
Two Novel Genera, Neostemphylium and Scleromyces (Pleosporaceae) from Freshwater Sediments and Their Global Biogeography


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolates
2.2. Phenotypic Study
2.3. DNA Extraction, Sequencing and Phylogenetic Analysis
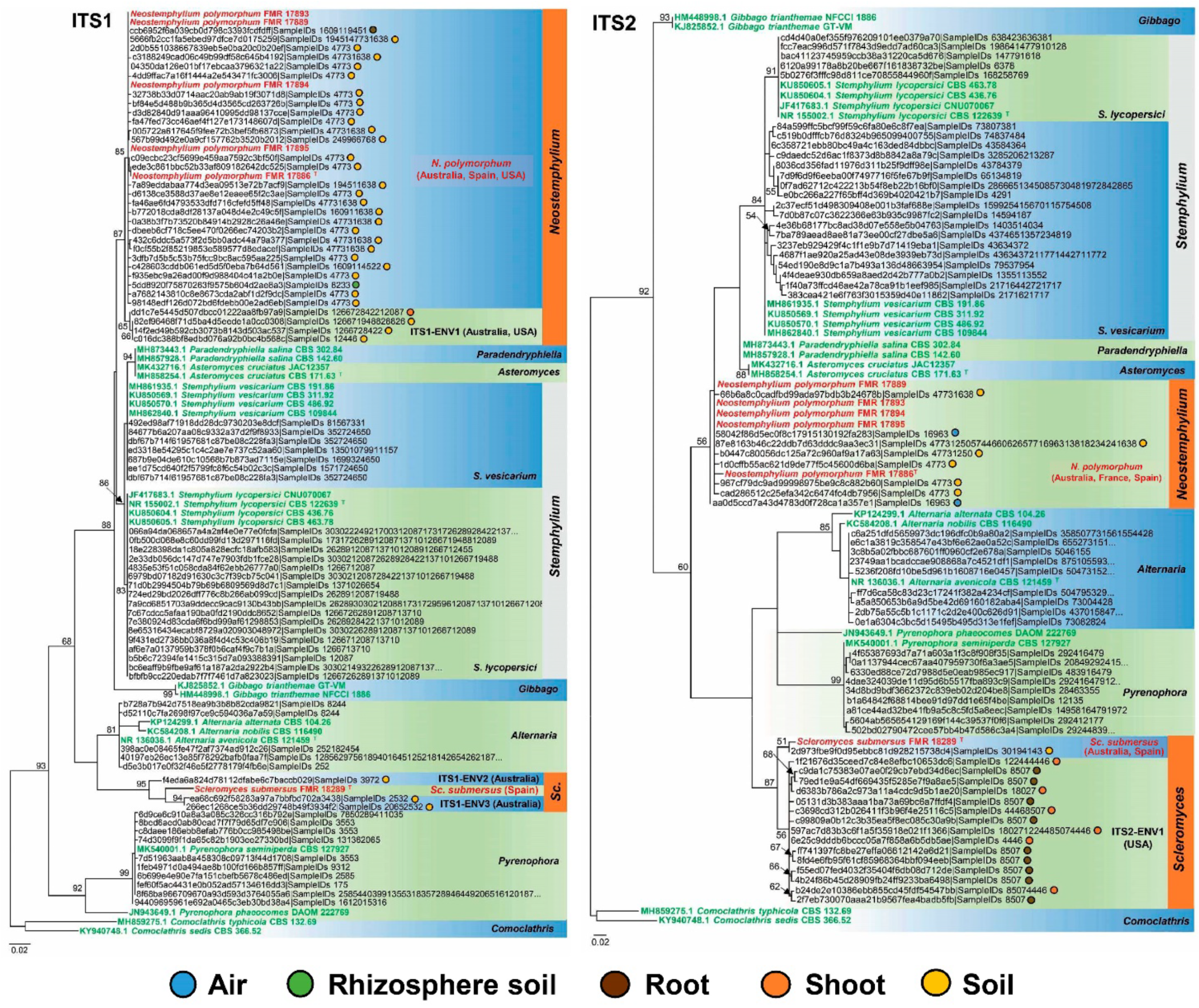
2.4. Phylogeny and Geographic Distribution of Allied Environmental Sequences
3. Results
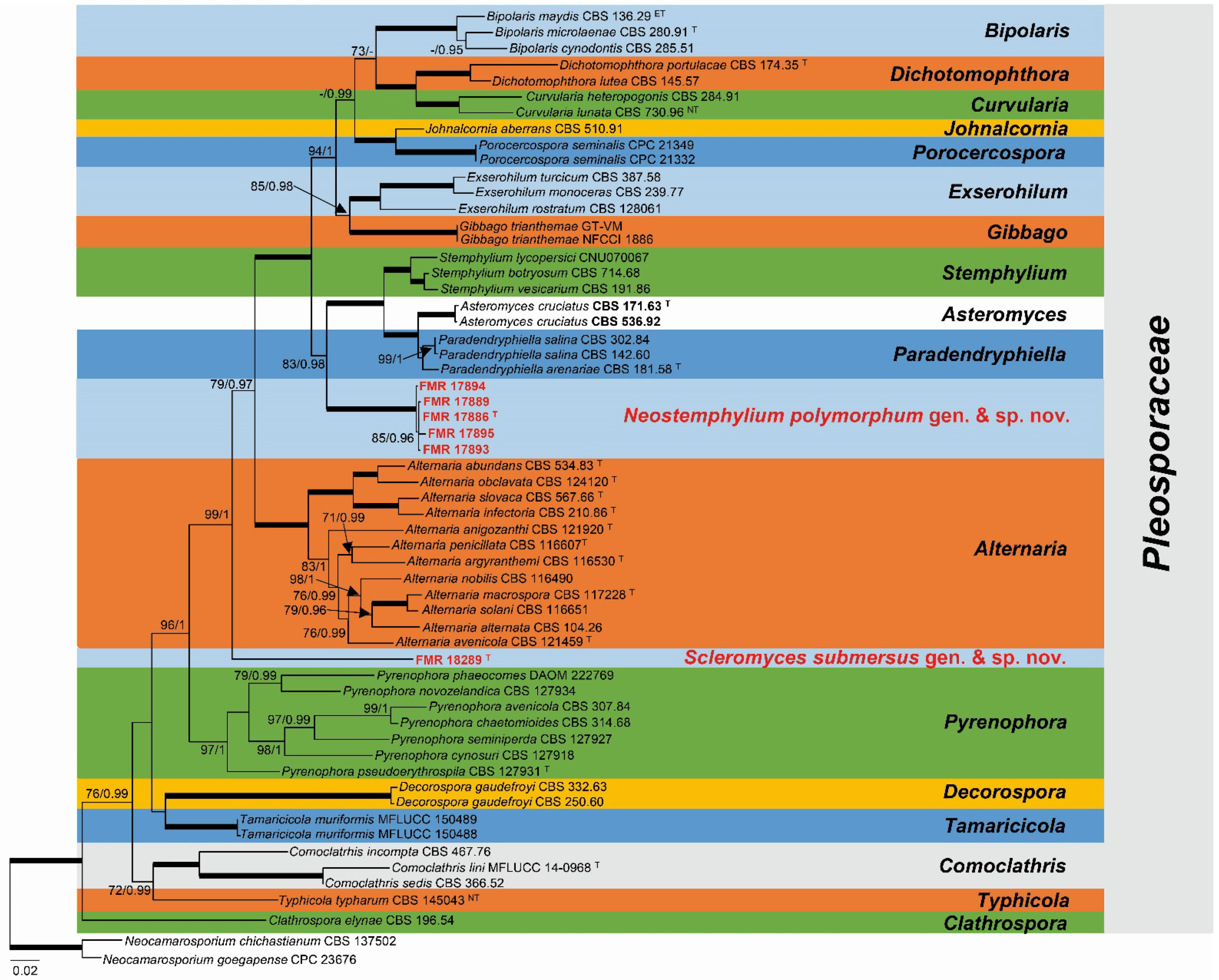
3.1. Phylogeny
3.2. Biogeography and Ecology
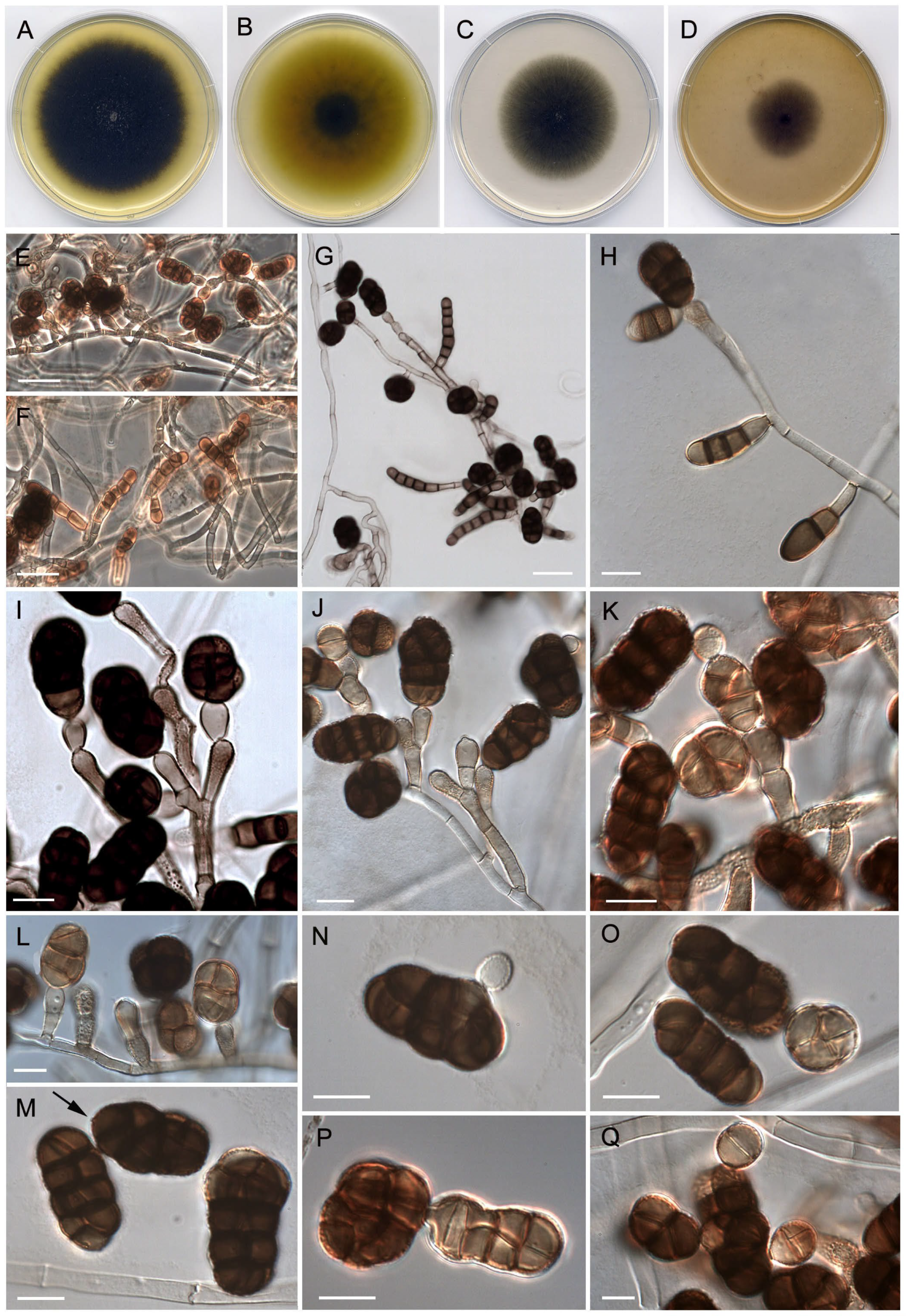
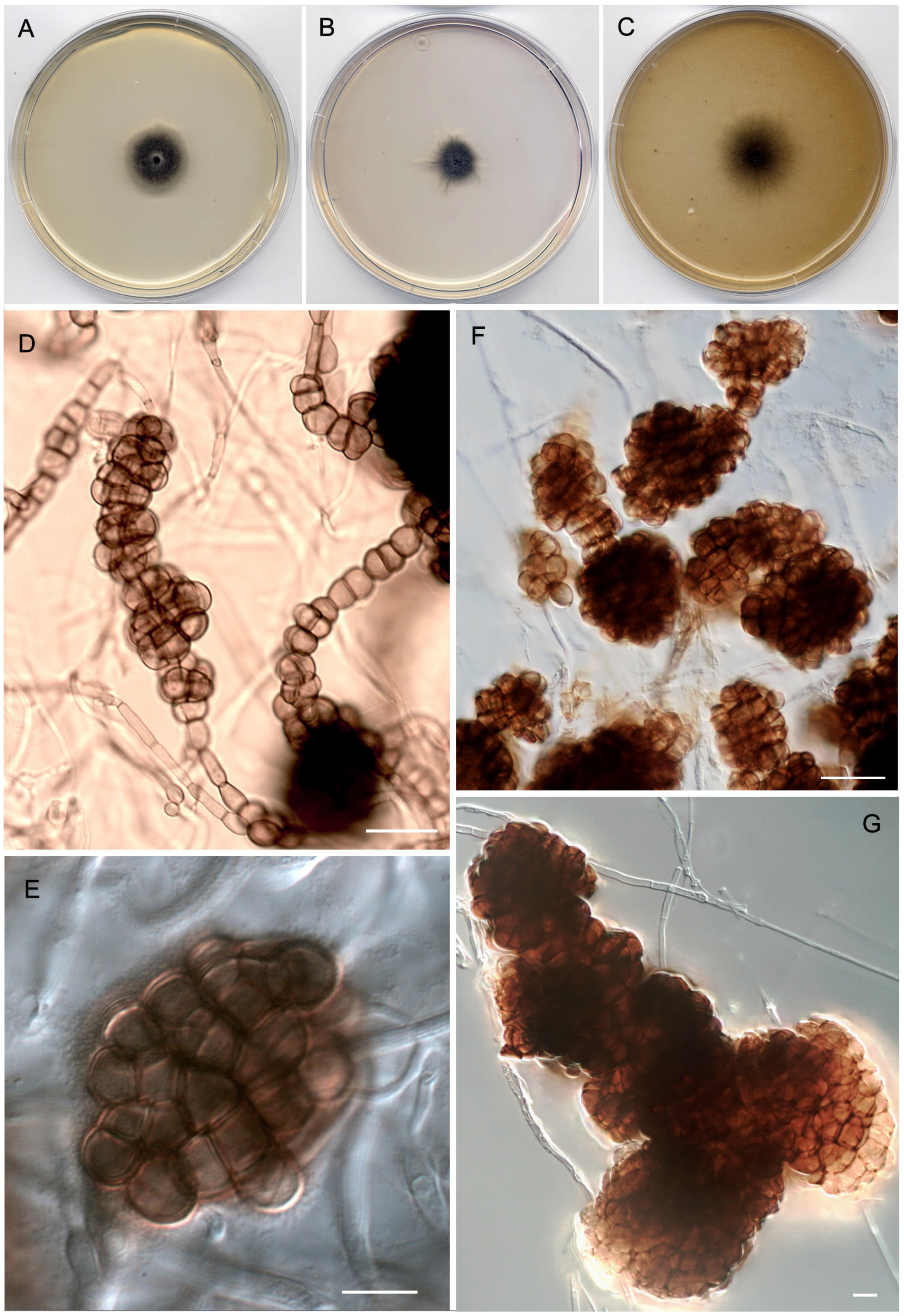
3.3. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nitschke, T.R.J. Grundlage eines systems der Pyrenomyceten. Verhandlungen des Naturhistorischen Vereins der Preussischen Rheinlande. Westfalens und des Regierungsbezirks Osnabrück 1869, 26, 70–77. [Google Scholar]
- Kodsueb, R.; Dhanasekaran, V.; Aptroot, A.; Lumyong, S.; McKenzie, E.H.C.; Hyde, K.D.; Jeewon, R. The family Pleosporaceae: Intergeneric relationships and phylogenetic perspectives based on sequence analyses of partial 28S rDNA. Mycologia 2006, 98, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Xie, N. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Fungal Divers. 2020, 105, 17–318. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Deng, C. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Zhang, Y.; Crous, P.W.; Schoch, C.L.; Hyde, K.D. Pleosporales. Fungal Divers. 2012, 52, 1–221. [Google Scholar] [CrossRef]
