
Sporobolomyces lactucae sp. nov. (Pucciniomycotina, Microbotryomycetes, Sporidiobolales): An Abundant Component of Romaine Lettuce Phylloplanes

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lettuce Leaf Preparation and Culturing of Fungal Isolates
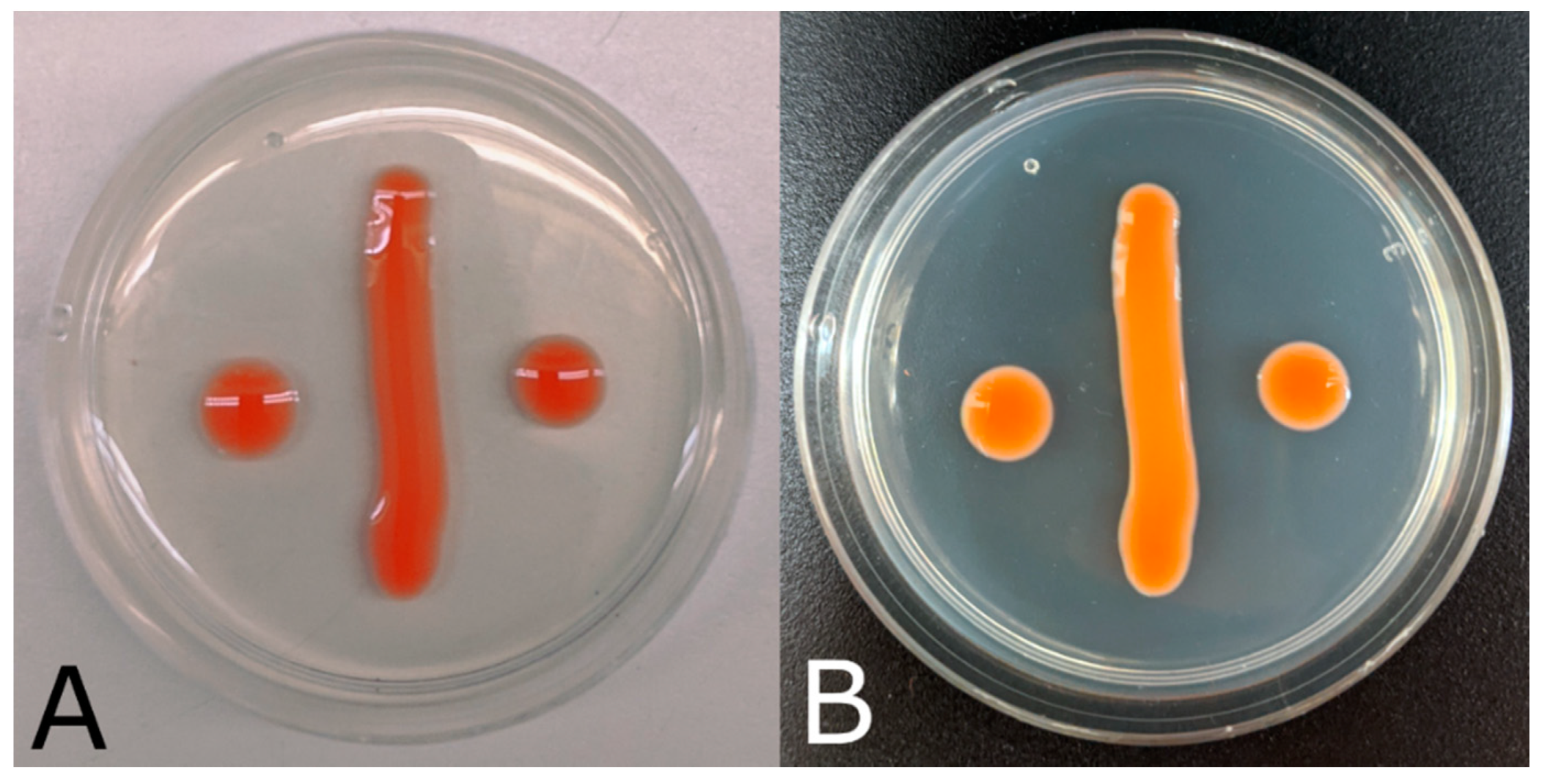
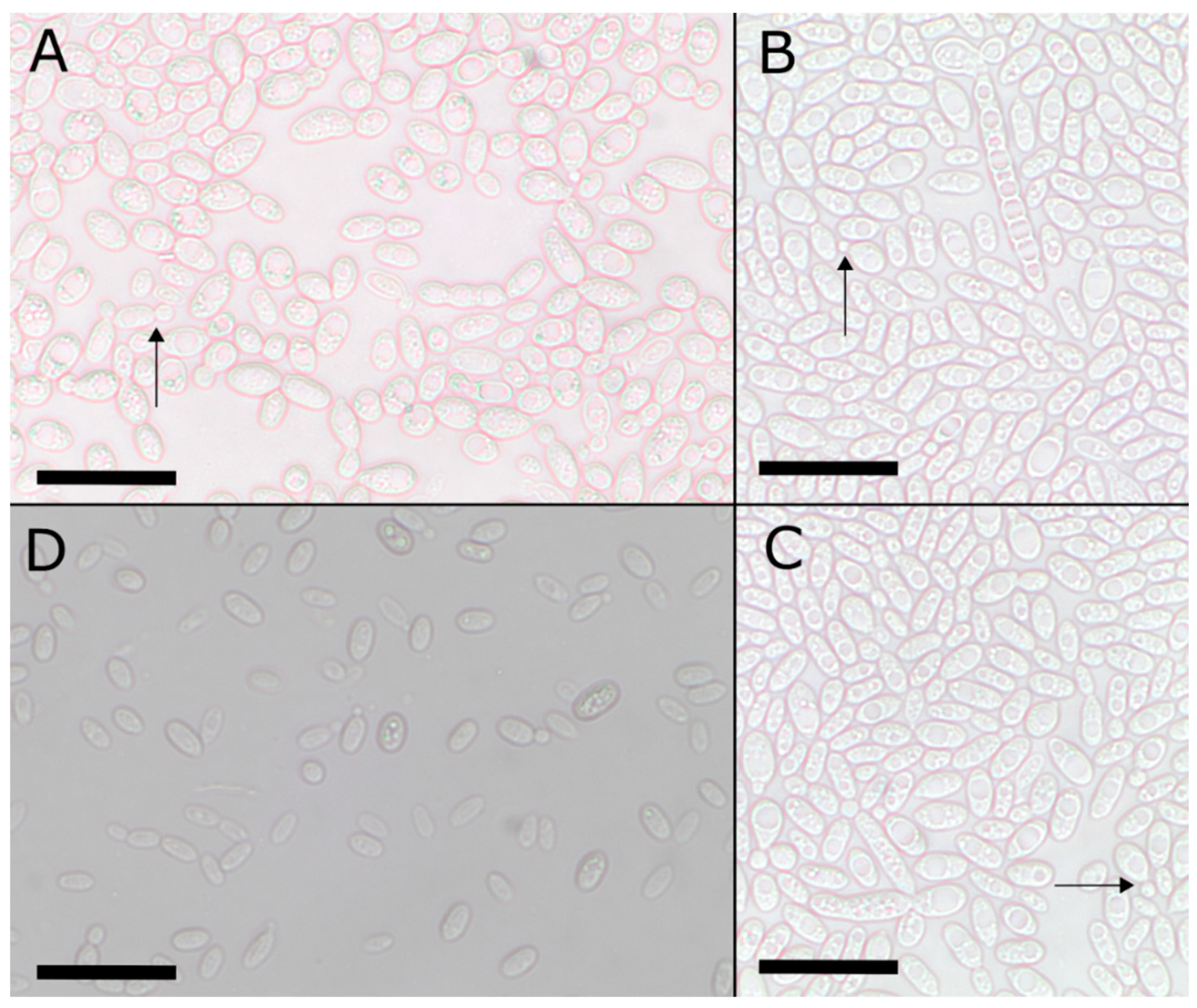
2.2. Morphological and Physiological Characterization
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Phylogenetic Inferences and Species Concepts
2.5. Environmental Determination of S. lactucae
3. Results
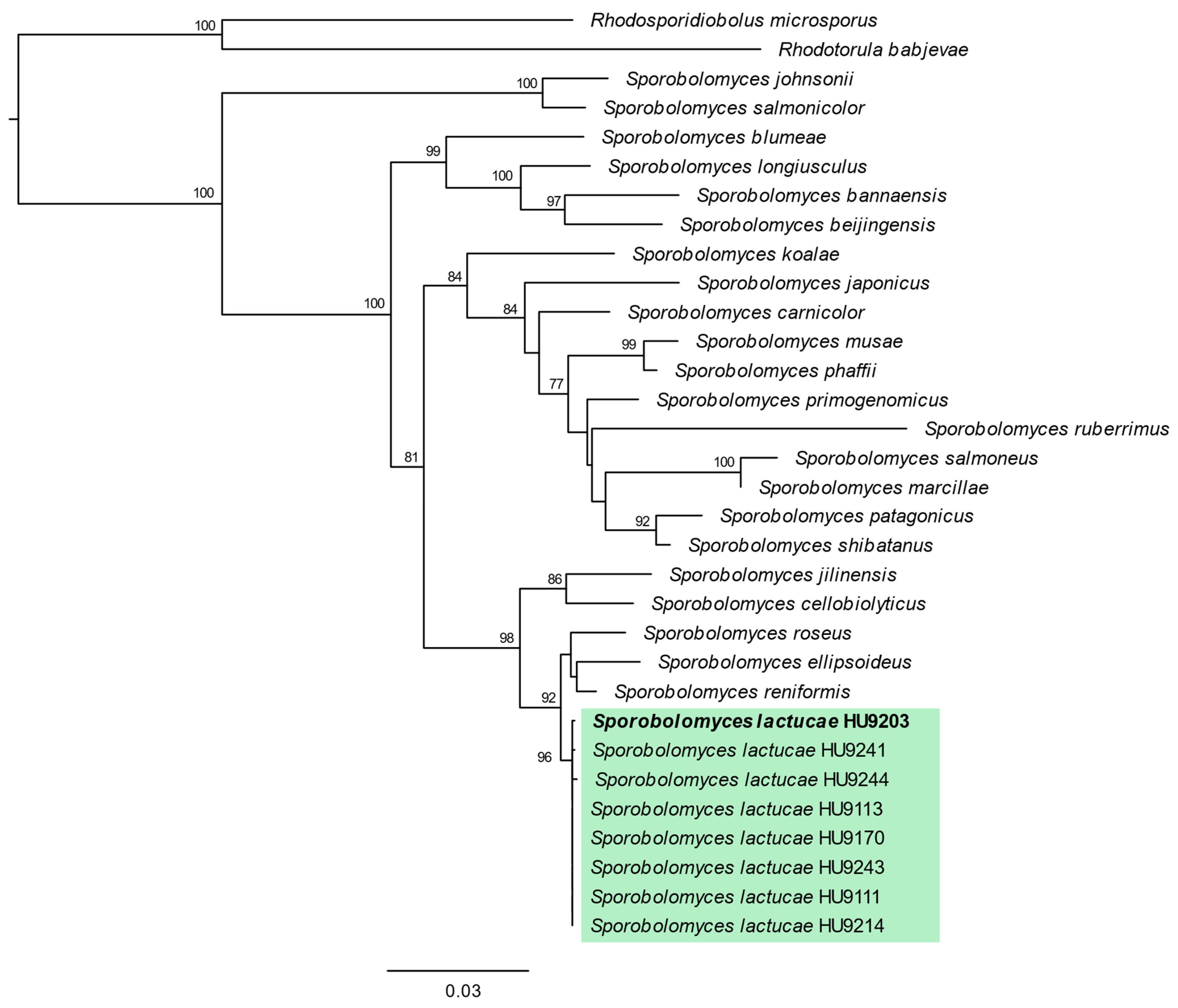
3.1. Phylogenetic Analyses
3.2. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kluyver, A.J.; van Niel, C.B. Über Spiegelbilder erzeugende Hefearten und die neue Hefegattung Sporobolomyces. Centbl. Bakt. ParasitKde 1924, 63, 19. [Google Scholar]
- Hamamoto, M.; Boekhout, T.; Nakase, T. Sporobolomyces Kluyver & van Niel (1924). In The Yeasts: A Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Burlington, MA, USA, 2011; Volume 3, pp. 1929–1990. [Google Scholar]
- Wang, Q.-M.; Yurkov, A.M.; Göker, M.; Lumbsch, H.T.; Leavitt, S.D.; Groenewald, M.; Theelen, B.; Liu, X.-Z.; Boekhout, T.; Bai, F.-Y. Phylogenetic classification of yeasts and related taxa within Pucciniomycotina. Stud. Mycol. 2015, 81, 149–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyland, G. Studies on Some Unusual Heterobasidiomycetes from Washington State. Mycologia 1949, 41, 686–701. [Google Scholar] [CrossRef]
- Valério, E.; Gadanho, M.; Sampaio, J.P. Reappraisal of the Sporobolomyces roseus species complex and description of Sporidiobolus metaroseus sp. nov. Int. J. Syst. Evol. Microbiol. 2008, 58, 736–741. [Google Scholar] [CrossRef] [Green Version]
- Aime, M.C.; Matheny, P.B.; Henk, D.A.; Frieders, E.M.; Nilsson, R.H.; Piepenbring, M.; McLaughlin, D.J.; Szabo, L.J.; Begerow, D.; Sampaio, J.P.; et al. An overview of the higher level classification of Pucciniomycotina based on combined analyses of nuclear large and small subunit rDNA sequences. Mycologia 2006, 98, 896–905. [Google Scholar] [CrossRef]
- Wang, Q.-M.; Groenewald, M.; Takashima, M.; Theelen, B.; Han, P.-J.; Liu, X.-Z.; Boekhout, T.; Bai, F.-Y. Phylogeny of yeasts and related filamentous fungi within Pucciniomycotina determined from multigene sequence analyses. Stud. Mycol. 2015, 81, 27–53. [Google Scholar] [CrossRef] [Green Version]
