
Identification and Characterization of Colletotrichum Species Causing Sorghum Anthracnose in Kenya and Screening of Sorghum Germplasm for Resistance to Anthracnose
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Diseased Sorghum Leaf Samples, Isolation and Purification of Fungi
2.2. Pathogenicity Tests of Fungal Isolates
2.3. Morpho-Cultural Characterization
2.4. DNA Extraction, Polymerase Chain Reaction (PCR) and Sequencing
2.5. Sequence Alignment and Phylogenetic Analysis
2.6. Screening of Sorghum Germplasm for Resistance to Anthracnose
2.6.1. Greenhouse Screening Trials

{kind=link}
{kind=link}
{kind=link}
| Severity Ratings | Symptoms and Lesion Type on the Leaves | DRC |
|---|---|---|
| 1 | 0 to <1% leaf area covered with hypersensitive reaction with mild yellow flecks | R |
| 2 | 1–5% leaf area covered with hypersensitive lesions without acervuli | MR |
| 3 | 6–10% leaf area covered with hypersensitive lesions without acervuli | MS |
| 4 | 11–20% leaf area covered with hypersensitive and restricted necrotic lesions with acervuli | |
| 5 | 21–30% leaf area covered with hypersensitive and restricted necrotic lesions with acervuli | S |
| 6 | 31–40% leaf area covered with coalescing necrotic lesions with acervuli | |
| 7 | 41–50% leaf area covered with coalescing necrotic lesions with acervuli | HS |
| 8 | 51–75% leaf area covered with coalescing necrotic lesions with acervuli | |
| 9 | 76–100% leaf area covered with coalescing necrotic lesions with acervuli |
2.6.2. Field Screening Trials
3. Results
3.1. Isolation and Pathogenicity Fungal Isolates
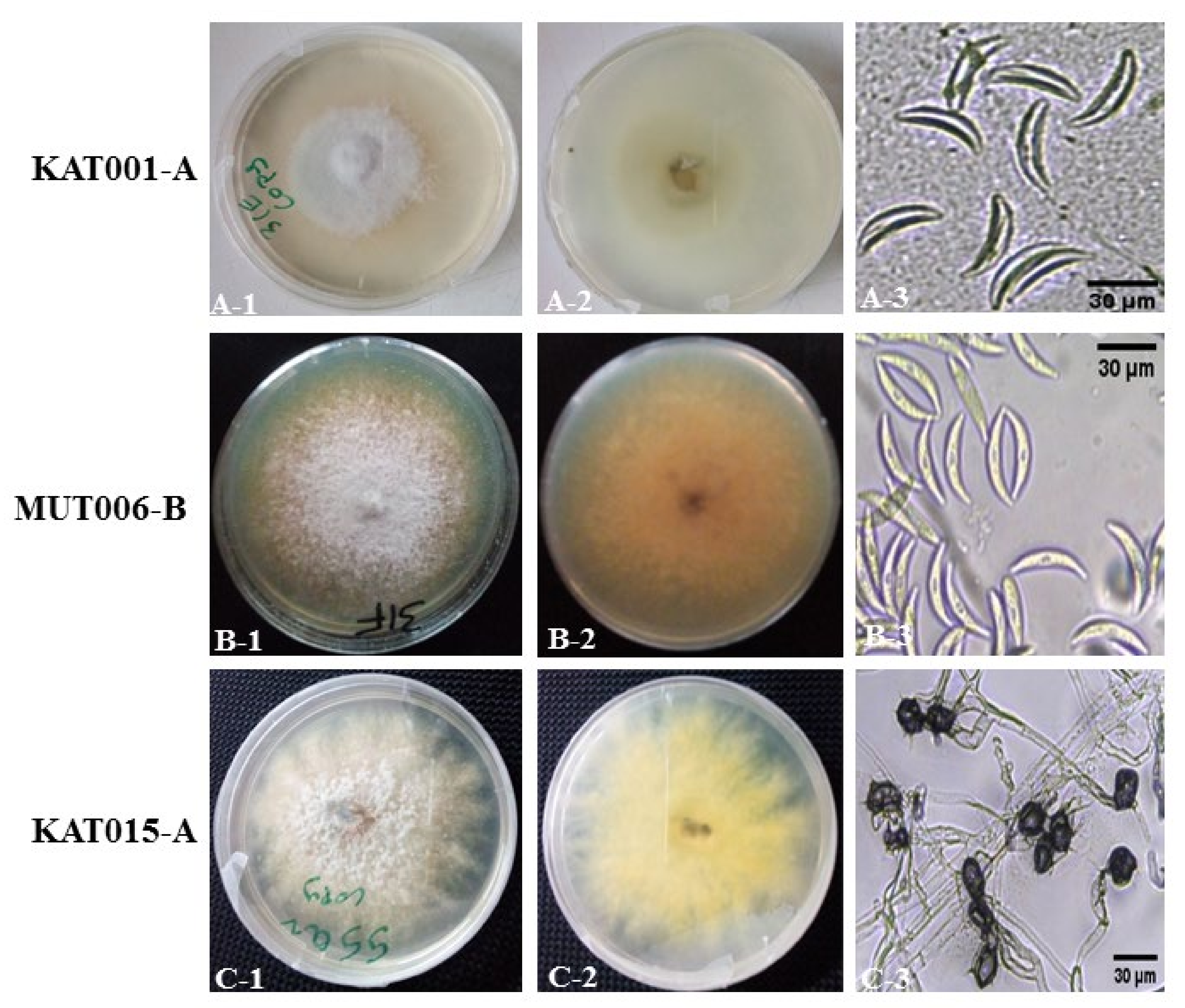
3.2. Morpho-Cultural Characterization of Colletotrichum spp. Isolates
3.3. PCR Amplification and DNA Sequencing
3.4. Phylogenetic Analysis of ITS Region
3.5. Reactions of Sorghum Genotypes to Colletotrichum Sublineola
3.6. Grain Yield, 100 Seed Weight and Harvest Index
3.7. Regression Analysis on Effect of Anthracnose on Grain Weight, Grain Yield and Weight of 100 Seeds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christopher, R.; Ramasamy, P.; Tesfaye, T.; Louis, K.; Gary, N.; Clint, W. Sorghum pathology and biotechnology—A fungal disease perspective: Part I. Grain mold, Head smut and Ergot. Eur. J. Plant Sci. Biotechnol. 2012, 6, 10–30. [Google Scholar]
- Thakur, R.P.; Mathur, K. Anthracnose. In Compendium of Sorghum Diseases; Frederiksen, R.A., Odvody, G.N., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 2000; pp. 10–12. [Google Scholar]
- Prom, L.K.; Cuevas, H.; Ramasamy, P.; Thomas, I.; Clint, M. Inheritance of resistance of three sorghum lines to pathotypes of Colletotrichumsublineola, causal agent of anthracnose. Plant Pathol. J. 2018, 17, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Mengistu, G.; Shimelis, H.; Laing, M.; Lule, D. Breeding for anthracnose (Colletotrichumsublineolum Henn.) resistance in sorghum: Challenges and opportunities. Aust. J. Crop. Sci. 2018, 12, 1911–1920. [Google Scholar] [CrossRef]
