
Taxonomy and Phylogeny of Fungi Associated with Mangifera indica from Yunnan, China

 ,
,  , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling, Isolation and Cultivation of Fungi
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Phylogenetic Analyses
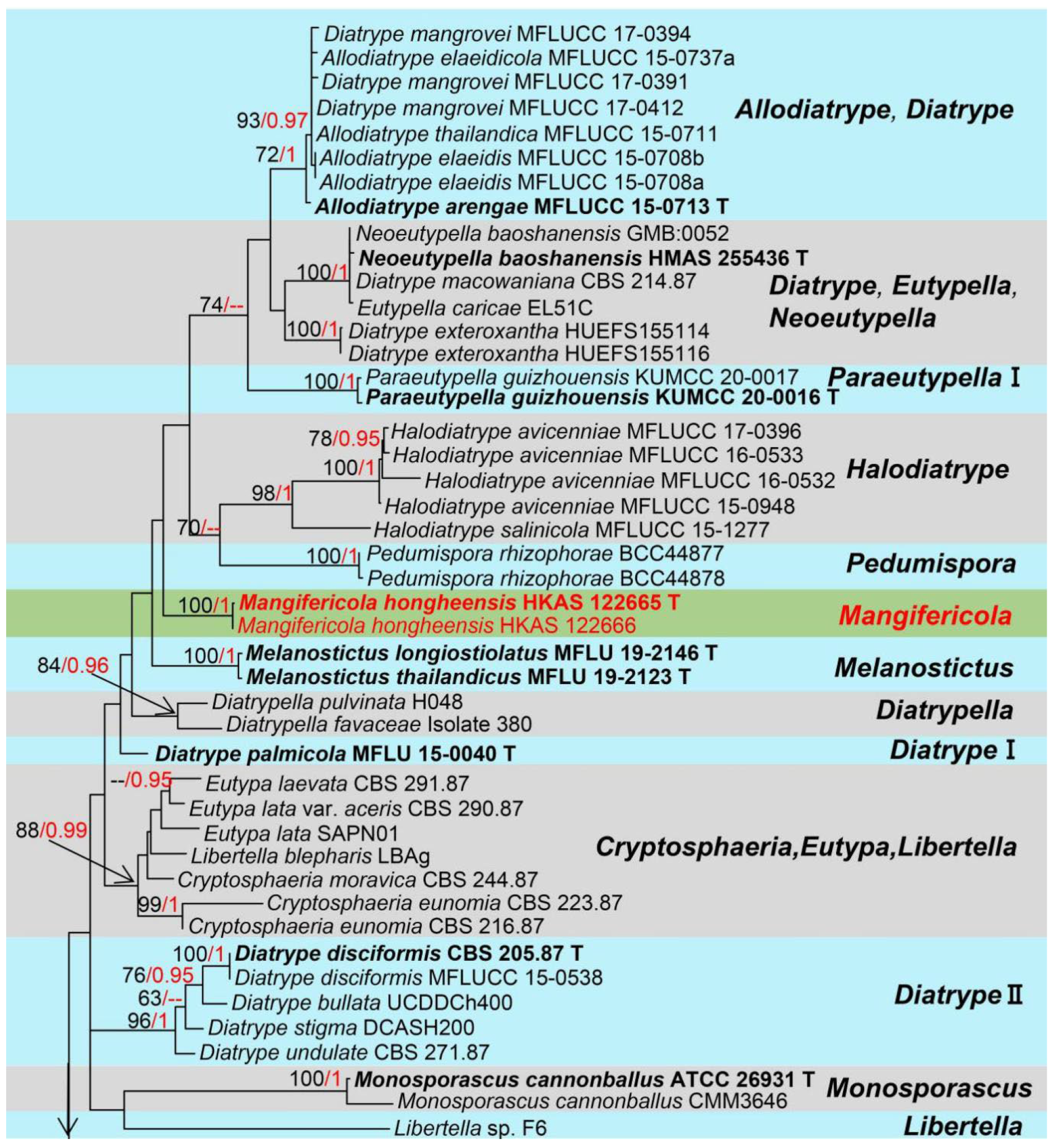
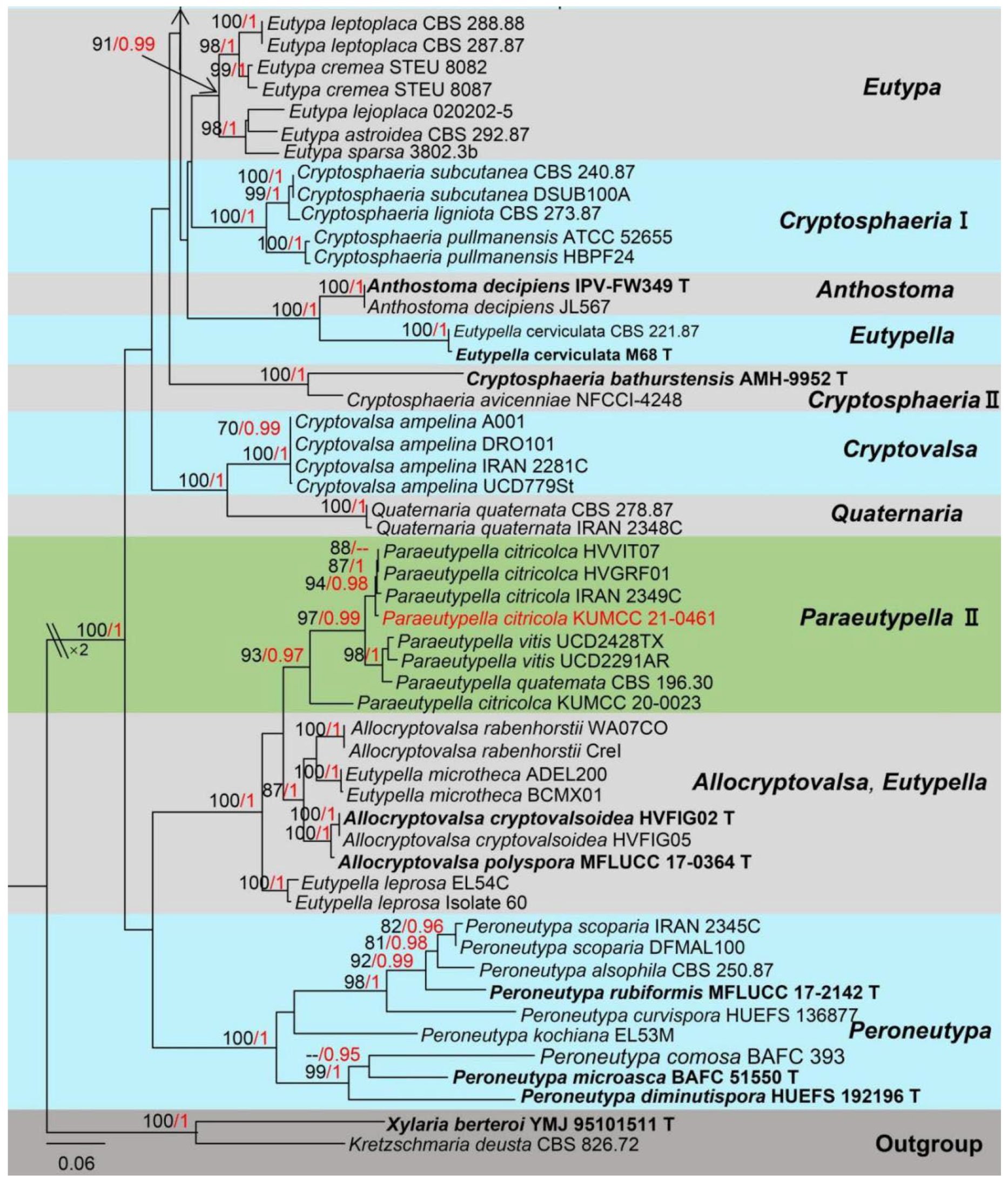
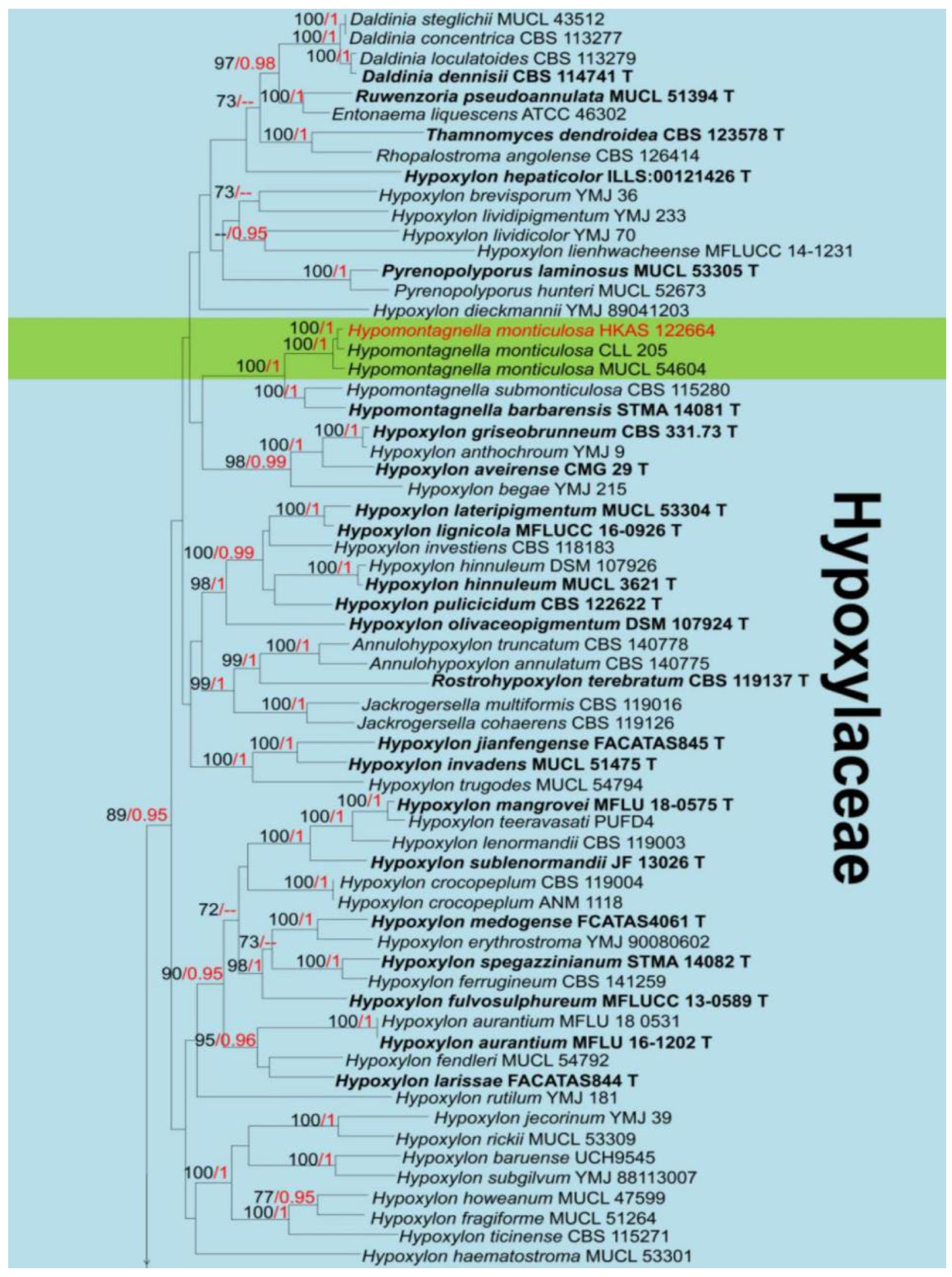
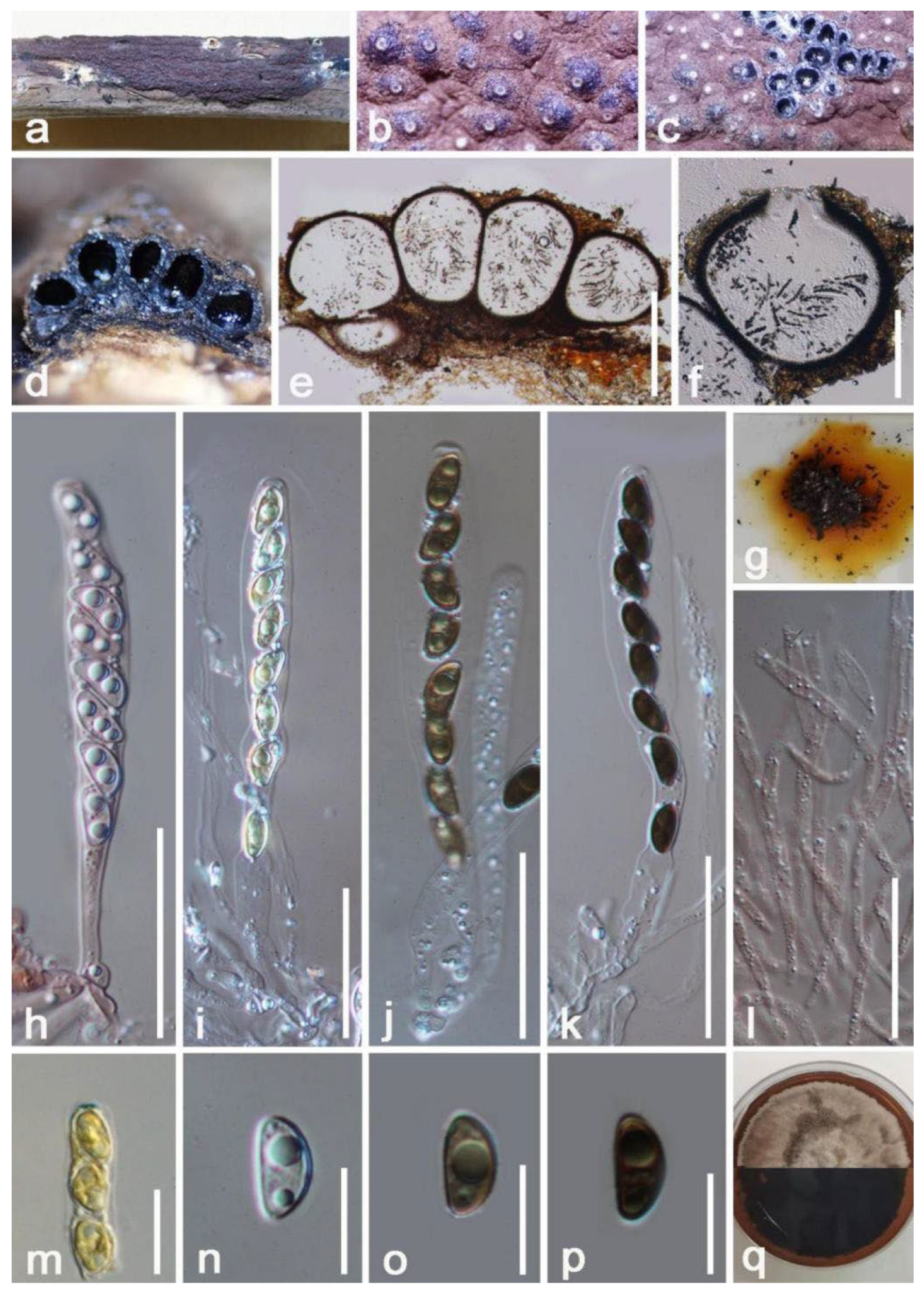
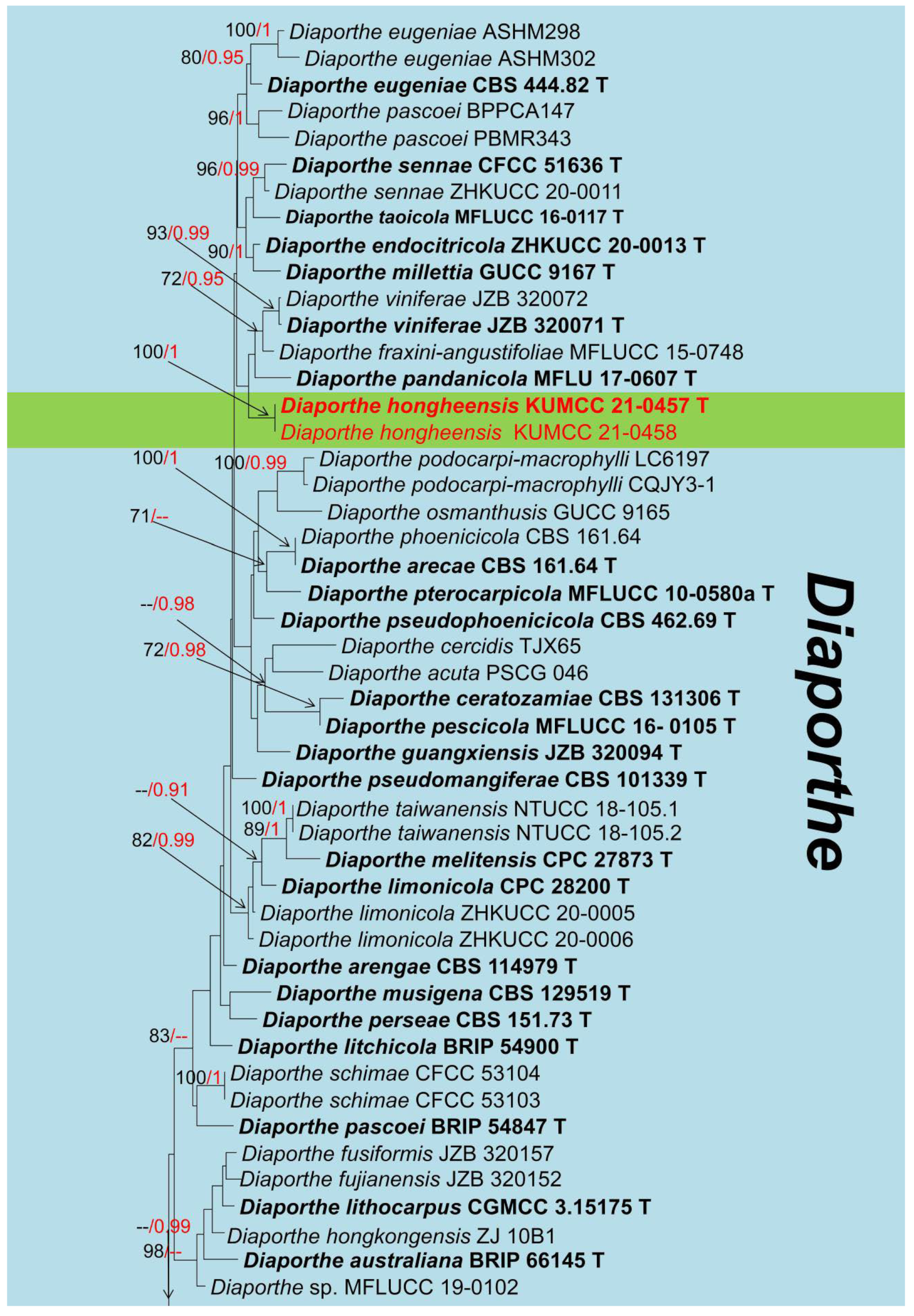
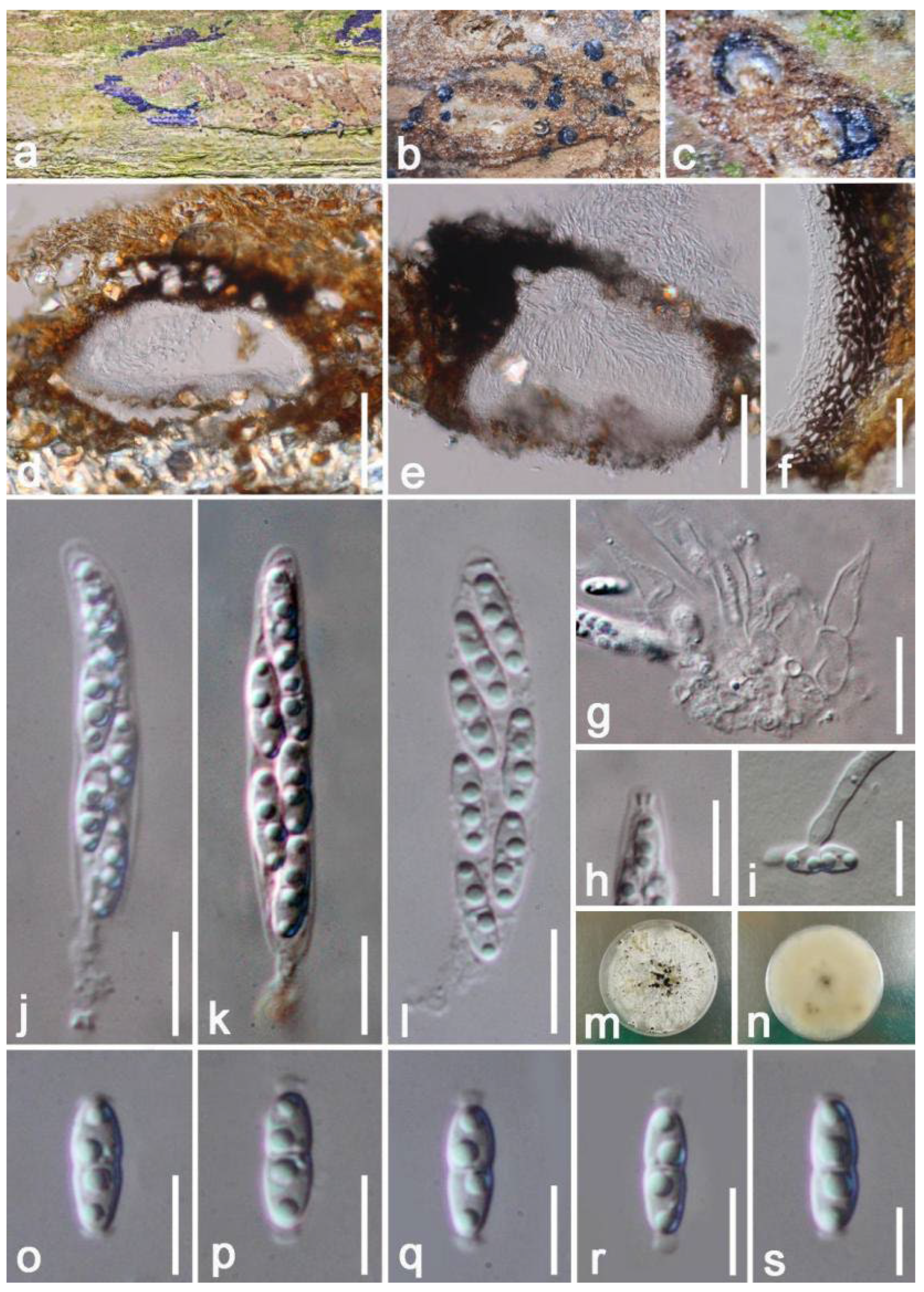
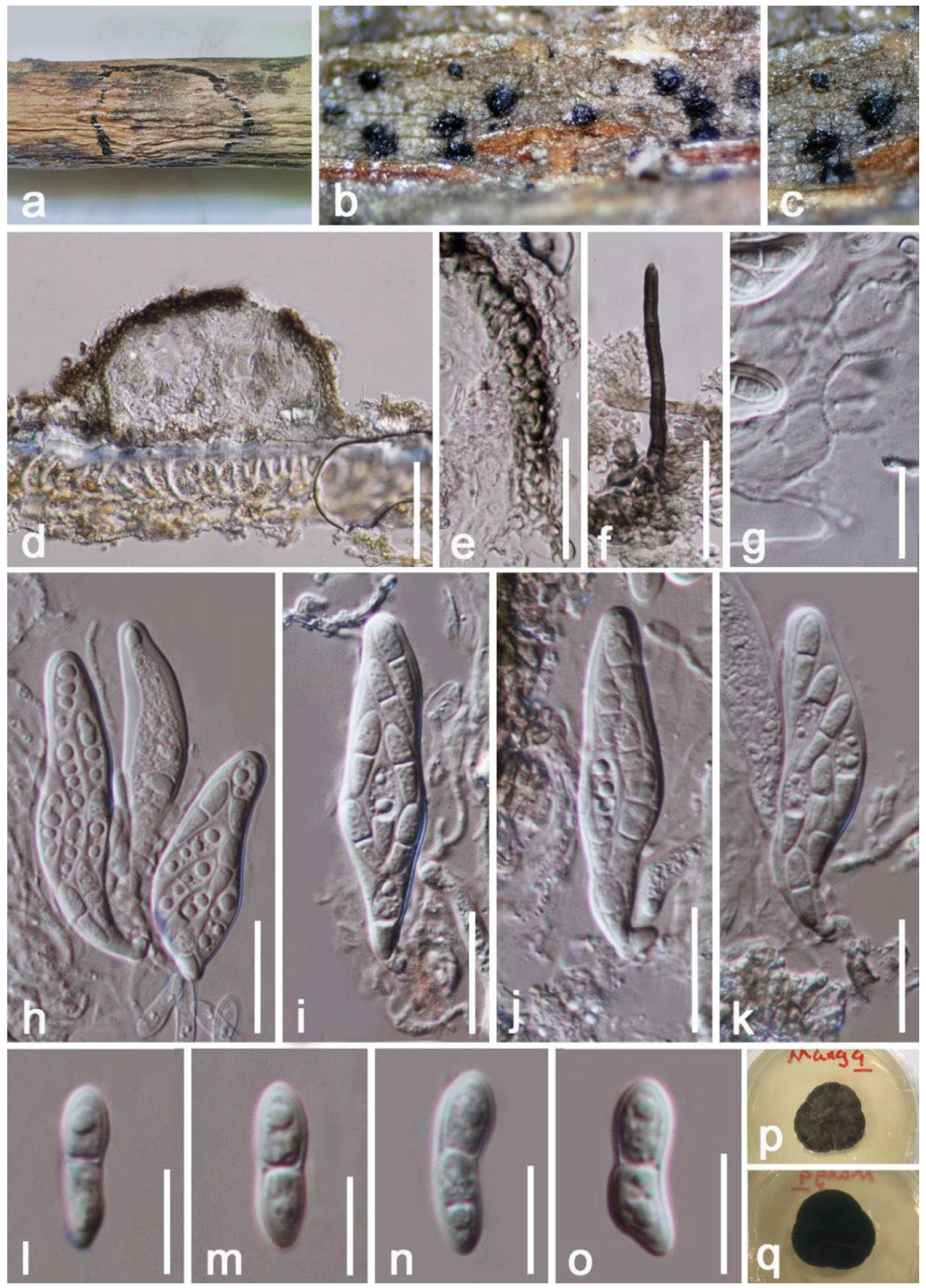
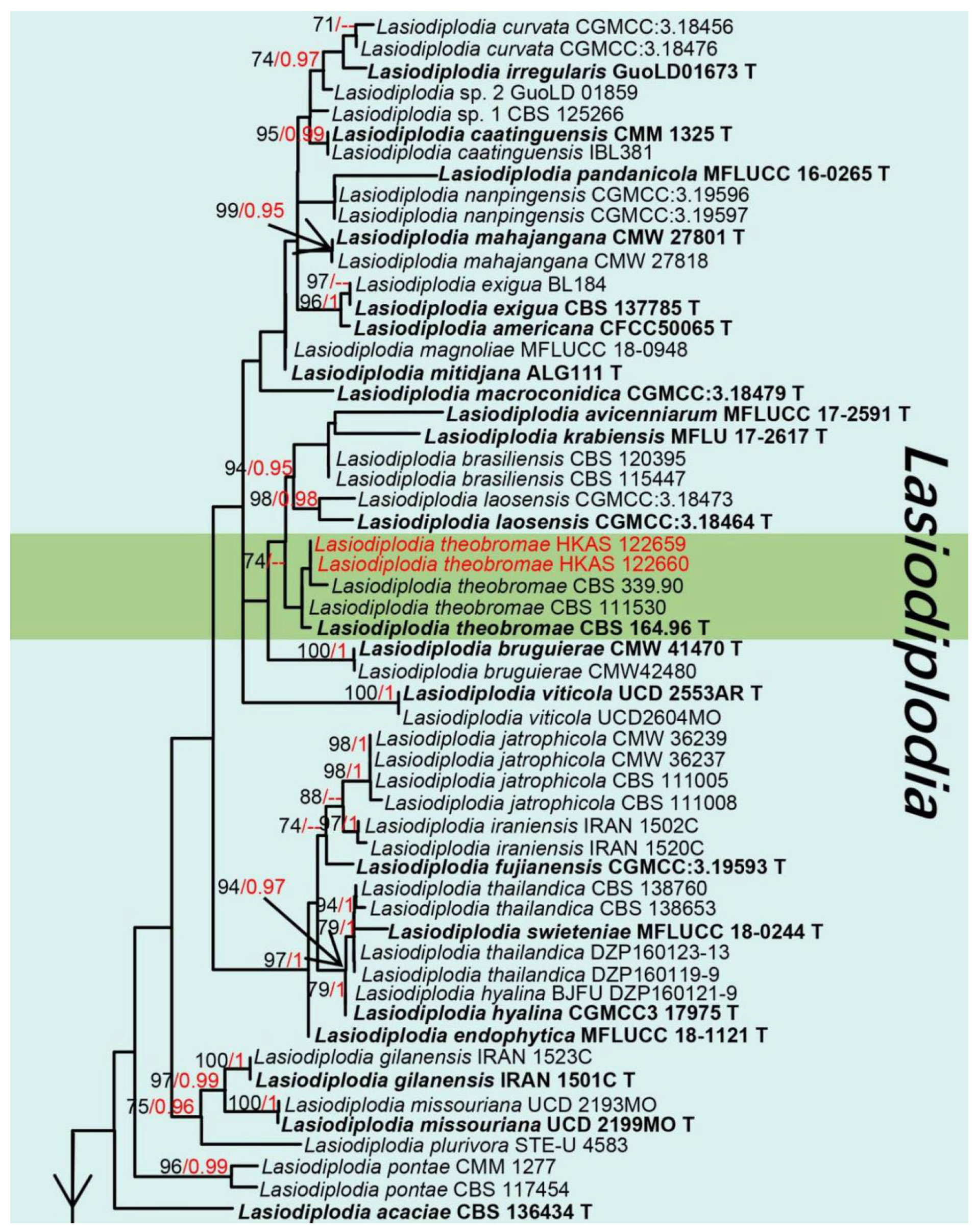