- McNeill, J.; Barrie, F.R.; Buck, W.R.; Demoulin, V.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Marhold, K.; Prado, J.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Melbourne Code): Appendices II-VIII; Turland, N., Ed.; Regnum Vegetabile: Melbourne, Australia, 2012; Volume 154, p. 492. [Google Scholar]
- Rossman, A.Y.; Crous, P.W.; Hyde, K.D.; Hawksworth, D.L.; Aptroot, A.; Bezerra, J.L.; Bhat, J.D.; Boehm, E.; Braun, U.; Boonmee, S.; et al. Recommended names for pleomorphic genera in Dothideomycetes. IMA Fungus 2015, 6, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Ariyawansa, H.A.; Thambugala, K.M.; Manamgoda, D.S.; Jayawardena, R.; Camporesi, E.; Boonmee, S.; Wanasinghe, D.N.; Phookamsak, R.; Hongsanan, S.; Singtripop, C.; et al. Towards a natural classification and backbone tree for Pleosporaceae. Fungal Divers. 2015, 71, 85–139. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Hanse, B.; Van Leeuwen, G.C.M.; Groenewald, J.Z.; Crous, P.W. Stemphylium revisited. Stud. Mycol. 2017, 87, 77–103. [Google Scholar] [CrossRef]
- Elkhateeb, W.A.; Kolaibe, A.G.A.; Elkhateeb, A.; Daba, G.M. Allergen, pathogen, or biotechnological tool? The dematiaceous fungi Alternaria what’s for it and what’s on it? J. Pharm. Pharmacol. Res. 2021, 4, 1–6. [Google Scholar] [CrossRef]
- De Hoog, G.S.; Guarro, J.; Gené, J.; Ahmed, S.; Al-Hatmi, A.M.S.; Figueras, M.; Vitale, R.G. Atlas of Clinical Fungi: The Ultimate Benchtool for Diagnostics. Introductions, Lower Fungi, Basidiomicetes, Yeasts, Filamentous Ascomycetes, 4th ed.; Foundation Atlas: Hilversum, The Netherlands, 2020; p. 776. [Google Scholar]
- Lepère, C.; Domaizon, I.; Humbert, J.F.; Hugoni, M.; Debroas, D. Diversity, spatial distribution and activity of fungi in freshwater ecosystems. PeerJ. 2019, 7, e6247. [Google Scholar] [CrossRef]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun. 2019, 10, 2369. [Google Scholar] [CrossRef]
- Mohapatra, M.; Yadav, R.; Rajput, V.; Dharne, M.S.; Rastogi, G. Metagenomic analysis reveals genetic insights on biogeochemical cycling, xenobiotic degradation, and stress resistance in mudflat microbiome. J. Environ. Manag. 2021, 292, 112738. [Google Scholar] [CrossRef] [PubMed]
- Chauvet, E.; Cornut, J.; Sridhar, K.R.; Selosse, M.-A.; Bärlocher, F. Beyond the water column: Aquatic hyphomycetes outside their preferred habitat. Fungal Ecol. 2016, 19, 112–127. [Google Scholar] [CrossRef]