- Little, R.; Perumal, R.; Tesso, T.; Kofoid, K.; Prasad, V.; Aiken, R.; Bean, S.; Wilson, J.; Herald, T. Registration of nine grain sorghum seed parent lines. J. Plant Regist. 2018, 9, 244–248. [Google Scholar]
- Tesso, T.; Perumal, R.; Little, R.; Adeyanju, A.; Radwan, G.; Prom, L.K.; Magill, C. Sorghum pathology and biotechnology—A fungal disease perspective: Part II. Anthracnose, stalk rot, and downy mildew. Eur. J. Plant Sci. Biotechnol. 2012, 6, 31–44. [Google Scholar]
- Chala, A.; Alemu, T.; Prom, L.K.; Tronsmo, A.M. Effect of host genotypes and weather variables on the severity and temporal dynamics of sorghum anthracnose in Ethiopia. Plant Pathol. J. 2010, 9, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Upadhyaya, H.; Manjunatha, S.; Rao, V.; Thakur, R. Resistance of foliar disease in a mini-core collations of sorghum germplasm. Plant Di. J. 2012, 96, 1629–1633. [Google Scholar] [CrossRef] [Green Version]
- Marley, P.; Diourte, M.; Neya, A.; Rattude, F. Sorghum anthracnose and sustainable management strategies in West and Cental Africa. J. Sustain. Agric. 2005, 25, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Ngugi, H.K.; Julian, A.M.; King, S.B.; Peacocke, B.J. Epidemiology of sorghum anthracnose (Colletotrichum sublineolum) and leaf blight (Exserohilum turcicum) in Kenya. Plant Pathol. 2000, 49, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Khodadadi, F.; Gonzalez, J.B.; Martin, P.L.; Giroux, E.; Bilodeau, G.J.; Peter, K.A.; Doyle, V.P.; Acimovic, S.G. Identification and characterization of Colletotrichums pecies causing apple bitter rot in New York and description of C. noveboracense sp. nov. Sci. Rep. 2020, 10, 11043. [Google Scholar] [CrossRef]
- Cai, L.; Hyde, K.D.; Taylor, P.W.J.; Weir, B.; Waller, J.; Abang, M.M.; Zhang, J.Z.; Yang, Y.L.; Phoulivong, S.; Liu, Z.Y.; et al. A polyphasic approach for studying Colletotrichum. Fungal Divers. 2009, 39, 183–204. [Google Scholar]
- Bailey, J.A.; O’Connell, R.J.; Pring, R.J.; Nash, C. Infection strategies of Colletotrichum species. In Colletotrichum: Biology, Pathology and Control; Bailey, J.A., Jeger, M.J., Eds.; CAB International: Wallingford, UK, 1992; pp. 88–120. [Google Scholar]
- Katan, T. Vegetative compatibility in Colletotrichum. In Colletotrichum: Host Specificity, Pathology and Host Pathogen Interaction; Prusky, D., Freeman, S., Dickman, M., Eds.; APS Press: St. Paul, MN, USA, 2000. [Google Scholar]
- Hyde, K.D.; Cai, L.; McKenzie, E.H.C.; Yang, Y.L.; Zhang, J.Z.; Prihastuti, H. Colletotrichum: A catalogue of confusion. Fungal Divers. 2009, 39, 1–17. [Google Scholar]
