.; Tibpromm, S.; et al. Yunnan–Guizhou Plateau: A mycological hotspot. Phytotaxa 2021, 523, 001–031. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Houbraken, J.; Boekhout, T.; Crous, P.W.; Robert, V.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.; Barber, P.A.; Alvarado, P.; Barnes, C.W.; Buchanan, P.K.; Heykoop, M.; Moreno, G.; et al. Fungal Planet description sheets: 558–624. Persoonia 2017, 38, 240–384. [Google Scholar] [CrossRef]
- Kobayasi, Y.; Shimizu, D. Monographic studies of cordyceps 2, group parasitic on cicadae. Bull. Natl. Sci. Mus. 1963, 6, 286–314. [Google Scholar]
- Wang, J.C.; Zhang, Z.Z.; Li, Z.L.; Wang, Y. Research Progress of Tolypocladium in Ophiocordycipitaceae. J. Fungal Res. 2019, 8, 1–10. [Google Scholar]
- Voigt, K.; James, T.Y.; Kirk, P.M.; Santiago, A.; Waldman, B.; Griffith, G.W.; Fu, M.; Radek, R.; Strassert, J.F.H.; Wurzbacher, C.; et al. Early-diverging fungal phyla: Taxonomy, species concept, ecology, distribution, anthropogenic impact, and novel phylogenetic proposals. Fungal Divers. 2021, 109, 59–98. [Google Scholar] [CrossRef] [PubMed]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal Identification Using Molecular Tools: A Primer for the Natural Products Research Community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Carlson, A.; Justo, A.; Hibbett, D.S. Species delimitation in Trametes: A comparison of ITS, RPB1, RPB2 and TEF1 gene phylogenies. Mycologia 2014, 106, 735–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, L.; Alves, A.; Alves, R. Evaluating multi-locus phylogenies for species boundaries determination in the genus Diaporthe. PeerJ 2017, 5, e3120. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.A.; Soares, G.G. Entomopathogenic Species of the Hyphomycete Genus Tolypocladium. J. Invertebr. Pathol. 1984, 43, 133–139. [Google Scholar] [CrossRef]
Figure 1.
Maximum-likelihood tree illustrating the phylogeny of Tolypocladium based on the combined dataset of nrSSU, nrLSU, tef-1α, rpb1 and rpb2 sequences. Polycephalomyces formosus ARSEF 1424 and Polycephalomyces sinensis CN 80-2 were used as outgroups. The maximum-likelihood bootstrap values (≥50) and Bayesian posterior probability values (≥0.50) are indicated above the branches. Isolates in bold type are those analyzed in this study.
Figure 1.
Maximum-likelihood tree illustrating the phylogeny of Tolypocladium based on the combined dataset of nrSSU, nrLSU, tef-1α, rpb1 and rpb2 sequences. Polycephalomyces formosus ARSEF 1424 and Polycephalomyces sinensis CN 80-2 were used as outgroups. The maximum-likelihood bootstrap values (≥50) and Bayesian posterior probability values (≥0.50) are indicated above the branches. Isolates in bold type are those analyzed in this study.

Figure 2.
Maximum parsimony, Bayesian analysis, and RAxML tree illustrating the phylogeny of Tolypocladium derived from ITS sequences. Statistical support values (MP bootstrap/Bayesian posterior probability/ML bootstrap ≥ 70%) are shown at the nodes. The indistinguishable species are in red and the isolates analyzed in this study are in bold.
Figure 2.
Maximum parsimony, Bayesian analysis, and RAxML tree illustrating the phylogeny of Tolypocladium derived from ITS sequences. Statistical support values (MP bootstrap/Bayesian posterior probability/ML bootstrap ≥ 70%) are shown at the nodes. The indistinguishable species are in red and the isolates analyzed in this study are in bold.

Figure 3.
Morphology of Tolypocladium pseudoalbum (YFCC 875, ex-type living culture). (A,B) Culture characteristics on PDA medium incubated at 22 °C for 14 days; (C–I) phialides; (J) conidia; (K) chlamydospore. Scale bars: (A,B) = 10 mm; (C–H) = 20 μm; (I–K) = 10 μm.
Figure 3.
Morphology of Tolypocladium pseudoalbum (YFCC 875, ex-type living culture). (A,B) Culture characteristics on PDA medium incubated at 22 °C for 14 days; (C–I) phialides; (J) conidia; (K) chlamydospore. Scale bars: (A,B) = 10 mm; (C–H) = 20 μm; (I–K) = 10 μm.

Figure 4.