- Su, H.-Y.; Udayanga, D.; Luo, Z.; Manamgoda, D.; Zhao, Y.-C.; Yang, J.; Hyde, K. Hyphomycetes from aquatic habitats in Southern China: Species of Curvularia (Pleosporaceae) and Phragmocephala (Melannomataceae). Phytotaxa 2015, 226, 201. [Google Scholar] [CrossRef]
- Kambura, A.K.; Mwirichia, R.K.; Kasili, R.W.; Karanja, E.N.; Makonde, H.M.; Boga, H.I. Diversity of fungi in sediments and water sampled from the hot springs of Lake Magadi and Little Magadi in Kenya. Afr. J. Microbiol. Res. 2016, 10, 330–338. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Větrovský, T.; Morais, D.; Kohout, P.; Lepinay, C.; Algora, C.; Hollá, S.A.; Bahnmann, B.D.; Bílohnědá, K.; Brabcová, V.; D’Alò, F.; et al. GlobalFungi, a global database of fungal occurrences from high-throughput-sequencing metabarcoding studies. Sci. Data 2020, 7, 228. [Google Scholar] [CrossRef]
- Ulfig, K.; Guarro, J.; Cano, J.; Gené, J.; Vidal, P.; Figueras, M.J. General assessment of the occurrence of keratinolytic fungi in river and marine beach sediments of Catalonian waters (Spain). Water Air Soil Pollut. 1997, 94, 275–287. [Google Scholar] [CrossRef]
- Chupp, C. Further notes on double cover-glass mounts. Mycologia 1940, 32, 269–270. Available online: https://10.2307/3754504 (accessed on 12 January 2022).