- Crouch, J.A.; Clarke, B.B.; White, J.F.; Hillman, B.I. Systematic analysis of the falcate-spored graminicolous Colletotrichum and a description of six new species of the fungus from warm season grasses. Mycologia 2009, 101, 717–732. [Google Scholar] [CrossRef]
- Crouch, J.A.; Clarke, B.B.; Hillman, B.I. What is the value of ITS sequence data in Colletotrichum systematics and species diagnosis? A case study using the falcate-spored graminicolous Colletotrichum group. Mycologia 2009, 101, 648–656. [Google Scholar] [CrossRef]
- Cannon, P.; Damm, U.; Johnston, R.; Weir, B. Colletotrichum—current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef]
- Freeman, S.; Katan, T.; Shabi, E. Characterization of Colletotrichum species responsible for anthracnose diseases of various fruit. Plant Dis. 1998, 82, 596–605. [Google Scholar] [CrossRef] [Green Version]
- Peres, N.A.R.; Souza, N.L.; Peever, T.L.; Timmer, L.W. Benomyl sensitivity of isolates of Colletotrichumacutatum and C. gloeosporioides from citrus. Plant Dis. 2004, 88, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Vieira, W.A.S.; Lima, W.G.; Nascimento, E.S.; Michereff, S.J.; Camara, M.P.S.; Doyle, V.P. The impact of phenotypic and molecular data on the inference of Colletotrichum diversity associated with Musa. Mycologia 2017, 109, 912–934. [Google Scholar] [CrossRef]
- Hess, D.E.; Bandyopadhyay, R.; Sissoko, I. Pattern analysis of sorghum genotype × environment interaction for leaf, panicle, and grain anthracnose in Mali. Plant Dis. 2002, 86, 1374–1382. [Google Scholar] [CrossRef] [Green Version]
- Mofokeng, M.; Hussein, S.; Mark, L.; Nemera, S. Sorghum (Sorghum bicolor) breeding for resistance to leaf and stalk anthracnose, Colletotrichum sublineolum, and improved yield: Progress and prospects. Aust. J. Crop Sci. 2017, 11, 1078–1085. [Google Scholar] [CrossRef]
- Cuevas, H.E.; Prom, L.K.; Erpelding, J.E.; Brotons, V. Assessments of genetic diversity and anthracnose disease response among Zimbabwe sorghum germplasm. Plant Breed. 2014, 133, 234–242. [Google Scholar] [CrossRef]
- Cuevas, H.E.; Prom, L.K.; Isakeit, T.; Radwan, G. Assessment of sorghum germplasm from Burkina Faso and South Africa to identify new sources of resistance to grain mold and anthracnose. Crop Protect. 2016, 79, 43–50. [Google Scholar] [CrossRef]
- Erpelding, J.E.; Prom, L.K. Evaluation of Malian sorghum germplasm for resistance against anthracnose. Plant Pathol. J. 2004, 3, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Erpelding, J.E.; Prom, L.K. Variation for anthracnose resistance within the sorghum germplasm collection from Mozambique, Africa. Plant Pathol. J. 2006, 5, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Prom, L.K.; Perumal, R.; Erattaimuthu, S.; Erpelding, J.; Montes, N.; Odvody, G.; Greenwald, C.; Jin, Z.; Frederiksen, R.; Magill, C. Virulence and molecular genotyping studies of Sporisoriumreilianum isolates in sorghum. J. Plant Dis. 2011, 95, 523–529. [Google Scholar] [CrossRef]
