
Species Diversity, Distribution, and Phylogeny of Exophiala with the Addition of Four New Species from Thailand

 ,
,  , and
, and 
Abstract
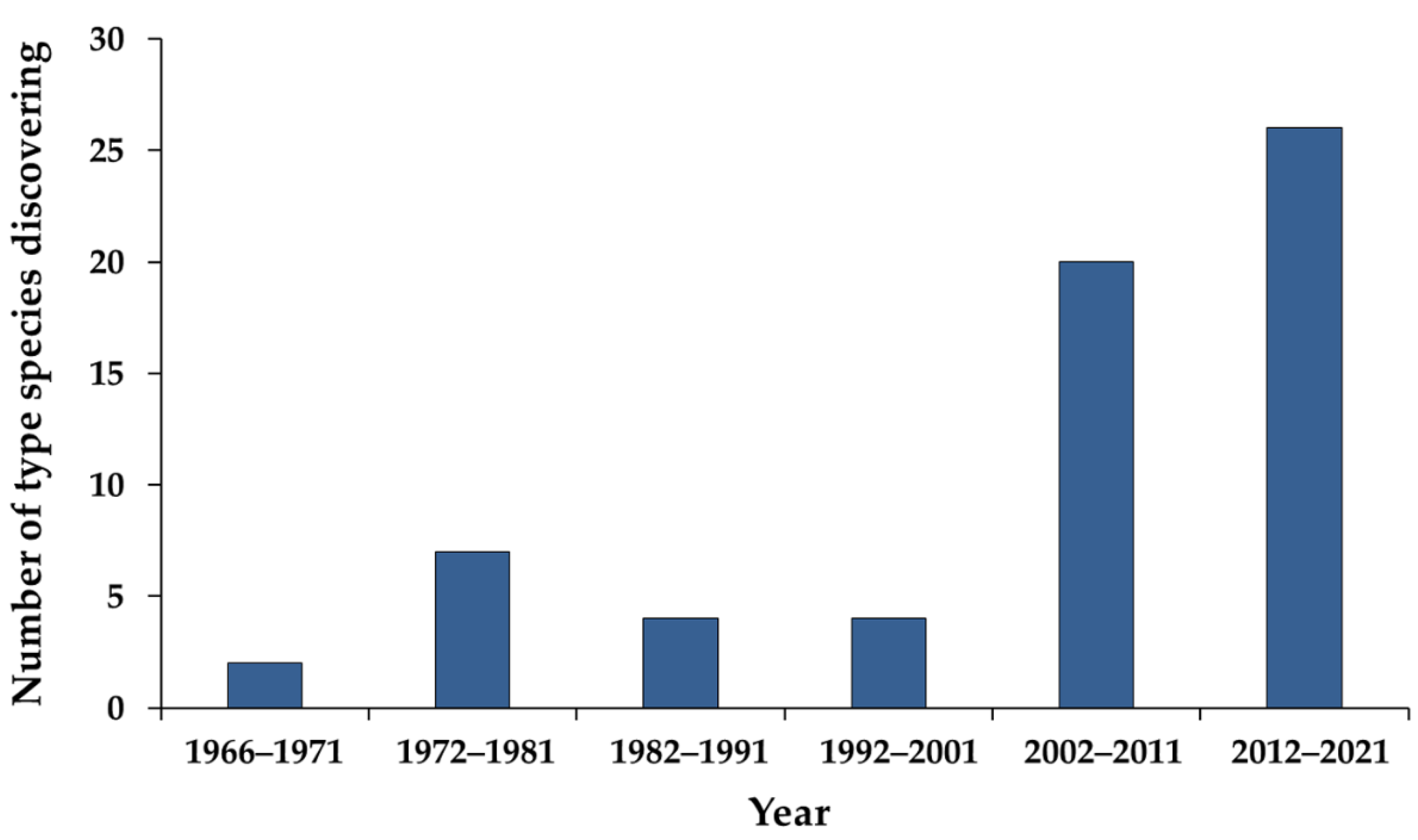
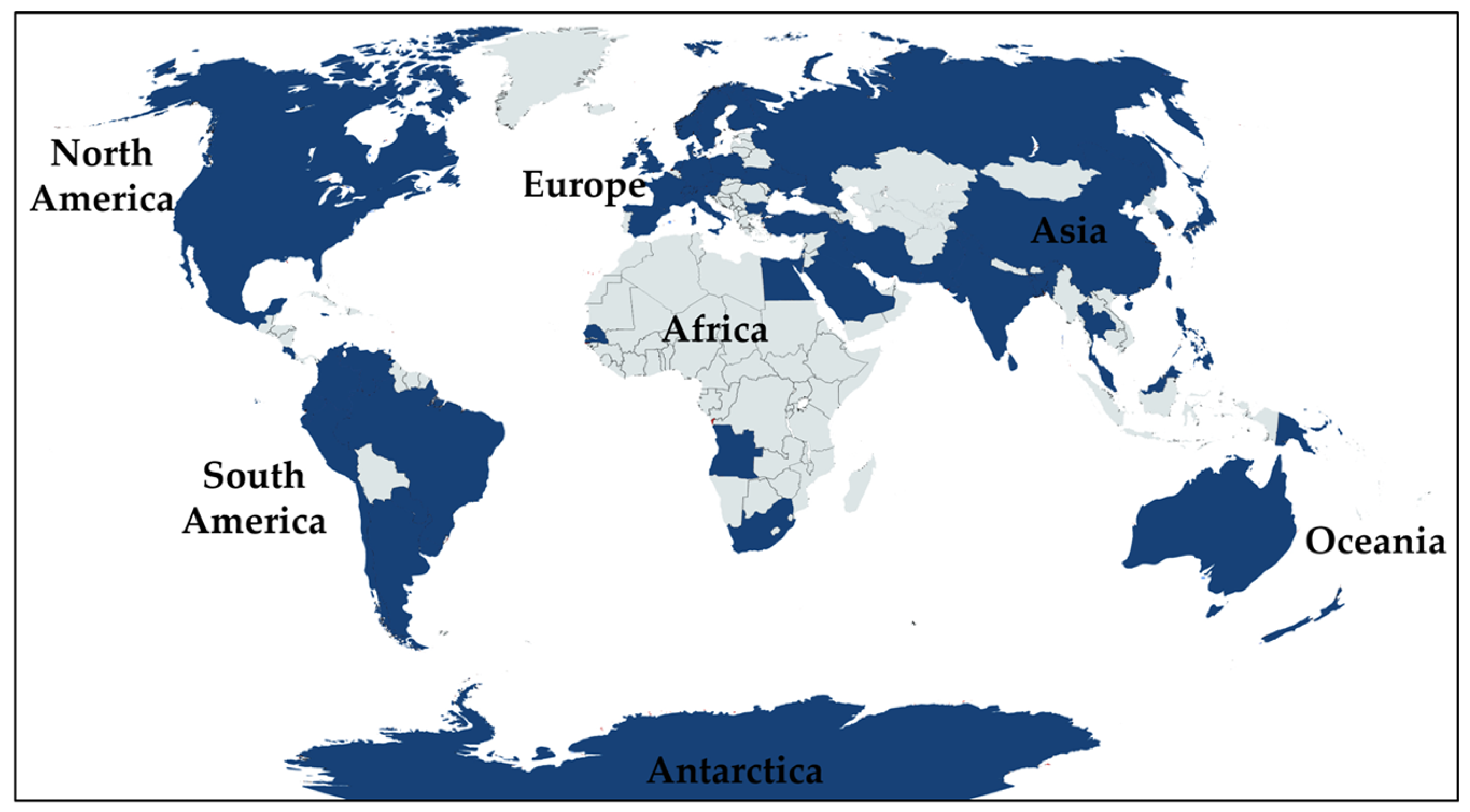
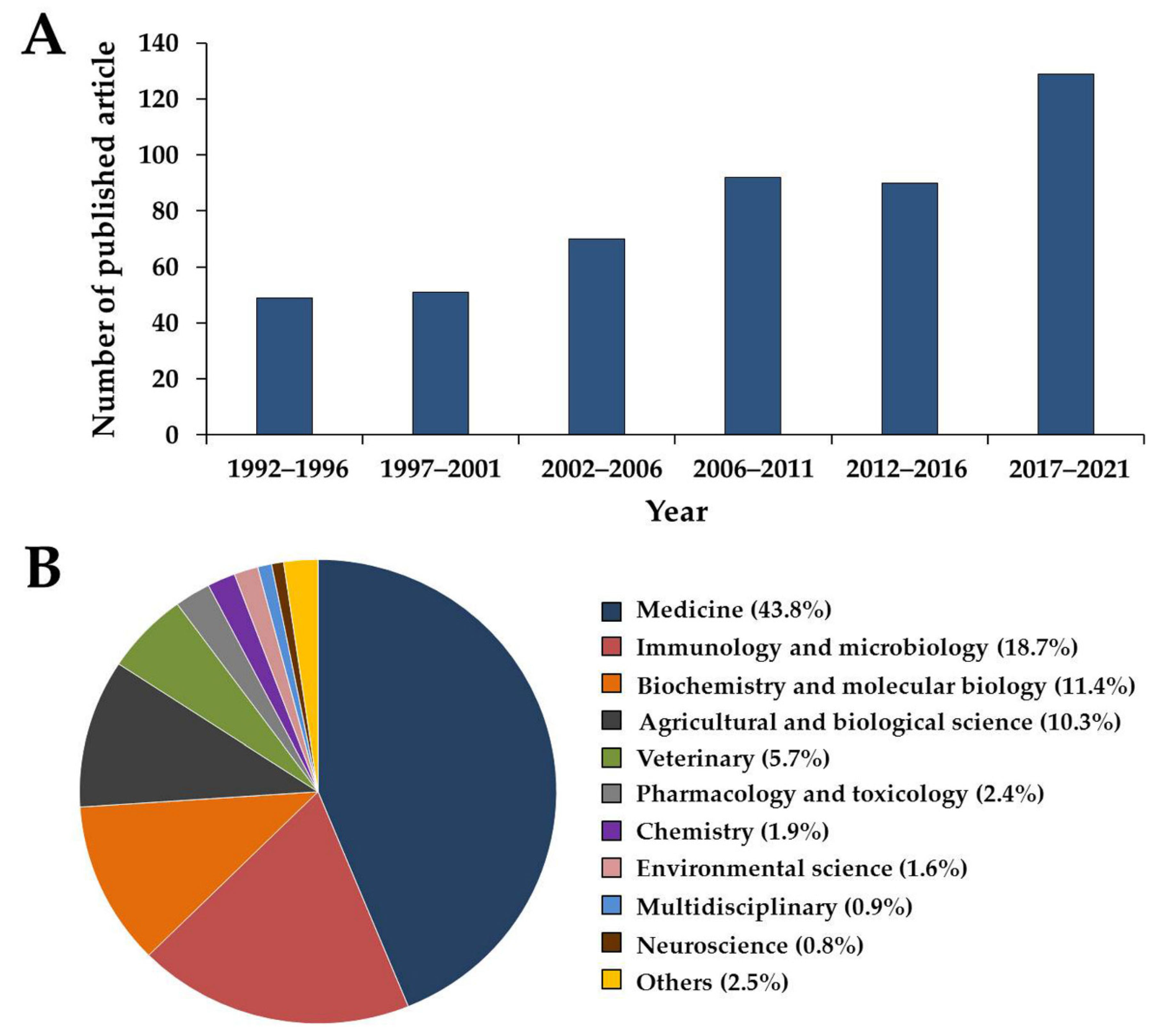
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Fungal Isolation
2.2. Morphological and Growth Observations
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Sequence Alignment
2.5. Phylogenetic Analyses
3. Results
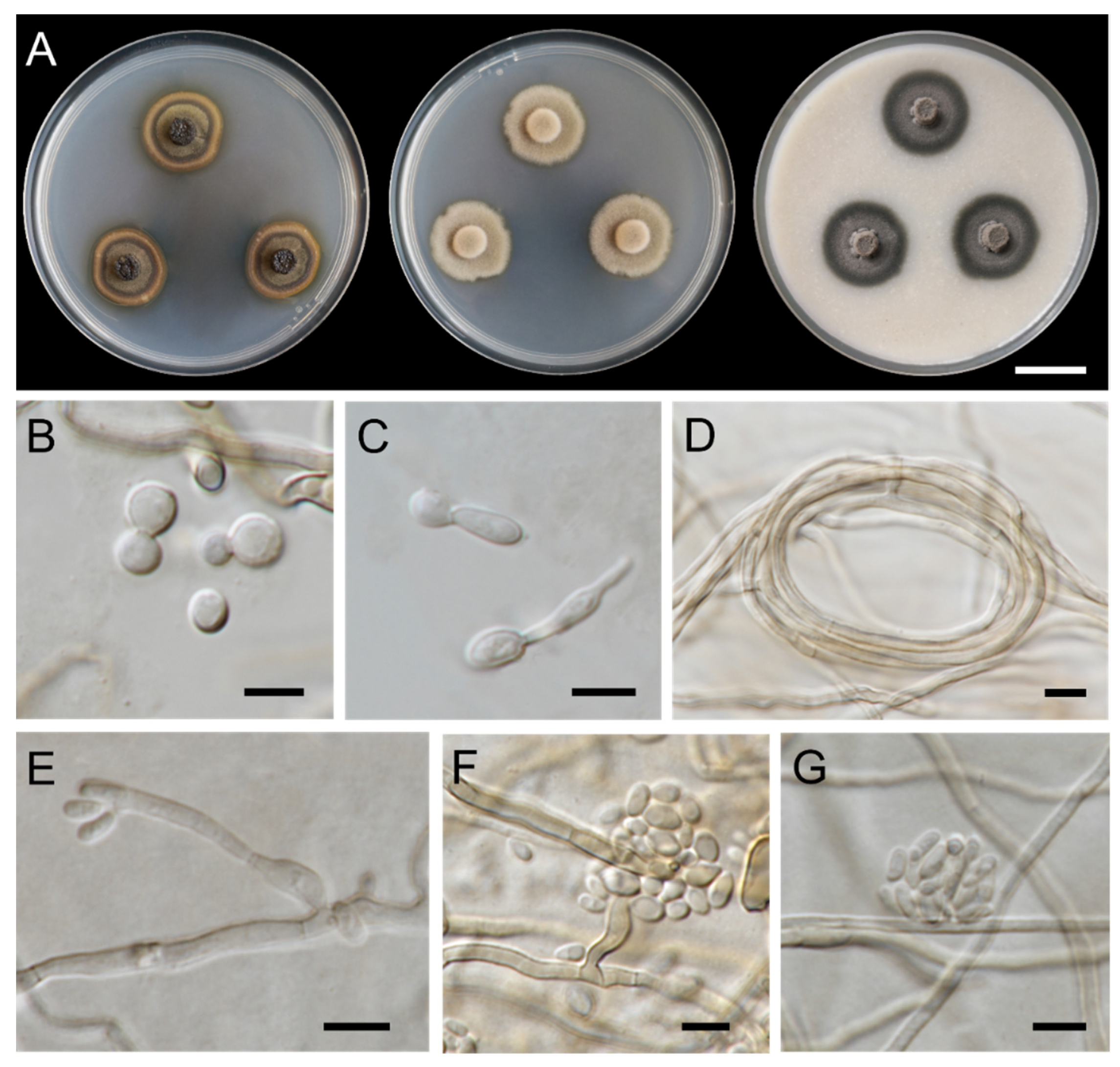
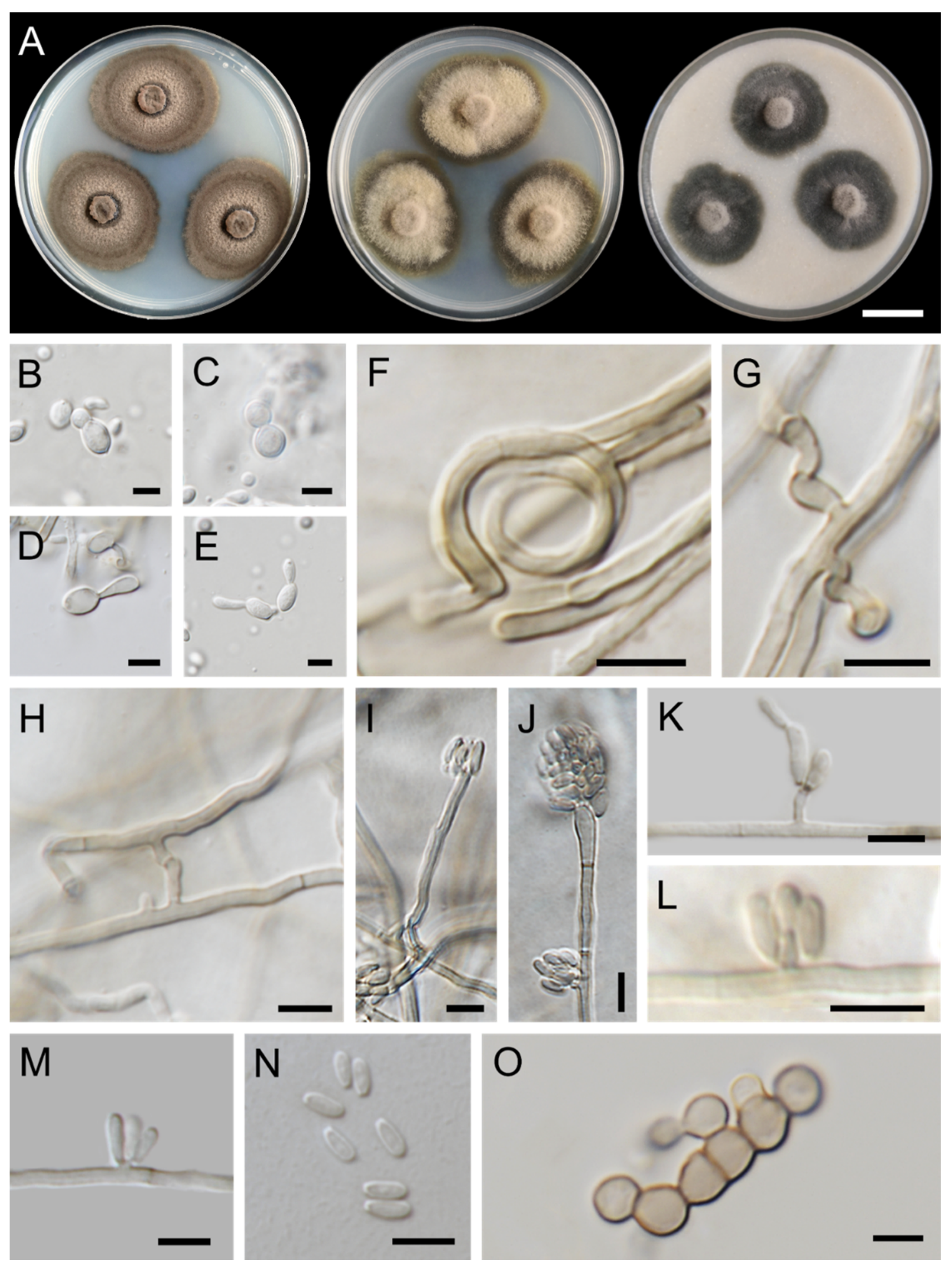
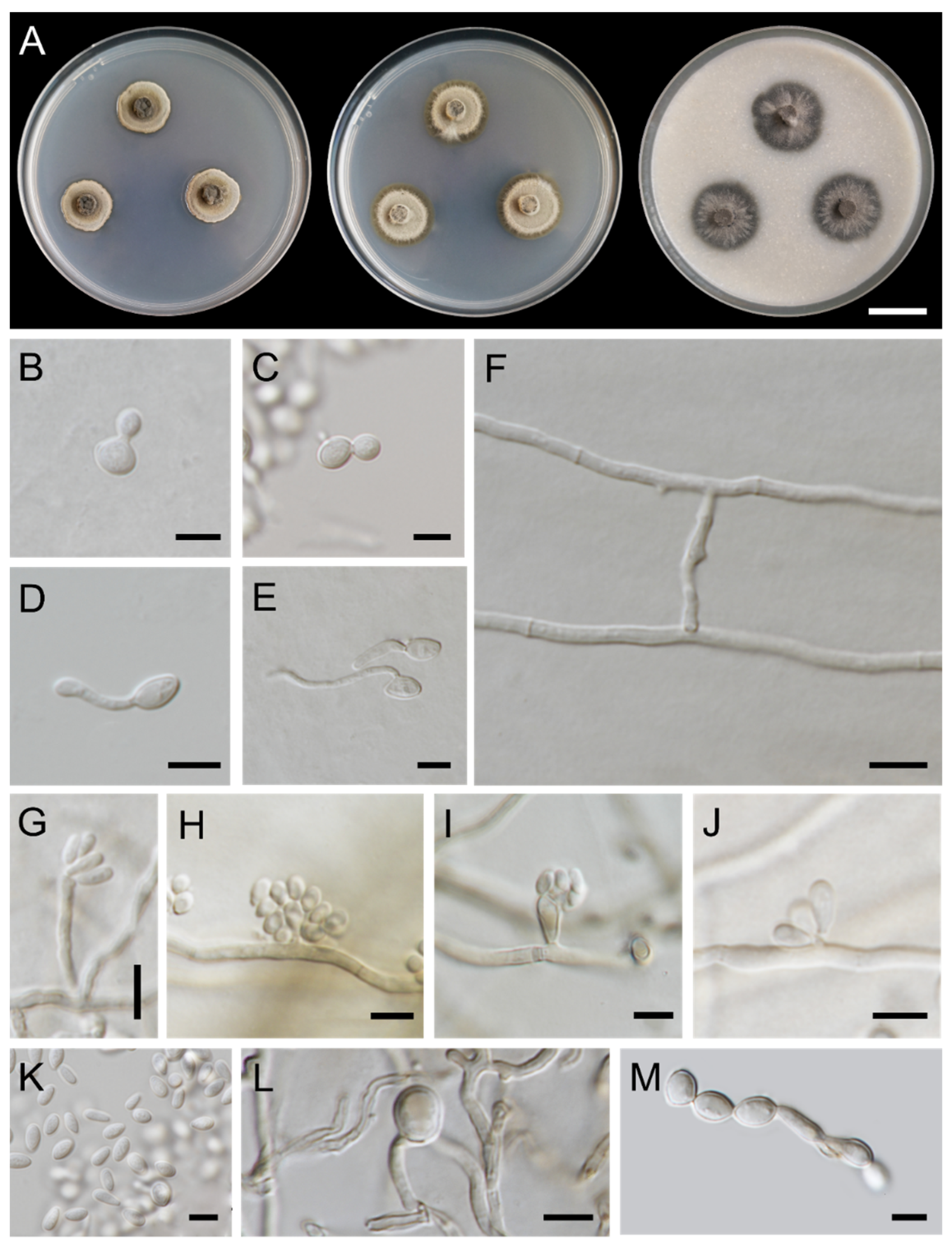
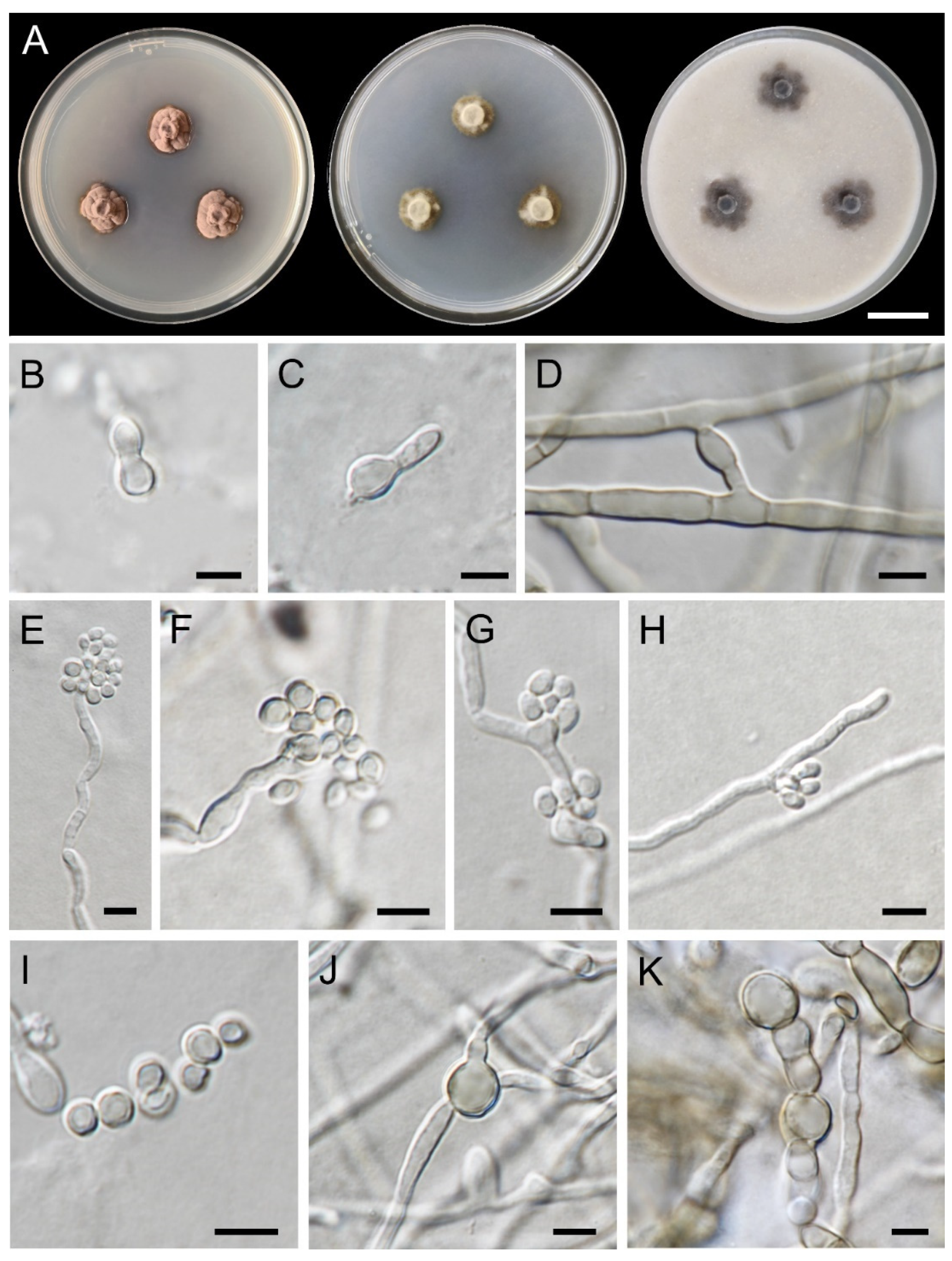
3.1. Fungal Isolation and Morphological Observations
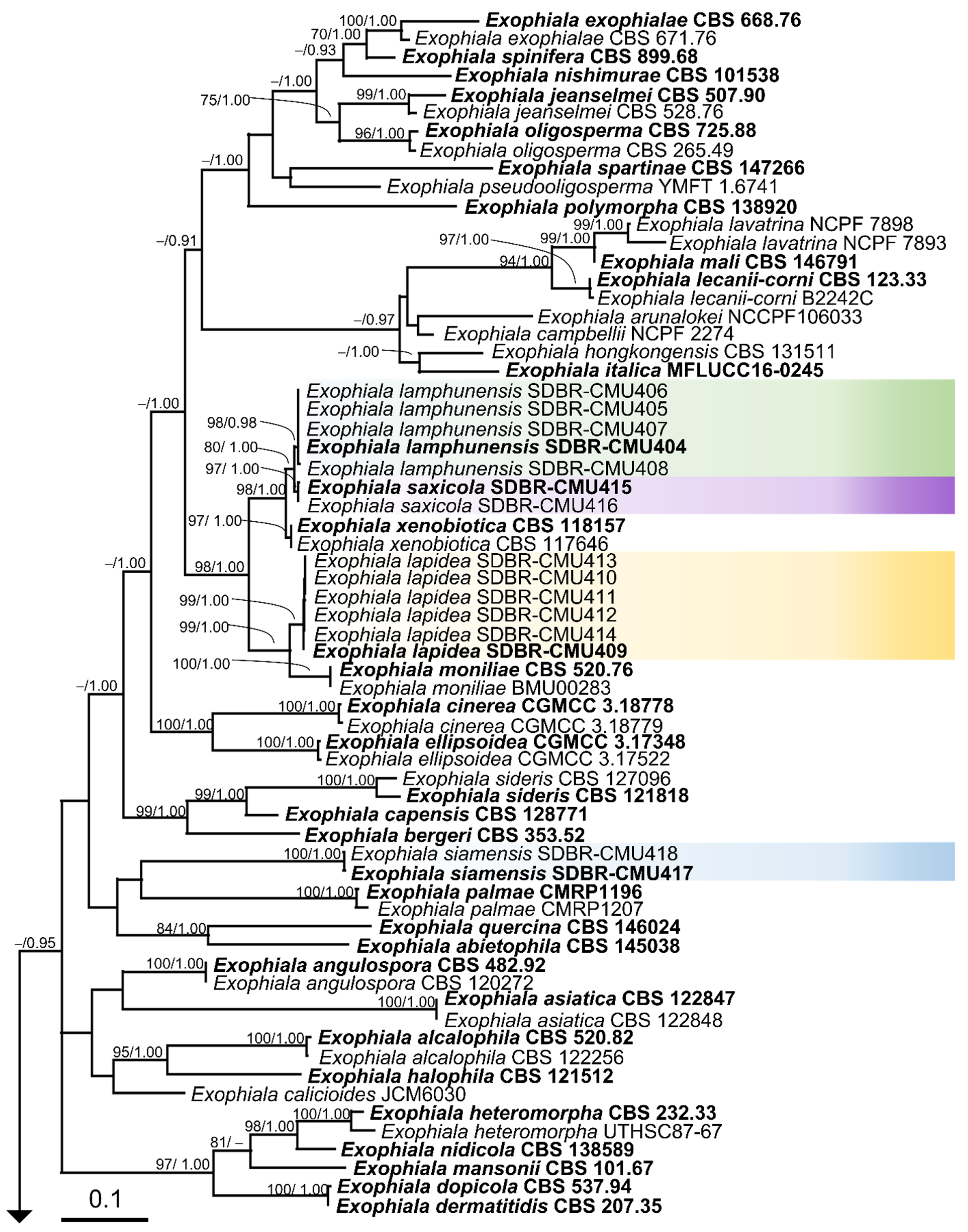
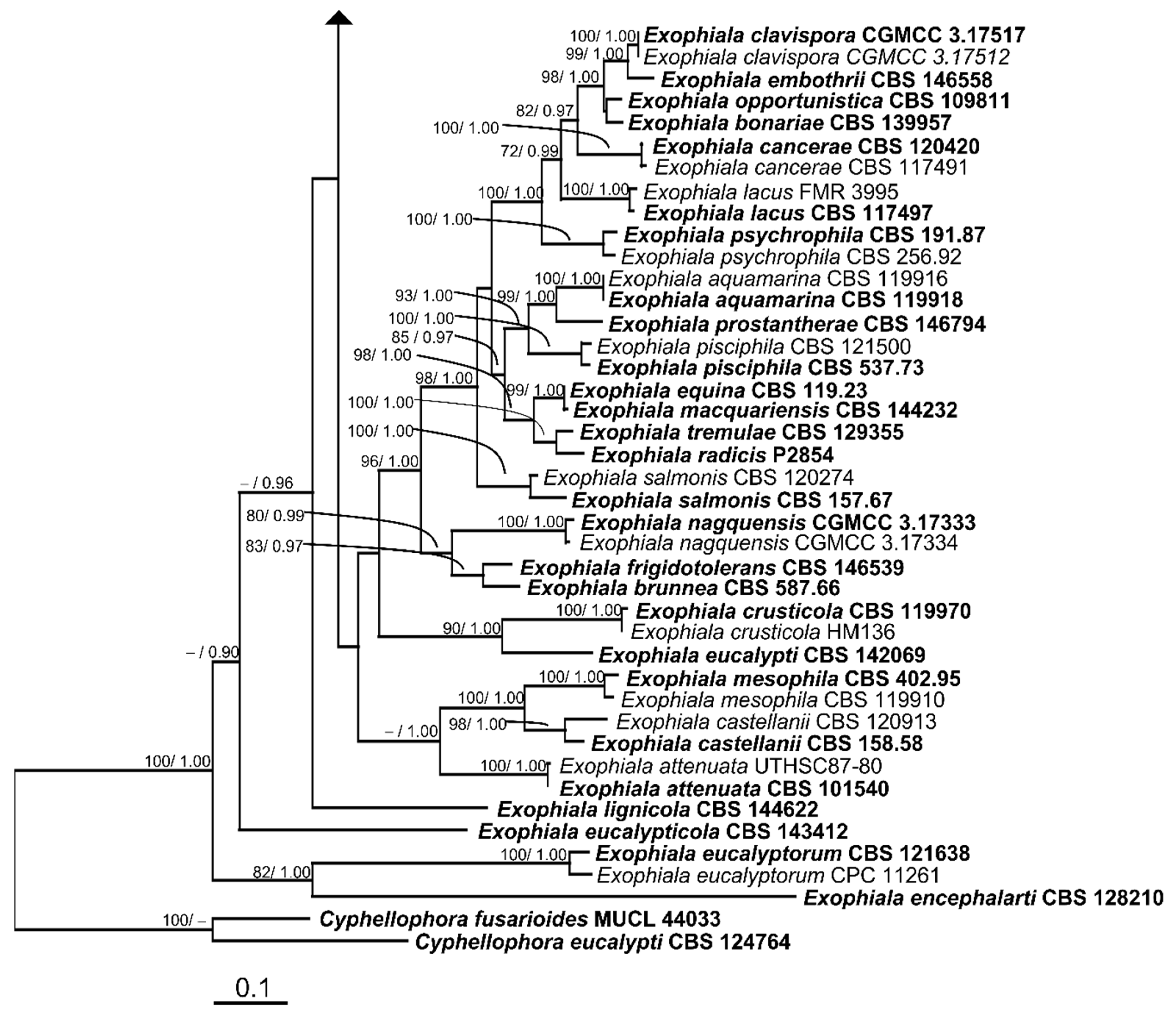
3.2. Phylogenetic Results
3.3. Taxonomic Descriptions
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carmichael, J.W. Cerebral mycetoma of trout due to a phialophora-like fungus. Sabouraudia 1966, 5, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Sirijovski, N.; Adler, L.; Ferrari, B.C. Exophiala macquariensis sp. nov., a cold adapted black yeast species recovered from a hydrocarbon contaminated sub-Antarctic soil. Fungal Biol. 2019, 123, 151–158. [Google Scholar] [CrossRef] [PubMed]
- de Hoog, G.S.; Vicente, V.A.; Najafzadeh, M.J.; Harrak, M.J.; Badali, H.; Seyedmousavi, S. Waterborne Exophiala species causing disease in cold-blooded animals. Persoonia 2011, 27, 46–72. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.Q.; Feng, M.Y.; Yu, Z.F. Exophiala pseudooligosperma sp. nov., a novel black yeast from soil in southern China. Int. J. Syst. Evol. Microbiol. 2021, 71, e005116. [Google Scholar] [CrossRef]