- Haelewaters, D.; Peterson, R.A.; Nevalainen, H.; Aime, M.C. Inopinatum lactosum gen. & comb. nov., the first yeast-like fungus in Leotiomycetes. Int. J. Syst. Evol. Microbiol. 2021, 71, 004862. [Google Scholar] [CrossRef]
- Aime, M.C.; Castlebury, L.A.; Abbasi, M.; Begerow, D.; Berndt, R.; Kirschner, R.; Marvanová, L.; Ono, Y.; Padamsee, M.; Scholler, M.; et al. Competing sexual and asexual generic names in Pucciniomycotina and Ustilaginomycotina (Basidiomycota) and recommendations for use. IMA Fungus 2018, 9, 75–89. [Google Scholar] [CrossRef]
- Lorenzini, M.; Zapparoli, G.; Azzolini, M.; Carvalho, C.; Sampaio, J.P. Sporobolomyces agrorum sp. nov. and Sporobolomyces sucorum sp. nov., two novel basidiomycetous yeast species isolated from grape and apple must in Italy. Int. J. Syst. Evol. Microbiol. 2019, 69, 3385–3391. [Google Scholar] [CrossRef]
- Li, A.-H.; Yuan, F.-X.; Groenewald, M.; Bensch, K.; Yurkov, A.M.; Li, K.; Han, P.-J.; Guo, L.-D.; Aime, M.C.; Sampaio, J.; et al. Diversity and phylogeny of basidiomycetous yeasts from plant leaves and soil: Proposal of two new orders, three new families, eight new genera and one hundred and seven new species. Stud. Mycol. 2020, 96, 17–140. [Google Scholar] [CrossRef]
- Urbina, H.; Aime, M.C. A closer look at Sporidiobolales: Ubiquitous microbial community members of plant and food biospheres. Mycologia 2018, 110, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. The Yeasts, a Taxonomic Study, 5th ed.; Elsevier: Burlington, MA, USA, 2011; Volume 3. [Google Scholar]
- Davoli, P.; Mierau, V.; Weber, R.W.S. Carotenoids and Fatty Acids in Red Yeasts Sporobolomyces roseus and Rhodotorula glutinis. Appl. Biochem. Microbiol. 2004, 40, 392–397. [Google Scholar] [CrossRef]
- Mannazzu, I.; Landolfo, S.; da Silva, T.L.; Buzzini, P. Red yeasts and carotenoid production: Outlining a future for non-conventional yeasts of biotechnological interest. World J. Microbiol. Biotechnol. 2015, 31, 1665–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, C.; Wu, T.; Ao, H.; Qian, X.; Wang, Z.; Sun, J. Increased torulene production by the red yeast, Sporidiobolus pararoseus, using citrus juice. Prep. Biochem. Biotechnol. 2020, 50, 66–73. [Google Scholar] [CrossRef] [PubMed]