- Nishikawa, J.; Nakashima, C. Japanese species of Alternaria and their species boundaries based on host range. Fungal Syst. Evol. 2020, 5, 197–282. [Google Scholar] [CrossRef] [PubMed]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Methuen: London, UK, 1978. [Google Scholar]
- Müller, F.M.; Werner, K.E.; Kasai, M.; Francesconi, A.; Chanock, S.J.; Walsh, T.J. Rapid extraction of genomic DNA from medically important yeasts and filamentous fungi by high-speed cell disruption. J. Clin. Microbiol. 1998, 36, 1625–1629. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications, 1st ed.; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenetics Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Seidl, M.F.; Groenewald, J.Z.; De Vries, M.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 81, 1–21. [Google Scholar] [CrossRef]
- Madrid, H.; Da Cunha, K.C.; Gené, J.; Dijksterhuis, J.; Cano, J.; Sutton, D.A.; Crous, P. Novel Curvularia species from clinical specimens. Persoonia 2014, 33, 48–60. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Gareth, J.E.B.; Camporesi, E.; Hyde, K.D. A new species of the genus Comoclathris (Pleosporaceae). J. Fungal Res. 2015, 13, 260–268. [Google Scholar]
- Ismail, S.I.; Batzer, J.C.; Harrington, T.C.; Crous, P.W.; Lavrov, D.V.; Li, H.; Gleason, M.L. Ancestral state reconstruction infers phytopathogenic origins of sooty blotch and flyspeck fungi on apple. Mycologia 2016, 108, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Restrepo, M.; Madrid, H.; Tan, Y.P.; Da Cunha, K.C.; Gené, J.; Guarro, J.; Crous, P.W. Multi-locus phylogeny and taxonomy of Exserohilum. Persoonia 2018, 41, 71–108. [Google Scholar] [CrossRef] [PubMed]
- Chilvers, M.I.; Rogers, J.D.; Dugan, F.M.; Stewart, J.E.; Chen, W.; Peever, T.L. Didymella pisi sp. nov., the teleomorph of Ascochyta pisi. Mycol. Res. 2009, 113, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Pem, D.; Jeewon, R.; Bhat, D.J.; Doilom, M.; Boonmee, S.; Hongsanan, S.; Promputtha, I.; Xu, J.C.; Hyde, K.D. Mycosphere notes 275-324, A morpho-taxonomic revision and typification of obscure Dothideomycetes genera (incertae sedis). Mycosphere 2019, 10, 1115–1246. [Google Scholar] [CrossRef]
- Kumar, V.; Kumar, N.; Aneja, K.R.; Kaur, M. Gibbago trianthemae, phaeodictyoconidial genus, cause leaf spot disease of Trianthema portulacastrum. Arch. Phytopathol. Plant Prot. 2016, 49, 48–58. [Google Scholar] [CrossRef]
- Crous, P.W.; Shivas, R.G.; Quaedvlieg, W.; van der Bank, M.; Zhang, Y.; Summerell, B.A.; Guarro, J.; Wingfield, M.J.; Wood, A.R.; Alfenas, A.C.; et al. Fungal Planet description sheets: 214–280. Persoonia 2014, 32, 184–306. [Google Scholar] [CrossRef]
- Amaradasa, B.S.; Madrid, H.; Groenewald, J.Z.; Crous, P.W.; Amundsen, K. Porocercospora seminalis gen. et comb. nov., the causal organism of buffalograss false smut. Mycologia 2014, 106, 77–85. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Iturrieta-González, I.; García, D.; Gené, J.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Quaedvlieg, W.; Schumacher, R.K.; et al. Genera of phytopathogenic fungi: GOPHY 3. Stud. Mycol. 2019, 94, 1–124. [Google Scholar] [CrossRef]
- Deng, J.X.; Paul, N.C.; Park, M.S.; Yu, S.H. Molecular characterization, morphology, and pathogenicity of Alternaria panax from araliaceous plants in Korea. Mycol. Prog. 2013, 12, 383–396. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.; Bulgakov, T.S.; Bhat, D.J.; Camporesi, E.; Bahkali, A.H.; Eungwanichayapant, P.D.; Liu, Z.L.; Hyde, K.D. Microfungi on Tamarix. Fungal Divers. 2017, 82, 239–306. [Google Scholar] [CrossRef]
- Crous, P.W.; Schumacher, R.K.; Akulov, A.; Thangavel, R.; Hernández-Restrepo, M.; Carnegie, A.J.; Cheewangkoon, R.; Wingfield, M.J.; Summerell, B.A.; Quaedvlieg, W.; et al. New and interesting fungi. 2. Fungal Syst. Evol. 2019, 3, 57–134. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Farris, J.S.; Kallersjo, M.; Kluge, A.G.; Bult, C. Testing significance of incongruence. Cladistics 1994, 10, 315–319. [Google Scholar] [CrossRef]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A simple, fast and accurate method to estimate large phylogenies by maximum-likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeifferm, W.; Schwartz, T. The CIPRES science gateway: Enabling high-impact science for phylogenetics researchers with limited resources. In Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the Extreme to the Campus and Beyond, Chicago, IL, USA, 16–20 July 2012; Association for Computing Machinery: New York, NY, USA, 2012; pp. 1–8. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Hespanhol, L.; Vallio, C.S.; Costa, L.M.; Saragiotto, B.T. Understanding and interpreting confidence and credible intervals around effect estimates. Braz. J. Phys. Ther. 2019, 23, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Réblová, M.; Kolařík, M.; Nekvindová, J.; Miller, A.N.; Hernández-Restrepo, M. Phylogeny, global biogeography and pleomorphism of Zanclospora. Microorganisms 2021, 9, 706. [Google Scholar] [CrossRef] [PubMed]