3. Results
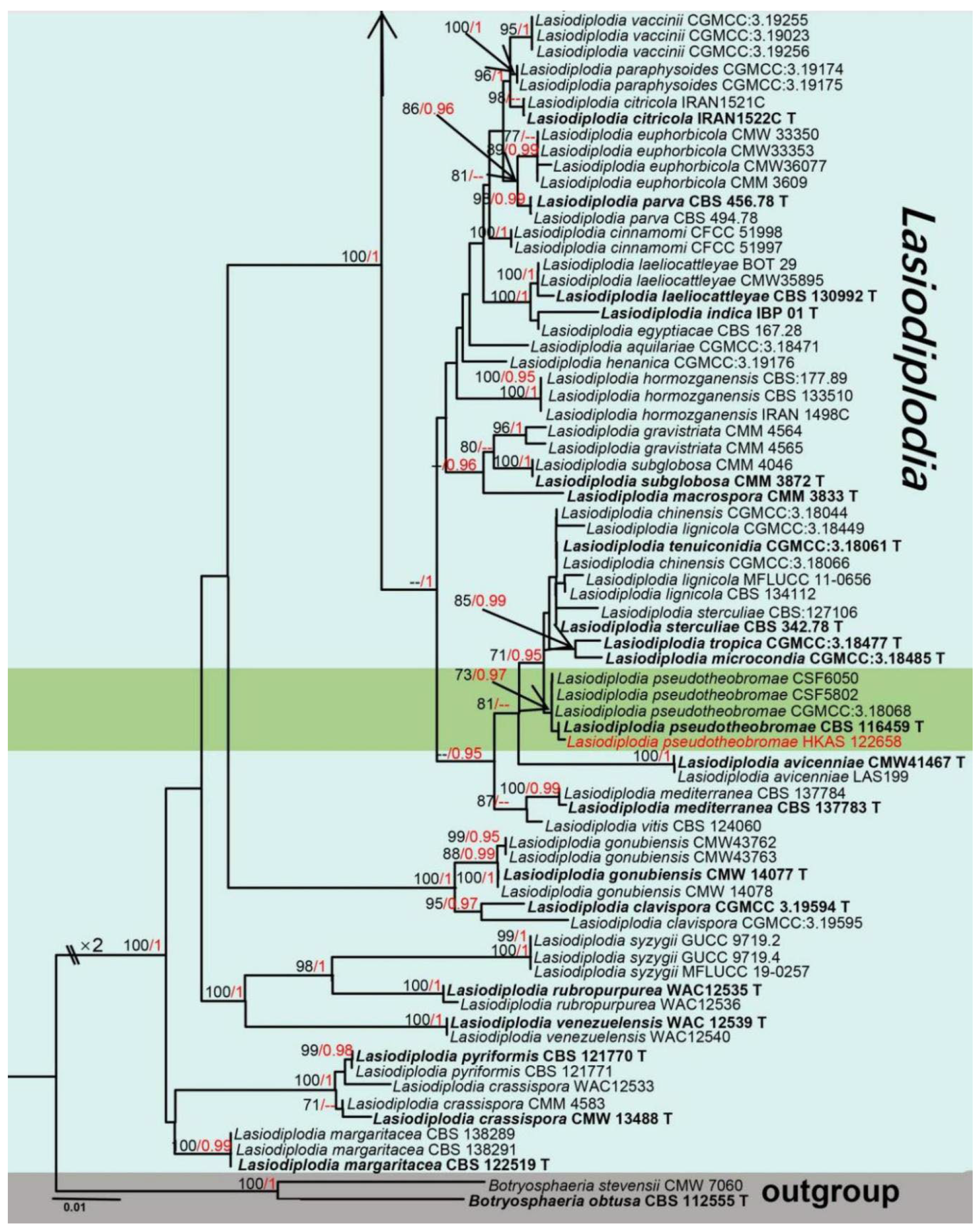
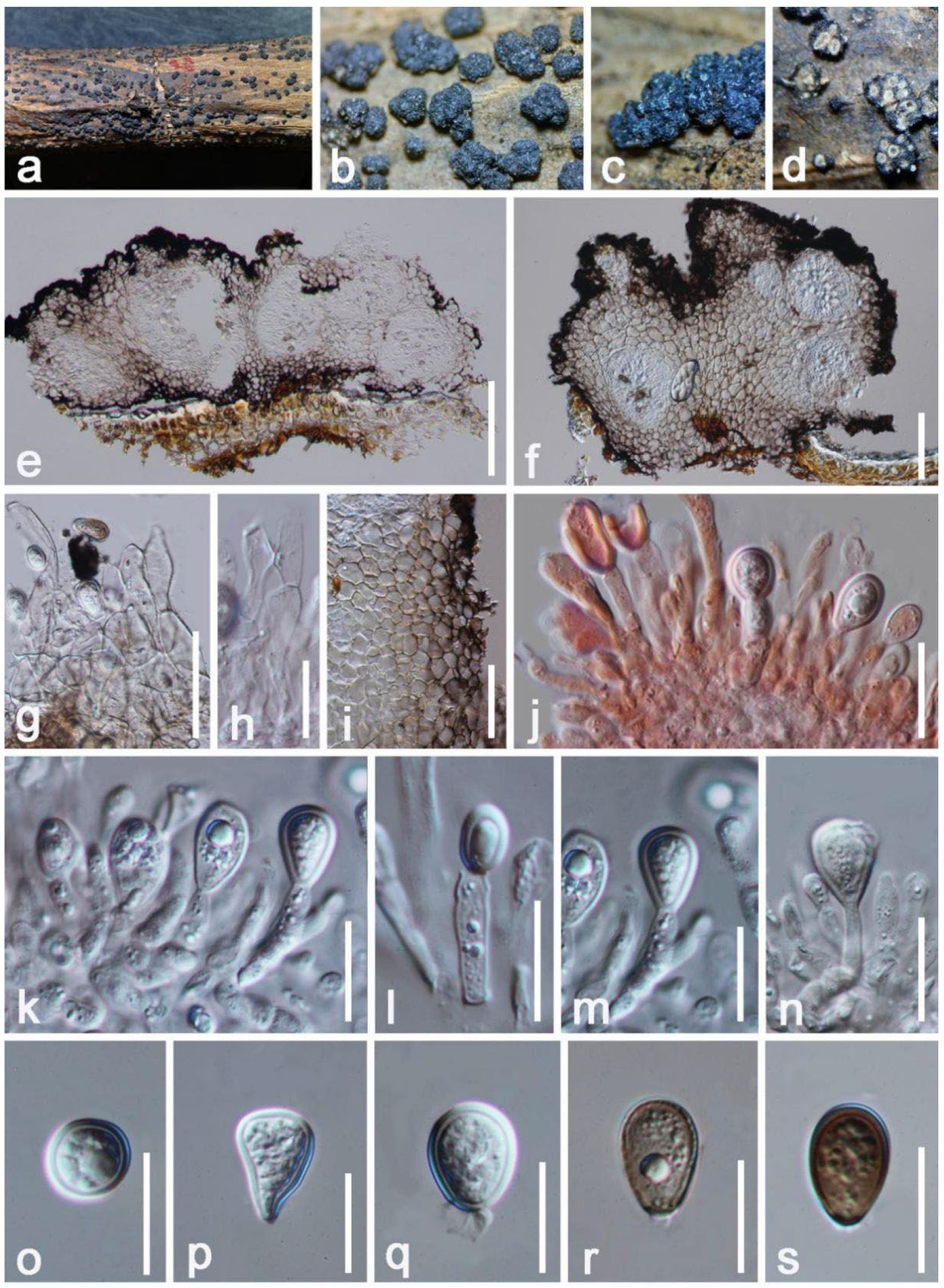
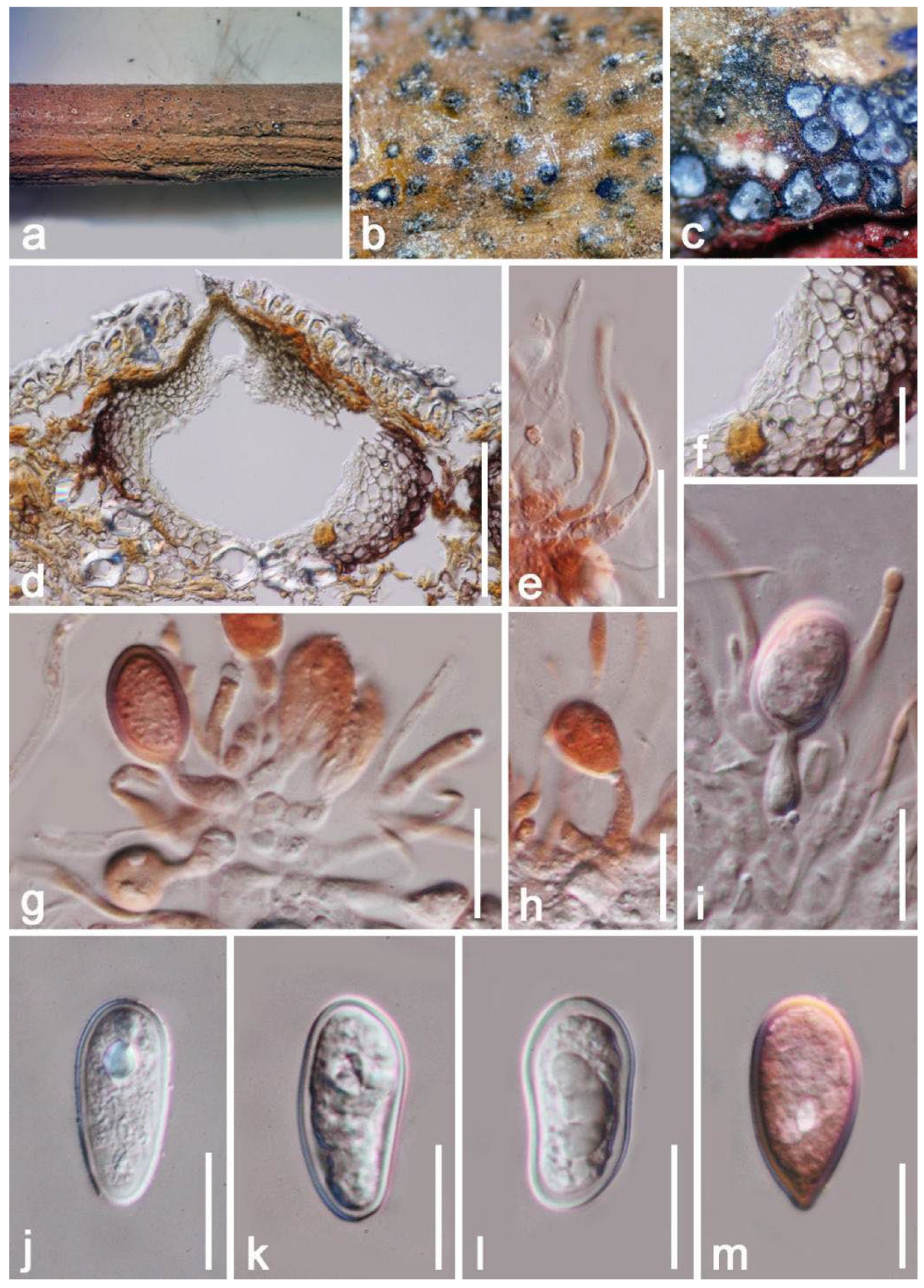
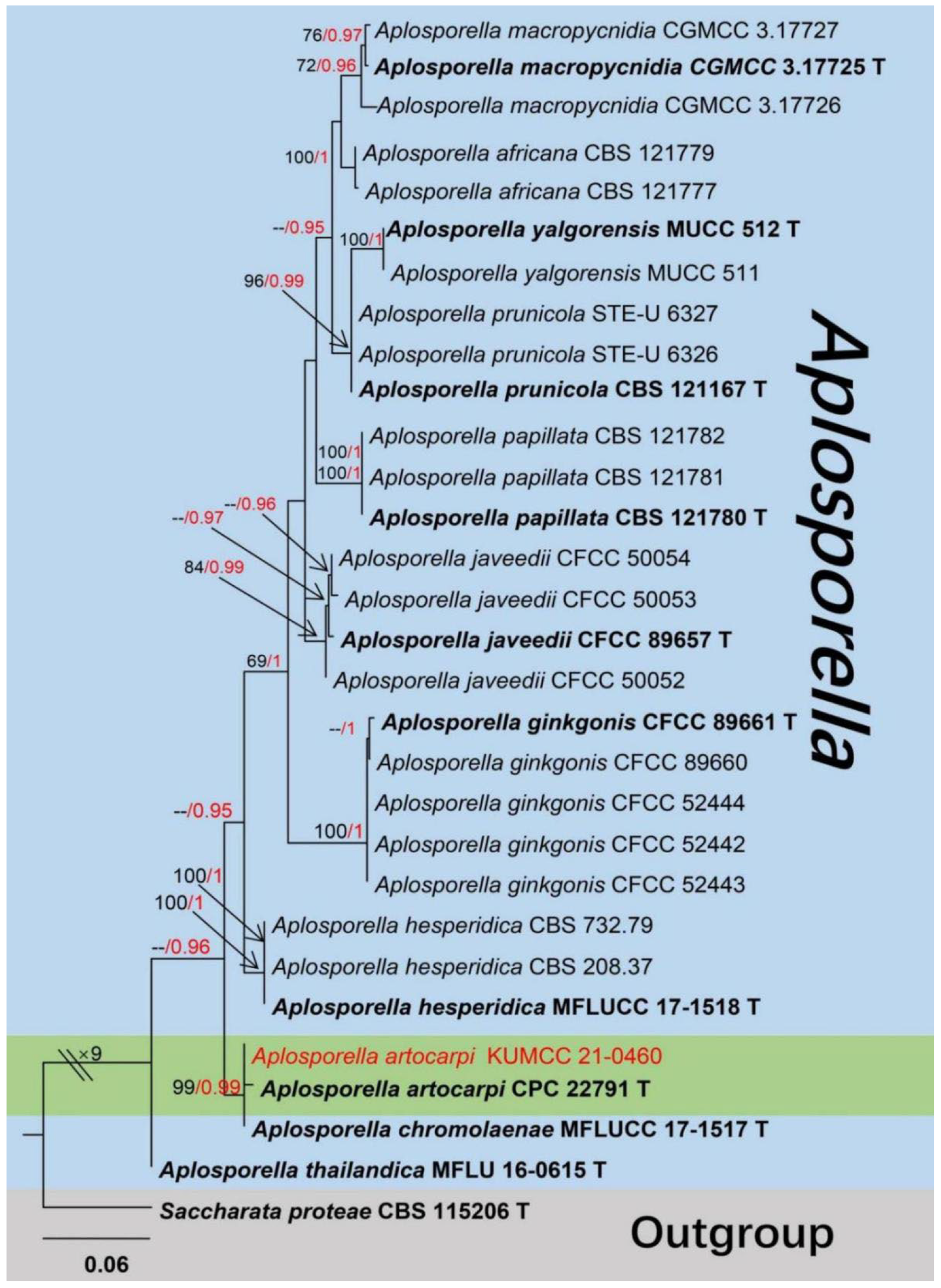
Taxonomy and Phylogenetic Analyses Results












4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Solís-Fuentes, J.A.; del Carmen Durán-de-Bazúa, M. Mango (Mangifera indica L.) Seed and Its Fats. In Nuts and Seeds in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2011; pp. 741–748. [Google Scholar]