- McCormack, P.J.; Wildman, H.G.; Jeffries, P. Production of antibacterial compounds by phylloplane-inhabiting yeasts and yeastlike fungi. Appl. Environ. Microbiol. 1994, 60, 927–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashi, E.; Fokkema, N.J. Environmental factors limiting growth of Sporobolomyces roseus, an antagonist of Cochliobolus sativus, on wheat leaves. Trans. Br. Mycol. Soc. 1977, 68, 17–25. [Google Scholar] [CrossRef]
- Di Menna, M.E. Yeasts from the leaves of pasture plants. N. Z. J. Agric. Res. 1959, 2, 394–405. [Google Scholar] [CrossRef]
- Hamamoto, M.; Nakase, T. Ballistosporous yeasts found on the surface of plant materials collected in New Zealand. 1. Six new species in the genus Sporobolomyces. Antonie Van Leeuwenhoek 1995, 67, 151–171. [Google Scholar] [CrossRef]
- Wang, Q.-M.; Bai, F.-Y. Four new yeast species of the genus Sporobolomyces from plant leaves. FEMS Yeast Res. 2004, 4, 579–586. [Google Scholar] [CrossRef] [Green Version]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The microbial ecology of wine grape berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef]
- Hunter, P.J.; Pink, D.A.C.; Bending, G.D. Cultivar-level genotype differences influence diversity and composition of lettuce (Lactuca sp.) phyllosphere fungal communities. Fungal Ecol. 2015, 17, 183–186. [Google Scholar] [CrossRef]
- Haelewaters, D.; Urbina, H.; Brown, S.; Newerth-Henson, S.; Aime, M. Isolation and Molecular Characterization of the Romaine Lettuce Phylloplane Mycobiome. J. Fungi 2021, 7, 277. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.H.; Harris, R.F. The Ecology and Biogeography of Microorganisms on Plant Surfaces. Annu. Rev. Phytopathol. 2000, 38, 145–180. [Google Scholar] [CrossRef]
- Nguyen-The, C.; Carlin, F. The microbiology of minimally processed fresh fruits and vegetables. Crit. Rev. Food Sci. Nutr. 1994, 34, 371–401. [Google Scholar] [CrossRef]
- Albu, S.; Toome, M.; Aime, M.C. Violaceomyces palustris gen. et sp. nov. and a new monotypic lineage, Violaceomycetales ord. nov. in Ustilaginomycetes. Mycologia 2015, 107, 1193–1204. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.-O.; Zhang, N.; Nguyen, N.; Gross, S.; Blackwell, M. Lab Manual for Yeast Study; Louisiana State University: Baton Rouge, LA, USA, 2008. [Google Scholar]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T.; Robert, V. Methods for Isolation, Phenotypic Characterization, and Maintenance of Yeasts. In The Yeasts: A Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Burlington, MA, USA, 2011; pp. 87–110. [Google Scholar]
