
Stilbocrea banihashemiana sp. nov. a New Fungal Pathogen Causing Stem Cankers and Twig Dieback of Fruit Trees


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Fungal Isolation
2.2. Morphological Characterization
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Phylogenetic Analyses
2.5. Pathogenicity Tests
3. Results
3.1. Field Surveys and Disease Symptoms
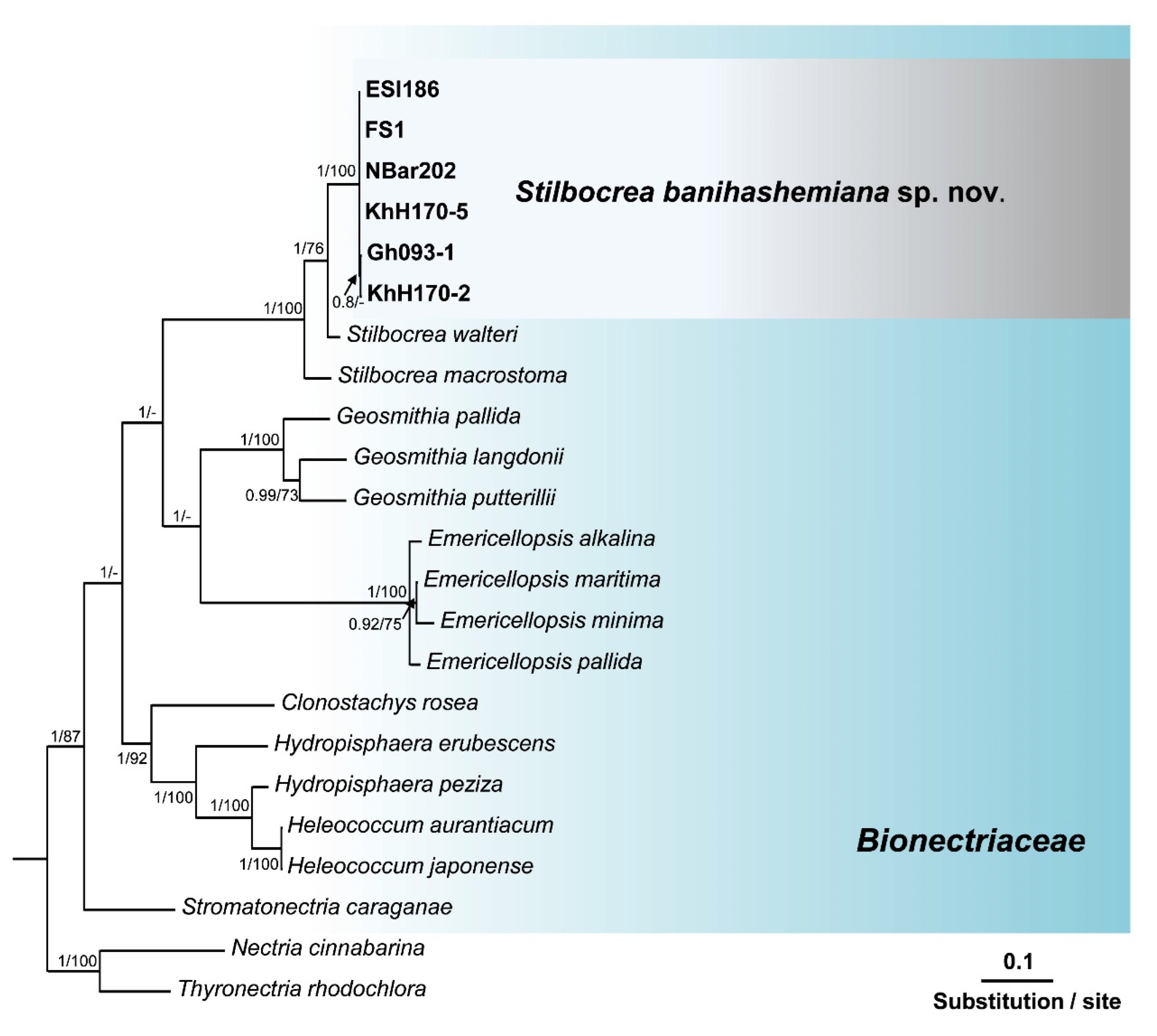
3.2. Molecular Identification and Phylogenetic Analyses
Taxonomy
3.3. Pathogenicity Tests
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jafari, M.; Rahemi, M.; Kamgar Haghighi, A.A. Role of fig rootstock on changes of water status and nutrient concentrations in ‘Sabz’ cultivar under drought stress condition. Sci. Hortic. 2018, 230, 56–61. [Google Scholar] [CrossRef]
- Jokar, A.; Zare, H.; Zakerin, A.; Aboutalebi Jahromi, A. Effects of shade net colors on mineral elements and postharvest shelf life and quality of fresh fig (Ficus carica L.) under rain-fed condition. Horticulturae 2021, 7, 93. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Organization of the United Nations, Statistical Databases. Available online: http://www.fao.org (accessed on 12 December 2021).