- Jahurul, M.H.A.; Zaidul, I.S.M.; Ghafoor, K.; Al-Juhaimi, F.Y.; Nyam, K.L.; Norulaini, N.A.N.; Omar, A.M. Mango (Mangifera indica L.) by-products and their valuable components: A review. Food Chem. 2015, 183, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.; Singh, S.P. Mango: History origin and distribution. J. Pharmacogn. Phytochem. 2017, 6, 1257–1262. [Google Scholar]
- Lauricella, M.; Emanuele, S.; Calvaruso, G.; Giuliano, M.; D’Anneo, A. Multifaceted health benefits of Mangifera indica L. (Mango): The inestimable value of orchards recently planted in Sicilian rural areas. Nutrients 2019, 9, 525. [Google Scholar] [CrossRef] [PubMed]
- Chay, K.G.; Workeneh, A.; Shifera, B. A review on production and marketing of mango fruit. World J. Agric. Soil Sci. 2019, 2, 1–7. [Google Scholar] [CrossRef]
- Shah, K.A.; Patel, M.B.; Patel, R.J.; Parmar, P.K. Mangifera indica (mango). Phcog. Rev. 2010, 4, 42. [Google Scholar] [CrossRef]
- Rossman, A.Y.; Farr, D.F. Towards a virtual reality for plant-associated fungi in the United States and Canada. Biodivers. Conserv. 1997, 6, 739–751. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA, 2022. Available online: https://nt.arsgrin.gov/fungaldatabases/ (accessed on 4 November 2022).