- Prom, L.K.; Erpelding, J.; Perumal, R.; Isakeit, T.; Cuevas, H. Response of sorghum accessions from four African countries against, causal agent of sorghum anthracnose. Am. J. Plant Sci. 2012, 3, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Dean, R.; Erpelding, J.; Pederson, G. Molecular genetics evaluation of sorghum germplasm differing in response to fungal diseases: Rust (Pucciniapurpurea) and anthracnose (Colletotrichumgramicola). Euphytica 2006, 148, 319–330. [Google Scholar] [CrossRef]
- Xavier, K.V.; Mizubuti, E.S.G.; Queiroz, M.V.; Chopra, S.; Vaillancourt, L. Genotypic and pathogenic diversity of Colletotrichum sublineola isolates from sorghum (Sorghum bicolor) and Johnson grass (S. halepense) in the south eastern United States. Plant Dis. J. 2018, 102, 2341–2351. [Google Scholar] [CrossRef] [Green Version]
- Kutawa, A.B.; Sijam, K.; Ahmd, K.; Zulkifli, A.S.; Sharif, M.F.; Norzihan, A. Characterization and pathological variability of Exserohilum turcicum responsible for causing northern corn leaf blight (NCLB) disease in Malaysia. Malays. J. Microbiol. 2017, 13, 41–49. [Google Scholar]
- Binyam, T.; Girma, A.; Fikre, L. Distribution and importance of sorghum anthracnose (Colletotrichum sublineolum) in south western Ethiopia. Plant Pathol. J. 2016, 15, 75–85. [Google Scholar]
- Kimaru, S.; Monda, E.; Cheruiyot, R.; Mbaka, J.; Alakonya, A. Morphologicaland molecular identification of the causal agent of anthracnose disease of Avocado in Kenya. Int. J. Microbiol. 2018, 4, 4568520. [Google Scholar]
- Ouoba, L.; Vouidibio-Mbozo, B.; Anyogu, A. Environmental heterogeneity of Staphylococcus species from alkaline fermented foods and associated toxins and antimicrobial resistance genetic elements. Int. J. Food Microbiol. 2019, 311, 108356. [Google Scholar] [CrossRef] [Green Version]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfaud, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; Volume 1, pp. 315–322. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. J. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ramathani, T.; Mahmoud, M.; Samawaty, A.; Sayed, S. Non liquid nitrogen-based method for isolation of DNA from filamentous fungi. Afr. J. Biotechnol. 2011, 10, 14337–14341. [Google Scholar]
- Prom, L.K.; Cuevas, T.; Isakeit, T.; Droleskey, R. Excised leaf method for high volume evaluation of sorghum germplasm for resistance against Colletotrichum sublineolum. Plant Pathol. J. 2016, 15, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Thakur, R.P. Anthracnose. In Screening Techniques for Sorghum Diseases; Thakur, R.P., Reddy, B., Mathur, K., Eds.; International Crops Research Institute for The Semi-Arid Tropics: Patancheru, India, 2007; pp. 53–57. [Google Scholar]