- Maciá-Vicente, J.G.; Glynou, K.; Piepenbring, M. A new species of Exophiala associated with roots. Mycol. Prog. 2016, 15, e18. [Google Scholar] [CrossRef]
- Zeng, J.S.; de Hoog, G.S. Exophiala spinifera and its allies: Diagnostics from morphology to DNA barcoding. Med. Mycol. 2008, 46, 193–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borman, A.M.; Fraser, M.; Szekely, A.; Larcombe, D.E.; Johnson, E.M. Rapid identification of clinically relevant members of the genus Exophiala by matrix-assisted laser desorption ionization–time of flight mass spectrometry and description of two novel species, Exophiala campbellii and Exophiala lavatrina. J. Clin. Microbiol. 2017, 55, 1162–1176. [Google Scholar] [CrossRef] [Green Version]
- Yong, L.K.; Wiederhold, N.P.; Sutton, D.A.; Sandoval-Denis, M.; Lindner, J.R.; Fan, H.; Sanders, C.; Guarro, J. Morphological and molecular characterization of Exophiala polymorpha sp. nov. isolated from sporotrichoid lymphocutaneous lesions in a patient with Myasthenia Gravis. J. Clin. Microbiol. 2015, 53, 2816–2822. [Google Scholar] [CrossRef] [Green Version]
- Seyedmousavi, S.; Badali, H.; Chlebicki, A.; Zhao, J.; Prenafeta-Boldú, F.X.; De Hoog, G.S. Exophiala sideris, a novel black yeast isolated from environments polluted with toxic alkyl benzenes and arsenic. Fungal Biol. 2011, 115, 1030–1037. [Google Scholar] [CrossRef]
- Julou, T.; Burghardt, B.; Gebauer, G.; Berveiller, D.; Damesin, C.; Selosse, A. Mixotrophy in orchids: Insights from a comparative study of green individuals and nonphotosynthetic individuals of Cephalanthera damasonium. New Phytol. 2005, 166, 639–653. [Google Scholar] [CrossRef]