- Online Auction Color Chart Co. The Online Auction Color Chart: The New Language of Color for Buyers and Sellers; Online Auction Color Chart Co.: Palo Alto, CA, USA, 2004. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phy-logenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Hopple, J.S., Jr. Phylogenetic Investigations in the Genus Coprinus Based on Morphological and Molecular Characters. Ph.D. Thesis, Duke University, Durham, NC, USA, 1994. [Google Scholar]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K.; Yokoyama, K.; Nishimura, K.; Miyaji, M. Molecular phylogenetics of the genus Rhodotorula and related basidiomycetous yeasts inferred from the mitochondrial cytochrome b gene. Int. J. Syst. Evol. Microbiol. 2001, 51, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- Toome, M.; Roberson, R.W.; Aime, M.C. Meredithblackwellia eburnean gen. et sp. nov., Kriegeriaceae fam. nov. and Kriegeriales ord. nov.—Toward resolving higher-level classification in Microbotryomycetes. Mycologia 2013, 105, 486–495. [Google Scholar] [CrossRef]
- Wang, Q.M.; Theelen, B.; Groenewald, M.; Bai, F.-Y.; Boekhout, T. Moniliellomycetes and Malasseziomycetes, two new classes in Ustilaginomycotina. Persoonia-Mol. Phylogeny Evol. Fungi 2014, 33, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.-M.; Bai, F.-Y. Molecular phylogeny of basidiomycetous yeasts in the Cryptococcus luteoluslineage (Tremellales) based on nuclear rRNA and mitochondrial cytochromebgene sequence analyses: Proposal of Derxomyces gen. nov. and Hannaella gen. nov., and description of eight novel Derxomyces species. FEMS Yeast Res. 2008, 8, 799–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.P.; Bishop-Hurley, S.L.; Marney, T.S.; Shivas, R.G. Nomenclatural novelties. Index Fungorum 2021, 503, 1–8. Available online: http://www.indexfungorum.org/Publications/Index%20Fungorum%20no.503.pdf (accessed on 27 January 2022).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; Institute of Electrical and Electronics Engineers: Piscataway, NJ, USA, 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding needles in haystacks: Linking scientific names, reference specimens and molecular data for Fungi. Database 2014, 2014, bau061. [Google Scholar] [CrossRef] [PubMed]