- Ahmadi, K.; Gholizadeh, H.; Ebadzadeh, H.R.; Hatami, F.; Hoseinpour, R.; Kazemifard, R.; Abdeshah, H. Agricultural Statistics; Horticultural Products; Ministry of Agriculture Jihad, Department of Planning and Economy, Center for Information and Communication Technology: Tehran, Iran, 2016; p. 3. [Google Scholar]
- Banihashemi, Z.; Javadi, A.R. Further investigations on the biology of Phomopsis cinerascens, the cause of fig canker in Iran. Phytopathol. Mediterr. 2009, 48, 454–460. [Google Scholar]
- Çeliker, N.M.; Michailides, T.J. First report of Lasiodiplodia theobromae causing canker and shoot blight of fig in Turkey. New Dis. Rep. 2012, 25, 12. [Google Scholar] [CrossRef]
- Giambra, S.; Piazza, G.; Alves, A.; Mondello, V.; Berbegal, M.; Armengol, J.; Burruano, S. Botryosphaeriaceae species associated with diseased loquat trees in Italy and description of Diplodia rosacearum sp. nov. Mycosphere 2017, 7, 978–989. [Google Scholar] [CrossRef]
- Gonzalez-Dominguez, E.; Alves, A.; León, M.; Armengol, J. Characterization of Botryosphaeriaceae species associated with diseased loquat (Eriobotrya japonica) in Spain. Plant Pathol. 2017, 66, 77–89. [Google Scholar] [CrossRef]
- Fahim Abbas, M.; Batool, S.; Khaliq, S.; Mubeen, S.; Azziz-ud-Din; Ullah, N.; Zafar, K.; Rafiq, M.; Al-Sadi, A.M.; Alotaibi, S.S.; et al. Diversity of fungal pathogens associated with loquat and development of novel virulence scales. PLoS ONE 2021, 16, e0257951. [Google Scholar] [CrossRef]
- Kajitani, Y.; Masuya, H. Ceratocystis ficicola sp. nov., a causal fungus of fig canker in Japan. Mycoscience 2011, 52, 349–353. [Google Scholar] [CrossRef]
- Alfieri, S.A., Jr.; Schoulties, C.L.; El-Gholl, N.E. Nectriella (Kutilakesa) pironii, a pathogen of ornamental plants. Annu. Meet. Fla. State Hortic. Soc. 1980, 93, 218–219. [Google Scholar]
- Fisher, J.R.; McCann, D.P.; Taylor, N.J. Geosmithia morbida, thousand cankers disease of black walnut pathogen, was found for the first time in southwestern Ohio. Plant Health Prog. 2013, 14, 36. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, D.; Moricca, S.; Bracalini, M.; Benigno, A.; Bernardo, U.; Luchi, N.; Da Lio, D.; Nugnes, F.; Cappellini, G.; Salemi, C.; et al. Rapid detection of Pityophthorus juglandis (Blackman) (Coleoptera, Curculionidae) with the loop-mediated isothermal amplification (LAMP). Method. Plants 2021, 10, 1048. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, D.; Aglietti, C.; Benigno, A.; Bracalini, M.; Da Lio, D.; Bartolini, L.; Cappellini, G.; Aronadio, A.; Francia, C.; Luchi, N.; et al. Loop-mediated isothermal amplification (LAMP) and SYBR Green qPCR for fast and reliable detection of Geosmithia morbida (Kolařik) in infected walnut. Plants 2022, 11, 1239. [Google Scholar] [CrossRef] [PubMed]
- Lechat, C.; Fournier, J. New insights into Stilbocrea (Hypocreales, Bionectriaceae): Recognition of S. colubrensis, a new species from Martinique (French West Indies), and observations on lifestyle and synnematous asexual more. Ascomycete.org 2019, 11, 183–190. [Google Scholar] [CrossRef]
- Voglmayr, H.; Jaklitsch, W.M. Stilbocrea walteri sp. nov., an unusual species of Bionectriaceae. Mycol. Prog. 2019, 18, 91–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esparham, N.; Mohammadi, H.; Gramaje, D. A survey of trunk disease pathogens within citrus trees in Iran. Plants 2020, 9, 754. [Google Scholar] [CrossRef]
- Di Lecce, R.; Bashiri, S.; Masi, M.; Alioto, D.; Tuzi, A.; Abdollahzadeh, J.; Evidente, A. Phytotoxic metabolites from Stilbocrea macrostoma, a fungal pathogen of Quercus brantii in Iran. Nat. Prod. Res. 2021, 35, 5857–5861. [Google Scholar] [CrossRef]
- Mirsoleimani, Z.; Mostowfizadeh-Ghalamfarsa, R. Characterization of Phytophthora pistaciae, the causal agent of pistachio gummosis, based on host range, morphology, and ribosomal genome. Phytopathol. Mediterr. 2013, 52, 501–516. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets forspeciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E.A. Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- BAST. Available online: https://www.blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 10 January 2021).