- Ploetz, R.C. Diseases of mango. In Diseases of Tropical Fruit Crops; Ploetz, R.C., Ed.; CAB International: Wallingford, UK, 2003; pp. 327–363. [Google Scholar] [CrossRef]
- Haggag, W.M. Mango diseases in Egypt. Agric. Biol. J. N. Am. 2010, 1, 285–289. [Google Scholar] [CrossRef]
- Vieira, W.A.; Michereff, S.J.; de Morais, M.A.; Hyde, K.D.; Câmara, M.P. Endophytic species of Colletotrichum associated with mango in northeastern Brazil. Fungal Divers. 2014, 67, 181–202. [Google Scholar] [CrossRef]
- Dashyal, M.S.; Sangeetha, C.G.; Appanna, V.; Halesh, G.K.; Devappa, V. Isolation and Morphological Characterization of endophytic fungi isolated from Ten different varieties of Mango. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 717–726. [Google Scholar] [CrossRef]
- Yang, E.F.; Tibpromma, S.; Karunarathna, S.C.; Phookamsak, R.; Xu, J.C.; Zhao, Z.X.; Karunanayake, C.; Promputtha, I. Taxonomy and phylogeny of novel and extant taxa in Pleosporales associated with Mangifera indica from Yunnan, China (Series I). J. Fungi 2022, 8, 152. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Wang, F.; Wang, K.; Cai, L.; Zhao, M.; Kirk, P.M.; Fan, G.; Sun, Q.; Li, B.; Wang, S.; Yu, Z.; et al. Fungal names. 2022. Available online: https://nmdc.cn/fungalnames/ (accessed on 13 November 2022).
- Kirk, P.M. Index Fungorum. Available online: http://www.indexfungorum.org/names/names.asp (accessed on 5 May 2022).
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Liu, S.; Shen, L.L.; Wang, Y.; Xu, T.M.; Gates, G.; Cui, B.K. Species diversity and molecular phylogeny of Cyanosporus (Polyporales, Basidiomycota). Front. Microbiol. 2021, 12, 631166. [Google Scholar] [CrossRef]
- Cruywagen, E.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Phylogenetic species recognition and hybridisation in Lasiodiplodia: A case study on species from baobabs. Fungal Biol. 2017, 121, 420–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A.J.L. Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–13. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Dissanayake, L.S.; Wijayawardene, N.N.; Dayarathne, M.C.; Samarakoon, M.C.; Dai, D.Q.; Hyde, K.D.; Kang, J.C. Paraeutypella guizhouensis gen. et sp. nov. and Diatrypella longiasca sp. nov. (Diatrypaceae) from China. Biodivers. Data J. 2021, 9, e63864. [Google Scholar] [CrossRef]
- Crous, P.W.; Osieck, E.R.; Jurjević, Ž.; Boers, J.; van Iperen, A.L.; Starink-Willemse, M.; Dima, B.; Balashov, S.; Bulgakov, T.S.; Johnston, P.R.; et al. Fungal planet description sheets: 1284–1382. Persoonia 2021, 47, 178–374. [Google Scholar] [CrossRef]
- Dong, Z.Y.; Manawasinghe, I.S.; Huang, Y.H.; Shu, Y.X.; Phillips, A.J.L.; Dissanayake, A.J.; Hyde, K.D.; Xiang, M.M.; Luo, M. Endophytic Diaporthe associated with Citrus grandis cv. Tomentosa in China. Front. Microbiol. 2021, 11, 3621. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.K.; Zhu, A.H.; Liu, Z.D.; Qu, Z.; Li, Y.; Ma, H.X. Three New Species of Hypoxylon (Xylariales, Ascomycota) on a Multigene Phylogeny from Medog in Southwest China. J. Fungi 2022, 8, 500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar] [CrossRef] [PubMed]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Bhat, D.J.; Jeewon, R.; Stadler, M.; Samarakoon, M.C.; Malaithong, M.; Tanunchai, B.; et al. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 1–175. [Google Scholar] [CrossRef]
- Tsang, C.C.; Chan, K.F.; Chan, W.; Chan, J.F.; Au-Yeung, R.K.; Ngan, A.H.Y.; Lin, K.P.K.; Woo, P.C. Hepatic phaeohyphomycosis due to a novel dematiaceous fungus, Pleurostoma hongkongense sp. nov., and importance of antifungal susceptibility testing. Emerg. Microbes Infect. 2021, 10, 81–96. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Glez-Peña, D.; Gómez-Blanco, D.; Reboiro-Jato, M.; Fdez-Riverola, F.; Posada, D. FALTER: Program oriented conversion of DNA and protein alignments. Nucleic Acids Res. 2010, 38, 14–18. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analysis with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zhaxybayeva, O.; Gogarten, J.P. Bootstrap, Bayesian probability and maximum likelihood mapping: Exploring new tools for comparative genome analyses. Genomics 2002, 3, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J.A.; Wilgenbusch, J.C.; Warren, D.L.; Swofford, D.L. AWTY (are we there yet?): A system for graphical exploration of MCMC convergence in Bayesian phylogenetics. Bioinformatics 2008, 24, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v1.4.0. A Graphical Viewer of Phylogenetic Trees. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 25 June 2022).
- Vasilyeva, L.N.; Stephenson, S.L. Pyrenomycetes of the Great Smoky Mountains National Park. III. Cryptosphaeria, Eutypa and Eutypella (Diatrypaceae). Fungal Divers. 2006, 22, 243–254. [Google Scholar] [CrossRef]
- Moyo, P.; Mostert, L.; Halleen, F. Diatrypaceae species overlap between vineyards and natural ecosystems in South Africa. Fungal Ecol. 2019, 39, 142–151. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa-2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Liu, J.K.; Hyde, K.D.; Jones, E.B.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Bulliard, P. Histoire des Champignons; L’imprimerie de la Societe Typographique: Paris, France, 1791; Volume 1, pp. 1–368. [Google Scholar]
- Palapathwala, P.L.E.S.; Daranagama, D.A.; Abeywickrama, K.; Kannangara, B.T.S.D.P. New records of Hypoxylon hypomiltum and Hypomontagnella monticulosa from Pilikuththuwa lowland wet zone forest, Sri Lanka. Stud. Fungi 2019, 4, 142–152. [Google Scholar] [CrossRef]
- Ju, Y.M.; Rogers, J.D. A Revision of the Genus Hypoxylon; Mycologia Memoir n. 20; APS Press: St. Paul, MN, USA, 1996; p. 365. [Google Scholar]
- Sir, E.B.; Kuhnert, E.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Stadler, M. New species and reports of Hypoxylon from argentina recognized by a polyphasic approach. Mycol. Prog. 2016, 15, 1–19. [Google Scholar] [CrossRef]
- Hsieh, H.M.; Ju, Y.M.; Rogers, J.D. Molecular phylogeny of Hypoxylon and closely related genera. Mycologia 2005, 97, 844–865. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.; Wendt, L.; Hladki, A.I.; Romero, A.I.; Stadler, M.; Sir, E.B. Hypomontagnella (Hypoxylaceae): A new genus segregated from Hypoxylon by a polyphasic taxonomic approach. Mycol. Prog. 2019, 18, 187–201. [Google Scholar] [CrossRef]
- Wendt, L.; Sir, E.B.; Kuhnert, E.; Heitkämper, S.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Luangsa-ard, J.J.; Srikitikulchai, P.; Peršoh, D.; et al. Resurrection and emendation of the Hypoxylaceae, recognised from amultigene phylogeny of the Xylariales. Mycol. Prog. 2018, 17, 115–154. [Google Scholar] [CrossRef] [Green Version]
