
Polyphasic Systematics of the Fungicolous Genus Cladobotryum Based on Morphological, Molecular and Metabolomics Data

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Material and Culture Conditions
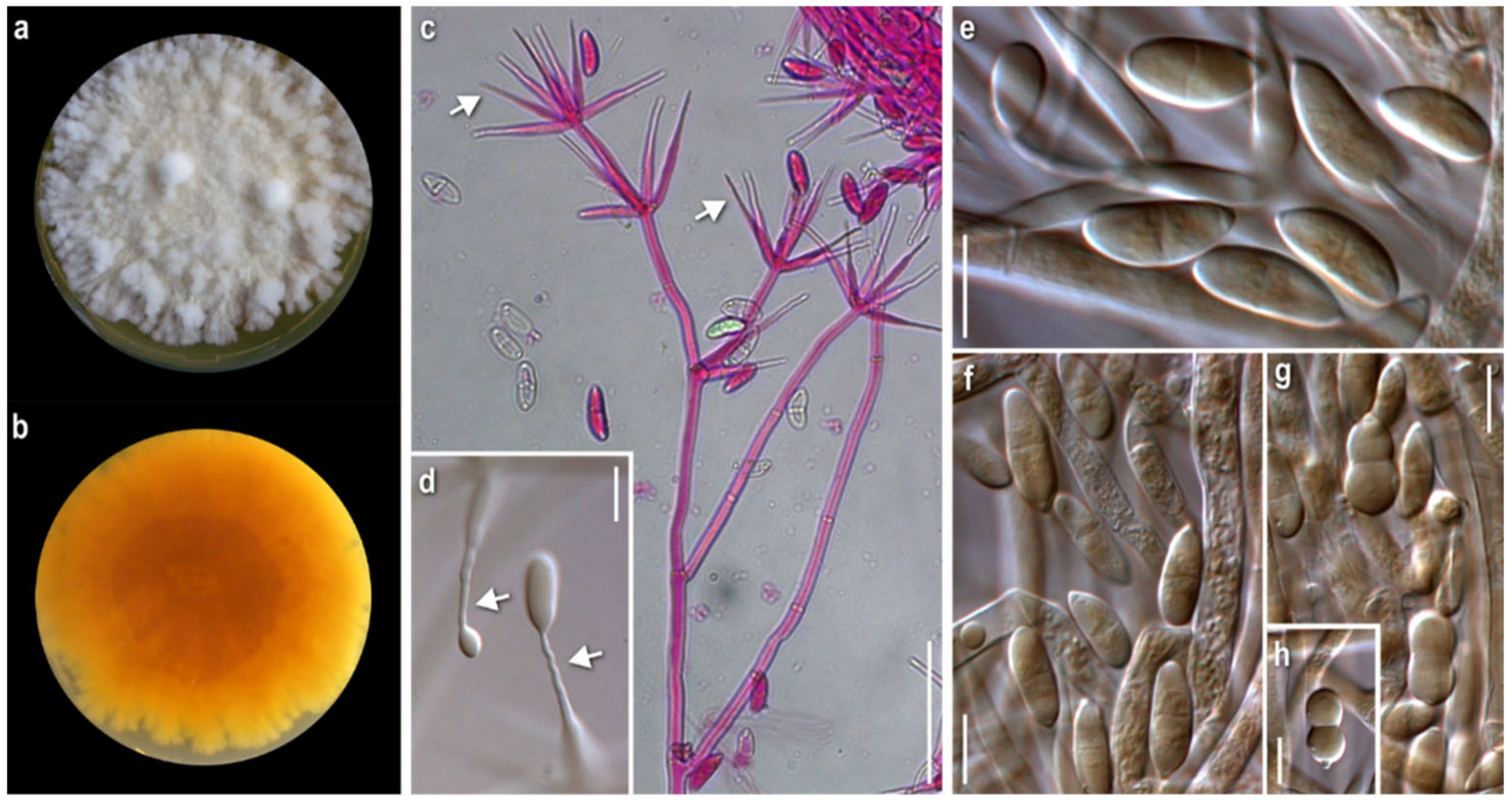
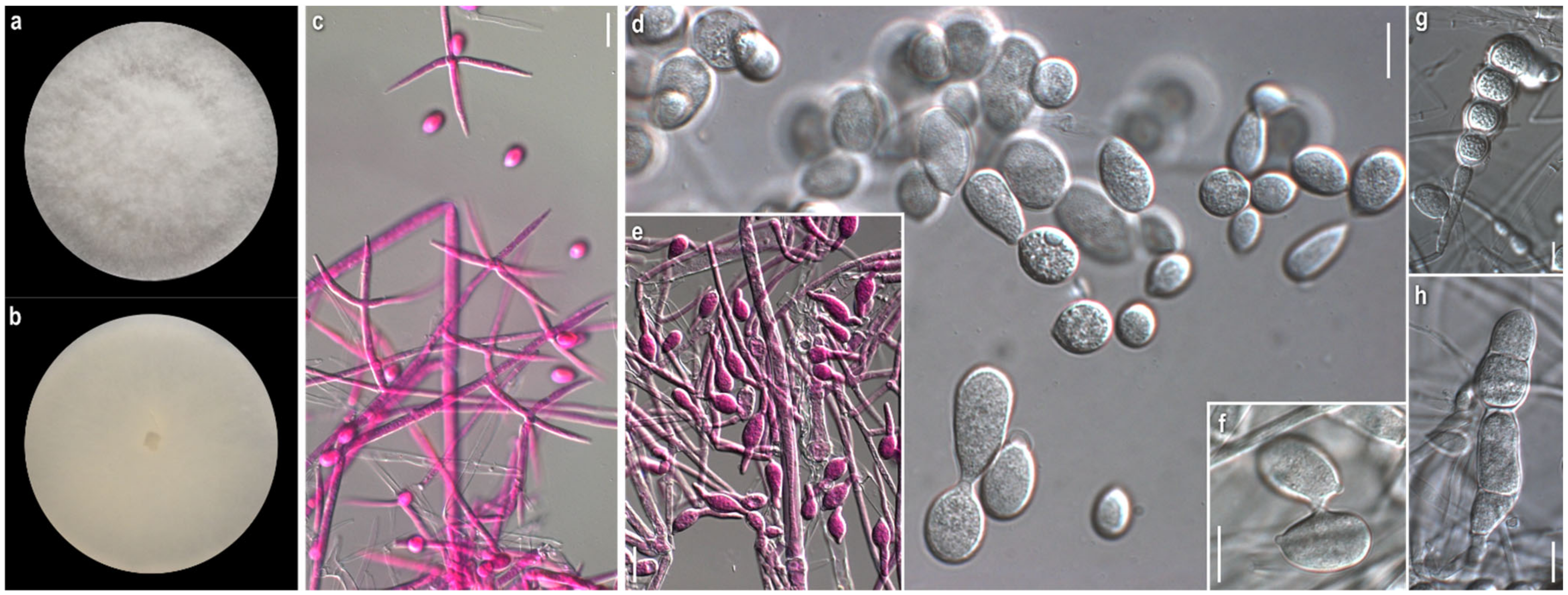
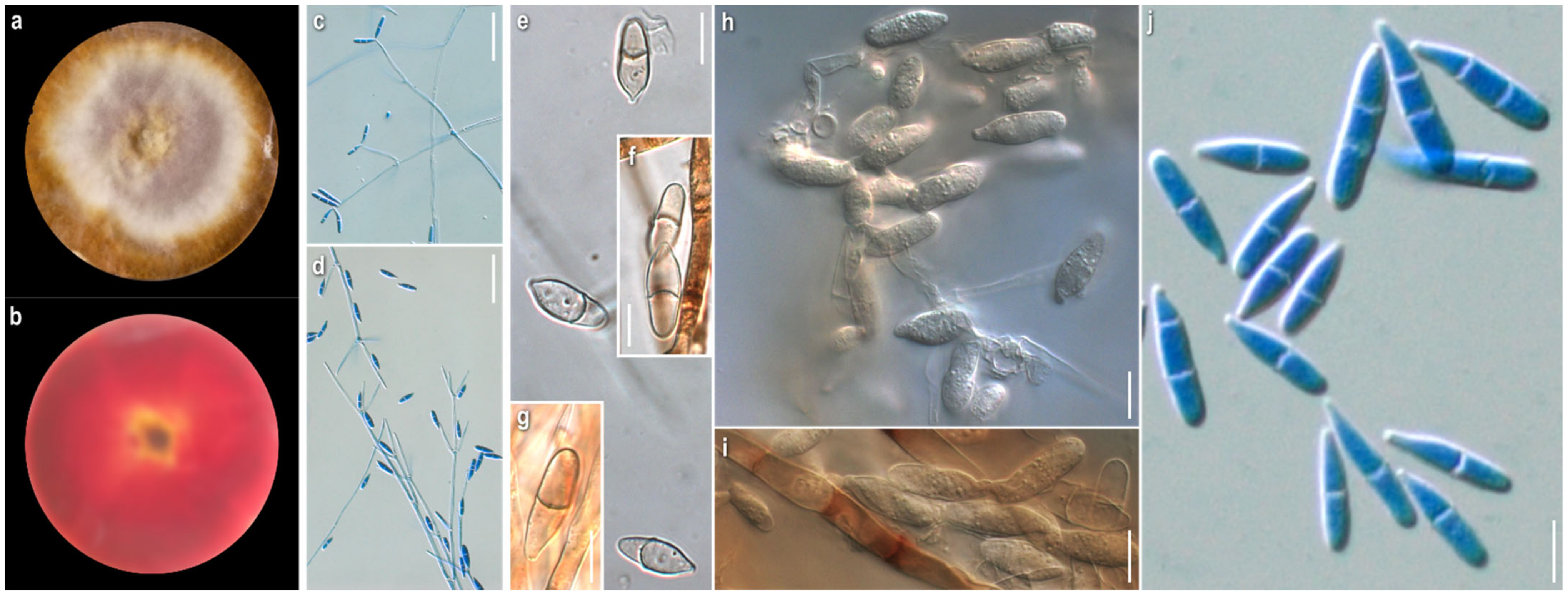
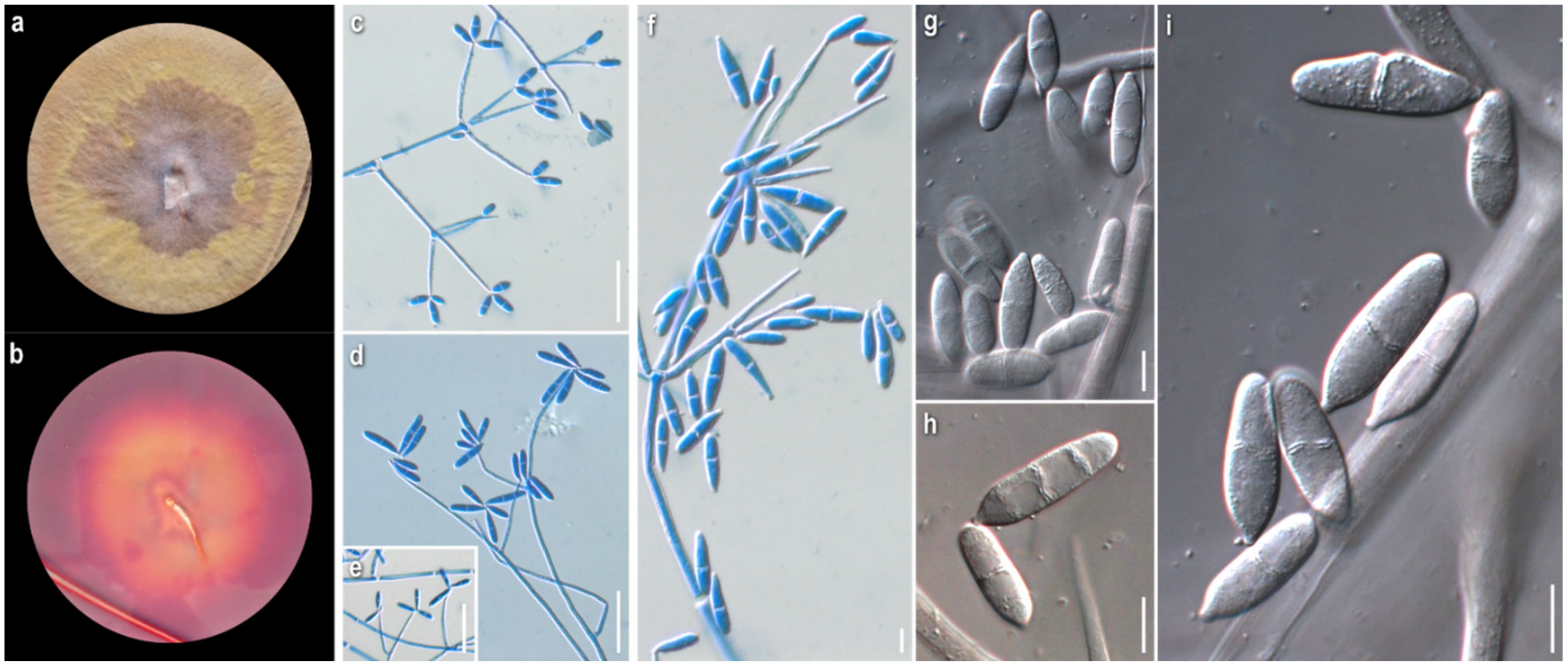
2.2. Morphological Study
2.3. Molecular Phylogenetic Analyses
2.3.1. DNA Extraction and PCR Reaction
2.3.2. Phylogenetic Tree Reconstruction
2.4. NMR-Based Metabolomics
2.4.1. Metabolite Extraction
2.4.2. NMR Spectroscopy
2.4.3. Statistical Analysis
3. Results
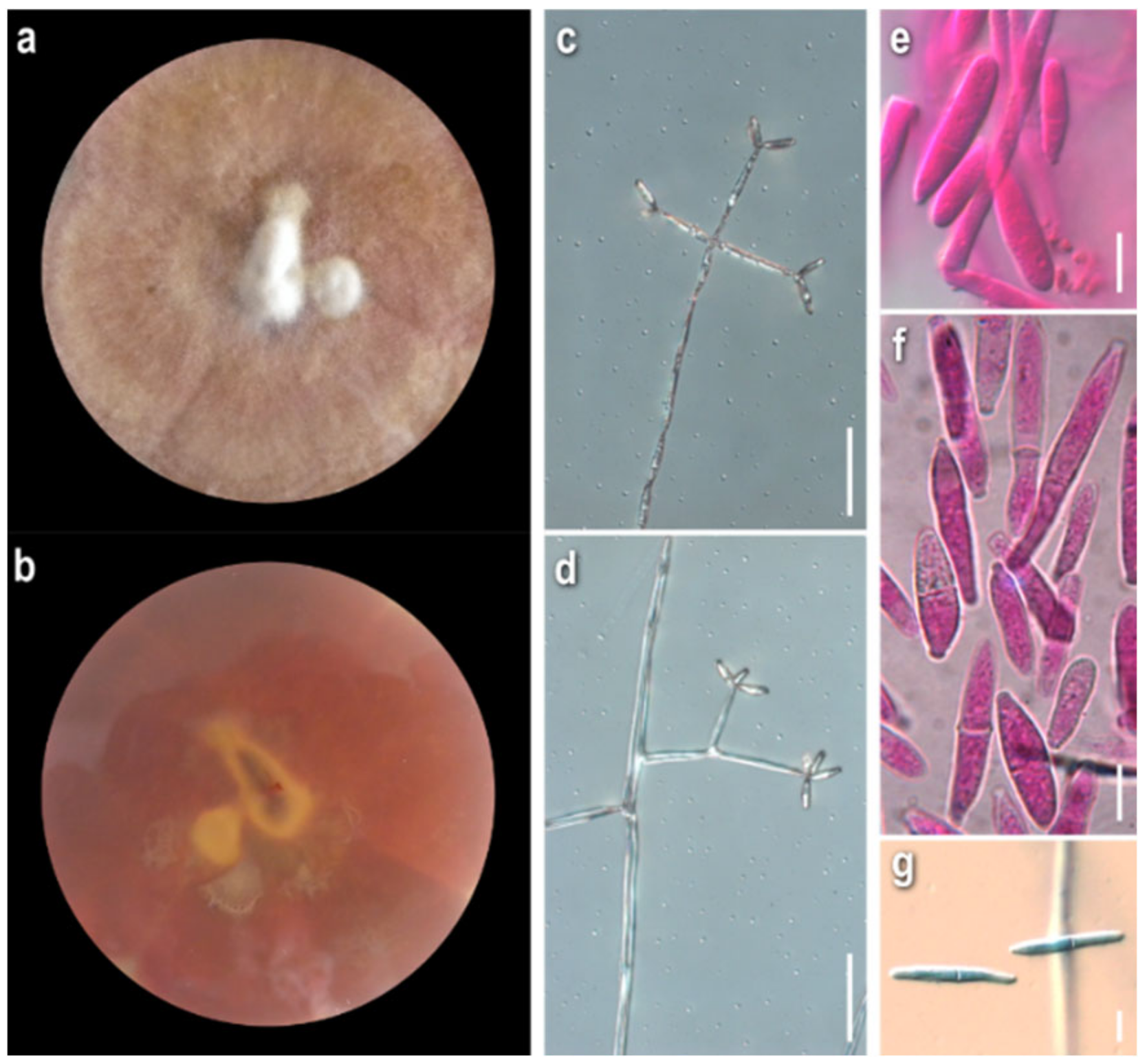
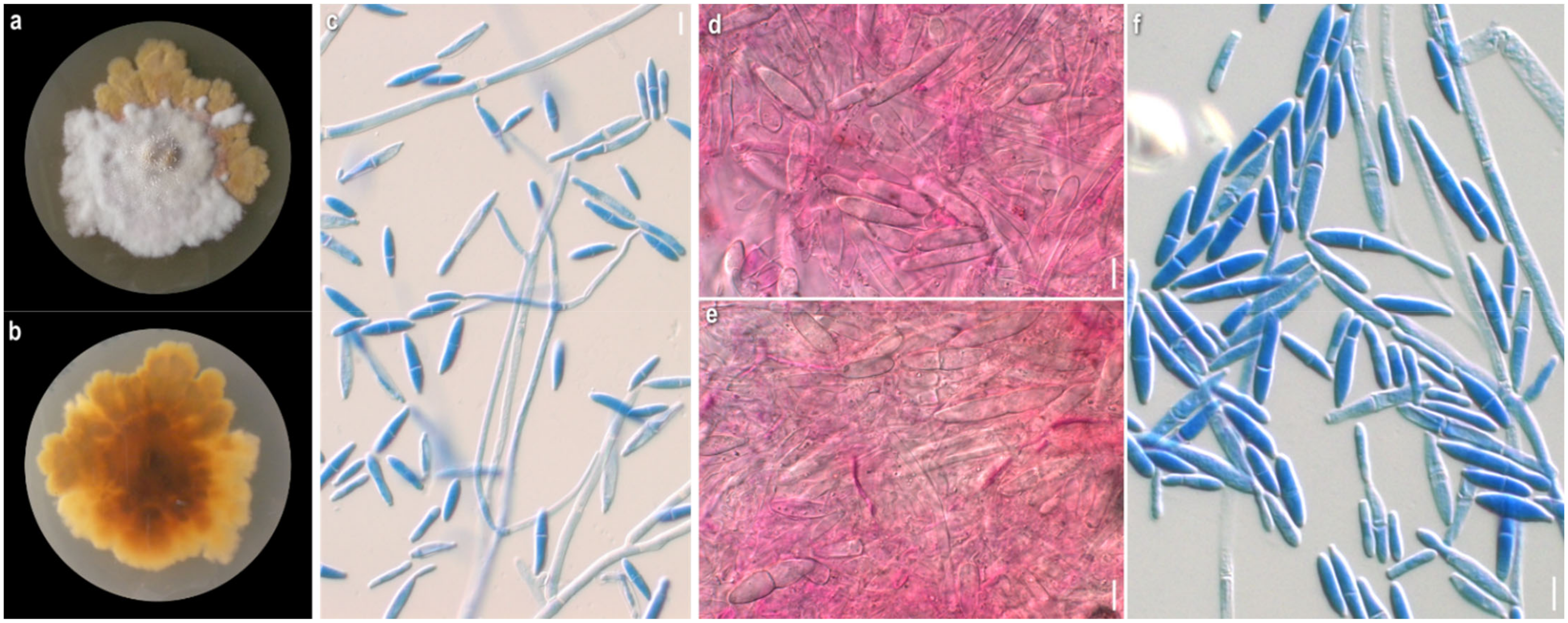
3.1. Morphological Study
3.2. Molecular Analyses
3.3. NMR-Based Metabolomics/Chemotaxonomy
4. Discussion
4.1. Contribution of Data Derived from Morphology and Notes on Ecology
4.2. Contribution of Data Derived from Molecular Phylogenetic Analyses
4.3. Contribution of Data Derived from NMR-Based Metabolomics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Samuels, G.J.; Seifert, K.A. Taxonomic Implications of Variation among Hypocrealean Anamorphs. In Pleomorphic Fungi: The Diversity and Its Taxonomic Implications; Sugiyama, J., Ed.; Elsevier: Tokyo, Japan, 1987; pp. 29–56. ISBN 978-0-444-98966-6. [Google Scholar] [CrossRef]
- Põldmaa, K. Generic Delimitation of the Fungicolous Hypocreaceae. Stud. Mycol. 2000, 45, 83–94. [Google Scholar]
- Rossman, A.Y.; Seifert, K.A.; Samuels, G.J.; Minnis, A.M.; Schroers, H.-J.; Lombard, L.; Crous, W.; Põldmaa, K.; Cannon, P.F.; Summerbell, R.C.; et al. Genera in Bionectriaceae, Hypocreaceae, and Nectriaceae (Hypocreales) Proposed for Acceptance or Rejection. IMA Fungus 2013, 4, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Samuels, G.J. Molecular Systematics of the Hypocreales: A Teleomorph Gene Phylogeny and the Status of Their Anamorphs. Can. J. Bot. 1995, 73, 816–823. [Google Scholar] [CrossRef]
- Sutton, B.C.; Hennebert, G.L. Interconnections amongst Anamorphs and Their Possible Contribution to Ascomycete Systematics. In Ascomycete Systematics; Hawksworth, D.L., Ed.; Springer: Boston, MA, USA, 1994; pp. 77–100. ISBN 978-1-4757-9292-8. [Google Scholar] [CrossRef]
- Rogerson, C.T.; Samuels, G.J. Polyporicolous Species of Hypomyces. Mycologia 1993, 85, 213–272. [Google Scholar] [CrossRef]
- Helfer, W. Pilze Auf Pilzfruchtkörpern: Untersuchungen Zur Ökologie, Systematik Und Chemie; Libri botanici, Band 1; IHW-Verlag: Eching, Germany, 1991. [Google Scholar]
- Gams, W.; Hoozemans, A.C.M. Cladobotryum-Konidienformen von Hypomyces-Arten. Pers.-Mol. Phylogeny Evol. Fungi 1970, 6, 95–110. [Google Scholar]
- Põldmaa, K. The Genus Hypomyces and Allied Fungicolous Fungi in Estonia. I. Species Growing on Aphyllophoralean Basidiomycetes. Folia Cryptogam. Est. 1999, 34, 15–31. [Google Scholar]
- Arnold, G. Über Eine Neue Hypomycetazee, Hypomyces odoratus Arnold Sp. Nov. Čes. Mykol. 1964, 18, 144–146. [Google Scholar]
- Arnold, G.R.W. Über Sibirina Und Sympodiophora Zwei Neue Gattungen Der Moniliales. Nova Hedwig. 1970, 19, 299–304. [Google Scholar]