- GenBank. Available online: http://www.ncbi.nlm.nih.gov/genbank (accessed on 10 January 2022).
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; pp. 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, M.P. PAUP*: Phylogenetic Analysis Using Parsimony (*and other methods). J. Bioinform. Comput. Biol. 2004, 1. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J.A.; Ronquist, F.; Huelsenbeck, J.P.; Nieves-Aldrey, J. Bayesian phylogenetic analysis of combined data. Syst. Biol. 2004, 53, 47–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baum, B.R. PHYLIP (phylogeny inference package), version 3.5 c. Q. Rev. Biol. 1998, 64, 539–541. [Google Scholar] [CrossRef]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- TreeBASE. Available online: http://www.treebase.org (accessed on 14 January 2022).
- Roux, J.; Eisenberg, B.; Kanzler, A.; Nel, A.; Coetzee, V.; Kietzka, E.; Wingfield, M.J. Testing of selected South African Pinus hybrids and families for tolerance to the pitch canker pathogen, Fusarium circinatum. New For. 2007, 33, 109–123. [Google Scholar] [CrossRef]
- Sakayaroj, J.; Preedanon, S.; Supaphon, O.; Jones, E.G.; Phongpaichit, S. Phylogenetic diversity of endophyte assemblages associated with the tropical seagrass Enhalus acoroides in Thailand. Fungal Divers. 2010, 42, 27–45. [Google Scholar] [CrossRef]
- Sung, G.H.; Poinar, G.O., Jr.; Spatafora, J.W. The oldest fossil evidence of animal parasitism by fungi supports a Cretaceous diversification of fungal–arthropod symbioses. Mol. Phylogenet. Evol. 2008, 49, 495–502. [Google Scholar] [CrossRef]
- McKnight, K.H.; Rayner, R.W. A mycological colour chart. Mycologia 1972, 64, 230–233. [Google Scholar] [CrossRef]
- Elshafie, A.E.; Ba-Omar, T. First report of Albizia lebbeck dieback caused by Scytalidium dimidiatum in Oman. Mycopathologia 2002, 154, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.D.; Burgess, T.; Lanoiselet, V.M. First record of Neoscytalidium dimidiatum and N. novaehollandiae on Mangifera indica and N. dimidiatum on Ficus carica in Australia. Australas. Plant Dis. Notes 2010, 5, 48–50. [Google Scholar] [CrossRef] [Green Version]
- Al-Bedak, O.A.; Mohamed, R.A.; Seddek, N.H. First detection of Neoscytalidium dimidiatum associated with canker disease in Egyptian Ficus trees. For. Pathol. 2018, 48, e12411. [Google Scholar] [CrossRef]
- Aloi, F.; Riolo, M.; Parlascino, R.; Pane, A.; Cacciola, S.O. Bot gummosis of lemon (Citrus × limon) caused by Neofusicoccum parvum. J. Fungi 2021, 7, 294. [Google Scholar] [CrossRef]
- Yeganeh, S.; Mohammadi, H. Sooty canker, a destructive disease of banyan (Ficus benghalensis L.) trees in landscapes of Kish Island (Iran). Urban For. Urban Green. 2022, 72, 127573. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Leavitt, G.M.; Guerrero, J.C.; Guevara, J.; Gubler, W.D. Identification and pathogenicity of Lasiodiplodia theobromae and Diplodia seriata, the causal agents of bot canker disease of grapevines in Mexico. Plant Dis. 2008, 92, 519–529. [Google Scholar] [CrossRef] [Green Version]