- Takashima, M.; Nakase, T. Four new species of the genus Sporobolomyces isolated from leaves in Thailand. Mycoscience 2000, 41, 357–369. [Google Scholar] [CrossRef]
- Bai, F.Y. Reclassification of the Sporobolomyces roseus and Sporidiobolus pararoseus complexes, with the description of Sporobolomyces phaffii sp. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 2309–2314. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.; Groenewald, M.; Szöke, S.; Cardinali, G.; Eberhardt, U.; Stielow, B.; de Vries, M.; Verkleij, G.J.M.; Crous, P.W.; Boekhout, T.; et al. DNA barcoding analysis of more than 9000 yeast isolates contributes to quantitative thresholds for yeast species and genera delimitation. Stud. Mycol. 2016, 85, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Fell, J.W.; Scorzetti, G.; Statzell-Tallman, A.; Pinel, N.; Yarrow, D. Recognition of the basidiomycetous yeast Sporobolomyces ruberrimus sp. nov. as a distinct species based on molecular and morphological analyses. FEMS Yeast Res. 2002, 1, 265–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, K.; Makimura, K. Sporobolomyces koalae sp. nov., a basidiomycetous yeast isolated from nasal smears of Queensland koalas kept in a Japanese zoological park. Int. J. Syst. Evol. Microbiol. 2008, 58, 2983–2986. [Google Scholar] [CrossRef] [PubMed]
- Libkind, D.; Gadanho, M.; Van Broock, M.; Sampaio, J.P. Sporidiobolus longiusculus sp. nov. and Sporobolomyces patagonicus sp. nov., novel yeasts of the Sporidiobolales isolated from aquatic environments in Patagonia, Argentina. Int. J. Syst. Evol. Microbiol. 2005, 55, 503–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maria, J.S. Sporobolomyces marcillae nov. spec., isolated from the air. Arch Mikrobiol. 1958, 32, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M. Phylogenetic Species Recognition and Species Concepts in Fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis, Version 3.61. 2019. Available online: https://www.mesquiteproject.org (accessed on 27 January 2022).