- Arnold, G.R.W. De Nonnullis Fungis Raris Vel Novis Vel pro URSS Primum Indicatis. Nov. Sist. Nizshikh Rastenii 1971, 8, 130–138. [Google Scholar]
- McNeill, J.; Turland, N.J. Major Changes to the Code of Nomenclature—Melbourne, July 2011. TAXON 2011, 60, 1495–1497. [Google Scholar] [CrossRef]
- Turland, N. The Code Decoded: A User’s Guide to the International Code of Nomenclature for Algae, Fungi, and Plants, 2nd ed.; Pensoft Publishers: Sofia, Bulgaria, 2019; ISBN 978-3-946583-16-5. [Google Scholar] [CrossRef]
- Kirk, P.M.; Stalpers, J.A.; Braun, U.; Crous, P.W.; Hansen, K.; Hawksworth, D.L.; Hyde, K.D.; Lücking, R.; Lumbsch, T.H.; Rossman, A.Y.; et al. A without-prejudice list of generic names of fungi for protection under the International Code of Nomenclature for algae, fungi, and plants. IMA Fungus 2013, 4, 381–443. [Google Scholar] [CrossRef] [PubMed]
- Rudakov, O.L. Physiological Groups in Mycophilic Fungi. Mycologia 1978, 70, 150–159. [Google Scholar] [CrossRef]
- Rossman, A.Y. Morphological and Molecular Perspectives on Systematics of the Hypocreales. Mycologia 1996, 88, 1–19. [Google Scholar] [CrossRef]
- Rossman, A.Y.; Samuels, G.I.; Rogerson, C.T.; Lowen, R. Genera of Bionectriaceae, Hypocreaceae and Nectriaceae (Hypocreales, Ascomycetes). Stud. Mycol. 1999, 42, 1–248. [Google Scholar]
- Hawksworth, D.L. A Survey of the Fungicolous Conidial Fungi. In Biology of Conidial Fungi; Cole, G.T., Kendrick, B., Eds.; Elsevier: New York, NY, USA, 1981; Volume 1, pp. 171–244. [Google Scholar]
- Gams, W.; Diederich, P.; Põldmaa, K. Fungicolous Fungi. In Biodiversity of Fungi; Mueller, G.M., Bills, G.F., Foster, M.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 343–392. ISBN 978-0-12-509551-8. [Google Scholar] [CrossRef]
- Sun, J.-Z.; Liu, X.-Z.; McKenzie, E.H.C.; Jeewon, R.; Liu, J.-K.; Zhang, X.-L.; Zhao, Q.; Hyde, K.D. Fungicolous Fungi: Terminology, Diversity, Distribution, Evolution, and Species Checklist. Fungal Divers. 2019, 95, 337–430. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases/fungushost/fungushost.cfm (accessed on 2 March 2022).
- Põldmaa, K. Three Species of Hypomyces Growing on Basidiomata of Stereaceae. Mycologia 2003, 95, 921–933. [Google Scholar] [CrossRef]
- Carrasco, J.; Navarro, M.-J.; Gea, F.J. Cobweb, a Serious Pathology in Mushroom Crops: A Review. Span. J. Agric. Res. 2017, 15, e10R01. [Google Scholar] [CrossRef]
- Fletcher, J.T.; Gaze, R.H. Fungal Diseases. In Mushroom Pest and Disease Control: A Colour Handbook; Fletcher, J.T., Gaze, R.H., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 63–91. ISBN 978-1-84076-505-2. [Google Scholar] [CrossRef]
- Gea, F.J.; Navarro, M.J.; Santos, M.; Diánez, F.; Carrasco, J. Control of Fungal Diseases in Mushroom Crops While Dealing with Fungicide Resistance: A Review. Microorganisms 2021, 9, 585. [Google Scholar] [CrossRef]
- Baars, J.; Hendrickx, P.; Sonnenberg, A.; Korsten, L. Global Collection of Mushroom Pathogens; Plant Research International, Part of Wageningen UR Business Unit Plant Breeding: Wageningen, The Netherlands, 2013; p. 41. [Google Scholar]