- Lambert, C.; Pourmoghaddam, M.J.; Cedeo-Sanchez, M.; Surup, F.; Khodaparast, S.A.; Krisai-Greilhuber, I.; Voglmayr, H.; Stradal, T.E.B.; Stadler, M. Resolution of the Hypoxylon fuscum complex (hypoxylaceae, xylariales) and discovery and biological characterization of two of its prominent secondary metabolites. J. Fungi 2021, 7, 131. [Google Scholar] [CrossRef] [PubMed]
- Khodaparast, S.A. Contribution to the knowledge of Hypoxylon and Annulohypoxylon in Guilan province (N Iran). Rostaniha 2012, 13, 197–206. [Google Scholar]
- Kout, J.; Zíbarová, L. Dřevomor Hypoxylon perforatum v České republice. Acta Mus. Richnov. Sect. Natur. 2016, 23, 39–43. [Google Scholar]
- Nitschke, T. Pyrenomycetes Germanici, 2nd ed.; Eduard Trewendt: Breslau, Germany, 1870; pp. 161–320. [Google Scholar]
- Senanayake, I.C.; Crous, P.W.; Groenewald, J.Z.; Maharachchikumbura, S.S.N.; Jeewon, R.; Phillips, A.J.L.; Bhat, D.J.; Perera, R.H.; Li, Q.R.; Li, W.J. Families of Diaporthales based on morphological and phylogenetic evidence. Stud. Mycol. 2017, 86, 217–296. [Google Scholar] [CrossRef]
- Rossman, A.Y.; Adams, G.C.; Cannon, P.F.; Castlebury, L.A.; Crous, P.W.; Gryzenhout, M.; Jaklitsch, W.M.; Mejia, L.C.; Stoykov, D.; Udayanga, D. Recommendations of generic names in Diaporthales competing for protection or use. IMA Fungus 2015, 6, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Udayanga, D.; Liu, X.; McKenzie, E.H.; Chukeatirote, E.; Bahkali, A.H.; Hyde, K.D. The genus Phomopsis: Biology, applications, species concepts and names of common phytopathogens. Fungal Divers. 2011, 50, 189–225. [Google Scholar] [CrossRef]
- Sun, W.X.; Huang, S.T.; Xia, J.W.; Zhang, X.G.; Li, Z. Morphological and molecular identification of Diaporthe species in south-western china, with description of eight new species. MycoKeys 2021, 77, 65–95. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Tsai, I.; Withee, P.; Tanjira, M.; Yen, C.Y.; Al-Rashed, S.; Elgorban, A.M.; Cheewangkoon, R. Diaporthe taiwanensis: A new taxon causing leaf spots and necrosis on Ixora chinensis in Taiwan. Phytotaxa 2020, 461, 155–165. [Google Scholar] [CrossRef]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vesterlund, S.R.; Helander, M.; Faeth, S.H.; Hyvönen, T.; Saikkonen, K. Environmental conditions and host plant origin override endophyte effects on invertebrate communities. Fungal Divers. 2011, 47, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Ash, G.J. The science, art and business of successful bioherbicides. Biol. Control 2010, 52, 230–240. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Tsai, I.; Wang, J.Y.; Withee, P.; Tanjira, M.; Lin, S.R.; Suwannarach, N.; Kumla, J.; Elgorban, A.M.; Cheewangkoon, R. Molecular phylogenetic diversity and biological characterization of Diaporthe species associated with leaf spots of Camellia sinensis in Taiwan. Plants 2021, 10, 1434. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. Insights into the genus Diaporthe: Phylogenetic species delimitation in the D. eres species complex. Fungal Divers. 2014, 67, 203–229. [Google Scholar] [CrossRef] [Green Version]
- De Vries, G.A. Cyphellophora laciniata nov. gen., nov. sp. and Dactylium Fusarioides Fragoso et Ciferri. Mycopathol. Mycol. Appl. 1962, 16, 47–54. [Google Scholar] [CrossRef]
- Yang, H.; Hyde, K.D.; Karunarathna, S.C.; Deng, C.; Gu, C.H.; Yang, S.A.; Zhang, Z.C. New species of Camptophora and Cyphellophora from China, and first report of sexual morphs for these genera. Phytotaxa 2018, 343, 149–159. [Google Scholar] [CrossRef]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Larsson, E.; Angelini, C.; Brandrud, T.E.; Dearnaley, J.D.W.; Dima, B.; Dovana, F.; et al. Fungal Planet description sheets: 1112–1181. Persoonia 2020, 45, 251–409. [Google Scholar] [CrossRef]
- Mayfield, D.A.; Karakaya, A.; Batzer, J.C.; Blaser, J.M.; Gleason, M.L. Diversity of sooty blotch and flyspeck fungi from apples in northeastern Turkey. Eur. J. Plant Pathol. 2012, 135, 805–815. [Google Scholar] [CrossRef]
- Ellis, J.B.; Everhart, B.M. New west American fungi. Erythea 1894, 2, 17–27. [Google Scholar]
- Tennakoon, D.S.; Phillips, A.J.L.; Phookamsak, R.; Ariyawansa, H.A.; Bahkali, A.H.; Hyde, K.D. Sexual morph of Lasiodiplodia pseudotheobromae (Botryosphaeriaceae, Botryosphaeriales, Dothideomycetes) from China. Mycosphere 2016, 7, 990–1000. [Google Scholar] [CrossRef]
- Dou, Z.P.; He, W.; Zhang, Y. Does morphology matter in taxonomy of Lasiodiplodia? An answer from Lasiodiplodia hyalina sp nov. Mycosphere 2017, 8, 1014–1027. [Google Scholar] [CrossRef]
- Sakalidis, M.L.; Ray, J.D.; Lanoiselet, V.; Hardy, G.E.; Burgess, T.I. Pathogenic Botryosphaeriaceae associated with Mangifera indica in the Kimberley Region of Western Australia. Eur. J. Plant Pathol. 2011, 130, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Gálvez, E.; Guerrero, P.; Barradas, C.; Crous, P.W.; Alves, A. Phylogeny and pathogenicity of Lasiodiplodia species associated with dieback of mango in Peru. Fungal Biol. 2017, 121, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.W.; Wang, C.S.; Wang, G.L. First report of bark cracking of Koelreuteria bipinnata var. integrifoliola caused by Lasiodiplodia theobromae in China. Plant Dis. 2012, 96, 1579. [Google Scholar] [CrossRef]
- Burgess, T.I.; Tan, Y.P.; Garnas, J.; Edwards, J.; Scarlett, K.A.; Shuttleworth, L.A.; Daniel, R.; Dann, E.K.; Parkinson, L.E.; Dinh, Q.; et al. Current status of the Botryosphaeriaceae in Australia. Australas. Plant Pathol. 2019, 48, 35–44. [Google Scholar] [CrossRef]
- Ma, Y.; Ahmad, T.; Zheng, Y.; Nie, C.; Liu, Y. First report of postharvest stem end rot of mango fruit (Mangifera indica) caused by Lasiodiplodia theobromae in China. Plant Dis. 2021, 105, 2715. [Google Scholar] [CrossRef]
- Zhao, J.P.; Lu, Q.; Liang, J.; Decock, C.; Zhang, X.Y. Lasiodiplodia pseudotheobromae, a new record of pathogenic fungus from some subtropical and tropical trees in southern China. Cryptogam. Mycol. 2010, 31, 431–439. [Google Scholar]
- Trakunyingcharoen, T.; Lombard, L.; Groenewald, J.Z.; Cheewangkoon, R.; To-anun, C.; Crous, P.W. Caulicolous Botryosphaeriales from Thailand. Persoonia 2015, 34, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Urbez-Torres, J.R.; Castro-Medina, F.; Mohali, S.R.; Gubler, W.D. Botryosphaeriaceae species associated with cankers and dieback symptoms of Acacia mangium and Pinus caribaea var. hondurensis in Venezuela. Plant Dis. 2016, 100, 2455–2464. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L.; Jayawardena, R.S.; Xu, W.; Hu, M.; Li, X.H.; Liu, J.H.; Hyde, K.D.; Yan, J. Lasiodiplodia theobromae and L. pseudotheobromae causing leaf necrosis on Camellia sinensis in Fujian Province, China. Can. J. Plant Pathol. 2019, 41, 277–284. [Google Scholar] [CrossRef]
- Scarlett, K.A.; Shuttleworth, L.A.; Collins, D.; Rothwell, C.T.; Guest, D.I.; Daniel, R. Botryosphaeriales associated with stem blight and dieback of blueberry (Vaccinium spp.) in New South Wales and Western Australia. Australas. Plant Pathol. 2019, 48, 45–57. [Google Scholar] [CrossRef]