- Tavakolian, B. Etiology of Loquat Canker in Fars Province. Master’s Thesis, Shiraz University, Shiraz, Iran, 2021. [Google Scholar]
- Hirooka, Y.; Rossman, A.Y.; Samuels, G.J.; Lechat, C.; Chaverri, P. A monograph of Allantonectria, Nectria, and Pleonectria (Nectriaceae, Hypocreales, Ascomycota) and their pycnidial, sporodochial, and synnematous anamorphs. Stud. Mycol. 2012, 71, 1–210. [Google Scholar] [CrossRef] [Green Version]
- Summerbell, R.C.; Gueidan, C.; Schroers, H.J.; de Hoog, G.S.; Starink, M.; Rosete, Y.A.; Guarro, J.; Scott, J.A. Acremonium phylogenetic overview and revision of Gliomastix, Sarocladium, and Trichothecium. Stud. Mycol. 2011, 68, 139–162. [Google Scholar] [CrossRef]
- Mugnai, L.; Graniti, A.; Surico, G. Esca (black measles) and brownwood-streaking: Two old and elusive diseases of grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar] [CrossRef] [Green Version]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine trunk diseases: A review of fifteen years of trials for their control with chemicals and biocontrol agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef] [Green Version]
- Cacciola, S.O.; Gullino, M.L. Emerging and re-emerging fungus and oomycete soil-borne plant diseases in Italy. Phytopathol. Mediterr. 2019, 58, 451–472. [Google Scholar] [CrossRef]
- Aloi, F.; Giambra, S.; Schena, L.; Surico, G.; Pane, A.; Gusella, G.; Stracquadanio, C.; Burruano, S.; Cacciola, S.O. New insights into scabby canker of Opuntia ficus-indica, caused by Neofusicoccum batangarum. Phytopathol. Mediterr. 2020, 59, 269–284. [Google Scholar] [CrossRef]
- Mostowfizadeh-Ghalamfarsa, R.; Salmaninezhad, F. Early detection of fungal plant pathogen invasions in natural ecosystems based on bridge-head effect with the emphasis on Phytophthora species. Strateg. Res. J. Agric. Sci. Nat. Resour. 2020, 5, 81–96. [Google Scholar]
- Kamali-Sarvestani, S.; Mostowfizadeh-Ghalamfarsa, R.; Salmaninezhad, F.; Cacciola, S.O. Fusarium and Neocosmospora species associated with rot of Cactaceae and other succulent plants. J. Fungi 2022, 8, 364. [Google Scholar] [CrossRef]
- Roccotelli, A.; Schena, L.; Sanzani, S.M.; Cacciola, S.O.; Mosca, S.; Faedda, R.; Ippolito, A.; di San Lio, G.M. Characterization of basidiomycetes associated with wood rot of citrus in southern Italy. Phytopathology 2014, 104, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Agustí-Brisach, C.; Moldero, D.; Raya, M.D.C.; Lorite, I.J.; Orgaz, F.; Trapero, A. Water stress enhances the progression of branch dieback and almond decline under field conditions. Plants 2020, 9, 1213. [Google Scholar] [CrossRef]
- Luo, Y.; Ma, R.; Barrera, E.; Gusella, G.; Michailides, T. Effects of temperature on development of canker-causing pathogens in almond and prune. Plant Dis. 2022, 107. [Google Scholar] [CrossRef]
- Schoeneweiss, D.F. Role of environmental stress in diseases of woody plants. Plant Dis. 1981, 65, 308–314. [Google Scholar] [CrossRef]
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.W.; Hesse, C.N.; et al. Botryosphaeria dothidea: A latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef]