Morphology of Tolypocladium subparadoxum (YFCC 879, ex-type living culture). (A,B) Culture characteristics on PDA medium incubated at 22 °C for 21 days; (C–F) phialides and conidia. Scale bars: (A,B) = 10 mm; (C–E) = 50 μm; (F) = 20 μm.
Figure 4.
Morphology of Tolypocladium subparadoxum (YFCC 879, ex-type living culture). (A,B) Culture characteristics on PDA medium incubated at 22 °C for 21 days; (C–F) phialides and conidia. Scale bars: (A,B) = 10 mm; (C–E) = 50 μm; (F) = 20 μm.

Figure 5.
Morphology of Tolypocladium yunnanense (YFCC 877, ex-type living culture). (A,B) Culture characteristics on PDA medium incubated at 22 °C for 14 days; (C–J) phialides and conidia; (K) chlamydospore. Scale bars: A–B = 10 mm; C, H = 10 μm; D–G, I–K = 20 μm.
Figure 5.
Morphology of Tolypocladium yunnanense (YFCC 877, ex-type living culture). (A,B) Culture characteristics on PDA medium incubated at 22 °C for 14 days; (C–J) phialides and conidia; (K) chlamydospore. Scale bars: A–B = 10 mm; C, H = 10 μm; D–G, I–K = 20 μm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Specimen information and GenBank accession numbers of sequences used in this study.
| Taxon | Voucher Information | GenBank Accession Number | Reference | ||||
|---|---|---|---|---|---|---|---|
| nrSSU | nrLSU | tef-1α | rpb1 | rpb2 | |||
| Drechmeria balanoides | CBS 250.82T | AF339588 | AF339539 | DQ522342 | DQ522388 | DQ522442 | [47,48] |
| Drechmeria campanulata | IMI 356051T | AF339592 | AF339543 | - | - | - | [47] |
| Drechmeria coniospora | ARSEF 6962 | - | LAYC01000003 | LAYC01000001 | LAYC01000003 | LAYC01000002 | [49] |
| Drechmeria gunnii | OSC 76404 | AF339572 | AF339522 | AY489616 | AY489650 | DQ522426 | [7,47] |
| Drechmeria panacis | CBS 142798T | MF588890 | MF588897 | MF614144 | - | - | [50] |
| Drechmeria sinensis | CBS 567.95 | AF339594 | AF339545 | DQ522343 | DQ522389 | DQ522443 | [47,48] |
| Drechmeria sphaerospora | CBS 522.80T | AF339590 | AF339541 | - | - | - | [47] |
| Drechmeria zeospora | CBS 335.80T | AF339589 | AF339540 | EF469062 | EF469091 | EF469109 | [7,47] |
| Harposporium anguillulae | ARSEF 5407 | - | AY636080 | - | - | - | [51] |
| Harposporium anguillulae | ARSEF 5593 | - | AY636081 | - | - | - | [51] |
| Harposporium harposporiferum | ARSEF 5472T | AF339569 | AF339519 | DQ118747 | DQ127238 | - | [47,48] |
| Harposporium helicoides | ARSEF 5354 | AF339577 | AF339527 | - | - | - | [47] |
| Hirsutella citriformis | ARSEF 1446 | KM652065 | KM652106 | KM651990 | KM652031 | - | [52] |
| Hirsutella cryptosclerotium | ARSEF 4517T | KM652066 | KM652109 | KM651992 | KM652032 | - | [52] |
| Hirsutella fusiformis | ARSEF 5474 | KM652067 | KM652110 | KM651993 | KM652033 | - | [52] |
| Hirsutella guyana | ARSEF 878 | KM652068 | KM652111 | KM651994 | KM652035 | - | [52] |
| Hirsutella illustris | ARSEF 5539 | KM652069 | KM652112 | KM651996 | KM652037 | - | [52] |
| Hirsutella lecaniicola | ARSEF 8888 | KM652071 | KM652114 | KM651998 | KM652038 | - | [52] |
| Hirsutella minnesotensis | 3608 | JPUM01000376 | JPUM01000376 | JPUM01000211 | JPUM01000139 | JPUM01000138 | [53] |
| Hirsutella necatrix | ARSEF 5549 | KM652073 | KM652116 | KM651999 | KM652039 | - | [52] |
| Hirsutella nodulosa | ARSEF 5473 | KM652074 | KM652117 | KM652000 | KM652040 | - | [52] |
| Hirsutella radiate | ARSEF 1369 | KM652076 | KM652119 | KM652002 | KM652042 | - | [52] |