- Réblová, M.; Kolařík, M.; Nekvindová, J.; Réblová, K.; Sklenář, F.; Miller, A.N.; Hernández-Restrepo, M. Phylogenetic reassessment, taxonomy, and biogeography of Codinaea and similar Fungi. J. Fungi 2022, 7, 1097. [Google Scholar] [CrossRef] [PubMed]
- Hennebert, G.L. Wardomyces and Asteromyces. Can. J. Bot. 1962, 40, 1203–1216. [Google Scholar] [CrossRef]
- Simmons, E.G. Gibbago, a new phaeodictyoconidial genus of hyphomycetes. Mycotaxon 1986, 27, 107–111. [Google Scholar]
- Campbell, M.A.; Medd, R.W.; Brown, J.F. Growth and sporulation of Pyrenophora semeniperda in vitro: Effects of culture media, temperature and pH. Mycol. Res. 1996, 100, 311–317. [Google Scholar] [CrossRef]
- Svobodová, Y. Chmelia slovaca gen. nov., a dematiaceous fungus, pathogenic for man and animals. Biológia 1966, 21, 81–88. [Google Scholar]
- Funnell-Harris, D.L.; Prom, L.K.; Pedersen, J.F. Isolation and characterization of the grain mold fungi Cochliobolus and Alternaria spp. from sorghum using semi-selective media and DNA sequence analyses. Can. J. Microbiol. 2013, 59, 87–96. [Google Scholar] [CrossRef]
- Gilgado, F.; Cano, J.; Gené, J.; Guarro, J. Molecular phylogeny of the Pseudallescheria boydii species complex: Proposal of two new species. J. Clin. Microbiol. 2005, 43, 4930–4942. [Google Scholar] [CrossRef]
- Pham, T.; Giraud, S.; Schuliar, G.; Rougeron, A.; Bouchara, J.P. Scedo-Select III: A new semi-selective culture medium for detection of the Scedosporium apiospermum species complex. Med. Mycol. 2015, 53, 512–519. [Google Scholar] [CrossRef]
- Nel, W.J.; Duong, T.A.; Wingfield, B.D.; Wingfield, M.J.; De Beer, Z.W. A new genus and species for the globally important, multihost root pathogen Thielaviopsis Basicola. Plant Pathol. 2018, 67, 871–882. [Google Scholar] [CrossRef]
- Wang, X.W.; Bai, F.Y.; Bensch, K.; Meijer, M.; Sun, B.D.; Han, Y.F.; Crous, P.W.; Samson, R.; Yang, F.Y.; Houbraken, J. Phylogenetic re-evaluation of Thielavia with the introduction of a new family Podosporaceae. Stud. Mycol. 2019, 93, 155–252. [Google Scholar] [CrossRef] [PubMed]
- Bhunjun, C.S.; Dong, Y.; Jayawardena, R.S.; Jeewon, R.; Phukhamsakda, C.; Bundhun, D.; Hyde, K.D.; Sheng, J. A polyphasic approach to delineate species in Bipolaris. Fungal Divers. 2020, 102, 225–256. [Google Scholar] [CrossRef]
- Raza, M.; Zhang, Z.F.; Hyde, K.D.; Diao, Y.Z.; Cai, L. Culturable plant pathogenic fungi associated with sugarcane in southern China. Fungal Divers. 2019, 99, 1–104. [Google Scholar] [CrossRef]
- Valenzuela-López, N.; Martin-Gómez, M.T.; Los-Arcos, I.; Stchigel, A.M.; Guarro, J.; Cano-Lira, J.F. A new pleosporalean fungus isolated from superficial to deep human clinical specimens. Med. Mycol. 2021, 59, 278–288. [Google Scholar] [CrossRef]
- Madrid, H.; Hernández-Restrepo, M.; Gené, J.; Cano, J.; Guarro, J.; Silva, V. New and interesting chaetothyrialean fungi from Spain. Mycol. Prog. 2016, 15, 1179–1201. [Google Scholar] [CrossRef]
- Chen, J.J.; Feng, X.X.; Xia, C.Y.; Kong, D.D.; Qi, Z.Y.; Liu, F.; Chen, D.; Lin, F.C.; Zhang, C.L. Confirming the phylogenetic position of the genus Muscodor and the description of a new Muscodor species. Mycosphere 2019, 10, 187–201. [Google Scholar] [CrossRef]
- Saxena, S.; Strobel, G.A. Marvellous Muscodor spp.: Update on their biology and applications. Microb. Ecol. 2021, 82, 5–20. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Hibbett, D.S.; Kirk, P.M.; Lücking, R. Proposals to permit DNA sequence data to serve as types of names of fungi. Taxon 2016, 65, 899–900. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Puusepp, R.; Nilsson, R.H.; James, T.Y. Novel soil-inhabiting clades fill gaps in the fungal tree of life. Microbiome 2017, 5, 42. [Google Scholar] [CrossRef]
- Ryberg, M.; Nilsson, R.H. New light on names and naming of dark taxa. MycoKeys 2018, 30, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Aoki, T.; Ariyawansa, H.A.; Cardinali, G.; Crous, P.W.; Druzshinina, I.S.; Geiser, D.M.; et al. Fungal taxonomy and sequence-based nomenclature. Nat. Microbiol. 2021, 6, 540–548. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain Number | Substrate | Country | GenBank Accession Number 1 | Citation | ||||