- SAS. SAS Guide for Personal Computers, Version 9.2; SAS Institute Inc.: Cary, NC, USA, 2008. [Google Scholar]
- Madden, L.V.; Hughes, G.; van den Bosch, F. The Study of Plant Disease Epidemics; APS Press: St. Paul, MN, USA, 2007. [Google Scholar]
- Xu, J.; Liu, K.J.; Hu, L.; Jiang, Y.; Zhang, M.H.; Xu, X.D. Rule for Evaluation of Sorghum Resistance to Anthracnose; Liaoning Provincial Bureau of Quality and Technical Supervision: Shenyang, China, 2017.
- Mathur, K.; Thakur, R.P.; Neya, A.; Marley, P.S.; Casela, C.R. Sorghum anthracnose—Problems and management strategies. In Sorghum and Millet Diseases; Leslie, J., Ed.; Iowa State Press: Ames, IA, USA, 2003; pp. 211–220. [Google Scholar]
- Jactzold, R.; Schmidt, H. Farm Management Handbook of Kenya; Ministry of agriculture: New Delhi, India, 2010.
- Choi, H.; Hong, K.; Lee, Y.; Yoon, Y. First report of Colletotrichum sublineola causing anthracnose on Sorghum bicolor in South Korea. Am. Physic. Soc. 2021, 10, 1094. [Google Scholar] [CrossRef]
- Gabriela, F.Z.; Gisele, M.A.N.; Luzia, D.P.M. Morphological Characterization of C. sublineola Strains, causal agent of anthracnose of sorghum. Trop. Plant Pathol. J. 2009, 34, 146–151. [Google Scholar]
- Prom, L.K.; Isakeit, T.; Cuevas, H.; Rooney, L.; Perumal, R.; Magill, W. Reaction of sorghum lines of zonate leaf spot and rough leaf spot. Plant Health Prog. J. 2015, 10, 15–0040. [Google Scholar] [CrossRef] [Green Version]
- Prom, L.K.; Perumal, R.; Isakeit, T.; Radwan, G.; Rooney, W.; Magill, C. The impact of weather conditions on response of sorghum genotypes to anthracnose (Colletotrichum sublineolum) infection. Am. J. Exp. Agric. 2015, 6, 242–250. [Google Scholar]
- Kamdoum, E.; Tsopmbeng, G.; Kuiate, J. Cultural and morphological variations of Colletotrichum spp. associated with anthracnose of various fruits in Cameroon. Int. J. Environ. Agric. Biotech. 2016, 1, 968–974. [Google Scholar]
- Hassan, O.; Lee, Y.S.; Chang, T. Colletotrichum species associated with Japanese plum (Prunussalicina) anthracnose in South Korea. Sci. Rep. 2019, 9, 12089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowling, M.; Peres, N.; Villani, S.; Schnabel, G. Managing Colletotrichum on fruit crops: A “complex” challenge. Plant Dis. 2020, 104, 2301–2316. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prom, L.K.; Ahn, E.; Isakeit, T.; Magill, C. GWAS analysis of sorghum association panel lines identifies SNPs associated with disease response to Texas isolates of Colletotrichum sublineola. Theor. Appl. Genet. 2019, 132, 1389–1396. [Google Scholar] [CrossRef]