| Hirsutella rhossiliensis | ARSEF 3747 | KM652080 | KM652123 | KM652006 | KM652045 | - | [52] |
| Hirsutella satumaensis | ARSEF 996 | KM652082 | KM652125 | KM652008 | KM652047 | - | [52] |
| Hirsutella strigose | ARSEF 2197 | KM652085 | KM652129 | KM652012 | KM652050 | - | [52] |
| Hirsutella subulata | ARSEF 2227 | KM652086 | KM652130 | KM652013 | KM652051 | - | [52] |
| Hirsutella thompsonii | MTCC 3556 | APKB01000383 | APKB01000383 | APKB01000061 | APKB01000125 | APKB01000164 | [54] |
| Hirsutella versicolor | ARSEF 1037 | KM652102 | KM652150 | KM652029 | KM652063 | - | [52] |
| Ophiocordyceps acicularis | OSC 110987 | EF468950 | EF468805 | EF468744 | EF468852 | - | [7] |
| Ophiocordyceps acicularis | OSC 110988 | EF468951 | EF468804 | EF468745 | EF468853 | - | [7] |
| Ophiocordyceps agriotidis | ARSEF 5692 | DQ522540 | DQ518754 | DQ522322 | DQ522368 | DQ522418 | [48] |
| Ophiocordyceps amazonica | HUA 186143T | KJ917562 | KJ917571 | KM411989 | KP212902 | KM411982 | [55] |
| Ophiocordyceps appendiculata | NBRC 106960 | JN941728 | JN941413 | AB968577 | JN992462 | AB968539 | [56,57] |
| Ophiocordyceps arborescens | NBRC 105891T | AB968386 | AB968414 | AB968572 | - | AB968534 | [56] |
| Ophiocordyceps bispora | ERS1123077 | FKNF01000183 | FKNF01000183 | FKNF01000002 | FKNF01000038 | FKNF01000031 | [58] |
| Ophiocordyceps blattarioides | HUA 186108T | KJ917558 | KJ917569 | - | KP212912 | KM411984 | [55] |
| Ophiocordyceps brunneanigra | TBRC 8093T | - | MF614654 | MF614638 | MF614668 | MF614681 | [59] |
| Ophiocordyceps brunneipunctata | OSC 128576 | DQ522542 | DQ518756 | DQ522324 | DQ522369 | DQ522420 | [48] |
| Ophiocordyceps cf. acicularis | OSC 128580 | DQ522543 | DQ518757 | DQ522326 | DQ522371 | DQ522423 | [48] |
| Ophiocordyceps crinalis | GDGM 17327 | KF226253 | KF226254 | KF226256 | KF226255 | - | [60] |
| Ophiocordyceps entomorrhiza | KEW 53484 | EF468954 | EF468809 | EF468749 | EF468857 | EF468911 | [7] |
| Ophiocordyceps geometridicola | TBRC 8095T | - | MF614648 | MF614632 | MF614663 | MF614679 | [59] |
| Ophiocordyceps gracilis | EFCC 8572 | EF468956 | EF468811 | EF468751 | EF468859 | EF468912 | [7] |
| Ophiocordyceps heteropoda | NBRC 100644 | JN941718 | JN941423 | AB968596 | JN992452 | AB968557 | [56,57] |
| Ophiocordyceps kniphofioides | HUA 186148 | KC610790 | KF658679 | KC610739 | KF658667 | KC610717 | [55] |
| Ophiocordyceps lanpingensis | YHOS0705 | KC417458 | KC417460 | KC417462 | KC417464 | KC456333 | [61] |
| Ophiocordyceps macroacicularis | NBRC 100685T | AB968388 | AB968416 | AB968574 | - | AB968536 | [56] |
| Ophiocordyceps multiperitheciata | BCC 69008T | - | MF614657 | MF614641 | - | MF614682 | [59] |
| Ophiocordyceps nigrella | EFCC 9247 | EF468963 | EF468818 | EF468758 | EF468866 | EF468920 | [7] |
| Ophiocordyceps nooreniae | BRIP 55363T | KX673811 | KX673810 | KX673812 | - | KX673809 | [62] |
| Ophiocordyceps pseudoacicularis | TBRC 8102T | - | MF614646 | MF614630 | MF614661 | MF614677 | [59] |
| Ophiocordyceps pruinosa | NHJ 12994 | EU369106 | EU369041 | EU369024 | EU369063 | EU369084 | [63] |
| Ophiocordyceps ravenelii | OSC 110995 | DQ522550 | DQ518764 | DQ522334 | DQ522379 | DQ522430 | [48] |
| Ophiocordyceps rhizoidea | NHJ 12522 | EF468970 | EF468825 | EF468764 | EF468873 | EF468923 | [7] |
| Ophiocordyceps rubiginosiperitheciata | NBRC 106966 | JN941704 | JN941437 | AB968582 | JN992438 | AB968544 | [56,57] |