- Haase, G.; Sonntag, L.; Melzer-Krick, B.; de Hoog, G.S. Phylogenetic inference by SSU gene analysis of members of the Herpotrichiellaceae, with special reference to human pathogenic species. Stud. Mycol. 1999, 43, 80–97. [Google Scholar]
- McGinnis, M.R.; Padhye, A.A. Exophiala jeanselmei, a new combination for Phialophora jeanselmei. Mycotaxon 1977, 5, 341–352. [Google Scholar]
- Okada, G.; Jacobs, K.; Kirisits, T.; Louis-Seize, G.W.; Seifert, K.A.; Sugita, T.; Takematsu, A.; Wingfield, M.J. Epitypification of Graphium penicillioides Corda, with comments on the phylogeny and taxonomy of graphium-like synnematous fungi. Stud. Mycol. 2000, 45, 169–188. [Google Scholar]
- McGinnis, M.R. Exophiala spinifera, a new combination for Phialophora spinifera. Mycotaxon 1977, 5, 337–340. [Google Scholar]
- de Hoog, G.S.; Hermanides-Nijhof, E.J. The black yeasts and allied Hyphomycetes. Stud. Mycol. 1977, 15, e222. [Google Scholar]
- Matos, T.; Haase, G.; Gerrits van den Ende, A.H.G.; de Hoog, G.S. Molecular diversity of oligotrophic and neurotropic members of the black yeast genus Exophiala, with accent on E. dermatitidis. Antonie Van Leeuwenhoek 2003, 83, 293–303. [Google Scholar] [CrossRef]
- de Hoog, G.S.; Vicente, V.; Caligiorne, R.B.; Kantarcioglu, S.; Tintelnot, K.; Gerrits van den Ende, A.H.; Haase, G. Species diversity and polymorphism in the Exophiala spinifera clade containing opportunistic black yeast-like fungi. J. Clin. Microbiol. 2003, 41, 4767–4778. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Su, L.; Yang, S.; Sun, J.; Liu, B.; Fu, R.; Wu, B.; Lui, X.; Cai, L.; Guo, L.; et al. Unveiling the hidden diversity of rock-inhabiting fungi: Chaetothyriales from China. J. Fungi 2020, 6, e187. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: http://www.indexfungorum.org (accessed on 22 March 2022).