|---|---|---|---|---|---|---|---|---|---|
| ITS | LSU | rpb2 | tef1 | gapdh | |||||
| Alternaria abundans | CBS 534.83 T | Fragaria stolon | New Zealand | MH861639 | MH873354 | KC584448 | KC584707 | KC584154 | [18] |
| Alternaria alternata | CBS 104.26 | Unknown | Uknown | KP124299 | KP124450 | KP124767 | KP125074 | KP124156 | [32] |
| Alternaria anigozanthi | CBS 121920 T | Anigozanthus cultivar | Australia | KC584180 | NG069123 | KC584376 | KC584635 | KC584097 | [18] |
| Alternaria argyranthemi | CBS 116530 | Argyranthemum sp. | New Zealand | KC584181 | KC584254 | KC584378 | KC584637 | KC584098 | [18] |
| Alternaria avenicola | CBS 121459 T | Avena sp. | Norway | KC584183 | KC584256 | KC584380 | KC584639 | KC584100 | [18] |
| Alternaria infectoria | CBS 210.86 T | Triticum aestivum | USA | FM958526 | MH873633 | KC584404 | KC584662 | KY038017 | [18] |
| Alternaria macrospora | CBS 117228 T | Gossypium barbadense | USA | NR136045 | NG069135 | KC584410 | KC584668 | KC584124 | [18] |
| Alternaria nobilis | CBS 116490 T | Dianthus caryophyllus | New Zealand | KC584208 | KC584291 | KC584415 | KC584673 | KC584127 | [18] |
| Alternaria obclavata | CBS 124120 T | Air | USA | NR165505 | MH874877 | KC584443 | KC584701 | KC584149 | [18] |
| Alternaria penicillata | CBS 116607 T | Papaver rhoeas | Austria | KC584229 | KC584322 | KC584447 | KC584706 | - | [18] |
| Alternaria slovaca | CBS 567.66 T | Human clinical specimen | Slovakia | KC584226 | KC584319 | KC584444 | KC584702 | KC584150 | [18] |
| Alternaria solani | CBS 116651 | Solanum tuberosum | USA | KC584217 | KC584306 | KC584430 | KC584688 | KC584139 | [18] |
| Asteromyces cruciatus | CBS 171.63 T | Sand of dunes | France | MH858254 | MH869856 | ON703247 | ON542234 | ON542232 | [19]; this study |
| CBS 536.92 | Composting seaweed | USA | ON773141 | ON773155 | ON703248 | ON542235 | ON542233 | This study | |
| Bipolaris cynodontis | CBS 285.51 | Cynodon transvaalensis | Kenya | MH856862 | MH868380 | HF934831 | - | HG779081 | [33] |
| Bipolaris maydis | CBS 136.29 ET | Zea mays | Japan | MH855024 | MH866491 | HF934828 | - | HG779086 | [33] |
| Bipolaris microlaenae | CBS 280.91 T | Microlaena stipoides | Australia | NR137073 | HF934877 | HF934835 | - | HG779092 | [33] |
| Clathrospora elynae | CBS 196.54 | Carex curvula | Switzerland | MH857290 | MH872973 | KC584496 | - | - | [18] |
| Comoclathris incompta | CBS 467.76 | Olaea europaea branch | Grece | KY940770 | MH871007 | KC584504 | - | - | [18] |
| Comoclathris linis | MFLUCC 145047 T | Dead stems of Linum sp. | Italy | NR153904 | NG058917 | - | - | - | [34] |
| Comoclathris sedis | CBS 366.52 | Unknown | USA | KY940748 | MH871007 | KT216533 | - | - | [35] |
| Curvularia heteropogonis | CBS 284.91 T | Heteropogon contorus | Australia | MH862253 | LT631396 | HF934821 | - | HG779121 | [33] |
| Curvularia lunata | CBS 730.96 NT | Human lung biopsy | USA | MG722981 | LT631416 | HF934813 | - | LT715821 | [36] |
| Decorospora gaudefroyi | CBS 332.63 | Unknown | France | MH858305 | MH869915 | - | - | - | [19] |
| CBS 250.60 | Unknown | UK | MH857974 | MH869526 | - | - | - | [19] | |
| Dichotomophthora lutea | CBS 145.57 T | Unknown | Unknown | MH857676 | NG069497 | LT990634 | - | LT990663 | [19] |
| Dichotomophthora portulacae | CBS 174.35 T | Unknown | Unknown | NR158421 | MH867137 | LT990638 | LT990668 | - | [19] |
| Didymella exigua | CBS 183.55 | Rumex arifolius | France | MH857436 | MH871007 | EU874850 | - | - | [37] |
| Exserohilum monoceras | CBS 239.77 | Echinochloa colona | Australia | LT837474 | LT883405 | LT852506 | - | LT883547 | [36] |
| Exserohilum rostratum | CBS 128061 | Zea mays | USA | KT265240 | MH877986 | LT715752 | - | LT715900 | [36] |
| Exserohilum turcicum | CBS 387.58 | Zea mays | USA | MH857820 | LT883412 | LT852514 | - | LT883554 | [36] |
| Gibbago trianthemae | NFCCI 1886 | Trianthema portulacastrum | India | HM448998 | MH870931 | - | - | - | [19,38] |
| GT-VM | Trianthema portulacastrum | Pakistan | KJ825852 | MH870931 | - | - | - | [39] | |
| Johnalcornia aberrans | CBS 510.91 | Unknown | Australia | MH862272 | KM243286 | LT715737 | - | KM257056 | [36] |
| Neocamarosporium chichastianum | CBS 137502 | Unknown | Iran | KJ869163 | MH877648 | - | - | - | [40] |
| Neocamarosporium goegapense | CPC 23676 | Mesembryanthemum sp. | South Africa | KJ869163 | KJ869220 | - | - | - | [40] |
| Paradendriphyella arinariae | CBS 181.58 T | Unknown | France | MH857747 | KC793338 | DQ435065 | - | - | [19] |
| Paradendriphyella salina | CBS 302.84 T | Cancer pagurus shell | Denmark | MH873443 | KC584325 | KC584450 | KC584709 | - | [19] |