- Riolo, M.; Aloi, F.; Pane, A.; Cara, M.; Cacciola, S.O. Twig and shoot dieback of citrus, a new disease caused by Colletotrichum species. Cells 2021, 10, 449. [Google Scholar] [CrossRef]
- Lade, S.B.; Štraus, D.; Oliva, J. Variation in fungal community in grapevine (Vitis vinifera) nursery stock depends on nursery, variety and rootstock. J. Fungi 2022, 8, 47. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene 1 | Number of Cycles | Initial Denaturation | Denaturation | Annealing | Extension | Final Extension |
|---|---|---|---|---|---|---|
| ITS | 30 | 95 (120) 2 | 95 (45) | 64 (45) | 72 (45) | 72 (600) |
| tef1 | 35 | 95 (120) | 95 (120) | 65 (60) | 72 (50) | 72 (600) |
| rpb2 | 35 | 95 (120) | 95 (120) | 66.5 (60) 3 | 72 (45) | 72 (600) |
| Isolate | Location | Host | Date of Sampling | Latitude | Longitude |
|---|---|---|---|---|---|
| ES184-1 | Estahban | Ficus carica cv. Sabz | 23 October 2019 | 29°06′.794″ N | 54°04′.533″ E |
| ES184-2 | Estahban | F. carica cv. Sabz | 23 October 2019 | 29°06′.794″ N | 54°04′.533″ E |
| ES184-3 | Estahban | F. carica cv Sabz | 23 October 2019 | 29°06′.794″ N | 54°04′.533″ E |
| ESH185-1 | Estahban | F. carica cv. Shah Anjeer | 23 October 2019 | 29°06′.852″ N | 54°04′.596″ E |
| ESH185-2 | Estahban | F. carica cv. Shah Anjeer | 23 October 2019 | 29°06′.852″ N | 54°04′.596″ E |
| ESi186-1 | Estahban | F. carica cv. Siah | 23 October 2019 | 29°06′.873″ N | 54°04′.563″ E |
| ESi186-2 | Estahban | F. carica cv. Siah | 23 October 2019 | 29°06′.873″ N | 54°04′.563″ E |
| ESi187-1 | Estahban | F. carica cv. Siah | 23 October 2019 | 29°06′.873″ N | 54°04′.584″ E |
| ESi187-2 | Estahban | F. carica cv. Siah | 23 October 2019 | 29°06′.873″ N | 54°04′.584″ E |
| ESi189-1 | Estahban | F. carica cv. Siah | 23 October 2019 | 29°06′.856″ N | 54°04′.583″ E |
| ESH191A-1 | Estahban | F. carica cv. Shah Anjeer | 23 October 2019 | 29°06′.852″ N | 54°04′.567″ E |
| ESH191A-2 | Estahban | F. carica cv. Shah Anjeer | 23 October 2019 | 29°06′.852″ N | 54°04′.567″ E |
| NS195-1 | Neyriz | F. carica cv. Sabz | 23 October 2019 | 29°08′.777″ N | 54°17′.480″ E |
| NS198-1 | Neyriz | F. carica cv. Sabz | 23 October 2019 | 29°08′.760″ N | 54°17′.514″ E |
| NS198-2 | Neyriz | F. carica cv. Sabz | 23 October 2019 | 29°08′.760″ N | 54°17′.514″ E |
| NS199-1 | Neyriz | F. carica cv. Sabz | 23 October 2019 | 29°08′.755″ N | 54°17′.500″ E |
| NS199-2 | Neyriz | F. carica cv. Sabz | 23 October 2019 | 29°08′.755″ N | 54°17′.503″ E |
| NS200-1 | Neyriz | F. carica cv. Sabz | 23 October 2019 | 29°08′.747″ N | 54°17′.503″ E |
| NS200-2 | Neyriz | F. carica cv. Sabz | 23 October 2019 | 29°08′.747″ N | 54°17′.503″ E |
| NS201-1 | Neyriz | F. carica cv. Sabz | 23 October 2019 | 29°08′.730″ N | 54°17′.512″ E |
| NS201-2 | Neyriz | F. carica cv. Sabz | 23 October 2019 | 29°08′.730″ N | 54°17′.512″ E |
| NS202 | Neyriz | F. carica cv. Sabz | 23 October 2019 | 29°08′.759″ N | 54°17′.470″ E |
| NBar202-1 | Neyriz | F. carica cv. Puzdonbali | 23 October 2019 | 29°08′.777″ N | 54°17′.480″ E |
| NBar202-2 | Neyriz | F. carica cv. Puzdonbali | 23 October 2019 | 29°08′.777″ N | 54°17′.480″ E |