- Singh, S.; Rudramurthy, S.M.; Padhye, A.A.; Hemashetter, B.M.; Iyer, R.; Hallur, V.; Sharma, A.; Agnihotri, S.; Gupta, S.; Ghosh, A.; et al. Clinical spectrum, molecular characterization, antifungal susceptibility testing of Exophiala spp. from India and description of a novel Exophiala species, E. arunalokei sp. nov. Front. Cell Infect. Microbiol. 2021, 11, e686120. [Google Scholar] [CrossRef]
- Crous, P.W.; Schumacher, R.K.; Akulov, A.; Thangavel, R.; Hernández-Restrepo, M.; Carnegie, A.J.; Cheewangkoon, R.; Wingfield, M.J.; Summerell, B.A.; Quaedvlieg, W. New and interesting fungi. 2. Fungal Syst. Evol. 2019, 3, 57–134. [Google Scholar] [CrossRef]
- Crous, P.W.; Schumacher, R.K.; Wingfield, M.J.; Akulov, A.; Denman, S.; Roux, J.; Braun, U.; Burgess, T.I.; Carnegie, A.J.; Váczy, K.Z. New and interesting fungi. 1. Fungal. Syst. Evol. 2018, 1, 169–216. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Schubert, K.; Braun, U.; Hoog, G.S.; de Hocking, A.D.; Shin, H.D.; Groenewald, J.Z. Opportunistic, human-pathogenic species in the Herpotrichiellaceae are phenotypically similar to saprobic or phytopathogenic species in the Venturiaceae. Stud. Mycol. 2007, 58, 185–217. [Google Scholar] [CrossRef] [PubMed]
- Tibpromma, S.; Hyde, K.D.; Jeewon, R.; Maharachchikumbura, S.S.N.; Liu, J.K.; Bhat, D.J.; Jones, E.B.G.; McKenzie, E.H.C.; Camporesi, E.; Bulgakov, T.S.; et al. Fungal diversity notes 491–602: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2017, 83, 1–261. [Google Scholar] [CrossRef]
- Estévez, E.; Veiga, M.C.; Kennes, C. Biodegradation of toluene by the new fungal isolates Paecilomyces variotii and Exophiala oligosperma. J. Ind. Microbiol. Biotechnol. 2005, 32, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Libert, X.; Chasseur, C.; Packeu, A.; Bureau, F.; Roosens, N.H.; De Keersmaecker, S.C.J. Exploiting the advantages of molecular tools for the monitoring of fungal indoor air contamination: First detection of Exophiala jeanselmei in indoor air of air-conditioned offices. Microorganisms 2019, 7, e674. [Google Scholar] [CrossRef] [Green Version]
- de Hoog, G.S.; Zeng, J.S.; Harrak, M.J.; Sutton, D.A. Exophiala xenobiotica sp. nov., an opportunistic black yeast inhabiting environments rich in hydrocarbons. Antonie van Leeuwenhoek 2006, 90, 257–268. [Google Scholar] [CrossRef]
- Isola, D.; Selbmann, L.; de Hoog, G.S.; Fenice, M.; Onofri, S.; Prenafeta-Boldú, F.X.; Zucconi, L. Isolation and screening of black fungi as degraders of volatile aromatic hydrocarbons. Mycopathologia 2013, 175, 369–379. [Google Scholar] [CrossRef]
- Listemann, H.; Freiesleben, H. Exophiala mesophila spec. nov. Mycoses 1996, 39, 1–3. [Google Scholar] [CrossRef]
- Li, D.M.; Li, R.Y.; de Hoog, G.S.; Wang, Y.X.; Wang, D.L. Exophiala asiatica, a new species from a fatal case in China. Med. Mycol. 2009, 47, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Woo, P.C.; Ngan, A.H.; Tsang, C.C.; Ling, I.W.; Chan, J.F.; Leung, S.Y.; Yuen, K.Y.; Lau, S.K. Clinical spectrum of Exophiala infections and a novel Exophiala species, Exophiala hongkongensis. J. Clin. Microbiol. 2013, 51, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Li, D.M.; Li, R.Y.; de Hoog, G.S.; Sudhadham, M.; Wang, D.L. Fatal Exophiala infections in China, with a report of seven cases. Mycoses 2011, 54, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Vitale, R.G.; de Hoog, G.S. Molecular diversity, new species and antifungal susceptibilities in the Exophiala spinifera clade. Med. Mycol. 2002, 40, 545–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.; Feng, P.; Gerrits van den Ende, A.H.G.; Xi, L.; Harrak, M.J.; de Hoog, G.S. Multilocus analysis of the Exophiala jeanselmei clade containing black yeasts involved in opportunistic disease in humans. Fungal Divers. 2014, 65, 3–16. [Google Scholar] [CrossRef]
- Barba-Gómez, J.F.; Mayorga, J.; McGinnis, M.R.; González-Mendoza, A. Chromoblastomycosis caused by Exophiala spinifera. J. Am. Acad. Derm. 1992, 26, 367–370. [Google Scholar] [CrossRef]
- Padhye, A.A.; Kaplan, W.; Neuman, M.A.; Case, P.; Radcliffe, G.N. Subcutaneous phaeohyphomycosis caused by Exophiala spinifera. J. Med. Vet. Mycol. 1984, 22, 493–500. [Google Scholar] [CrossRef]
- Padhye, A.A.; Hampton, A.A.; Hampton, M.T.; Hutton, N.W.; Prevost-Smith, E.; Davis, M.S. Chromoblastomycosis caused by Exophiala spinifera. Clin. Infect. Dis. 1996, 22, 331–335. [Google Scholar] [CrossRef] [Green Version]
- Lacaz, C.S.; Porto, E.; Andrade, J.G.; Telles Filho, F.Q. Feohifomicose disseminada por Exophiala spinifera. An. Bras. Derm. 1984, 59, 238–243. [Google Scholar]
- Rajam, R.V.; Kandhari, K.C.; Thirumalachar, M. Chromoblastomycosis caused by a rare yeast-like dematiaceous fungus. Mycopathol. Mycol. Appl. 1958, 9, 5–19. [Google Scholar] [CrossRef]
- Nielsen, H.S.; Conant, N.F. A new pathogenic Phialophora. Sabouraudia 1968, 6, 228–231. [Google Scholar] [CrossRef]
- Kettlewell, P.; McGinnis, M.R.; Wilkinson, G.T. Phaeohyphomycosis caused by Exophiala spinifera in two cats. J. Med. Vet. Mycol. 1989, 27, 257–264. [Google Scholar] [CrossRef]
- Conti-Díaz, I.A.; MacKinnon, J.E.; Civila, E. Isolation and identification of black yeasts from the external environment in Uruguay. Pan Am. Health Org. Sci. Publ. 1977, 356, 109–114. [Google Scholar]
- Song, Y.; Laureijssen-van de Sande, W.W.J.; Moreno, L.F.; Gerrits van den Ende, B.; Li, R.; de Hoog, S. Comparative ecology of capsular Exophiala species causing disseminated infection in humans. Front. Microbiol. 2017, 8, e2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenz, B.E.; Held, B.W.; Jurgens, J.A.; Farrell, R.L.; Blanchette, R.A. Fungal diversity in soils and historic wood from the Ross Sea Region of Antarctica. Soil Biol. Biochem. 2006, 38, 3057–3064. [Google Scholar] [CrossRef]
- Alimu, Y.; Ban, S.; Yaguchi, T. Molecular phylogenetic study of strains morphologically identified as Exophiala dermatitidis from clinical and environmental specimens in Japan. Med. Mycol. J. 2022, 63, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sudhadham, M.; Prakitsin, S.; Sivichai, S.; Chaiyarat, R.; Dorrestein, G.M.; Menken, S.B.; de Hoog, G.S. The neurotropic black yeast Exophiala dermatitidis has a possible origin in the tropical rain forest. Stud. Mycol. 2008, 61, 145–155. [Google Scholar] [CrossRef]
- Döğen, A.; Kaplan, E.; Ilkit, M.; de Hoog, G.S. Massive contamination of Exophiala dermatitidis and E. phaeomuriformis in railway stations in subtropical Turkey. Mycopathologia 2013, 175, 381–386. [Google Scholar] [CrossRef]
- Gümral, R.; Tümgör, A.; Saraçlı, M.A.; Yıldıran, Ş.T.; Ilkit, M.; de Hoog, G.S. Black yeast diversity on creosoted railway sleepers changes with ambient climatic conditions. Microb. Ecol. 2014, 68, 699–707. [Google Scholar] [CrossRef]
- Zupančič, J.; Novak Babič, M.; Zalar, P.; Gunde-Cimerman, N. The black yeast Exophiala dermatitidis and other selected opportunistic human fungal pathogens spread from dishwashers to kitchens. PLoS ONE 2016, 11, e0148166. [Google Scholar] [CrossRef]
- Yazdanparast, S.A.; Mohseni, S.; de Hoog, G.S.; Aslani, N.; Sadeh, A.; Badali, H. Consistent high prevalence of Exophiala dermatitidis, a neurotropic opportunist, on railway sleepers. J. Mycol. Med. 2017, 27, 180–187. [Google Scholar] [CrossRef]
- Sav, H.; Ozakkas, F.; Altınbas, R.; Kiraz, N.; Tümgör, A.; Gümral, R.; Döğen, A.; Ilkit, M.; de Hoog, G.S. Virulence markers of opportunistic black yeast in Exophiala. Mycoses 2016, 59, 343–350. [Google Scholar] [CrossRef]
- Jayaram, M.; Nagao, H. First report of environmental isolation of Exophiala spp. in Malaysia. Curr. Microbiol. 2020, 77, 2915–2924. [Google Scholar] [CrossRef] [PubMed]
- Klasinc, R.; Riesenhuber, M.; Bacher, A.; Willinger, B. Invasive fungal infection caused by Exophiala dermatitidis in a patient after lung transplantation: Case report and literature review. Mycopathologia 2019, 184, 107–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naka, W.; Harada, T.; Nishikawa, T.; Fukushiro, R. A case of chromoblastomycosis: With special reference to the mycology of the isolated Exophiala jeanselmei. Mykosen 1986, 29, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Langeron, M. Mycétome à Torula jeanselmei Langeron, 1928. Nouveau type de mycétome à grains noirs. Ann. Parasitol. Hum. Comp. 1928, 6, 385–403. [Google Scholar] [CrossRef] [Green Version]
- Murray, I.G.; Dunkerley, G.E.; Hughes, K.E.A. A case of madura foot caused by Phialophora jeanselmei. Sabouraudia 1964, 3, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, H.; Matsumoto, T.; Kimura, U.; Hiruma, M.; Kano, R.; Yaguchi, T.; Kubo, M.; Kashiwada-Nakamura, K.; Fukushima, S. Empiric antifungal therapy in patients with cutaneous and subcutaneous phaeohyphomycosis. J. Dermatol. 2022, 49, 564–571. [Google Scholar] [CrossRef]
- Pattanaprichakul, P.; Bunyaratavej, S.; Leeyaphan, C.; Sitthinamsuwan, P.; Sudhadham, M.; Muanprasart, C.; Feng, P.; Badali, H.; de Hoog, G.S. An unusual case of eumycetoma caused by Exophiala jeanselmei after a sea urchin injury. Mycoses 2013, 56, 491–494. [Google Scholar] [CrossRef]
- Valiente, C.; Quesada, E. Morphologic and physiologic characteristics of Costa Rica pathogenic fungi (Dermatiaceae). Rev. Biol. Trop. 1991, 39, 103–106. [Google Scholar]
- Ramírez Soto, M.C.; Malaga, G. Subcutaneous mycoses in Peru: A systematic review and meta-analysis for the burden of disease. Int. J. Dermatol. 2017, 56, 1037–1045. [Google Scholar] [CrossRef]
- Al-Tawfiq, J.A.; Amr, S.S. Madura leg due to Exophiala jeanselmei successfully treated with surgery and itraconazole therapy. Med. Mycol. 2009, 47, 648–652. [Google Scholar] [CrossRef] [Green Version]
- Thammayya, A.; Sanyal, M. Exophiala jeanselmei causing mycetoma pedis in India. Sabouraudia 1980, 18, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Brownell, I.; Pomeranz, M.; Ma, L. Eumycetoma. Dermatol. Online J. 2005, 11, e10. [Google Scholar] [CrossRef]
- Nielsen Jr, H.S.; Conant, N.F.; Weinberg, T.; Reback, J.F. Report of a mycetoma due to Phialophora jeanselmei and undescribed characteristics of the fungus. Saboraudia 1968, 6, 330–333. [Google Scholar] [CrossRef]
- Negroni, R. Estudio micologico del primer caso de micetoma por Phialophora jeanselmei observado en la Argentina. Med. Cut. Iberolat. 1970, 5, 625–630. [Google Scholar]
- Simpson, A.; Singh, S.R. A case of Madura foot. J. Roy. College. Surg. Edinburgh 1984, 29, 326–328. [Google Scholar]
- Badali, H.; Najafzadeh, M.J.; van Esbroeck, M.; van den Enden, E.; Tarazooie, B.; Meis, J.F.; de Hoog, G.S. The clinical spectrum of Exophiala jeanselmei, with a case report and in vitro antifungal susceptibility of the species. Med. Mycol. 2010, 48, 318–327. [Google Scholar] [CrossRef] [Green Version]