| CBS 142.60 | Stem of Spartina sp. | England | MH857928 | MH869472 | DQ435066 | - | - | [19] | |
| Pheosphaeria oryzae | CBS 110110 | Oryza sativa | Korea | MH862850 | MH871007 | - | - | - | [19] |
| Porocercospora seminalis | CBS 134907 | Bouteloua dactyloides | USA | HF934941 | HF934862 | HF934843 | - | - | [41] |
| CPC 21349 | B. dactyloides | USA | HF934945 | HF934861 | HF934845 | - | - | [41] | |
| Neostemphylium polymorphum | FMR 17886 T | Fluvial sediment | Spain | OU195609 | OU195892 | OU196009 | ON368192 | OU195960 | This study |
| FMR 17889 | Fluvial sediment | Spain | OU195610 | OU195914 | OU196957 | ON368193 | OU195977 | This study | |
| FMR 17893 | Fluvial sediment | Spain | OU195631 | OU195915 | OU197255 | ON368194 | OU195978 | This study | |
| FMR 17894 | Fluvial sediment | Spain | OU195879 | OU195937 | OU196956 | ON368195 | OU195998 | This study | |
| FMR 17895 | Fluvial sediment | Spain | OU195878 | OU195936 | OU197545 | ON368196 | OU195999 | This study | |
| Pyrenophora avenicola | CBS 307.84 | Avena seed | Sweden | MK539972 | MK540042 | - | - | MK540180 | [42] |
| Pyrenophora chaetomioides | CBS 314.68 | Avena sativa | Germany | MK539979 | MH870853 | MK540105 | - | MK540187 | [42] |
| Pyrenophora cynosuri | CBS 127918 | Seeds of Cynosurus | New Zealand | MK539980 | MK540047 | MK540106 | - | MK540188 | [42] |
| Pyrenophora novozelandica | CBS 127934 | Seeds of Triticum sp. | New Zealand | MK539997 | MK540061 | MK540125 | - | MK540209 | [42] |
| Pyrenophora phaecomes | DAOM 222769 | Unknown | Unknown | JN943649 | JN940093 | DQ497614 | - | - | [18] |
| Pyrenophora pseudoerythrospila | CBS 127931 T | Lolium sp. | Germany | NR164465 | NG066344 | - | - | MK540212 | [42] |
| Pyrenophora seminiperda | CBS 127927 | Unknown | Unknown | MK540001 | MH877966 | MK540128 | - | MK540213 | [42] |
| Scleromyces submersus | FMR 18289 T | Fluvial sediment | Spain | OU195893 | OU195959 | OU197244 | OU196982 | OU196008 | This study |
| Stemphylium botryosum | CBS 714.68 T | Medicago sativa | Canada | MH859208 | MH870931 | - | KC584729 | MH206176 | [18] |
| Stemphylium lycopersici | CNU 070067 | Capsicum annum | Korea | JF417683 | - | JF417698 | JX213347 | JF417693 | [43] |
| Stemphylium vesicarium | CBS 191.86 | Medicago sativa | India | MH861935 | JX681120 | KC584471 | KC584731 | - | [18] |
| Tamaricicola muriformis | MFLUCC 150488 | Tamarix sp. | Italy | KU752187 | KU561879 | KU820870 | - | - | [44] |
| MFLUCC 150489 | Tamarix sp. | Italy | KU752188 | KU729857 | - | - | - | [44] | |
| Typhicola typharum | CBS 145043 NT | Leaf of Typha sp. | Germany | MK442590 | MK442530 | MK442666 | MK442696 | - | [45] |
| Abundance | Other Data 4 | Geographical Origin 5 | Substrate 6 | Biomes 7 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Taxa | Samples 1 | Reads 2 | FR 3 | MAT | MAP | pH | USA | Europe | Australia | Soil | Rhiz. Soil * | Root | Others 8 | Wetland | Cropland | Forest | Woodland | Shrubland | Grassland | Aquatic |
| Neostemphylium polymorphum ITS1 | 3 | 45.56 | 0.079 | 13.0 | 700.2 | 5.8 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 |
| Neostemphylium ITS1-ENV1 | 2 | 2.50 | 0.004 | 15.2 | 935.6 | 5.7 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 |
| Neostemphylium polymorphum ITS2 | 2 | 19.88 | 0.064 | 9.7 | 854.4 | 6.0 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 |
| Scleromyces submersus ITS2 | 1 | 29.00 | 0.140 | 16.4 | 767.0 | 5.8 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Scleromyces ITS1-ENV2 | 1 | 2.00 | 0.012 | 15.0 | 681.0 | 5.8 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Scleromyces ITS1-ENV3 | 1 | 2.00 | 0.012 | 20.1 | 563.0 | 6.7 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Scleromyces ITS2-ENV1 | 2 | 26.87 | 0.044 | 16.8 | 731.4 | NA 9 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Garcia, D.; García, D.; Cano-Lira, J.F.; Gené, J. Two Novel Genera, Neostemphylium and Scleromyces (Pleosporaceae) from Freshwater Sediments and Their Global Biogeography. J. Fungi 2022, 8, 868. https://doi.org/10.3390/jof8080868
Torres-Garcia D, García D, Cano-Lira JF, Gené J. Two Novel Genera, Neostemphylium and Scleromyces (Pleosporaceae) from Freshwater Sediments and Their Global Biogeography. Journal of Fungi. 2022; 8(8):868. https://doi.org/10.3390/jof8080868
Chicago/Turabian StyleTorres-Garcia, Daniel, Dania García, José F. Cano-Lira, and Josepa Gené. 2022. "Two Novel Genera, Neostemphylium and Scleromyces (Pleosporaceae) from Freshwater Sediments and Their Global Biogeography" Journal of Fungi 8, no. 8: 868. https://doi.org/10.3390/jof8080868