- Santos, M.; Diánez, F.; Moreno-Gavíra, A.; Sánchez-Montesinos, B.; Gea, F.J. Cladobotryum mycophilum as Potential Biocontrol Agent. Agronomy 2019, 9, 891. [Google Scholar] [CrossRef]
- De Hoog, G.S. Notes Some Fungicolous Hyphomycetes and Their Relatives. Pers.-Mol. Phylogeny Evol. Fungi 1978, 10, 33–81. [Google Scholar]
- Rogerson, C.T.; Samuels, G.J. Agaricicolous Species of Hypomyces. Mycologia 1994, 86, 839–866. [Google Scholar] [CrossRef]
- Põldmaa, K.; Samuels, G.J. Aphyllophoricolous Species of Hypomyces with KOH-Negative Perithecia. Mycologia 1999, 91, 177–199. [Google Scholar] [CrossRef]
- Rogerson, C.T.; Samuels, G.J. Boleticolous Species of Hypomyces. Mycologia 1989, 81, 413–432. [Google Scholar] [CrossRef]
- Valadon, L.R.G.; Chapman, D.J. The Naphthoquinones (Fungal Vacuolation Factor) of Verticillium agaricinum. Microbios Lett. 1983, 24, 115–119. [Google Scholar]
- Nair, M.S.R.; Carey, S.T. Metabolites of Pyrenomycetes: XII. Polyketides from The Hypocreales. Mycologia 1979, 71, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Tamm, H.; Põldmaa, K. Diversity, Host Associations, and Phylogeography of Temperate Aurofusarin-Producing Hypomyces/Cladobotryum Including Causal Agents of Cobweb Disease of Cultivated Mushrooms. Fungal Biol. 2013, 117, 348–367. [Google Scholar] [CrossRef]
- Põldmaa, K. Tropical Species of Cladobotryum and Hypomyces Producing Red Pigments. Stud. Mycol. 2011, 68, 1–34. [Google Scholar] [CrossRef]
- Bastos, C.N.; Neill, S.J.; Horgan, R. A Metabolite from Cladobotryum amazonense with Antibiotic Activity. Trans. Br. Mycol. Soc. 1986, 86, 571–578. [Google Scholar] [CrossRef]
- Wagner, C.; Anke, H.; Sterner, O. Rubrobramide, a Cytotoxic and Phytotoxic Metabolite from Cladobotryum rubrobrunnescens. J. Nat. Prod. 1998, 61, 501–502. [Google Scholar] [CrossRef]
- Feng, Y.; Blunt, J.W.; Cole, A.L.J.; Cannon, J.F.; Robinson, W.T.; Munro, M.H.G. Two Novel Cytotoxic Cyclodepsipeptides from a Mycoparasitic Cladobotryum sp. J. Org. Chem. 2003, 68, 2002–2005. [Google Scholar] [CrossRef]
- Mitova, M.I.; Lang, G.; Blunt, J.W.; Cummings, N.J.; Cole, A.L.J.; Robinson, W.T.; Munro, M.H.G. Cladobotric Acids A−F: New Cytotoxic Polyketides from a New Zealand Cladobotryum sp. J. Org. Chem. 2006, 71, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Tsujii, E.; Abe, F.; Yamashitat, M.; Shigematsu, N.; IzuMift, S.; Okuhara, M. FR901483, a Novel Immunosuppressant Isolated from Cladobotryum sp. No. 11231. Taxonomy of the Producing Organism, fermentation, Isolation, Physico-chemical Properties and Biological Activities. J. Antibiot. 1996, 49, 8. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.-T.; Williams, K.; de Mattos-Shipley, K.M.J.; Song, Z.; Takebayashi, Y.; Simpson, T.J.; Spencer, J.; Bailey, A.M.; Willis, C.L. Cladobotric Acids: Metabolites from Cultures of Cladobotryum sp., Semisynthetic Analogues and Antibacterial Activity. J. Nat. Prod. 2022, 85, 572–580. [Google Scholar] [CrossRef]