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; Von Haeseler, A.; Minh, B.Q. Terrace Aware Data Structure for Phylogenomic Inference from Supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- USDA. Agricultural Statistics 2020; National Agricultural Statistics Service: Washington, DC, USA. Available online: https://usda.library.cornell.edu/concern/publications/j3860694x?locale=en (accessed on 13 April 2021).
- FDA. Guide to Minimize Microbial Food Safety Hazards for Fresh Fruits and Vegetables; US Department of Health and Human Services: Washington, DC, USA, 1998.
- Moulas, C.; Petsoulas, C.; Rousidou, K.; Perruchon, C.; Karas, P.; Karpouzas, D.G. Effects of Systemic Pesticides Imidacloprid and Metalaxyl on the Phyllosphere of Pepper Plants. BioMed Res. Int. 2013, 2013, 969750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martos, F.; Munoz, F.; Pailler, T.; Kottke, I.; Gonneau, C.; Selosse, M.-A. The role of epiphytism in architecture and evolutionary constraint within mycorrhizal networks of tropical orchids. Mol. Ecol. 2012, 21, 5098–5109. [Google Scholar] [CrossRef] [PubMed]
- Setati, M.E.; Jacobson, D.; Andong, U.-C.; Bauer, F.F. The Vineyard Yeast Microbiome, a Mixed Model Microbial Map. PLoS ONE 2012, 7, e52609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourret, T.B.; Grove, G.G.; VanDeMark, G.J.; Henick-Kling, T.; Glawe, D.A. Diversity and molecular determination of wild yeasts in a central Washington State vineyard. N. Am. Fungi 2013, 8, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Jroundi, F.; González-Muñoz, M.T.; Sterflinger, K.; Piñar, G. Molecular Tools for Monitoring the Ecological Sustainability of a Stone Bio-Consolidation Treatment at the Royal Chapel, Granada. PLoS ONE 2015, 10, e0132465. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Authority | Strain | tef1 | LSU | ITS | cytb | Source |
|---|---|---|---|---|---|---|---|---|
| Rhodosporidiobolus | microsporus | (Higham ex Fell, Blatt, and Statzell) Q.M. Wang, F.Y. Bai, M. Groenew., and Boekhout 2015 | CBS 7041 T | KJ707817 | NG_042344 | NR_073290 | KJ707724 | [7,46] |
| Rhodotorula | babjevae | (Golubev) Q.M. Wang, F.Y. Bai, M. Groenew., and Boekhout 2015 | CBS 7808 T | – | NG_042339 | NR_077096 | – | [46] |
| Sporobolomyces | bannaensis | F.Y. Bai and J.H. Zhao 2003 | CBS 9204 T | KJ707934 | NG_068721 | NR_073345 | KJ707581 | [7,46] |
| Sporobolomyces | beijingensis | F.Y. Bai and Q.M. Wang 2004 | CGMCC 2.2365 T | KJ707919 | AY364837 | NR_137663 | KJ707588 | [7,21] |
| Sporobolomyces | blumeae | M. Takash. and Nakase 2000 | CBS 9094 T | KJ707926 | KY109742 | NR_137641 | KJ707673 | [7,47] |
| Sporobolomyces | carnicolor | Yamasaki and H. Fujii ex F.Y. Bai and Boekhout | CBS 4215 T | KJ707912 | NG_067316 | NR_137659 | KJ707707 | [7,48] |
| Sporobolomyces | cellobiolyticus | Q.M. Wang, F.Y. Bai and A.H. Li (2020) | CGMCC 2.5675 T | MK849110 | MK050406 | MK050406 | MK848982 | [11] |
| Sporobolomyces | ellipsoideus | Q.M. Wang, F.Y. Bai and A.H. Li (2020) | CGMCC 2.5619 T | MK849088 | MK050409 | MK050409 | MK848957 | [11] |
| Sporobolomyces | japonicus | Iizuka and Goto 1965 | CBS 5744 T | KJ707932 | KY109745 | NR_155844 | KJ707578 | [7,49] |
| Sporobolomyces | jilinensis | F.Y. Bai and Q.M. Wang 2004 | CGMCC 2.2301 T | KJ707913 | NG_068244 | NR_137664 | KJ707583 | [21] |
| Sporobolomyces | johnsonii | (Nyland) Q.M. Wang, F.Y. Bai, M. Groenew., and Boekhout 2015 | CBS 5470 T | KJ707931 | NG_042343 | NR_077090 | – | [7,50] |
| Sporobolomyces | koalae | Satoh and Makimura 2008 | CBS 10914 T | KJ707850 | NG_067317 | NR_137556 | KJ707604 | [7,51] |