- Seifert, K.A.; Okada, G. Graphium anamorphs of Ophiostoma species and similar anamorphs of other Ascomycetes. In Ceratocystis and Ophiostoma: Taxonomy, Ecology, and Pathogenicity; Wingfield, M.J., Seifert, K.A., Webber, J.F., Eds.; The American Phytopathological Society Press: St. Paul, MN, USA, 1993; pp. 27–41. [Google Scholar]
- Isola, D.; Zucconi, L.; Onofri, S.; Caneva, G.; de Hoog, G.S.; Selbmann, L. Extremotolerant rock inhabiting black fungi from Italian monumental sites. Fungal Divers. 2016, 76, 75–96. [Google Scholar] [CrossRef]
- Borman, A.M.; Fraser, M.; Schilling, W.; Jones, G.; Pearl, R.; Linton, C.J.; Johnson, E.M. Exophiala campbellii causing a subcutaneous palmar cyst in an otherwise healthy UK resident. Med. Mycol. Case Rep. 2020, 29, 43–45. [Google Scholar] [CrossRef]
- Madrid, H.; Hernández-Restrepo, M.; Gené, J.; Cano, J.; Guarro, J.; Silva, V. New and interesting chaetothyrialean fungi from Spain. Mycol. Prog. 2016, 15, 1179–1201. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.; Gené, J.; Guarro, J.; Baseia, I.G.; García, D.; Gusmão, L.; Souza-Motta, C.M.; et al. Fungal planet description sheets: 716–784. Persoonia 2018, 40, 240–393. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Schumacher, R.K.; Akulov, A.; Bulgakov, T.S.; Carnegie, A.J.; Jurjević, Ž.; Decock, C.; Denman, S.; Lombard, L.; et al. New and interesting fungi. 3. Fungal Syst. Evol. 2020, 6, 157–231. [Google Scholar] [CrossRef] [PubMed]
- Kochkina, G.A.; Ivanushkina, N.E.; Lupachev, A.V.; Starodumova, I.P.; Vasilenko, O.V.; Ozerskaya, S.M. Diversity of mycelial fungi in natural and human-affected Antarctic soils. Polar Biol. 2019, 42, 47–64. [Google Scholar] [CrossRef]
- Kondratiuk, T.O.; Kondratyuk, S.Y.; Khimich, M.V.; Beregova, T.V.; Ostapchenko, L.I. Confirmation of taxonomic status of black yeast-like fungus by three gene phylogeny. Acta Bot. Hung. 2016, 58, 287–302. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, K.; Miyaji, M.; Taguchi, H.; Tanaka, R. Fungi in bathwater and sludge of bathroom drainpipes. 1. Frequent isolation of Exophiala species. Mycopathologia 1987, 97, 17–23. [Google Scholar] [CrossRef]
- Nyaoke, A.; Weber, E.S.; Innis, C.; Stremme, D.; Dowd, C.; Hinckley, L.; Gorton, T.; Wickes, B.; Sutton, D.; de Hoog, S. Disseminated phaeohyphomycosis in weedy seadragons (Phyllopteryx taeniolatus) and leafy seadragons (Phycodurus eques) caused by species of Exophiala, including a novel species. J. Vet. Diagn. Investig. 2009, 21, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, M.; Beckmann, M.J.; Pflaum, S.; Pearson, M.; Carcajona, D.; Treasurer, J.W.; West, P.V. Exophiala angulospora infection in hatchery-reared lumpfish (Cyclopterus lumpus) broodstock. J. Fish Dis. 2019, 42, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Scholz, F.; Ruane, N.M.; Marcos-López, M.; Mitchell, S.; Bolton-Warberg, M.; O’Connor, I.; Mirimin, L.; MacCarthy, E.; Rodger, H.D. Systemic mycoses in lumpfish (Cyclopterus lumpus L.) in Ireland: Aetiology and clinical presentation. Bull. Eur. Ass. Fish Pathol. 2018, 38, 202–212. [Google Scholar]
- Kanchan, C.; MuraosaKishio, Y.; Hatai, H. Exophiala angulospora infection found in cultured Japanese flounder Paralichthys olivaceus in Japan. Bull. Eur. Ass. Fish Pathol. 2014, 34, 187–194. [Google Scholar]
- Gjessing, M.C.; Davey, M.; Kvellestad, A.; Vrålstad, T. Exophiala angulospora causes systemic inflammation in Atlantic cod Gadus morhua. Dis. Aquat. Organ. 2011, 96, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Chermette, R.; Ferreiro, L.; De Bievre, C.; Camadro, J.P.; Mialot, M.; Vauzelle, P. Exophiala spinifera nasal infection in a cat and a literature review of feline phaeohyphomycosis. J. Mycol. Med. 1997, 7, 149–158. [Google Scholar]
- Bernhardt, A.; Bomhard, W.V.; Antweiler, E.; Tintelnot, K. Molecular identification of fungal pathogens in nodular skin lesions of cats. Med. Mycol. 2015, 53, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Overy, D.P.; Martin, C.; Muckle, A.; Lund, L.; Wood, J.; Hanna, P. Cutaneous phaeohyphomycosis caused by Exophiala attenuata in a domestic cat. Mycopathologia 2015, 180, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.C.; Gonçalves, S.S.; Santos, D.W.; Padovan, A.C.; Bizerra, F.C.; Melo, A.S. Species diversity, antifungal susceptibility and phenotypic and genotypic characterisation of Exophiala spp. infecting patients in different medical centres in Brazil. Mycoses 2017, 60, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Rakeman, J.L.; Bui, U.; LaFe, K.; Chen, Y.C.; Honeycutt, R.J.; Cookson, B.T. Multilocus DNA sequence comparisons rapidly identify pathogenic molds. J. Clin. Microbiol. 2005, 43, 3324–3333. [Google Scholar] [CrossRef] [Green Version]
- Figel, I.C.; Marangoni, P.R.; Tralamazza, S.M.; Vicente, V.A.; Dalzoto, P.; do Nascimento, M.M.; de Hoog, G.S.; Pimentel, I.C. Black yeasts-like fungi isolated from dialysis water in hemodialysis units. Mycopathologia 2013, 175, 413–420. [Google Scholar] [CrossRef]
- Vicente, V.A.; Orélis-Ribeiro, R.; Najafzadeh, M.J.; Sun, J.; Guerra, R.S.; Miesch, S.; Ostrensky, A.; Meis, J.F.; Klaassen, C.H.; de Hoog, G.S.; et al. Black yeast-like fungi associated with Lethargic Crab Disease (LCD) in the mangrove-land crab, Ucides cordatus (Ocypodidae). Vet. Microbiol. 2012, 158, 109–122. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z. Why everlastings don’t last. Persoonia 2011, 26, 70–84. [Google Scholar] [CrossRef]
- Bates, S.T.; Reddy, G.; Garcia-Pichel, F. Exophiala crusticola anam. nov. (affinity Herpotrichiellaceae), a novel black yeast from biological soil crusts in the Western United States. Int. J. Syst. Evol. Microbiol. 2006, 56, 2697–2702. [Google Scholar] [CrossRef] [Green Version]
- Katz, B.; McGinnis, R. A new species of Exophiala recovered from loblolly pine litter. Mycotaxon 1980, 11, 182–184. [Google Scholar]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Larsson, E.; Angelini, C.; Brandrud, T.E.; Dearnaley, J.; Dima, B.; Dovana, F.; et al. Fungal Planet description sheets: 1112–1181. Persoonia 2020, 45, 251–409. [Google Scholar] [CrossRef]
- Persoonial Reflections. Pers. Mol. Phylogeny Evol. Fungi 2010, 25, 117–159.
- Ávila, A.; Groenewald, J.Z.; Trapero, A.; Crous, P.W. Characterization and epitypification of Pseudocercospora cladosporioides, the causal organism of Cercospora-leaf spot of olives. Mycol. Res. 2005, 109, 881–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollacci, G. Miceti del corpo umano e degli animali. Atti dell’Istituto Botanico Università di Pavia 1923, 18, 1–9. [Google Scholar]
- Manharth, A.; Lemberger, K.; Mylniczenko, N.; Pinkerton, M.; Pessier, A.P.; Kammeyer, P.; de Hoog, S. Disseminated Phaeohyphomycosis due to Exophiala species in a Galapagos tortoise, Geochelone nigra. J. Herpetol. Med. Surg. 2005, 15, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Neubert, K.; Mendgen, K.; Brinkmann, H.; Wirsel, S.G.R. Only a few fungal species dominate highly diverse mycofloras associated with the common reed. Appl. Environ. Microbiol. 2006, 72, 1118–1128. [Google Scholar] [CrossRef] [Green Version]
- Najafzadeh, M.J.; Suh, M.K.; Lee, M.H.; Ha, G.Y.; Kim, J.R.; Kim, T.H.; Lee, H.J.; Choi, J.S.; Meis, J.F.; de Hoog, G.S. Subcutaneous phaeohyphomycosis caused by Exophiala equina, with susceptibility to eight antifungal drugs. J. Med. Microbiol. 2013, 62, 797–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.; Crane, C.; Barrett, S.; Cano-Lira, J.F.; Le Roux, J.J.; Thangavel, R.; Guarro, J.; et al. Fungal planet description sheets: 469–557. Persoonia 2016, 37, 218–403. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Chooi, Y.H.; Gilchrist, C.; Lacey, E.; Pitt, J.I.; Roets, F.; Swart, W.J.; Cano-Lira, J.F.; Valenzuela-Lopez, N.; et al. Fungal planet description sheets: 1042–1111. Persoonia 2020, 44, 301–459. [Google Scholar] [CrossRef]
- Benedek, T.; Specht, G. Mykologisch-bakteriologische Untersuchungen über Pilze und Bakterien als Symbionten bei Kerbtieren. Zentbl. f. Bakt. Parasitenk. Infekt. Abt. I Orig. 1933, 130, 74–90. [Google Scholar]
- Lian, X.; de Hoog, G.S. Indoor wet cells harbour melanized agents of cutaneous infection. Med. Mycol. 2010, 48, 622–628. [Google Scholar] [CrossRef] [Green Version]
- McGinnis, M.R.; Sorrell, D.F.; Miller, R.L.; Kaminski, G.W. Subcutaneous phaeohyphomycosis caused by Exophiala moniliae. Mycopathologia 1981, 73, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Tintelnot, K.; de Hoog, G.S.; Thomas, E.; Steudel, W.I.; Huebner, K.; Seeliger, H.P.R. Cerebral phaeohyphomycosis caused by an Exophiala species. Mycoses 1991, 34, 239–244. [Google Scholar] [CrossRef]
- Nucci, M.; Akiti, T.; Barreiros, G.; Silveira, F.; Revankar, S.G.; Sutton, D.A.; Patterson, T.F. Nosocomial fungemia due to Exophiala jeanselmei var. jeanselmei and a Rhinocladiella species: Newly described causes of bloodstream infection. J. Clin. Microbiol. 2001, 39, 514–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossler, A.D.; Richter, S.S.; Chavez, A.J.; Vogelgesang, S.A.; Sutton, D.A.; Grooters, A.M.; Rinaldi, M.G.; de Hoog, G.S.; Pfaller, M.A. Exophiala oligosperma causing olecranon bursitis. J. Clin. Microbiol. 2003, 40, 4779–4782. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, M.M.F.; Vicente, V.A.; Bittencourt, J.V.M.; Gelinski, J.M.L.; Prenafeta-Boldú, F.X.; Romero-Güiza, M.; Fornari, G.; Gomes, R.R.; Santos, G.D.; Gerrits Van Den Ende, A.H.G.; et al. Diversity of opportunistic black fungi on babassu coconut shells, a rich source of esters and hydrocarbons. Fungal Biol. 2017, 121, 488–500. [Google Scholar] [CrossRef] [PubMed]
- Gold, W.L.; Vellend, H.; Salit, I.E.; Campbell, I.; Summerbell, R.; Rinaldi, M.; Simor, A.E. Successful treatment of systemic and local infections due to Exophiala species. Clin. Infect. Dis. 1994, 19, 339–341. [Google Scholar] [CrossRef] [PubMed]
- Harsonowati, W.; Marian, M.; Surono, S.; Narisawa, K. The effectiveness of a dark septate endophytic fungus, Cladophialophora chaetospira SK51, to mitigate strawberry Fusarium-wilt disease and with growth promotion activities. Front. Microbiol. 2020, 11, e585. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Thangavel, R.; Wingfield, M.J.; Noordeloos, M.E.; Dima, B.; Brandrud, T.E.; Jansen, G.M.; et al. Fungal planet description sheets: 1182–1283. Persoonia 2021, 46, 313–528. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z.; Shivas, R.G.; Edwards, J.; Seifert, K.A.; Alfenas, A.C.; Alfenas, R.F.; Burgess, T.I.; Carnegie, A.J.; Hardy, G.E.; et al. Fungal planet description sheets: 69–91. Persoonia 2011, 26, 108–156. [Google Scholar] [CrossRef]
- Scopus Database. Available online: https://www.scopus.com (accessed on 9 May 2022).