- Watanabe, Y.; Yoshida, Y.; Tokiwa, T.; Higo, M.; Ban, S.; Ikeda, A.; Noguchi, Y.; Hirose, T.; Sunazuka, T.; Nonaka, K.; et al. Hakuhybotric Acid, a New Antifungal Polyketide Produced by a Mycoparasitic Fungus Hypomyces pseudocorticiicola FKI-9008. J. Gen. Appl. Microbiol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Breinholt, J.; Jensen, G.W.; Kjær, A.; Olsen, C.E.; Rosendahl, C.N.; Romerosa, A.; Robinson, W.T.; Roos, B.O.; Vallance, C.; Wood, B.R. Hypomycetin—an Antifungal, Tetracyclic Metabolite from Hypomyces aurantius: Production, Structure and Biosynthesis. Acta Chem. Scand. 1997, 51, 855–860. [Google Scholar] [CrossRef]
- McKay, G.J.; Egan, D.; Morris, E.; Scott, C.; Brown, A.E. Genetic and Morphological Characterization of Cladobotryum Species Causing Cobweb Disease of Mushrooms. Appl. Environ. Microbiol. 1999, 65, 606–610. [Google Scholar] [CrossRef]
- Põldmaa, K.; Larsson, E.; Kõljalg, U. Phylogenetic Relationships in Hypomyces and Allied Genera, with Emphasis on Species Growing on Wood-Decaying Homobasidiomycetes. Can. J. Bot. 1999, 77, 13. [Google Scholar] [CrossRef]
- Frisvad, J.C. Fungal Chemotaxonomy. In Biosynthesis and Molecular Genetics of Fungal Secondary Metabolites, Volume 2; Zeilinger, S., Martín, J.-F., García-Estrada, C., Eds.; Fungal Biology; Springer: New York, NY, USA, 2015; pp. 103–121. ISBN 978-1-4939-2530-8. [Google Scholar]
- Frisvad, J.C. Chemical Fungal Taxonomy, 1st ed.; Frisvad, J.C., Bridge, P.D., Arora, D.K., Eds.; CRC Press: Boca Raton, FL, USA, 1998; ISBN 978-1-00-306462-6. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Andersen, B.; Thrane, U. The Use of Secondary Metabolite Profiling in Chemotaxonomy of Filamentous Fungi. Mycol. Res. 2008, 112, 231–240. [Google Scholar] [CrossRef]
- Ząbek, A.; Klimek-Ochab, M.; Jawień, E.; Młynarz, P. Biodiversity in Targeted Metabolomics Analysis of Filamentous Fungal Pathogens by 1H NMR-Based Studies. World J. Microbiol. Biotechnol. 2017, 33, 132. [Google Scholar] [CrossRef]
- Kim, W.; Peever, T.L.; Park, J.-J.; Park, C.-M.; Gang, D.R.; Xian, M.; Davidson, J.A.; Infantino, A.; Kaiser, W.J.; Chen, W. Use of Metabolomics for the Chemotaxonomy of Legume-Associated Ascochyta and Allied Genera. Sci. Rep. 2016, 6, 20192. [Google Scholar] [CrossRef]
- Becker, K.; Wongkanoun, S.; Wessel, A.-C.; Bills, G.F.; Stadler, M.; Luangsa-ard, J.J. Phylogenetic and Chemotaxonomic Studies Confirm the Affinities of Stromatoneurospora phoenix to the Coprophilous Xylariaceae. J. Fungi 2020, 6, 144. [Google Scholar] [CrossRef] [PubMed]
- Maciá-Vicente, J.G.; Shi, Y.-N.; Cheikh-Ali, Z.; Grün, P.; Glynou, K.; Kia, S.H.; Piepenbring, M.; Bode, H.B. Metabolomics-Based Chemotaxonomy of Root Endophytic Fungi for Natural Products Discovery: Chemotaxonomy of Root Endophytic Fungi. Environ. Microbiol. 2018, 20, 1253–1270. [Google Scholar] [CrossRef] [PubMed]
- Kelman, M.J.; Renaud, J.B.; Seifert, K.A.; Mack, J.; Yeung, K.K.-C.; Sumarah, M.W. Chemotaxonomic Profiling of Canadian Alternaria Populations Using High-Resolution Mass Spectrometry. Metabolites 2020, 10, 238. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhou, L.; Wang, M.; Wei, L.; Qu, H.; Ma, J.; Ju, J.; Han, Z. Chemical Constituents from Marine Derived Fungus Talaromyces cellulolyticus SHJ-3 and Its Chemotaxonomic Significance. Biochem. Syst. Ecol. 2022, 100, 104377. [Google Scholar] [CrossRef]