- Li, G.; Slippers, B.; Wingfield, M.J.; Chen, S. Variation in Botryosphaeriaceae from Eucalyptus plantations in Yunnan Province in southwestern China across a climatic gradient. IMA Fungus 2020, 11, 1–49. [Google Scholar] [CrossRef] [PubMed]
- Spegazzini, C. Fungi argentini. Pugillus tertius (Continuacion). Anales Soc. Ci. Argent. 1880, 10, 145–168. [Google Scholar]
- Ekanayaka, A.H.; Dissanayake, A.J.; Jayasiri, S.C.; To-anun, C.; Jones, E.B.G.; Zhao, Q.; Hyde, K.D. Aplosporella thailandica; a novel species revealing the sexual-asexual connection in Aplosporellaceae (Botryosphaeriales). Mycosphere 2016, 7, 440–447. [Google Scholar] [CrossRef]
- Slippers, B.; Boissin, E.; Phillips, A.J.L.; Groenewald, J.Z.; Lombard, L.; Wingfield, M.J.; Postma, A.; Burgess, T.I.; Crous, P.W. Phylogenetic lineages in the Botryosphaeriales: A systematic and evolutionary framework. Stud. Mycol. 2013, 76, 31–49. [Google Scholar] [CrossRef] [Green Version]
- Montagne, J.P.F.C. Cryptogamia Guyanensis seu plantarum cellularium in Guyana gallica annis 1835–1849 a cl. Leprieur collectarum enumeration universalis. Ann. Sci. Nat. Bot. ser. 1850, 3, 283–309. [Google Scholar]
- Wibberg, D.; Stadler, M.; Lambert, C.; Bunk, B.; Spröer, C.; Rückert, C.; Kalinowski, J.; Cox, R.J.; Kuhnert, E. High quality genome sequences of thirteen Hypoxylaceae (Ascomycota) strengthen the phylogenetic family backbone and enable the discovery of new taxa. Fungal Divers. 2020, 106, 7–28. [Google Scholar] [CrossRef]
- Anisa, L.; Erman, M.; Yurnaliza, Y.; Mohammad, B. Chemical analysis and anticancer activity of sesterterpenoid from an endophytic fungus Hypomontagnella monticulosa Zg15SU and its host Zingiber griffithii Baker. Heliyon 2021, 7, e06292. [Google Scholar] [CrossRef]
- Ju, Y.M.; Rogers, J.D. The Xylariaceae of Taiwan (excluding Anthostomella). Mycotaxon 1999, 73, 343–440. [Google Scholar]
- Suwannasai, N.; Rodtong, S.; Thienhirun, S.; Whalley, A.J.S. New species and phylogenetic relationships of Hypoxylon species found in Thailand inferred from the internal transcribed spacer regions of ribosomal DNA sequences. Mycotaxon 2005, 94, 303–324. [Google Scholar]
- Rogers, J.D.; Ju, Y.M. Key to Hawaiian Rosellinia taxa and additions to host-fungus index. N. Am. Fungi 2015, 10, 1–4. [Google Scholar]
- U’Ren, J.M.; Miadlikowska, J.; Zimmerman, N.B.; Lutzoni, F.; Stajich, J.E.; Arnold, A.E. Contributions of North American endophytes to the phylogeny, ecology, and taxonomy of Xylariaceae (Sordariomycetes, Ascomycota). Mol. Phylogenet. Evol. 2016, 98, 210–232. [Google Scholar] [CrossRef] [Green Version]
- Zainee, N.F.A.; Ismail, A.; Ibrahim, N.; Ismail, A. Seaweed temporal distribution in southeast coast of Peninsular Malaysia and isolation of endophytic fungi. In AIP Conf.Proc.; AIP Publishing LLC: Melville, NY, USA, 2018; Volume 1940, p. 020069. [Google Scholar]
- Hsieh, C.W.; Chuang, Y.Y.; Lee, M.Z.; Kirschner, R. First inventory of fungi in symptomless and symptomatic Chinese Mesona indicates phytopathological threat. Plant Dis. 2020, 104, 2391–2397. [Google Scholar] [CrossRef]
- Chethana, K.W.T.; Niranjan, M.; Dong, W.; Samarakoon, M.C.; Bao, D.F.; Calabon, M.S.; Chaiwan, N.; Chuankid, B.; Dayarathne, M.C.; de Silva, N.I.; et al. AJOM new records and collections of fungi: 101–150. Asian. J. Mycol. 2021, 4, 113–260. [Google Scholar]
- Glawe, A.; Jacobs, K.A. Taxonomic notes on Eutypella vitis, Cryptosphaeria populina, and Diatrype stigma. Mycologia 1987, 79, 135–139. [Google Scholar] [CrossRef]
- Gao, L.; Ma, Y.Q.; Zhao, W.Y.; Wei, Z.Y.; Gleason, L.M.; Chen, H.C.; Hao, L.; Sun, G.Y.; Zhang, R. Three new species of Cyphellophora (Chaetothyriales) associated with Sooty Blotch and Flyspeck. PLoS ONE 2015, 10, e0136857. [Google Scholar] [CrossRef]
- Reinking, O.A. Citrus diseases of the Philippines, southern China, Indo-China and Siam. Philipp. Agric. 1921, 9, 121–179. [Google Scholar]
- Teodoro, N.G. An Enumeration of Philippine Fungi. Techn. Bull. Dept. Agric. Comm. Manila 1937, 4, 1–585. [Google Scholar]
- Pitt, W.M.; Trouillas, F.P.; Gubler, W.D.; Savocchia, S.; Sosnowski, M.R. Pathogenicity of Diatrypaceous fungi on grapevines in Australia. Plant Dis. 2013, 97, 749–756. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Purahong, W.; Zhang, W.; Wubet, T.; Li, X.H.; Liu, M.; Zhao, W.; Hyde, K.D.; Liu, L.H.; Yan, J. Biodiversity of fungi on Vitis vinifera L. revealed by traditional and high-resolution culture-independent approaches. Fungal Divers. 2018, 90, 1–84. [Google Scholar] [CrossRef] [Green Version]
- Mehrabi, M.; Asgari, B.; Hemmati, R. Two new species of Eutypella and a new combination in the genus Peroneutypa (Diatrypaceae). Mycol. Prog. 2019, 18, 1057–1069. [Google Scholar] [CrossRef]
- Sohrabi, M.; Mohammadi, H.; Leon, M.; Armengol, J.; Banihashemi, Z. Fungal pathogens associated with branch and trunk cankers of nut crops in Iran. Eur. J. Plant Pathol. 2020, 157, 327–351. [Google Scholar] [CrossRef]
- Mehrabi, M.; Hemmati, R.; Vasilyeva, L.N.; Trouillas, F.P. Diatrypella macrospora sp. nov. and new records of diatrypaceous fungi from Iran. Phytotaxa 2016, 252, 43–55. [Google Scholar] [CrossRef]
- Tulasne, E.L.R.; Tulasne, C. Selecta Fungorum Carpologia ea documenta et icones potissimum exhibens quae varia fructuum et seminum genera in eodem fungosimul aut vicissim adesse demonstrent; Imperiali Typographeo Excudebatur: Paris, France, 1863; Volume 2, p. 247. [Google Scholar]
- Gramaje, D.; Mostert, L.; Groenewald, J.Z.; Crous, P.W. Phaeoacremonium: From esca disease to phaeohyphomycosis. Fungal Biol. 2015, 119, 759–783. [Google Scholar] [CrossRef]
- Vijaykrishna, D.; Mostert, L.; Jeewon, R.; Gams, W.; Crous, P.W. Pleurostomophora, an anamorph of Pleurostoma (Calosphaeriales), a new anamorph genus morphologically similar to Phialophora. Stud. Mycol. 2004, 50, 387–395. [Google Scholar] [CrossRef]
- Martina, R.; Jaklitsch, W.M.; Kamila, R.; Václav, S.; Tamás, P. Phylogenetic reconstruction of the Calosphaeriales and togniniales using five genes and predicted rna secondary structures of its, and Flabellascus tenuirostris gen. et sp. nov. PLoS ONE 2015, 10, e0144616. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.K.; Hyde, K.D.; Mapook, A.; Maharachchikumbura, S.S.N.; Bhat, J.D.; McKenzie, E.H.C.; Jeewon, R.; Wen, T.C. Taxonomic studies of some often over-looked Diaporthomycetidae and Sordariomycetidae. Fungal Divers. 2021, 111, 443–572. [Google Scholar] [CrossRef]
- Lynch, S.C.; Zambino, P.J.; Scott, T.A.; Eskalen, A. Occurence, incidence and associations among fungal pathogens and Agrilus auroguttatus, and their roles in Quercus agrifolia decline in California. Forest Pathol. 2014, 44, 62–74. [Google Scholar] [CrossRef]
- Barr, M.E. Notes on the Calosphaeriales. Mycologia 1985, 77, 549–565. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa Names | Loci | The BLASTn Results |