- Xu, R.; Li, T.; Shen, M.; Yang, Z.L.; Zhao, Z.W. Evidence for a dark septate endophyte (Exophiala pisciphila, H93) enhancing phosphorus absorption by maize seedlings. Plant Soil 2020, 452, 249–266. [Google Scholar] [CrossRef]
- Xiao, Y.; Dai, M.X.; Zhang, G.Q.; Yang, Z.X.; He, Y.M.; Zhan, F.D. Effects of the dark septate endophyte (DSE) Exophiala pisciphila on the growth of root cell wall polysaccharides and the cadmium content of Zea mays L. under cadmium stress. J. Fungi 2021, 7, e1035. [Google Scholar] [CrossRef] [PubMed]
- Ondeyka, J.G.; Zink, D.L.; Dombrowski, A.W.; Polishook, J.D.; Felock, P.J.; Hazuda, D.J.; Singh, S.B. Isolation, structure and HIV-1 integrase inhibitory activity of exophillic acid, a novel fungal metabolite from Exophiala pisciphila. J. Antibiot. 2003, 56, 1018–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheikh-Ali, Z.; Glynou, K.; Ali, T.; Ploch, S.; Kaiser, M.; Thines, M.; Bode, H.B.; Maciá-Vicente, J.G. Diversity of exophillic acid derivatives in strains of an endophytic Exophiala sp. Phytochemistry 2015, 118, 83–93. [Google Scholar] [CrossRef]
- Zhang, D.; Yang, X.; Kang, J.S.; Choi, H.D.; Son, B.W. Chlorohydroaspyrones A and B, antibacterial aspyrone derivatives from the marine-derived fungus Exophiala sp. J. Nat. Prod. 2008, 71, 1458–1460. [Google Scholar] [CrossRef] [PubMed]
- Doshida, J.; Hasegawa, H.; Onuki, H.; Shimidzu, N. Exophilin A, a new antibiotic from a marine microorganism Exophiala pisciphila. J. Antibiot. 1996, 49, 1105–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ide-Pérez, M.R.; Fernández-López, M.G.; Sánchez-Reyes, A.; Leija, A.; Batista-García, R.A.; Folch-Mallol, J.L.; Sánchez-Carbente, M.d.R. Aromatic hydrocarbon removal by novel extremotolerant Exophiala and Rhodotorula spp. from an oil polluted site in Mexico. J. Fungi. 2020, 6, e135. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Chen, J.; Dissanayake, A.J.; Doilom, M.; Hongsanan, S.; Jayawardena, R.S.; Jeewon, R.; Perera, R.H.; Thongbai, B.; et al. Thailand’s amazing diversity: Up to 96% of fungi in northern Thailand may be novel. Fungal Divers. 2018, 93, 215–239. [Google Scholar] [CrossRef]
- Khuna, S.; Suwannarach, N.; Kumla, J.; Frisvad, J.C.; Matsui, K.; Nuangmek, W.; Lumyong, S. Growth enhancement of Arabidopsis (Arabidopsis thaliana) and onion (Allium cepa) with inoculation of three newly identified mineral-solubilizing fungi in the genus Aspergillus section Nigri. Front. Microbiol. 2021, 12, e705896. [Google Scholar] [CrossRef]
- Boonmee, S.; Wanasinghe, D.N.; Calabon, M.S.; Huanraluek, N.; Chandrasiri, A.K.U.; Jones, G.E.B.; Rossi, W.; Leonardi, M.; Singh, S.K.; Rana, S.; et al. Fungal diversity notes 1387–1511: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2021, 111, 1–335. [Google Scholar] [CrossRef]
- Selbmann, L.; Isola, D.; Egidi, E.; Zucconi, L.; Gueidan, C.; de Hoog, G.S.; Onofri, S. Mountain tips as reservoirs for new rock-fungal entities: Saxomyces gen. nov. and four new species from the Alps. Fungal Divers. 2014, 65, 167–182. [Google Scholar] [CrossRef] [Green Version]
- Voigt, K.; Wostemeyer, J. Reliable amplification of actin genes facilitates deep-level phylogeny. Microbiol. Res. 2000, 155, 179–195. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innes, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, e113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T. Bioedit Version 6.0.7. 2004. Available online: http://www.mbio.ncsu.edu/bioedit/bioedit.html (accessed on 20 November 2021).
- Zeng, J.S.; Sutton, D.A.; Fothergill, A.W.; Rinaldi, M.G.; Harrak, M.J.; de Hoog, G.S. Spectrum of clinically relevant Exophiala species in the United States. J. Clin. Microbiol. 2007, 45, 3713–3720. [Google Scholar] [CrossRef] [Green Version]
- Gueidan, C.; Villaseñor, C.R.; de Hoog, G.S.; Gorbushina, A.A.; Untereiner, W.A.; Lutzoni, F. A rock-inhabiting ancestor for mutualistic and pathogen-rich fungal lineages. Stud. Mycol. 2008, 61, 111–119. [Google Scholar] [CrossRef]
- Okada, G.; Seifert, K.A.; Takematsu, A.; Yamaoka, Y.; Miyazaki, S.; Tubaki, K. A molecular phylogenetic reappraisal of the Graphium complex based on 18 S rDNA sequences. Can. J. Bot. 1998, 76, 1495–1506. [Google Scholar] [CrossRef]
- Attili-Angelis, D.; Duarte AP, M.; Pagnocca, F.C.; Nagamoto, N.S.; De Vries, M.; Stielow, J.B.; de Hoog, G.S. Novel Phialophora species from leaf-cutting ants (tribe Attini). Fungal Divers. 2014, 65, 65–75. [Google Scholar] [CrossRef]
- Vicente, V.A.; Najafzadeh, M.J.; Sun, J.; Gomes, R.R.; Robl, D.; Marques, S.G.; Azevedo, C.M.P.S.; de Hoog, G.S. Environmental siblings of black agents of human chromoblastomycosis. Fungal Divers. 2014, 65, 47–63. [Google Scholar] [CrossRef]
- Untereiner, W.A.; Naveau, F.A. Molecular systematics of the Herpotrichiellaceae with an assessment of the phylogenetic positions of Exophiala dermatitidis and Phialophora americana. Mycologia 1999, 91, 67–83. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Réblová, M.; Untereiner, W.A.; Réblová, K. Novel evolutionary lineages revealed in the Chaetothyriales (fungi) based on multigene phylogenetic analyses and comparison of its secondary structure. PLoS ONE 2013, 8, e63547. [Google Scholar] [CrossRef] [PubMed]
- Vasse, M.; Voglmayr, H.; Mayer, V.; Gueidan, C.; Nepel, M.; Moreno, L.; de Hoog, S.; Selosse, M.A.; McKey, D.; Blatrix, R. A phylogenetic perspective on the association between ants (Hymenoptera: Formicidae) and black yeasts (Ascomycota: Chaetothyriales). Proc. R. Soc. B 2017, 284, e20162519. [Google Scholar] [CrossRef] [PubMed]
- Prenafeta-Boldú, F.X.; Summerbell, R.; de Hoog, G.S. Fungi growing on aromatic hydrocarbons: Biotechnology’s unexpected encounter with biohazard? FEMS Microbiol. Rev. 2006, 30, 109–130. [Google Scholar] [CrossRef] [Green Version]
- Tischner, Z.; Sebők, R.; Kredics, L.; Allaga, H.; Vargha, M.; Sebestyén, Á.; Dobolyi, C.; Kriszt, B.; Magyar, D. Mycological investigation of bottled water dispensers in healthcare facilities. Pathogens 2021, 10, e871. [Google Scholar] [CrossRef]
- Haase, G.; Sonntag, L.; van de Peer, Y.; Uijthof, J.M.; Podbielski, A.; Melzer-Krick, B. Phylogenetic analysis of ten black yeast species using nuclear small subunit rRNA gene sequences. Antonie van Leeuwenhoek 1995, 68, 19–33. [Google Scholar] [CrossRef]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding needles in haystacks: Linking scientific names, reference specimens and molecular data for fungi. Database 2014, 2014, bau061. [Google Scholar] [CrossRef]
- Cheewangkoon, R.; Groenewald, J.Z.; Summerell, B.A.; Hyde, K.D.; To-Anun, C.; Crous, P.W. Myrtaceae, a cache of fungal biodiversity. Persoonia 2009, 23, 55–85. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Braun, U.; Hunter, G.C.; Wingfield, M.J.; Verkley, G.J.; Shin, H.D.; Nakashima, C.; Groenewald, J.Z. Phylogenetic lineages in Pseudocercospora. Stud. Mycol. 2013, 75, 37–114. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Stamatakis, A. Raxml-vi-hpc: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Holder, M.T.; Vos, R.; Midford, P.E.; Liebowitz, T.; Chan, L.; Hoover, P.; Warnow, T. The CIPRES Portals. CIPRES, 2009. Available online: http://www.phylo.org/sub_sections/portal (accessed on 4 April 2022).
- Nylander, J.A.A. MrModeltest 2.0; Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree Tree Figure Drawing Tool Version 131, Institute of Evolutionary Biology, University of Edinburgh. Available online: http://treebioedacuk/software/figtree/ (accessed on 4 April 2022).