- Larsen, T.O.; Smedsgaard, J.; Nielsen, K.F.; Hansen, M.E.; Frisvad, J.C. Phenotypic Taxonomy and Metabolite Profiling in Microbial Drug Discovery. Nat. Prod. Rep. 2005, 22, 672. [Google Scholar] [CrossRef] [PubMed]
- Baker, P.M.; Roberts, J.C. Studies in Mycological Chemistry. Part XXI. The Structure of Aurofusarin, a Metabolite of Some Fusarium Species. J. Chem. Soc. C 1966, 2234–2237. [Google Scholar] [CrossRef]
- Shibata, S.; Morishita, E.; Takeda, T.; Sakata, K. The Structure of Aurofusarin. Tetrahedron Lett. 1966, 7, 4855–4860. [Google Scholar] [CrossRef]
- Birchall, G.R.; Bowden, K.; Weiss, U.; Whalley, W.B. The Chemistry of Fungi. Part LVI. Aurofusarin. J. Chem. Soc. C 1966, 2237–2239. [Google Scholar] [CrossRef]
- Gray, J.S.; Martin, G.C.J.; Rigby, W. Aurofusarin. J. Chem. Soc. C 1967, 2578–2580. [Google Scholar] [CrossRef]
- Gea, F.J.; Navarro, M.J.; Suz, L.M. First Report of Cladobotryum mycophilum Causing Cobweb on Cultivated King Oyster Mushroom in Spain. Plant Dis. 2011, 95, 1030. [Google Scholar] [CrossRef]
- Gea, F.J.; Navarro, M.J.; Carrasco, J.; González, A.J.; Suz, L.M. First Report of Cobweb on White Button Mushroom (Agaricus bisporus) in Spain Caused by Cladobotryum mycophilum. Plant Dis. 2012, 96, 1067. [Google Scholar] [CrossRef] [PubMed]
- Gea, F.J.; Carrasco, J.; Suz, L.M.; Navarro, M.J. Characterization and Pathogenicity of Cladobotryum mycophilum in Spanish Pleurotus Eryngii Mushroom Crops and Its Sensitivity to Fungicides. Eur. J. Plant Pathol. 2017, 147, 129–139. [Google Scholar] [CrossRef]
- Gea, F.J.; Navarro, M.J.; Suz, L.M. First Report of Cobweb Disease Caused by Cladobotryum dendroides on Shiitake Mushroom (Lentinula edodes) in Spain. Plant Dis. 2018, 102, 1030. [Google Scholar] [CrossRef]
- Potočnik, I.; Rekanović, E.; Milijasević, S.; Todorović, B.; Stepanović, M. Morphological and Pathogenic Characteristics of the Fungus Cladobotryum dendroides, the Causal Agent of Cobweb Disease of the Cultivated Mushroom Agaricus Bisporus in Serbia. Pestic. Fitomed. 2008, 23, 175–181. [Google Scholar] [CrossRef]
- Milic, N.; Kostidis, S.; Stavrou, A.; Gonou-Zagou, Z.; Kouvelis, V.; Mikros, E.; Fokialakis, N. A Polyphasic Approach (Metabolomics, Morphological and Molecular Analyses) in the Systematics of Cladobotryum Species in Greece. Planta Med. 2012, 78, PG14. [Google Scholar] [CrossRef]
- Riddell, R.W. Permanent Stained Mycological Preparations Obtained by Slide Culture. Mycologia 1950, 42, 265–270. [Google Scholar] [CrossRef]
- Mauri, M.; Elli, T.; Caviglia, G.; Uboldi, G.; Azzi, M. RAWGraphs: A Visualisation Platform to Create Open Outputs. In Proceedings of the 12th Biannual Conference on Italian SIGCHI Chapter, Cagliari, Italy, 18–20 September 2017; ACM: New York, NY, USA, 2017; pp. 1–5. [Google Scholar]
- Inkscape Project; 2021. Available online: https://inkscape.org (accessed on 28 July 2022).