| NSiDrj-1 | Neyriz | F. carica cv. Siah | 18 November 2020 | 29°08′.777″ N | 54°17′.480″ E |
| ESi217-1 | Estahban | F. carica cv. Siah | 18 November 2020 | 29°06′.794″ N | 54°04′.471″ E |
| ECH218-1 | Estahban | F. carica cv. Barg Chenary | 18 November 2020 | 29°06′.858″ N | 54°04′.549″ E |
| ECH218-2 | Estahban | F. carica cv. Barg Chenary | 18 November 2020 | 29°06′.858″ N | 54°04′.549″ E |
| KDS22-3 | Kazerun | F. carica cv. Sabz | 8 January2019 | 29°49′.482″ N | 51°47′.688″ E |
| KDS25-8 | Kazerun | F. carica cv. Sabz | 1 January2019 | 29°49′.503″ N | 51°47′.633″ E |
| FS1 * | Firuzabad | F. carica cv. Sabz | 18 November 2020 | 28°49′.198″ N | 52°33′.396″ E |
| FS2 | Firuzabad | F. cacrica cv. Sabz | 18 November 2020 | 28°49′.198″ N | 52°33′.396″ E |
| DMS1 | Darab | F. carica cv. Sabz | 3 March 2021 | 29°56′.446″ N | 53°18′.129″ E |
| DMS2 | Darab | F. carica cv. Sabz | 3 March 2021 | 29°56′.446″ N | 53°18′.129″ E |
| Gh093-1 | Shiraz- | Eryobotria japonica | 1 October 2019 | 29°40′.582″ N | 52°28′.553″ E |
| KhH170-1 | Khafr | E. japonica | 1 October 2019 | 28°59′.053″ N | 53°12′.299″ E |
| KhH170-2 | Khafr | E. japonica | 1 October 2019 | 28°59′.053″ N | 53°12′.299″ E |
| KhH170-3 | Khafr | E. japonica | 1 October 2019 | 28°59′.053″ N | 53°12′.299″ E |
| KhH170-4 | Khafr | E. japonica | 1 October 2019 | 28°59′.053″ N | 53°12′.299″ E |
| KhH170-5 | Khafr | E. japonica | 1 October 2019 | 28°59′.053″ N | 53°12′.299″ E |
| KhH170-6 | Khafr | E. japonica | 1 October 2019 | 28°59′.053″ N | 53°12′.299″ E |
| KhH170-7 | Khafr | E. japonica | 1 October 2019 | 28°59′.053″ N | 53°12′.299″ E |
| Isolate | GenBank Accession Number | Isolate | GenBank Accession Number | ||||
|---|---|---|---|---|---|---|---|
| ITS | tef1 | rpb2 | ITS | tef1 | rpb2 | ||
| ESH185-1 | OM615396 | N/A | OM876870 | KDS25-8 | N/A | N/A | OM876873 |
| ESi186-1 | OM615398 | OM876866 | OM876871 | FS1 * | OM615399 | OM876865 | OM876872 |
| ESH191A-1 | OM615397 | N/A | N/A | Gh093-1 | OM615379 | OM876867 | OM908433 |
| NS199-1 | OM615401 | N/A | N/A | KhH170-2 | OM615380 | OM876868 | OM908434 |
| NBar202-1 | OM615400 | OM876864 | OM876874 | KhH170-5 | OM615381 | OM876869 | OM908435 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolboli, Z.; Tavakolian, B.; Mostowfizadeh-Ghalamfarsa, R.; Jafari, M.; Cacciola, S.O. Stilbocrea banihashemiana sp. nov. a New Fungal Pathogen Causing Stem Cankers and Twig Dieback of Fruit Trees. J. Fungi 2022, 8, 694. https://doi.org/10.3390/jof8070694
Bolboli Z, Tavakolian B, Mostowfizadeh-Ghalamfarsa R, Jafari M, Cacciola SO. Stilbocrea banihashemiana sp. nov. a New Fungal Pathogen Causing Stem Cankers and Twig Dieback of Fruit Trees. Journal of Fungi. 2022; 8(7):694. https://doi.org/10.3390/jof8070694
Chicago/Turabian StyleBolboli, Zeinab, Behnaz Tavakolian, Reza Mostowfizadeh-Ghalamfarsa, Moslem Jafari, and Santa Olga Cacciola. 2022. "Stilbocrea banihashemiana sp. nov. a New Fungal Pathogen Causing Stem Cankers and Twig Dieback of Fruit Trees" Journal of Fungi 8, no. 7: 694. https://doi.org/10.3390/jof8070694