- Taj-Aldeen, S.J.; El Shafie, S.; Alsoub, H.; Eldeeb, Y.; de Hoog, G.S. Isolation of Exophiala dermatitidis from endotracheal aspirate of a cancer patient. Mycoses 2006, 49, 504–509. [Google Scholar] [CrossRef] [PubMed]
- McGinnis, M.R. Taxonomy of Exophiala jeanselmei (Langeron) McGinnis and Padhye. Mycopathologia 1978, 65, 79–87. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Isolation Resources | Location | Reference |
|---|---|---|---|
| Exophiala abietophila | Silver fir (Abies alba) | Norway | [21] |
| Exophiala alcalophila | Soil, soap container, washing machine, bathwater from households, and human skin | Brazil, Denmark, Germany, Japan, and Ukraine | [3,75,76] |
| Exophiala angulospora | Polluted soil, drinking water, Tilia wood, fish nursery, weedy seadragon, lumpfish skin and spleen, olive flounder (Paralichthys olivaceus), Atlantic cod (Gadus morhua), and human skin | Brazil, Denmark, Germany, Ireland, Japan, Netherlands, Norway, Russia, Scotland, and the USA | [3,77,78,79,80,81] |
| Exophiala aquamarina | Clown fish, leafy sea dragon, little tunnyfish, lumpfish, sand lance, weedy seadragon, and winter flounder | Canada, the UK, and the USA | [3,7] |
| Exophiala arunalokei | Subcutaneous lesion on human | India | [20] |
| Exophiala asiatica | Tonsil tissue of human | China | [30,32] |
| Exophiala attenuata | Soil, nasal granuloma of cat, cutaneous phaeohyphomycosis of cat, and human disease | Colombia, France, Germany, and the USA | [33,82,83,84] |
| Exophiala bergeri | Eye and skin of human | Brazil, Canada, Japan, Hong Kong, the UK, and the USA | [6,7,31,85] |
| Exophiala bonariae | Marble | Italy | [69] |
| Exophiala brunnea | Leaf of Acacia karroo | South Africa | [3] |
| Exophiala calicioides | Rotten wood | Japan | [68] |
| Exophiala campbellii | Subcutaneous lesion (foot ganglion) of human and human chest nodule | Germany and the UK | [7,70] |
| Exophiala cancerae | Water, water from tank, fruit drink, dialysis water Mangrove crab (Ucides cordatus), liver of green toad, sputum of human, and human finger | Australia, Brazil, Canada, Germany, Hong Kong, Israel, Netherlands, the UK, and the USA | [3,7,31,86,87,88] |
| Exophiala capensis | Leaf of Phaenocoma prolifera | Canada and South Africa | [7,30,31,32,33,68,69,70,82,83,84,85,86,87,88,89] |
| Exophiala castellanii | Decaying timber joinery, spoilt apple juice, drinking water, ice water, nematode, and human skin | Denmark, Germany, Netherlands, Sri Lanka, Switzerland, and the UK | [3,27] |
| Exophiala cinerea | Rock | China | [18] |
| Exophiala clavispora | Rock | China | [18] |
| Exophiala crusticola | Biological soil crust | the USA | [90] |
| Exophiala dermatitidis | Soil, dishwasher’s rubber, wood, internal organs of bat, chromoblastomycosis, knee fluid, lung, finger, and central nervous system fluid of human | Angola, Brazil, China, Finland, Germany, Hong Kong, Iran, Iraq, Japan, Korea, Malaysia, Mauritius, Qatar, Slovenia, South Africa, Taiwan, Thailand, Turkey, the UK, the USA, and Venezuela | [7,15,16,31,32,45,46,47,48,49,50,51,52,53] |
| Exophiala dopicola | Loblolly pine (Pinus taeda) | the USA | [91] |
| Exophiala ellipsoidea | Rock | China | [18] |
| Exophiala embothrii | Rhizosphere of Embothrium coccineum | Chile | [92] |
| Exophiala encephalarti | On leaves of Encephalartos transvenosus | South Africa | [93] |
| Exophiala equina | Soil, drinking water, bottled water, water from water machine, water system of packaging machine, wastewater, dialysis water bathroom-flask, bathroom-plate, silica gel, root mycorrhiza, Tilia root, Populus tremuloides, Cephalanthera damasonium, Phragmitis australis, Olea twig, nematode cyst (Heterodera sp.), subcutaneous infection of horse, Galapagos turtle, human stool, human sputum, human eye, and skin of human | Australia, Brazil, Canada, Denmark, Germany, Italy, Japan, Netherlands, Korea, the UK, and the USA | [3,7,10,87,94,95,96,97,98] |
| Exophiala eucalypti | Leaves of Eucalyptus sp. | South Africa | [99] |
| Exophiala eucalypticola | Leaf of Eucalyptus obliqua | Australia | [22] |
| Exophiala eucalyptorum | Leaf of Eucalyptus sp. | New Zealand | [23] |
| Exophiala exophialae | Soil, straw in armadillo’s burrow (Dasypus septemcinctus) | Colombia and Uruguay | [6,15,34] |
| Exophiala frigidotolerans | Soil | Ecuador | [100] |
| Exophiala halophila | Salty water, human skin, and human nail | Germany and the USA | [3] |
| Exophiala heteromorpha | Wood and human | Sweden and the USA | [7,15] |
| Exophiala hongkongensis | Big toenail infection of human | China and Hong Kong | [31] |
| Exophiala italica | Cytisus scoparius on dead branch | Italy | [24] |
| Exophiala jeanselmei | Subcutaneous abscesses, skin lesion, eumycetoma of human, peritoneal dialysis fluid, human blood, human sputum, and human eye | Australia, Bangladesh, Brazil, Canada, China, Costa Rica, France, Hong Kong, Jamaica, Japan, Martinique, Pakistan, Paraguay, Peru, Philippines, Saudi Arabia, Thailand, Trinidad, the UK, Uruguay, and the USA | [6,17,27,31,32,33,34,54,55,56,57,58,59,60,61,62,63,64,65,66,67] |
| Exophiala lacus | Lake water and river sediments | Netherlands and Spain | [3,71] |
| Exophiala lavatrina | Domestic bathroom | the UK | [7] |
| Exophiala lecanii-corni | Lecanium corni, domestic bathroom, dialysis fluid, subcutaneous abscess, knee cyst, skin lesion, eye sclera, finger fluid, skin scales, and human nail | Austria, Germany, Hong Kong, Japan, Netherlands, the UK, and the USA | [27,31,57,101,102] |
| Exophiala lignicola | Quercus sp. | Ukraine | [21] |
| Exophiala macquariensis | Island soil | Australia | [2] |
| Exophiala mali | Inner fruit tissue of Malus sp. | South Africa | [92] |
| Exophiala mansonii | Populus tremula | Sweden | [15] |
| Exophiala mesophila | Shower joint, swimming pool, dental waterline, bathroom, contact lens, phaeohyphomycotic cyst, subcutaneous nodule biopsy, immunosuppressed, bronchial endoscopy, finger, sinus, hip joint, hair, and nasal tissue of human | Brazil, France, Germany, Netherlands, the UK, and the USA | [3,7,85] |
| Exophiala moniliae | Branch of Quercus sp., sludge in bathroom drainpipes, and medicated bathwater | Australia, Japan, and Russia | [15,76,103] |
| Exophiala nagquensis | Rock | China and Tibet | [18] |
| Exophiala nidicola | Nest of bird | Spain | [72] |
| Exophiala nishimurae | Bark and human skin | the USA and Venezuela | [17,33] |
| Exophiala oligosperma | Soil, wood, swimming pool, water, polluted water, river sediments, sauna, silicone solution, ear swab, plastic foil, prosthetic contact lenses, cerebral mycosis, subcutaneous abscess, thigh abscess, skin lesion, sphenoid tumor, lung, sinus, and human sputum | Austria, Brazil, Canada, Finland, France, Germany, Hong Kong, Italy, Japan, Netherlands, Spain, Switzerland, the UK, Ukraine, the USA, and Venezuela | [6,7,17,31,34,57,71,104,105,106] |
| Exophiala opportunistica | Drinking water, rhizosphere (Triticum aestivum), polyvinyl alcohol, human nail, and human foot | Australia, Denmark, Germany, and Netherlands | [3] |
| Exophiala palmae | Decaying shell of babassu coconut (Orbignya phalerata) | Brazil | [107] |
| Exophiala phaeomuriformis | Natural hot spring, sauna, tile floor of swimming pool, bathroom tap, bathroom sink, cutaneous mycosis, blood culture, external ear channel, oral mucosa, nail, and human sputum, | Austria, Canada, Czech Republic, Germany, Japan, Netherlands, Slovenia, the UK, and the USA | [7,16,108] |
| Exophiala pisciphila | Swimming pool, water pipe, dialysis water, catfish (Ictalurus punctatus), Potbelly seahorse, crocodile, and human | Brazil, Germany, Japan, Israel, and the USA | [3,7,87,109] |
| Exophiala polymorpha | Subcutaneous lesion of human | the USA | [8] |
| Exophiala prostantherae | Leaves of Prostanthera sp. | Australia | [92] |
| Exophiala pseudooligosperma | Karst rocky desertification mountain soil | China | [4] |
| Exophiala psychrophila | Atlantic salmon smolt (Salmo salar) | Ireland and Norway | [3] |
| Exophiala quercina | Dead wood of Quercus sp. | Germany | [73] |
| Exophiala radicis | Soil, root endophyte of Microthlaspi perfoliatum, plant roots, Olea sp. twig, nematode cyst (Heterodera sp.), toenail, tinea on leg, and foot of human | Bulgaria, Denmark, France, Germany, Italy, the Netherlands, and Spain | [5,71] |
| Exophiala salmonis | Drinking water, drinking water tap and cerebral mycetoma of fingerling trout (Salmo clarkii) | Canada and the Netherlands | [1] |
| Exophiala sideris | Oak railway tie, creosoted tie, gold mine, and surface of wild berries of Sorbus aucuparia | the Netherlands and Poland | [9] |
| Exophiala spartinae | Spartina alterniflora root tissue in saltwater marsh | the USA | [110] |
| Exophiala spinifera | Soil, palm tree, wood, nest of Anumbius annumbi, armadillo burrow, maize kernel, apple juice, rotten cactus, skin lesion, foot abscess, neck lymph node, human sputum, and bark nasal granuloma of human | Antarctic, Argentina, Australia, Brazil, China, Colombia, Egypt, Germany, India, Mexico, Papua New Guinea, Senegal, Thailand, the UK, Uruguay, the USA, and Venezuela | [6,32,33,34,35,36,37,38,39,40,41,42,43,44] |
| Exophiala tremulae | Populus tremuloides roots | Canada | [111] |
| Exophiala xenobiotica | Soil, wood, oil sludge, chromoblastomycosis on back, phaeomycotic cyst, subcutaneous cyst, elbow pus, and skin lesions | Antarctic, Australia, Brazil, Canada, Germany, Hong Kong, Japan, the Netherlands, New Zealand, Switzerland, Sweden, the UK, the USA, and Venezuela | [6,7,27,31,34,57,74] |
| Target Gene | Primer | Primer Sequence (5′–3′) | Annealing Temperature (°C) | Reference |
|---|---|---|---|---|
| act | Act1 | TGGGACGATATGGAIAAIATCTGGCA | 52 | [124] |
| Act5ra | TTAGAAGCACTTNCGGTG | 52 | [124] | |
| ITS | ITS4 | TCCTCCGCTTATTGATATGC | 55 | [125] |
| ITS5 | GGAAGTAAAAGTCGTAACAAGG | 55 | [125] | |
| nrSSU | NS1 | GTAGTCATATGCTTGTCTC | 55 | [125] |
| NS4 | CTTCCGTCAATTCCTTTAAG | 55 | [125] | |
| tef | EF1-728F | CATCGAGAAGTTCGAGAAGG | 57 | [126] |
| EF1-986R | TACTTGAAGGAACCCTTACC | 57 | [126] | |
| tub | Bt2a | GGTAACCAAATCGGTGCTGCTTTC | 52 | [127] |
| Bt2b | ACCCTCAGTGTAGTGACCCTTGGC | 52 | [127] |
| Species | Strains | GenBank Accession No. | References | ||||
|---|---|---|---|---|---|---|---|
| ITS | nrSSU | tub | tef | act | |||
| Exophiala abietophila | CBS 145038 T | NR163357 | – | – | – | – | [21] |
| CBS 520.82 T | JF747041 | JN856010 | JN112423 | JN128771 | JN112379 | [3] | |
| CBS 122256 | JF747044 | – | JN112425 | JN128773 | JN112381 | [3] | |
| Exophiala angulospora | CBS 482.92 T | JF747046 | JN856011 | JN112426 | JN128780 | JN112383 | [3] |
| CBS 120272 | JF747045 | – | JN112427 | JN128781 | JN112382 | [3] | |
| Exophiala aquamarina | CBS 119918 T | JF747054 | JN856012 | JN112434 | – | JN112388 | [3] |
| CBS 119916 | JF747055 | – | JN112435 | – | JN112389 | [3] | |
| Exophiala arunalokei | NCCPF106033 | MW724320 | – | – | – | – | [20] |
| Exophiala asiatica | CBS 122847 T | NR111332 | – | – | – | – | [30] |
| CBS 122848 | MW222182 | – | – | – | – | [30] | |
| Exophiala attenuata | CBS 101540 T | AF549446 | – | – | – | – | [33] |