- Kouvelis, V.N.; Ghikas, D.V.; Typas, M.A. The Analysis of the Complete Mitochondrial Genome of Lecanicillium muscarium (Synonym Verticillium lecanii) Suggests a Minimum Common Gene Organization in MtDNAs of Sordariomycetes: Phylogenetic Implications. Funga Genet. Biol. 2004, 41, 930–940. [Google Scholar] [CrossRef]
- Kouvelis, V.N.; Sialakouma, A.; Typas, M.A. Mitochondrial Gene Sequences Alone or Combined with ITS Region Sequences Provide Firm Molecular Criteria for the Classification of Lecanicillium Species. Mycol. Res. 2008, 112, 829–844. [Google Scholar] [CrossRef]
- Burland, T.G. DNASTAR’s Lasergene Sequence Analysis Software. In Bioinformatics Methods and Protocols; Misener, S., Krawetz, S.A., Eds.; Humana Press: Totowa, NJ, USA, 2000; Volume 132, pp. 71–91. ISBN 978-1-59259-192-3. [Google Scholar] [CrossRef]
- Kuraku, S.; Zmasek, C.M.; Nishimura, O.; Katoh, K. ALeaves Facilitates On-Demand Exploration of Metazoan Gene Family Trees on MAFFT Sequence Alignment Server with Enhanced Interactivity. Nucleic Acids Res. 2013, 41, W22–W28. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Wilgenbusch, J.C.; Swofford, D. Inferring Evolutionary Trees with PAUP. Curr. Protoc. Bioinform. 2003, 6.4.1–6.4.28. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Teng, Q.; Huang, W.; Collette, T.W.; Ekman, D.R.; Tan, C. A Direct Cell Quenching Method for Cell-Culture Based Metabolomics. Metabolomics 2009, 5, 199–208. [Google Scholar] [CrossRef]
- Duportet, X.; Aggio, R.B.M.; Carneiro, S.; Villas-Bôas, S.G. The Biological Interpretation of Metabolomic Data Can Be Misled by the Extraction Method Used. Metabolomics 2012, 8, 410–421. [Google Scholar] [CrossRef]
- Matsushima, T. Icones Microfungorum a Matsushima Lectorum; Matsushima: Kobe, Japan, 1975. [Google Scholar]
- Arnold, G.R.W.; Yurchenko, E.O. The First Contribution on Mycophilous Fungi from Belarus. Mycena 2007, 7, 4–19. [Google Scholar]
- Cole, G.T.; Kendrick, B. Conidium Ontogeny in Hyphomycetes. Development and Morphology of Cladobotryum. Can. J. Bot. 1971, 49, 595–599. [Google Scholar] [CrossRef]
- Nair, M.S.R.; Carey, S.T. Metabolites of Pyrenomycetes. XII. Structure of siribinone, a new α-pyrone from Hypomyces. Phytochemistry 1977, 16, 1613. [Google Scholar] [CrossRef]
- Helfer, W. Morphologische, Chemische Und Okologische Characterisierung Mykophiler Pilze. Diploma Thesis, Universitat Regensburg, Regensburg, Germany, 1986. [Google Scholar]
- Nair, M.S.R.; Carey, S.T. Metabolites of Pyrenomycetes IV: Rosellisin, an Antibiotic α-Pyrone from Hypomyces rosellus. Tetrahedron Lett. 1975, 16, 3517–3518. [Google Scholar] [CrossRef]
- Nair, M.S.R. Biogenesis and revised structure of rosellisin. Structure of rosellisin aldhyde. Phytochemistry 1976, 15, 1090–1091. [Google Scholar] [CrossRef]
- Osman, M.; Valadon, L.R.G. Fungistatic Naphthoquinones of Verticillium agaricinum. Trans. Br. Mycol. Soc. 1984, 82, 713–714. [Google Scholar] [CrossRef]
- Huot, R.; Brassard, P. Skyrin from Hypomyces lactifluorum. Phytochemistry 1972, 11, 2879. [Google Scholar] [CrossRef]
- Tezuka, Y.; Huang, Q.; Kikuchi, T.; Nishi, A.; Tubaki, K. Studies on the Metabolites of Mycoparasitic Fungi. I. Metabolites of Cladobotryum varium. Chem. Pharm. Bull. 1994, 42, 2612–2617. [Google Scholar] [CrossRef]
- Tulasne, L.R.; Tulasne, C. Selecta Fungorum Carpologia, Ea Documenta et Icones Potissimum Exhibens; Imperatoris Jussu, In Imperiali Typographeo Excudebatur: Paris, France, 1863; Volume 2, p. 101. [Google Scholar] [CrossRef]
- Roca, M.G.; Davide, L.C.; Mendes-Costa, M.C.; Wheals, A. Conidial Anastomosis Tubes in Colletotrichum. Fungal Genet. Biol. 2003, 40, 138–145. [Google Scholar] [CrossRef]
- Roca, M.G.; Arlt, J.; Jeffree, C.E.; Read, N.D. Cell Biology of Conidial Anastomosis Tubes in Neurospora crassa. Eukaryot. Cell 2005, 4, 911–919. [Google Scholar] [CrossRef]
- Roca, M.G.; Read, N.D.; Wheals, A.E. Conidial Anastomosis Tubes in Filamentous Fungi. FEMS Microbiol. Lett. 2005, 249, 191–198. [Google Scholar] [CrossRef]
- Shulaev, V. Metabolomics Technology and Bioinformatics. Brief. Bioinform. 2006, 7, 128–139. [Google Scholar] [CrossRef]
- Alhadrami, H.A.; Sayed, A.M.; El-Gendy, A.O.; Shamikh, Y.I.; Gaber, Y.; Bakeer, W.; Sheirf, N.H.; Attia, E.Z.; Shaban, G.M.; Khalifa, B.A.; et al. A Metabolomic Approach to Target Antimalarial Metabolites in the Artemisia annua Fungal Endophytes. Sci. Rep. 2021, 11, 2770. [Google Scholar] [CrossRef]
- Baert, F.; Lefevere, P.; D’hooge, E.; Stubbe, D.; Packeu, A. A Polyphasic Approach to Classification and Identification of Species within the Trichophyton benhamiae Complex. J. Fungi 2021, 7, 602. [Google Scholar] [CrossRef]
- Bills, G.F.; Gloer, J.B. Biologically Active Secondary Metabolites from the Fungi. Microbiol. Spectr. 2016, 4, 1–32. [Google Scholar] [CrossRef]
- Moore, D.; Robson, G.D.; Trinci, A.P.J. 21st Century Guidebook to Fungi, 2nd ed.; Cambridge University Press: Cambridge, UK, 2020; ISBN 978-1-108-77638-7. [Google Scholar]
- Keller, N.P. Fungal Secondary Metabolism: Regulation, Function and Drug Discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Pennycook, S.R. The Correct Authorship of the Genus Hypomyces and Its Original Species. Mycotaxon 2009, 108, 185–195. [Google Scholar] [CrossRef]
- Gray, D.J.; Morgan-Jones, G. Notes on Hyphomycetes. XXXIV. Some Mycoparasitic Species. Mycotaxon 1980, 10, 375–404. [Google Scholar]