| Ophiocordyceps sinensis | EFCC 7287 | EF468971 | EF468827 | EF468767 | EF468874 | EF468924 | [7] |
| Ophiocordyceps sinensis | YN07-8 | JX968027 | JX968032 | JX968017 | JX968007 | JX968012 | [64] |
| Ophiocordyceps sinensis | YHH 1805 | MK984568 | MK984580 | MK984572 | MK984587 | MK984576 | [11] |
| Ophiocordyceps spataforae | BCC 86480T | - | MG831747 | MG831746 | MG831748 | MG831749 | [59] |
| Ophiocordyceps stylophora | OSC 110999 | EF468982 | EF468837 | EF468777 | EF468882 | EF468931 | [7] |
| Ophiocordyceps unilateralis | OSC 128574 | DQ522554 | DQ518768 | DQ522339 | DQ522385 | DQ522436 | [48] |
| Ophiocordyceps unituberculata | YFCC HU1301T | KY923214 | KY923212 | KY923216 | KY923218 | KY923220 | [65] |
| Ophiocordyceps variabilis | ARSEF 5365 | DQ522555 | DQ518769 | DQ522340 | DQ522386 | DQ522437 | [48] |
| Ophiocordyceps xuefengensis | GZUH2012HN14T | KC631789 | - | KC631793 | KC631798 | - | [66] |
| Polycephalomyces formosus | ARSEF 1424 | KF049615 | AY259544 | DQ118754 | DQ127245 | KF049671 | [43,51,67,68] |
| Polycephalomyces sinensis | CN 80-2 | HQ832887 | HQ832886 | HQ832890 | HQ832888 | HQ832889 | [69] |
| Purpureocillium atypicolum | CBS 744.73 | EF468987 | EF468841 | EF468786 | EF468892 | - | [7] |
| Purpureocillium atypicolum | OSC 151901 | KJ878914 | KJ878880 | KJ878961 | KJ878994 | - | [8] |
| Purpureocillium lavendulum | CBS 128677T | - | FR775489 | FR775516 | FR775512 | FR775538 | [70] |
| Purpureocillium lilacinum | CBS 284.36T | AY526475 | FR775484 | EF468792 | EF468898 | EF468941 | [7,70,71] |
| Purpureocillium lilacinum | NHJ 3497 | EU369096 | EU369033 | EU369014 | EU369053 | EU369074 | [63] |
| Purpureocillium takamizusanense | NHJ 3582 | EU369097 | EU369034 | EU369015 | - | - | [63] |
| Tolypocladium amazonense | CBS 136895T | KF747314 | KF747134 | KF747099 | KF747214 | - | [72] |
| Tolypocladium bacillisporum | C23 | LC684522 | LC684522 | LC684525 | [13] | ||
| Tolypocladium capitatum | NBRC 100997 | JN941740 | JN941401 | AB968597 | JN992474 | AB968558 | [56,57] |
| Tolypocladium capitatum | NBRC 106325 | JN941739 | JN941402 | AB968598 | JN992473 | AB968559 | [56,57] |
| Tolypocladium capitatum | YFCC 881 | OP207711 | OP207731 | OP223145 | OP223123 | OP223133 | Present study |
| Tolypocladium cucullae | GZU A-77 | MW798785 | MW798787 | - | - | - | [73] |
| Tolypocladium cucullae | HKAS 55588 | MW798784 | MW798786 | - | - | - | [73] |
| Tolypocladium cylindrosporum | ARSEF 2920T | - | MH871712 | MG228390 | MG228384 | MG228387 | [15,74] |
| Tolypocladium cylindrosporum | YFCC 1805001 | MK984565 | MK984577 | MK984569 | MK984584 | MK984573 | [11] |
| Tolypocladium endophyticum | MS337 | KF747315 | KF747136 | KF747101 | KF747215 | - | [72] |
| Tolypocladium endophyticum | MX486 | KF747321 | KF747152 | KF747116 | KF747232 | - | [72] |
| Tolypocladium flavonigrum | BCC 66576 | - | MN337287 | MN338495 | - | - | [14] |
| Tolypocladium flavonigrum | BCC 66580 | - | MN337289 | MN338497 | MN338494 | - | [14] |
| Tolypocladium fractum | OSC 110990 | DQ522545 | DQ518759 | DQ522328 | DQ522373 | DQ522425 | [48] |
| Tolypocladium fumosum | CBS H-22968T | - | KU985053 | - | - | - | [75] |
| Tolypocladium geodes | CBS 126054 | - | MH875520 | - | - | - | [74] |
| Tolypocladium inegoense | SU-15 | - | DQ118741 | DQ118752 | DQ127243 | - | [51] |