| UTHSC87-80 | EF025392 | – | – | – | – | [130] | |
| Exophiala bergeri | CBS 353.52 T | EF551462 | FJ358308 | EF551497 | EF551524 | EF551464 | [131] |
| Exophiala bonariae | CBS 139957 T | JX681046 | – | – | – | – | [69] |
| Exophiala brunnea | CBS 587.66 T | JF747062 | JN856013 | JN112442 | JN128783 | JN112393 | [3] |
| Exophiala calicioides | JCM6030 | – | AB007655 | – | – | – | [132] |
| Exophiala campbellii | NCPF 2274 | LT594703 | – | – | LT594739 | – | [7] |
| Exophiala cancerae | CBS 120420 T | JF747064 | – | JN112444 | JN128800 | JN112394 | [3] |
| CBS 117491 | KF928439 | – | KF928567 | JN128799 | JN112396 | [3] | |
| Exophiala capensis | CBS 128771 T | JF499841 | – | – | – | – | [89] |
| Exophiala castellanii | CBS 158.58 T | JF747070 | JN856014 | KF928586 | JN128766 | – | [3,133] |
| CBS 120913 | JF747144 | – | JN112506 | JN128750 | – | [3] | |
| Exophiala cinerea | CGMCC 3.18778 T | MG012695 | MG012724 | MG012745 | MG012704 | MG012714 | [18] |
| CGMCC 3.18779 | MG012696 | MG012725 | MG012746 | MG012705 | MG012715 | [18] | |
| Exophiala clavispora | CGMCC 3.17512 | KP347940 | MG012733 | KP347931 | KP347909 | MG012712 | [18] |
| CGMCC 3.17517 T | KP347942 | KP347967 | KP347932 | KP347911 | KP347893 | [18] | |
| Exophiala crusticola | CBS 119970 T | AM048755 | KF155199 | – | – | – | [90,134] |
| HM136 | MK281393 | – | – | – | – | Unpublished | |
| Exophiala dermatitidis | CBS 207.35 T | AF050269 | – | KF928572 | – | – | [133,134,135] |
| CBS 120473 | MF320159 | – | MF320217 | MF320196 | – | [43] | |
| Exophiala dopicola | CBS 537.94 T | MH862483 | – | – | – | – | [136] |
| Exophiala ellipsoidea | CGMCC 3.17348 T | KP347955 | KP347965 | KP347921 | KP347901 | MG012713 | [18] |
| CGMCC 3.17522 | KP347954 | MG012735 | KP347919 | – | KP347884 | [18] | |
| Exophiala embothrii | CBS 146558 T | NR171982 | – | MW055976 | MW055980 | – | [92] |
| Exophiala encephalarti | CBS 128210 T | HQ599588 | – | – | – | – | [93] |
| Exophiala equina | CBS 119.23 T | JF747094 | JN856017 | JN112462 | JN128814 | JN112401 | [3] |
| CBS 120906 | JF747093 | – | JN112461 | JN128813 | JN112400 | [3] | |
| Exophiala eucalypti | CBS 142069 | KY173411 | – | – | – | – | [99] |
| Exophiala eucalypticola | CBS 143412 T | NR158438 | – | MH108039 | MH108016 | – | [22] |
| Exophiala eucalyptorum | CBS 121638 T | NR132882 | KC455302 | KC455228 | – | – | [137] |
| CPC 11261 | EU035417 | – | – | – | – | [23] | |
| Exophiala exophialae | CBS 668.76 T | AY156973 | KX822287 | EF551499 | EF551526 | EF551466 | [33,138] |
| CBS 671.76 | AY156975 | – | EF551500 | EF551525 | EF551467 | [33] | |
| Exophiala frigidotolerans | CBS 146539 T | LR699566 | – | – | – | – | [100] |
| Exophiala halophila | CBS 121512 T | NR111628 | NG062077 | JN112473 | JN128774 | – | [3] |
| Exophiala heteromorpha | CBS 232.33 T | AY857524 | – | – | – | – | [139] |
| U THSC87-67 | EF025400 | – | – | – | – | [130] | |
| Exophiala hongkongensis | CBS 131511 | JN625231 | – | JN625236 | JN625246 | JN625241 | [31] |
| Exophiala italica | MFLUCC16-0245 T | KY496744 | KY501114 | – | KY514393 | - | [24] |
| Exophiala jeanselmei | CBS 507.90 T | AY156963 | FJ358310 | EF551501 | EF551530 | - | [33,131] |
| CBS 528.76 | AY857530 | – | EF551502 | EF551531 | EF551469 | [139] | |
| Exophiala lacus | FMR 3995 | KU705830 | – | – | – | – | [71] |
| CBS 117497 T | JF747110 | – | – | JN128776 | JN112407 | [3] | |
| Exophiala lamphunensis | SDBR-CMU404 T | ON555798 | ON555813 | ON544227 | ON544242 | ON544257 | This study |
| SDBR-CMU405 | ON555799 | ON555814 | ON544228 | ON544243 | ON544258 | This study | |
| SDBR-CMU406 | ON555800 | ON555815 | ON544229 | ON544244 | ON544259 | This study | |
| SDBR-CMU407 | ON555801 | ON555816 | ON544230 | ON544245 | ON544260 | This study | |
| SDBR-CMU408 | ON555802 | ON555817 | ON544231 | ON544246 | ON544261 | This study | |
| Exophiala lapidea | SDBR-CMU409 T | ON555803 | ON555818 | ON544232 | ON544247 | ON544262 | This study |
| SDBR-CMU410 | ON555804 | ON555819 | ON544233 | ON544248 | ON544263 | This study | |
| SDBR-CMU411 | ON555805 | ON555820 | ON544234 | ON544249 | ON544264 | This study | |
| SDBR-CMU412 | ON555806 | ON555821 | ON544235 | ON544250 | ON544265 | This study | |
| SDBR-CMU413 | ON555807 | ON555822 | ON544236 | ON544251 | ON544266 | This study | |
| SDBR-CMU414 | ON555808 | ON555823 | ON544237 | ON544252 | ON544267 | This study | |
| Exophiala lavatrina | NCPF 7893 | LT594696 | – | – | LT594729 | – | [7] |
| NCPF 7898 | LT594697 | – | – | LT594731 | – | [7] | |
| Exophiala lecanii-corni | CBS 123.33 T | AY857528 | FJ358311 | – | – | – | [131,139] |
| B2242C | MT320770 | – | – | MZ190330 | – | [140] | |
| Exophiala lignicola | CBS:144622 T | NR163358 | – | – | MK442694 | – | [21] |
| Exophiala macquariensis | CBS 144232 T | MF619956 | – | MH297438 | MH297439 | – | [2] |
| Exophiala mali | CBS 146791 T | MW175341 | – | – | – | – | [92] |
| Exophiala mansonii | CBS 101.67 T | AF050247 | X79318 | – | – | – | [135,141] |
| Exophiala mesophila | CBS 402.95 T | JF747111 | JN856016 | JN112476 | JN128761 | – | [3] |
| CBS 119910 | JF747113 | – | JN112478 | JN128753 | – | [3] | |
| Exophiala moniliae | CBS 520.76 T | KF881967 | – | – | – | – | Unpublished |
| BMU00283 | MW222184 | – | – | – | – | Unpublished | |
| Exophiala nagquensis | CGMCC 3.17333 T | KP347948 | KP347970 | KP347924 | KP347914 | KP347895 | [18] |
| CGMCC 3.17334 | KP347949 | MG012741 | KP347923 | KP347915 | KP347896 | [18] | |
| Exophiala nidicola | CBS 138589 T | NR161045 | – | – | – | – | [72] |
| Exophiala nishimurae | CBS 101538 T | AY163560 | KX822288 | JX482552 | EF551523 | JX482553 | [33] |
| Exophiala oligosperma | CBS 725.88 T | AY163551 | FJ358313 | EF551508 | EF551534 | EF551474 | [17,131] |
| CBS 265.49 | MH856519 | – | EF551507 | EF551536 | EF551473 | [136] | |
| Exophiala opportunistica | CBS 109811 T | JF747123 | – | JN112486 | JN128792 | JN112408 | [3] |
| Exophiala palmae | CMRP1196 T | KY680434 | – | KY689829 | – | – | [107] |
| CMRP1207 | KY680433 | – | KY689828 | – | – | [107] | |
| Exophiala phaeomuriformis | CBS 131.88 T | AJ244259 | – | – | – | – | Unpublished |
| Exophiala pisciphila | CBS 537.73 T | NR121269 | JN856018 | JN112493 | JN128788 | JN112412 | [3,142] |
| CBS 121500 | JF747134 | – | JN112496 | JN128789 | JN112414 | [3] | |
| Exophiala polymorpha | CBS 138920 T | KP070763 | – | – | – | – | [8] |
| Exophiala prostantherae | CBS 146794 T | NR171990 | – | – | – | – | [92] |
| Exophiala pseudooligosperma | YMFT 1.6741 | MW616557 | MW616558 | MZ127830 | – | – | [4] |
| Exophiala psychrophila | CBS 191.87 T | JF747135 | JN856019 | JN112497 | JN128798 | – | [3] |
| CBS 256.92 | JF747136 | – | JN112498 | – | – | [3] | |
| Exophiala quercina | CBS 146024 T | NR170053 | – | – | MT223713 | – | [73] |
| Exophiala radicis | P2854 T | KT099204 | KT723453 | KT723463 | KT723458 | KT723443 | [5] |
| Exophiala salmonis | CBS 157.67 T | AF050274 | JN856020 | JN112499 | JN128747 | JN112415 | [3,135] |
| CBS 120274 | JF747138 | – | KF928562 | JN128802 | JN112416 | [3] | |
| Exophiala saxicola | SDBR-CMU415 T | ON555809 | ON555824 | ON544238 | ON544253 | ON544268 | This study |
| SDBR-CMU416 | ON555810 | ON555825 | ON544239 | ON544254 | ON544269 | This study | |
| Exophiala siamensis | SDBR-CMU417 T | ON555811 | ON555826 | ON544240 | ON544255 | ON544270 | This study |
| SDBR-CMU418 | ON555812 | ON555827 | ON544241 | ON544256 | ON544271 | This study | |
| Exophiala sideris | CBS 121818 T | HQ452311 | HQ441174 | HQ535833 | HQ452336 | – | [9] |
| CBS 127096 | HQ452312 | HQ441175 | HQ535834 | HQ452337 | – | [9] | |
| Exophiala spartinae | CBS 147266 T | NR174648 | – | – | – | – | [110] |
| Exophiala spinifera | CBS 899.68 T | AY156976 | – | EF551516 | EF551541 | EF551482 | [33] |
| Exophiala tremulae | CBS 129355 T | FJ665274 | KT894147 | KT894148 | KT894149 | KT894146 | [5,89] |
| Exophiala xenobiotica | CBS 118157 T | DQ182587 | – | – | – | – | [27] |
| CBS 117646 | KP132146 | – | – | – | – | [27] | |
| Cyphellophora eucalypti | CBS:124764 T | GQ303274 | NG062860 | KF928601 | GU384510 | JQ325009 | [133,137,143,144] |
| Cyphellophora fusarioides | MUCL 44033 | NR132879 | NG065006 | KC455224 | – | – | [137] |
| Fungal Strains | Colony Diameter (mm) * | |||||||
|---|---|---|---|---|---|---|---|---|
| 10 °C | 15 °C | 20 °C | 25 °C | 28 °C | 30 °C | 35 °C | 37 °C | |
| SDBR-CMU404 | 10.25 ± 0.27 | 17.92 ± 0.92 | 18.17 ± 0.52 | 23.83 ± 0.41 | 24.42 ± 0.58 | 22.83 ± 0.68 | 13.08 ± 0.58 | 8.00 ± 0.55 |
| SDBR-CMU405 | 10.54 ± 0.33 | 17.88 ± 0.56 | 19.08 ± 0.12 | 24.27 ± 0.42 | 24.85 ± 0.57 | 23.09 ± 0.41 | 12.47 ± 0.52 | 8.12 ± 0.14 |
| SDBR-CMU406 | 11.42 ± 0.52 | 16.12 ± 0.16 | 19.78 ± 0.72 | 24.05 ± 0.97 | 25.41 ± 0.44 | 22.79 ± 0.85 | 12.55 ± 0.55 | 7.36 ± 0.22 |
| SDBR-CMU407 | 11.25 ± 0.42 | 16.33 ± 0.41 | 19.25 ± 0.52 | 25.58 ± 0.58 | 25.92 ± 0.38 | 23.17 ± 0.52 | 12.33 ± 0.41 | 6.92 ± 0.20 |
| SDBR-CMU408 | 10.12 ± 0.22 | 16.45 ± 0.87 | 18.44 ± 0.61 | 25.03 ± 0.45 | 25.25 ± 0.62 | 22.81 ± 0.43 | 12.74 ± 0.40 | 7.45 ± 0.39 |
| SDBR-CMU409 | 17.75 ± 0.27 | 20.08 ± 1.07 | 28.33 ± 0.98 | 36.24 ± 1.44 | 37.00 ± 1.26 | 40.83 ± 1.33 | 10.08 ± 0.38 | 6.08 ± 0.20 |
| SDBR-CMU410 | 16.11 ± 0.18 | 24.35 ± 0.84 | 26.65 ± 0.88 | 35.91 ± 1.36 | 36.02 ± 1.31 | 38.42 ± 1.44 | 8.27 ± 0.45 | 5.96 ± 0.22 |
| SDBR-CMU411 | 14.98 ± 0.12 | 20.03 ± 0.41 | 27.78 ± 1.23 | 34.78 ± 0.97 | 35.43 ± 1.28 | 36.92 ± 1.96 | 9.04 ± 0.36 | 5.23 ± 0.27 |
| SDBR-CMU412 | 15.97 ± 0.52 | 26.27 ± 0.92 | 27.56 ± 0.71 | 36.77 ± 1.22 | 37.11 ± 1.45 | 38.82 ± 0.79 | 9.19 ± 0.24 | 5.71 ± 0.13 |
| SDBR-CMU413 | 14.42 ± 0.38 | 19.25 ± 0.27 | 25.08 ± 1.07 | 32.25 ± 1.44 | 34.33 ± 2.04 | 35.42 ± 0.86 | 8.42 ± 0.49 | 5.58 ± 0.49 |
| SDBR-CMU414 | 14.23 ± 0.47 | 25.78 ± 0.74 | 25.71 ± 0.88 | 35.04 ± 1.47 | 35.47 ± 1.42 | 36.96 ± 0.65 | 10.12 ± 0.56 | 5.44 ± 0.39 |
| SDBR-CMU415 | 9.92 ± 0.20 | 14.42 ± 0.49 | 15.50 ± 0.55 | 21.33 ± 0.26 | 23.75 ± 1.37 | 24.17 ± 1.66 | 12.75 ± 0.27 | 8.92 ± 0.20 |
| SDBR-CMU416 | 9.83 ± 0.41 | 14.08 ± 0.20 | 16.58 ± 0.38 | 21.75 ± 1.13 | 24.67 ± 0.41 | 26.88 ± 1.28 | 11.42 ± 0.20 | 8.17 ± 0.26 |
| SDBR-CMU417 | 8.08 ± 0.38 | 10.08 ± 0.49 | 11.75 ± 1.17 | 10.42 ± 0.80 | 9.42 ± 0.58 | 7.33 ± 0.26 | – | – |
| SDBR-CMU418 | 8.08 ± 0.38 | 10.50 ± 0.77 | 11.83 ± 0.68 | 10.35 ± 1.17 | 9.92 ± 0.20 | 7.58 ± 0.20 | – | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thitla, T.; Kumla, J.; Khuna, S.; Lumyong, S.; Suwannarach, N. Species Diversity, Distribution, and Phylogeny of Exophiala with the Addition of Four New Species from Thailand. J. Fungi 2022, 8, 766. https://doi.org/10.3390/jof8080766
Thitla T, Kumla J, Khuna S, Lumyong S, Suwannarach N. Species Diversity, Distribution, and Phylogeny of Exophiala with the Addition of Four New Species from Thailand. Journal of Fungi. 2022; 8(8):766. https://doi.org/10.3390/jof8080766
Chicago/Turabian StyleThitla, Tanapol, Jaturong Kumla, Surapong Khuna, Saisamorn Lumyong, and Nakarin Suwannarach. 2022. "Species Diversity, Distribution, and Phylogeny of Exophiala with the Addition of Four New Species from Thailand" Journal of Fungi 8, no. 8: 766. https://doi.org/10.3390/jof8080766