- Lenti, I. Mikoparazita Gombák A Bátorligeti Természetvédelmi Területeken. Mikológiai Közlemények Clusiana 2008, 47, 21–30. [Google Scholar]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | ATHUM Acc. Nos. | Cladobotryum Species | Fungal Host 1 | Habitat, Geographic Origin |
|---|---|---|---|---|
| 1 | 6907 | C. apiculatum | Russula sp. (RUS, Rus) | Fagus sylvatica forest; Mt. Zigourolivado, Ag. Nikolaos, Karditsa |
| 2 | 6847 | C. dendroides | Flammulina sp. (AGA, Phy) | National Gardens, Athens, Attiki |
| 3 | 6848 | C. dendroides | Polyporus varius (POL, Pol) | Fagus sylvatica forest with Juniperus communis; Mt. Leivaditis, Xanthi |
| 4 | 6849 | C. dendroides | Tricholoma sp. (AGA, Tri) | Abies cephalonica forest; Mt. Parnitha, Attiki |
| 5 | 6909 | C. dendroides | polypore basidiome (POL) | Fagus sylvatica forest; Mt. Zigourolivado, Ag. Nikolaos, Karditsa |
| 6 | 7998 | C. dendroides | Hohenbuehelia sp. (AGA, Ple) | Abies cephalonica forest; Mt. Parnitha, Attiki |
| 7 | 7999 | C. dendroides | Helvella lacunosa (ASC) | Platanus orientalis forest with Quercus sp., Abies cephalonica and Castanea sativa (sporadically); Ag. Nikolaos, Eurytania |
| 8 | 6855 | C. fungicola | Cortinarius sp. (AGA, Cor) | Quercus frainetto, Q. pubescens and Q. coccifera forest with Abies cephalonica (sporadically); Mt. Oiti, Mesochori, Fthiotida |
| 9 | 6906 | C. mycophilum | Hypholoma sp. (AGA, Str) | Castanea sp. forest; Mt. Pilio, Magnisia |
| 10 | 7994 | C. mycophilum | Inocybe sp. (AGA, Ino) | Abies borisii-regis forest; Belοkomitis, Karditsa |
| 11 | 8000 | C. mycophilum | Mycena sp. (AGA, Myc) | Platanus orientalis forest with Quercus sp., Abies cephalonica and Castanea sativa (sporadically); Ag. Nikolaos, Eurytania |
| 12 | 8001 | C. mycophilum | Agaricus bisporus (cultivated) (AGA, Aga) | mushroom cultivation unit |
| 13 | CBS 176.92 ex-type | C. rubrobrunnescens | Inocybe sp. (AGA, Ino) | Regensburg, Germany |
| 14 | CBS 152.92 ex-type | C. tenue | agaric basidiome (AGA) | Keilberg at Regensburg, Germany |
| 15 | 6514 | C. varium | Inocybe sp. (AGA, Ino) | Abies borisii-regis forest; near Krikello, Eurytania |
| 16 | 6845 | C. varium | Clitocybula familia (AGA, Mar) | Abies cephalonica forest; Mt. Parnitha, Attiki |
| 17 | 6846 | C. varium | Panellus sp. (AGA, Myc) | Abies cephalonica forest; Mt. Parnitha, Attiki |
| 18 | 6908 | C. varium | agaric basidiome (AGA) | Fagus sylvatica forest; Mt. Zigourolivado, Ag. Nikolaos, Karditsa |
| 19 | 7995 | C. varium | polypore hymenophore (POL) | no data |
| 20 | 7996 | C. varium | polypore basidiome (POL) | Picea abies, Pinus sylvestris and Betula sp. Mt. Frakto, Rodopi |
| 21 | 8002 | C. varium | Ganoderma sp. (POL, Pol) | Abies borisii-regis forest with Juniperus oxycedrus (sporadically); Mt. Tymfristos, Eurytania |
| 22 | 8003 | C. varium | polypore basidiome (POL) | Abies borisii-regis forest; near Krikello, Eurytania |
| 23 | 6850 | C. verticillatum | Lactarius subumbonatus (RUS, Rus) | Quercus sp. forest; Mt. Lykaio, Megalopoli, Arkadia |
| 24 | 6920 | C. verticillatum | Lactarius subumbonatus (RUS, Rus) | Quercus sp. forest; Mt. Lykaio, Megalopoli, Arkadia |
| 25 | 6921 | C. verticillatum | Lactarius subumbonatus (RUS, Rus) | Quercus sp. forest; Mt. Lykaio, Megalopoli, Arkadia |
| 26 | 6851 | Cladobotryum sp. | Laccaria laccata (AGA, Hyd) | Quercus frainetto, Q. pubescens and Q. coccifera forest with Abies cephalonica (sporadically); Mt. Oiti, Mesochori, Fthiotida |
| 27 | 6852 | Cladobotryum sp. | Lactarius salmonicolor (RUS, Rus) | Quercus frainetto, Q. pubescens and Q. coccifera forest with Abies cephalonica (sporadically); Mt. Oiti, Mesochori, Fthiotida |
| 28 | 6853 | Cladobotryum sp. | Russula sp. (RUS, Rus) | Quercus frainetto, Q. pubescens and Q. coccifera forest with Abies cephalonica (sporadically); Mt. Oiti, Mesochori, Fthiotida |
| 29 | 6904 | Cladobotryum sp. | Lactarius sp. (RUS, Rus) | Fagus sylvatica forest; Mt. Zigourolivado, Ag. Nikolaos, Karditsa |
| 30 | 6912 | Cladobotryum sp. | Tricholoma atrosquamosum (AGA, Tri) | Abies cephalonica forest; Mt. Parnitha, Attiki |
| 31 | 6913 | Cladobotryum sp. | Lactarius salmonicolor (RUS, Rus) | Abies cephalonica forest; Mt. Parnitha, Attiki |
| 32 | 6914 | Cladobotryum sp. | Cortinarius sp. (AGA, Cor) | Abies cephalonica forest; Mt. Parnitha, Attiki |
| # | Species | Isolate or Specimen Number | ITS GenBank Acc. Nos. | Country of Origin |
|---|---|---|---|---|
| 1 | C. amazonense | CBS 47080 | MH861285.1 | South Africa |
| 2 | C. apiculatum | CBS 829.69 | MH859447.1 | South Africa |
| 3 | C. apiculatum | CBS 174.56 | NR_159770.1 | Japan |
| 4 | C. apiculatum | ATHUM6907 | OM993303 | Greece |
| 5 | C. asterophorum | TFC 97-23 | Y17089.1 | Australia |
| 6 | C. asterophorum | CBS 676.77 | NR_111426.1 | Japan |
| 7 | C. croceum | CBS 23195 | MH862511.1 | South Africa |
| 8 | C. cubitense | CBS 416.85 | NR_111427.1 | Cuba |
| 9 | C. dendroides | ATHUM6909 | OM993302 | Greece |
| 10 | C. dendroides | ATHUM6848 | OM993306 | Greece |
| 11 | C. dendroides | ATHUM6849 | OM993310 | Greece |
| 12 | C. dendroides | ATHUM7999 | OM993313 | Greece |
| 13 | C. dendroides | ATHUM7998 | OM993314 | Greece |
| 14 | C. dendroides | ATHUM6847 | OM993320 | Greece |
| 15 | C. dendroides | PSU 177 | Y17091.1 | United States |
| 16 | C. dendroides | MUCL 28202 | Y17092.1 | Luxembourg |
| 17 | C. fungicola | ATHUM6855 | OM993323 | Greece |
| 18 | C. heterosporum | CBS 71988 | FN859398.1 | Cuba |