| Tolypocladium inflatum | OSC 71235 | EF469124 | EF469077 | EF469061 | EF469090 | EF469108 | [7] |
| Tolypocladium inusitaticapitatum | HKAS 112152 | MW537733 | MW537718 | MW507527 | - | MW507529 | [12] |
| Tolypocladium inusitaticapitatum | HKAS 112153 | MW537734 | MW537719 | MW507528 | - | MW507530 | [12] |
| Tolypocladium japonicum | NBRC 9647 | OP207712 | OP207732 | OP223146 | OP223124 | OP223134 | Present study |
| Tolypocladium jezoense | NBRC 106328 | OP207713 | OP207733 | OP223147 | OP223125 | OP223135 | Present study |
| Tolypocladium longisegmentum | OSC 110992 | - | EF468816 | - | EF468864 | EF468919 | [7] |
| Tolypocladium nubicola | CBS 568.84T | - | MH873478 | - | - | - | [74] |
| Tolypocladium ophioglossoides | CBS 100239 | KJ878910 | KJ878874 | KJ878958 | KJ878990 | KJ878944 | [8] |
| Tolypocladium ophioglossoides | NBRC 100998 | JN941735 | JN941406 | AB968602 | JN992469 | AB968563 | [56,57] |
| Tolypocladium ophioglossoides | NBRC 106330 | JN941734 | JN941407 | AB968603 | JN992468 | AB968564 | [56,57] |
| Tolypocladium paradoxum | NBRC 100945 | JN941731 | JN941410 | AB968599 | JN992465 | AB968560 | [56,57] |
| Tolypocladium paradoxum | YFCC 882 | OP207714 | OP207734 | OP223148 | OP223126 | OP223136 | Present study |
| Tolypocladium pseudoalbum | YFCC 875T | OP207717 | OP207737 | OP223151 | OP223129 | OP223139 | Present study |
| Tolypocladium pseudoalbum | YFCC 876 | OP207718 | OP207738 | OP223152 | OP223130 | OP223140 | Present study |
| Tolypocladium pustulatum | MRL GB6597 | - | AF389190 | - | - | - | [18] |
| Tolypocladium pustulatum | MRL MF5368LR | - | AF373282 | - | - | - | [18] |
| Tolypocladium reniformisporum | YFCC 1805002T | MK984566 | MK984578 | MK984570 | MK984585 | MK984574 | [11] |
| Tolypocladium sp. | YFCC 201803 | MK984567 | MK984579 | MK984571 | MK984586 | MK984575 | [11] |
| Tolypocladium subparadoxum | NBRC 106958 | OP207715 | OP207735 | OP223149 | OP223127 | OP223137 | Present study |
| Tolypocladium subparadoxum | YFCC 879T | OP207716 | OP207736 | OP223150 | OP223128 | OP223138 | Present study |
| Tolypocladium tropicale | CBS 136897T | - | KF747125 | KF747090 | KF747204 | - | [72] |
| Tolypocladium tropicale | MX338 | KF747318 | KF747149 | KF747113 | KF747229 | - | [72] |
| Tolypocladium tundrense | CBS 569.84T | - | MH873479 | - | - | - | [74] |
| Tolypocladium yunnanense | YFCC 877T | OP207719 | OP207739 | OP223153 | OP223131 | - | Present study |
| Tolypocladium yunnanense | YFCC 878 | OP207720 | OP207740 | OP223154 | OP223132 | - | Present study |
Boldface: data generated in this study. T ex-type material.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dong, Q.-Y.; Wang, Y.; Wang, Z.-Q.; Liu, Y.-F.; Yu, H. Phylogeny and Systematics of the Genus Tolypocladium (Ophiocordycipitaceae, Hypocreales). J. Fungi 2022, 8, 1158. https://doi.org/10.3390/jof8111158
AMA Style
Dong Q-Y, Wang Y, Wang Z-Q, Liu Y-F, Yu H. Phylogeny and Systematics of the Genus Tolypocladium (Ophiocordycipitaceae, Hypocreales). Journal of Fungi. 2022; 8(11):1158. https://doi.org/10.3390/jof8111158
Chicago/Turabian StyleDong, Quan-Ying, Yao Wang, Zhi-Qin Wang, Yan-Fang Liu, and Hong Yu. 2022. "Phylogeny and Systematics of the Genus Tolypocladium (Ophiocordycipitaceae, Hypocreales)" Journal of Fungi 8, no. 11: 1158. https://doi.org/10.3390/jof8111158
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.