|---|---|---|
| Aplosporellaartocarpi (KUMCC 21-0460) | LSU(LR0R/LR5): | 100% (571/571 bp; 0 gap) similar to Aplosporella chromolaenae (MFLUCC 17-1517) |
| ITS(ITS4/ITS5): | 100% (554/554 bp; 0 gap) similar to Aplosporella artocarpi (CPC 22791) and A. chromolaenae (MFLUCC 17-1517) | |
| tef1-α(688F/1251R): | 100% (250/250 bp; 0 gap) similar to Aplosporella artocarpi (CPC 22791) | |
| Cyphellophorahongheensis (HKAS 122661) | SSU(NS1/NS4): | 99% (964/966 bp; 0 gap) similar to Cyphellophora sessilis (CBS 238.93, CBS 243.85) |
| LSU(LR0R/LR5): | 98% (876/893 bp; 5 gaps) similar to Cyphellophora sessilis (SP12386Ca) | |
| ITS(ITS4/ITS5): | 92% (509/556 bp; 8 gaps) similar to Cyphellophora livistonae (PhliHN2702, CPC 19433) | |
| rpb1(AR/CF): | 75% (495/660 bp; 21 gaps) similar to Cyphellophora oxyspora (CBS 416.89, CBS 698.73) | |
| Diaporthehongheensis (KUMCC 21-0457) | ITS(ITS4/ITS5): | 99% (556/562 bp; 2 gaps) similar to Diaporthe eugeniae (ASHM304) and D. phaseolorum (B3147) |
| tef1-α(728F/986R): | 96% (330/342 bp; 3 gaps) similar to Diaporthe pascoei (PBMR343) | |
| β-tubulin (Bt2a/Bt 2b): | 100% (495/496 bp; 1 gap) similar to Diaporthe endocitricola (ZHKUCC 20-0013) | |
| CAL (228F/373R): | 97% (468/480 bp; 3 gaps) similar to Diaporthe sp. (CFCC 53101) | |
| Hypomontagnellmonticulosa (HKAS 122664) | LSU(LR0R/LR5): | 100% (841/842 bp; 0 gap) similar to Hypomontagnella monticulosa (MFLUCC 18-0362, XY02480) |
| ITS(ITS4/ITS5): | 99% (571/576 bp; 1 gap) similar to Hypomontagnella monticulosa (67.3.3) | |
| β-tubulin (Bt2a/Bt 2b): | 97% (392/403 bp; 1 gap) similar to Hypomontagnella monticulosa (EK13014, YMJ 90080806) | |
| rpb2(5F/7cR): | 99% (1074/1077 bp; 1 gap) similar to Hypomontagnella monticulosa (CLL 205) | |
| Hypoxylonhongheensis (HKAS 122663) | LSU(LR0R/LR5): | 100% (840/841 bp; 1 gap) similar to Hypoxylon perforatum (XZ142) |
| ITS(ITS4/ITS5): | 98% (625/637 bp; 11 gaps) similar to Hypoxylon perforatum (MoEF023, KoRLI047347) | |
| β-tubulin(T1/T22): | 97% (1386/1424 bp; 5 gaps) similar to Hypoxylon perforatum (STMA 14051) and H. rubiginosum (YMJ 4) | |
| rpb2(5F/7cR): | 98 % (973/993 bp; 0 gap) similar to Hypoxylon perforatum (DSM:107930) and H. rubiginosum (FL1170) | |
| Lasiodiplodiatheobromae (HKAS 122660) | LSU(LR0R/LR5): | 100% (846/847 bp; 0 gap) similar to Lasiodiplodia krabiensis (MFLU 17-2617) and L. theobromae (CBS 129758) |
| ITS(ITS4/ITS5): | 100% (498/498 bp; 0 gap) similar to Lasiodiplodia theobromae (ZW 50-1, IRNBS73) | |
| tef1-α(668F/986R): | 100% (315/315 bp; 0 gap) similar to Lasiodiplodia theobromae (ZWLT 482, BOM230) | |
| rpb2(LasF/LasR): | 100% (570/570 bp; 0 gap) similar to Lasiodiplodia theobromae (DAR82930) | |
| β-tubulin(Bt2a/Bt 2b): | 99% (464/469 bp; 2 gaps) similar to Lasiodiplodia theobromae (LTHEOB 7940) | |
| Lasiodiplodiapseudotheobromae (HKAS 122658) | LSU(LR0R/LR5): | 100% (862/862 bp; 0 gap) similar to Lasiodiplodia hyalina (CGMCC 3.17975) and L. pseudotheobromae (CBS 447.62) |
| ITS(ITS4/ITS5): | 100% (498/498; 0 gap) similar to Lasiodiplodia pseudotheobromae (AY-11, KoRLI047143) | |
| tef1-α(668F/986R: | 100% (307/307 bp; 0 gap) similar to Lasiodiplodia pseudotheobromae (MPMR65, MFLUCC 17-2289, ZK201) | |
| rpb2(LasF/LasR): | 100% (569/569 bp; 0gap) similar to Lasiodiplodia pseudotheobromae (DAR83095, CERC 3496) | |
| β-tubulin(Bt2a/Bt 2b): | 100% (429/429 bp; 0 gap) similar to Lasiodiplodia pseudotheobromae (G32) | |
| Mangifericolahongheensis (HKAS 122665) | LSU(LR0R/LR5): | 99% (844/856bp; 1 gap) similar to Diatrype dalbergiae (CBS 147068) |
| ITS(ITS4/ITS5): | 99% (552/557bp; 2 gaps) similar to Diatrypella pulvinata (B1B085-3-EM2CC568) | |
| β-tubulin(Bt2a/Bt2b): | 90% (352/393bp; 18 gaps) similar to Melanostictus thailandicus (MFLU 19-2123, MFLU 19-2146) | |
| Paraeutypellacitricola (HKAS 122667) | LSU(LR0R/LR5): | 100% (841/841 bp; 1 gap) similar to Paraeutypella citricola (CBS 128334, KUMCC 21-0461) |
| ITS(ITS4/ITS5): | 100% (531/531 bp; 0 gap) similar to Paraeutypella citricola (STEU 8186, KUMCC 21-0461, BRPET19) | |
| β-tubulin(Bt2a/Bt2b): | 100% (370/373 bp; 1 gap) similar to Paraeutypella citricola (HUEFS 194248, HUEFS 131041) | |
| Pleurostoma ootheca (HKAS 122679) | SSU(NS1/NS4): | 100% (1022/1022 bp; 0 gap) similar to Pleurostoma ootheca (CBS 115329) and P. repens (CBS 294.39) |
| LSU(LR0R/LR5): | 100% (559/560 bp; 0 gap) similar to Pleurostoma ootheca (CBS:115329, CMU 23858) | |
| ITS(ITS4/ITS5): | 100% (519/519 bp; 0 gap) similar to Pleurostoma ootheca (CBS 126089) and P. ootheca (CBS 115329) | |
| rpb2(5F/7cR): | 100% (940/947 bp; 4 gaps) similar to Pleurostoma ootheca (CBS 115329) | |
| β-tubulin(Bt2a/Bt 2b): | 100% (451/451 bp; 0 gap) similar to Pleurostoma ootheca (CBS 115329) |
| Species | Conidiomata | Conidia | References |
|---|---|---|---|
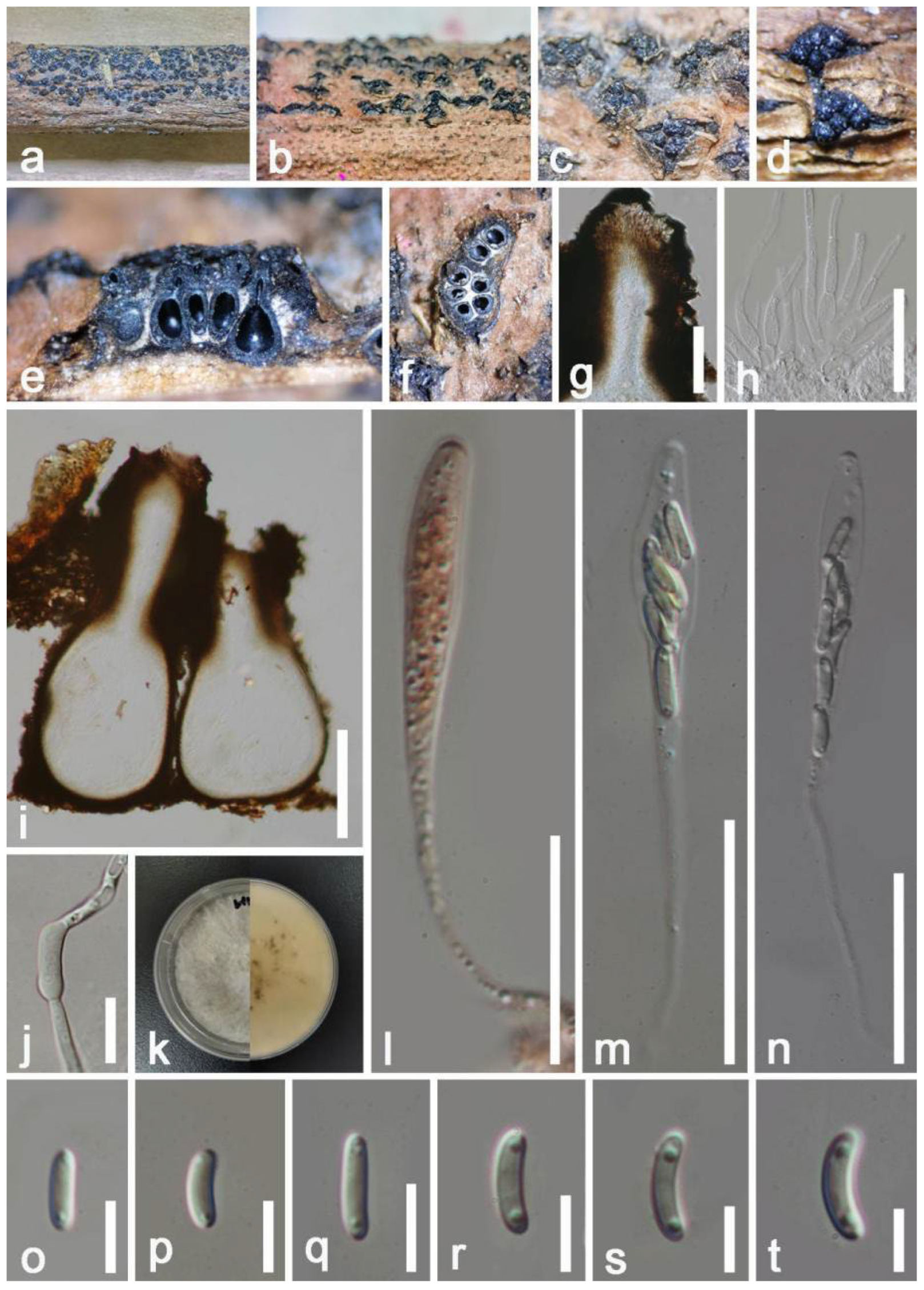
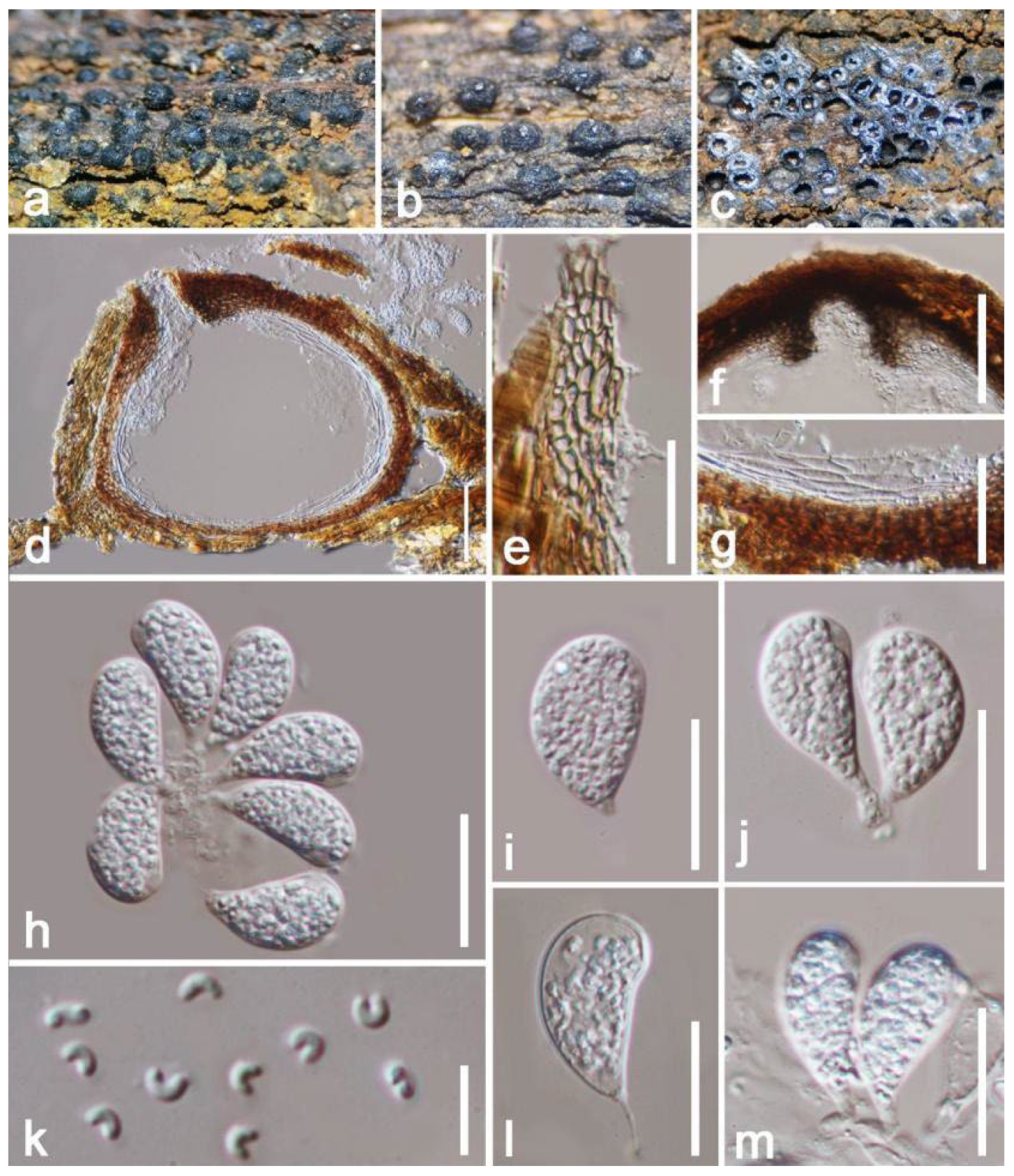
| A.artocarpi (HKAS 122656) | 340–430 × 620–670 μm, erumpent to complete immersed, discoid or irregular in shape, dark brown to black, multilocular, with a pore-like ostiolar. | 14–19 × 7–10 μm, hyaline to dark brown, broadly ellipsoidal to subcylindrical, aseptate, blunt ends, rough-walled, guttulate. | This study |
| A.artocarpi (CBS H-21931) | (350–)540–550(–650) × (490–)540–600(–700) µm, pycnidial, semi-immersed, mostly solitary, dark brown to black, with globose base, multilocular. | (17–)18–21(–22) × (9–)10–11 µm, ellipsoid to ovoid, smooth, moderately thick-walled, with granular content, aseptate. | [81] |
| A.chromolaenae (MFLU 20-0298) | 75–145 × 80–160 µm, gregarious, with 2–3 locules forming groups immersed in conidiostroma, globose to subglobose. | (13–)16–20 × 8.5–12 µm, hyaline to brown to dark brown, aseptate, ellipsoid or oval to reniform, thick-walled, verruculose. | [31] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, E.-F.; Karunarathna, S.C.; Dai, D.-Q.; Stephenson, S.L.; Elgorban, A.M.; Al-Rejaie, S.; Xiong, Y.-R.; Promputtha, I.; Samarakoon, M.C.; Tibpromma, S. Taxonomy and Phylogeny of Fungi Associated with Mangifera indica from Yunnan, China. J. Fungi 2022, 8, 1249. https://doi.org/10.3390/jof8121249
Yang E-F, Karunarathna SC, Dai D-Q, Stephenson SL, Elgorban AM, Al-Rejaie S, Xiong Y-R, Promputtha I, Samarakoon MC, Tibpromma S. Taxonomy and Phylogeny of Fungi Associated with Mangifera indica from Yunnan, China. Journal of Fungi. 2022; 8(12):1249. https://doi.org/10.3390/jof8121249
Chicago/Turabian StyleYang, Er-Fu, Samantha C. Karunarathna, Dong-Qin Dai, Steven L. Stephenson, Abdallah M. Elgorban, Salim Al-Rejaie, Yin-Ru Xiong, Itthayakorn Promputtha, Milan C. Samarakoon, and Saowaluck Tibpromma. 2022. "Taxonomy and Phylogeny of Fungi Associated with Mangifera indica from Yunnan, China" Journal of Fungi 8, no. 12: 1249. https://doi.org/10.3390/jof8121249