| 19 | C. heterosporum | CBS 719.88 | NR_111428.1 | Cuba |
| 20 | C. indoafrum | CBS 127163 | MH864452.1 | South Africa |
| 21 | C. multiseptatum | CBS 472.71 | NR_121423.1 | New Zealand |
| 22 | C. mycophilum | ATHUM6906 | OM993304 | Greece |
| 23 | C. mycophilum | ATHUM8001 | OM993311 | Greece |
| 24 | C. mycophilum | ATHUM8000 | OM993312 | Greece |
| 25 | C. mycophilum | ATHUM7994 | OM993316 | Greece |
| 26 | C. mycophilum | IMI 267134 | Y17095.1 | United Kingdom |
| 27 | C. mycophilum | CBS 818.69 | Y17096.1 | The Netherlands |
| 28 | C. obconicum | CBS 52881 | MH861373.1 | South Africa |
| 29 | C. obconicum | CBS 528.81 | NR_172286.1 | Belgium |
| 30 | C. paravirescens | TFC 97-23 | NR_121424.1 | Thailand |
| 31 | C. penicillatum | CBS 40780 | FN859407.1 | The Netherlands |
| 32 | C. pinarense | CBS 40086 | MH861973.1 | South Africa |
| 33 | C. protrusum | IMI 165503 | FN859412.1 | Zimbabwe |
| 34 | C. protrusum | TFC 201316 | FN859414.1 | Madagascar |
| 35 | C. purpureum | CBS 154.78 | FN859415.1 | USA |
| 36 | C. rubrobrunnescens | CBS 176.92 | FN859416.1 | Germany |
| 37 | C. rubrobrunnescens | CBS 176.92 | NR_111429.1 | Germany |
| 38 | C. semicirculare | TFC 03-3 | FN859418.1 | Sri Lanka |
| 39 | C. soroaense | INIFATC86/7 | HE792978.1 | Cuba |
| 40 | C. soroaense | INIFATC86/7 | NR_145041.1 | Cuba |
| 41 | Cladobotryum sp. | TFC 201294 | FN859423.1 | Madagascar |
| 42 | Cladobotryum sp. | PPRI 11269 | KY069830.1 | N/A |
| 43 | Cladobotryum sp. | PPRI 13579 | KY069856.1 | N/A |
| 44 | Cladobotryum sp. | PPRI 13554 | KY767656.1 | N/A |
| 45 | Cladobotryum sp. | KZN-2 | MH455274.1 | South Africa |
| 46 | Cladobotryum sp. | GP-2 | MH489096.1 | South Africa |
| 47 | Cladobotryum sp. | WC-4 | MH489098.1 | South Africa |
| 48 | Cladobotryumsp. | ATHUM6904 | OM993297 | Greece |
| 49 | Cladobotryumsp. | ATHUM6914 | OM993299 | Greece |
| 50 | Cladobotryumsp. | ATHUM6913 | OM993300 | Greece |
| 51 | Cladobotryumsp. | ATHUM6912 | OM993301 | Greece |
| 52 | Cladobotryumsp. | ATHUM6852 | OM993308 | Greece |
| 53 | Cladobotryumsp. | ATHUM6851 | OM993309 | Greece |
| 54 | Cladobotryumsp. | ATHUM6853 | OM993321 | Greece |
| 55 | C. stereicola | CBS 457.71 | NR_159773.1 | Russia |
| 56 | C. tchimbelense | TFC 201146 | NR_121426.1 | Gabon |
| 57 | C. tchimbelense | TFC 201146 | FN859419.1 | Gabon |
| 58 | C. tenue | CBS 152.92 | NR_121427.1 | Germany |
| 59 | C. varium | ATHUM6846 | OM993307 | Greece |
| 60 | C. varium | ATHUM6514 | OM993317 | Greece |
| 61 | C. varium | ATHUM7995 | OM993318 | Greece |
| 62 | C. varium | ATHUM6908 | OM993319 | Greece |
| 63 | C. varium | ATHUM8002 | OM993322 | Greece |
| 64 | C. varium | ATHUM6845 | OM993324 | Greece |
| 65 | C. varium | ATHUM8003 | OM993325 | Greece |
| 66 | C. varium | ATHUM7996 | OM993326 | Greece |
| 67 | C. verticillatum | ATHUM6921 | OM993298 | Greece |
| 68 | C. verticillatum | ATHUM6850 | OM993305 | Greece |
| 69 | H. australis | FJ810168 | China | |
| 70 | H. aconidialis | TFC 201334 | FN859457.1 | Madagascar |
| 71 | H. armeniacus | TFC 02-86/2 | FN859424.1 | France |
| 72 | H. aurantius | AB298708.1 | Japan | |
| 73 | H. aurantius | CV01 | MW709638.1 | China |
| 74 | H. aurantius | MUCL 8223 | Y17088.1 | Canada |
| 75 | H. australasiaticus | TFC 03-8 | NR_121428.1 | Sri Lanka |
| 76 | H. australasiaticus | TFC 99-95 | FN859427.1 | Australia |
| 77 | H. dactylarioides | NLB 1512 | MT537005.1 | Australia |
| 78 | H. dactylarioides | CBS 141.78 | NR_111430.1 | New Zealand |
| 79 | H. gabonensis | TFC 201156 | NR_121429.1 | Gabon |
| 80 | H. khaoyaiensis | GJS01-304 | FN859431.1 | Thailand |
| 81 | H. lactifluorum | TAAM170476 | FN859432.1 | USA |
| 82 | H. odoratus | CBS 819.69 | MH859441.1 | The Netherlands |
| 83 | H. odoratus | CBS 820.69 | MH859442.1 | The Netherlands |
| 84 | H. odoratus | MK044830.1 | China | |
| 85 | H. rosellus | isolate LE01 | MF992212.1 | Spain |
| 86 | H. samuelsii | InBio3-233 | FN859450.1 | Costa Rica |
| 87 | H. samuelsii | CBS 536.88 | FN859444.1 | Cuba |
| 88 | H. samuelsii | CBS 127157 | NR_121430.1 | Puerto Rico |
| 89 | H. semicircularis | CBS 705.88 | NR_121425.1 | Cuba |
| 90 | H. semitranslucens | CBS 821.70 | MH859960.1 | Sweden |
| 91 | H. semitranslucens | CBS 458.71 | MH860218.1 | Russia |
| 92 | H. subiculosus | TFC 97-166 | FN859452.1 | Puerto Rico |
| 93 | Fusarium sp. | C1032B | OM993315 | Greece |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milic, N.; Christinaki, A.C.; Benaki, D.; Stavrou, A.A.; Tsafantakis, N.; Fokialakis, N.; Kouvelis, V.N.; Gonou-Zagou, Z. Polyphasic Systematics of the Fungicolous Genus Cladobotryum Based on Morphological, Molecular and Metabolomics Data. J. Fungi 2022, 8, 877. https://doi.org/10.3390/jof8080877
Milic N, Christinaki AC, Benaki D, Stavrou AA, Tsafantakis N, Fokialakis N, Kouvelis VN, Gonou-Zagou Z. Polyphasic Systematics of the Fungicolous Genus Cladobotryum Based on Morphological, Molecular and Metabolomics Data. Journal of Fungi. 2022; 8(8):877. https://doi.org/10.3390/jof8080877
Chicago/Turabian StyleMilic, Nikola, Anastasia C. Christinaki, Dimitra Benaki, Aimilia A. Stavrou, Nikolaos Tsafantakis, Nikolas Fokialakis, Vassili N. Kouvelis, and Zacharoula Gonou-Zagou. 2022. "Polyphasic Systematics of the Fungicolous Genus Cladobotryum Based on Morphological, Molecular and Metabolomics Data" Journal of Fungi 8, no. 8: 877. https://doi.org/10.3390/jof8080877