| Sporobolomyces | lactucae | – | HU 9214 | MG589084 | MG588949 | MG470917 | MG589043 | [12] |
| Sporobolomyces | lactucae | Fatemi, Urbina, Haelew., and Aime 2022 | HU 9203 T | MG589082 | MG588947 | MG470912 | MG589041 | [12] |
| Sporobolomyces | lactucae | – | HU 9170 | MG589079 | MG588944 | MG470903 | MG589039 | [12] |
| Sporobolomyces | lactucae | – | HU 9113 | MG589077 | MG588942 | MG470889 | MG589037 | [12] |
| Sporobolomyces | lactucae | – | HU 9241 | MG589086 | MG588951 | MG470921 | MG589045 | [12] |
| Sporobolomyces | lactucae | – | HU 9243 | MG589087 | MG588952 | MG470922 | MG589046 | [12] |
| Sporobolomyces | lactucae | – | HU 9111 | MG589076 | MG588941 | MG470888 | MG589036 | [12] |
| Sporobolomyces | lactucae | – | HU 9244 | MG589088 | MG588953 | MG470923 | MG589047 | [12] |
| Sporobolomyces | longiusculus | (Libkind, Van Broock, and J.P. Samp.) Q.M. Wang, F.Y. Bai, M. Groenew., and Boekhout 2015 | PYCC 5818 T | KJ707929 | NG_068720 | NR_155773 | KJ707668 | [7,52] |
| Sporobolomyces | marcillae | Santa María 1958 | JCM 6883 T | KJ707933 | – | – | KJ707725 | [53] |
| Sporobolomyces | musae | Y.P. Tan, Marney, and R.G. Shivas 2021 | BRIP 28276 T | – | OK483137 | OK483138 | – | [41] |
| Sporobolomyces | patagonicus | Libkind, Van Broock, and J.P. Samp. 2005 | CBS 9657 T | KJ707928 | KY109759 | NR_137666 | KP216520 | [7,52] |
| Sporobolomyces | phaffii | F.Y. Bai, M. Takash., and Nakase 2002 | CGMCC 2.2137 T | KJ707918 | NG_068245 | NR_137660 | KJ707577 | [7,48] |
| Sporobolomyces | primogenomicus | Q.M. Wang and F.Y. Bai (2020) | JCM 8242 T | MK848998 | MK050417 | MK050417 | MK848872 | [11] |
| Sporobolomyces | reniformis | Q.M. Wang, F.Y. Bai, and A.H. Li (2020) | CGMCC 2.5627 T | MK849096 | MK050408 | MK050408 | MK848965 | [11] |
| Sporobolomyces | roseus | Kluyver and C.B. Niel 1924 | CBS 486 T | HM014022 | NG_069417 | NR_155845 | KJ707569 | D.A. Henk unpubl.; [7,49] |
| Sporobolomyces | ruberrimus | Yamasaki and H. Fujii ex Fell, Pinel, Scorzetti, Statzell, and Yarrow 2002 | CBS 7500 T | HM014017 | NG_067252 | NR_136959 | KJ707643 | D.A. Henk unpubl.; [50] |
| Sporobolomyces | salmoneus | Derx 1930 | CGMCC 2.2195 T | KJ707920 | KY109767 | KY105530 | KJ707580 | [7] |
| Sporobolomyces | salmonicolor | (B. Fisch. and Brebeck) Kluyver and C.B. Niel 1924 | JCM 1841 T | KJ707923 | NG_056268 | NR_149325 | KJ707701 | [7] |
| Sporobolomyces | shibatanus | (Okun.) Verona and Cif. | CBS 491 T | HM014019 | NG_067256 | NR_155770 | – | D.A. Henk unpubl.; [50] |
| Species | S. lactucae | S. jilinensis | S. roseus | |
|---|---|---|---|---|
| Type Strain | HU 9203 (CBS 16795) | CGMCC 2.2301 | CBS 486 | |
| Reference | This Paper | [21] | [13] | |
| CARBON ASSIMILATIONS | Glucose | + | + | + |
| Galactose | + (w, d) | + | + | |
| Sucrose | + | + | + | |
| Maltose | + | + | + | |
| Cellobiose | + (w, d) | - | + | |
| Lactose | - | - | - | |
| Inulin | - | - | - | |
| Soluble starch | + (w) | + | + | |
| myo-Inositol | - | - | - | |
| Glucono-1,5-lactone | + (w) | n/a | n/a | |
| Glucuronate | + | n/a | - | |
| Galacturonic Acid | (w) | n/a | - | |
| Lactate | - | -(as DL-lactic acid) | + | |
| Citrate | - | -(as citric acid) | + | |
| Methanol | - | - | - | |
| Ethanol | (w) | + | + | |
| NITROGEN ASSIMILATIONS | Propane-1,2-diol | (w) | n/a | n/a |
| K Nitrate | + | + | + | |
| Na Nitrite | + | + | + | |
| Ethylamine | (w) | - | n/a | |
| L-lysine | + | + | n/a | |
| Cadaverine | (w) | + | n/a | |
| Creatine | (w) | n/a | n/a | |
| Creatinine | (w) | n/a | n/a | |
| D-glucosamine | + | - | - | |
| Imidazole | - | n/a | n/a | |
| D-tryptophan | (w) | n/a | n/a | |
| OTHER | 10% (w/v) NaCl | (w) | n/a | n/a |
| 16% (w/v) NaCl | (w) | n/a | n/a | |
| 50% (w/v) Glucose | + | - | v | |
| 60% (w/v) Glucose | + | n/a | n/a |
| Accession Number | Genbank Identification | Strain Identification | Our Identification | Percent Identification | Locality | Substrate | Reference |
|---|---|---|---|---|---|---|---|
| AY070006 | Sporobolomyces sp. | AS 2.2108 | Sporobolomyces lactucae | 99.82% | Yunnan, China | wilting leaf of Parthenocissus sp. | [48] |
| HF947090 | Sporobolomyces sp. (as Sporidiobolus sp.) | – | Sporobolomyces lactucae | 99.65% | Greece | phylloplane of Capsicum annuum | [62] |
| JF691061 | Atractiellales | – | Sporobolomyces lactucae | 99.46% | Réunion Island | orchid roots | [63] |
| JQ425363 | Sporobolomyces sp. (as Sporidiobolus sp.) | JPS-2007a | Sporobolomyces lactucae | 99.65% | Egypt | air, grapevine plantation | Z.S.M. Soliman unpubl. |
| JQ993369 | Sporobolomyces roseus | IWBT-Y808 | Sporobolomyces lactucae | 99.47% | South Africa | wine grape berries | [64] |
| JX188234 | Sporobolomyces sp. (as Sporidiobolus sp.) | JPS-2007a | Sporobolomyces lactucae | 99.82% | Pullman, WA, USA | on Vitis vinifera | [65] |
| KM062084 | Sporobolomyces sp. | 2H-7 | Sporobolomyces lactucae | 99.65% | Granada | stone (biotreated) | [66] |
| KU168778 | Sporobolomyces roseus (as Sporidiobolus metaroseus) | T11-22 | Sporobolomyces lactucae | 99.82% | Antarctica | rock | S. Barahona et al. unpubl. |
| KX376263 | Sporobolomyces roseus (as Sporidiobolus metaroseus) | AUMC 10722 | Sporobolomyces lactucae | 99.65% | Egypt | yogurt | Z.S.M. Soliman unpubl. |
| KY105475 | Sporobolomyces roseus (as Sporidiobolus metaroseus) | CBS 10225 | Sporobolomyces lactucae | 99.82% | Portugal | plant | [49] |
| KY495743 | Sporobolomyces roseus | AUMC 10775 | Sporobolomyces lactucae | 99.12% | Egypt | strawberry juice | Z.S.M. Soliman unpubl. |
| KY495777 | Sporobolomyces roseus | AUMC 11209 | Sporobolomyces lactucae | 99.82% | Egypt | guava juice | Z.S.M. Soliman unpubl. |
| KY611818 | Sporobolomyces roseus | AUMC 11213 | Sporobolomyces lactucae | 99.47% | Egypt | flower, chamomile | Z.S.M. Soliman unpubl. |
| KY611834 | Sporobolomyces roseus | AUMC 11233 | Sporobolomyces lactucae | 99.65% | Egypt | flower, mango (Mangifera indica) | Z.S.M. Soliman unpubl. |
| MF071283 | Sporobolomyces roseus (as Sporidiobolus metaroseus) | AUMC 11218 | Sporobolomyces lactucae | 99.64% | Egypt | flower of Rosaceae plant | Z.S.M. Soliman unpubl. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fatemi, S.; Haelewaters, D.; Urbina, H.; Brown, S.; Houston, M.L.; Aime, M.C. Sporobolomyces lactucae sp. nov. (Pucciniomycotina, Microbotryomycetes, Sporidiobolales): An Abundant Component of Romaine Lettuce Phylloplanes. J. Fungi 2022, 8, 302. https://doi.org/10.3390/jof8030302
Fatemi S, Haelewaters D, Urbina H, Brown S, Houston ML, Aime MC. Sporobolomyces lactucae sp. nov. (Pucciniomycotina, Microbotryomycetes, Sporidiobolales): An Abundant Component of Romaine Lettuce Phylloplanes. Journal of Fungi. 2022; 8(3):302. https://doi.org/10.3390/jof8030302
Chicago/Turabian StyleFatemi, Samira, Danny Haelewaters, Hector Urbina, Samuel Brown, Makenna L. Houston, and M. Catherine Aime. 2022. "Sporobolomyces lactucae sp. nov. (Pucciniomycotina, Microbotryomycetes, Sporidiobolales): An Abundant Component of Romaine Lettuce Phylloplanes" Journal of Fungi 8, no. 3: 302. https://doi.org/10.3390/jof8030302