- Abreha, K.B.; Ortiz, R.; Carlsson, A.S.; Geleta, M. Understanding the sorghum–Colletotrichum sublineola interactions for enhanced host resistance. Front. Plant Sci. 2021, 12, 641969. [Google Scholar] [CrossRef]
- Xavier, K.V.; Pfeiffer, T.; Parreira, D.; Chopra, S.; Vaillancourt, L. Aggressiveness of Colletotrichum sublineola strains from Sorghum bicolor and S. halepenseto sweet sorghum variety Sugar Drip, and their impact on yield. Plant Dis. 2017, 101, 1578–1587. [Google Scholar] [CrossRef] [Green Version]
- Crouch, J.A.; Tomaso, P.M. Anthracnose disease of centipegrass turf caused by Colletotrichum eremochloae, a fungal species closely related to Colletotrichum sublineola. Mycologia 2021, 104, 1085–1096. [Google Scholar] [CrossRef]



| Severity Ratings | Symptoms and Lesion Type on the Leaves | DRC |
|---|---|---|
| 1 | No visible symptoms/chlorotic flecks | R |
| 2 | 1–25% leaf area covered with small, restricted lesions | MR |
| 3 | 26−50% leaf area covered with small, restricted lesions | MS |
| 4 | 51−75% leaf area covered with large, coalescing lesions | S |
| 5 | >75% leaf area covered with large, coalescing lesions | HS |
| Isolate ID Code | County | Location | Mycelial Characteristics | Colony Size (cm) | Colony Growth Rate/Day (cm) | Conidia | Macroscopic Characteristics | Microscopic Characteristics | Pathogen Identity | |
|---|---|---|---|---|---|---|---|---|---|---|
| Length (µm) (Average ± SD *) | Width (µm) (Average ± SD *) | |||||||||
| KAT015-A | Makueni | Kathonzweni | Raised | 7.46 b | 0.82 c | 34.47 ± 2.14 a | 5.12 ± 1.10 a | Top—White Reverse—Yellowish | Falcate, No septation and hyaline | Colletotrichum sublineola |
| KT001-A | Kitui | Kitui township | Cottony | 8.5 ab | 0.98 a | 32.47 ± 2.14 a | 4.59 ± 1.10 b | Top—Cotton Reverse—Whitish centers | Falcate, hyaline with no septation | Colletotrichum sublineola |
| W008-L | Makueni | Wote | Cottony | 7.6 b | 0.9 b | 31.41 ± 5.37 a | 4.47 ± 0.53 b | Top—Cotton Reverse—Whitish with dark centers | Falcate, hyaline with no septation | Colletotrichum sublineola |
| MUT006-B | Kitui | Mutomo | Raised and cottony | 7.69 b | 0.88 bc | 35.27 ± 2.14 a | 5.02 ± 1.14 ab | Top—white Reverse-pale orange | Falcate, slender, hyaline and no septation | Colletotrichum sublineola |
| MAK017-D | Makueni | Makindu | Raised and cottony | 6.03 bc | 0.74 cd | 34.47 ± 2.14 a | 5.12 ± 1.10 a | Top—White Reverse—Yellowish | Falcate, No septation and hyaline | Colletotrichum sublineola |
| IKA009-G | Kitui | Ikanga | Raised and cottony | 7.39 b | 0.88 bc | 23 ± 1.92 b | 4.91 ± 0.51 ab | Top—White Reverse—Yellowish | Falcate, No septation and hyaline | Colletotrichum sublineola |
| MTH022-A | Machakos | Muthesya | Raised and cottony | 5.44 c | 0.54 d | 31.41 ± 5.37 a | 5.29 ± 0.81 a | Top—White Reverse—Yellowish | Falcate, No septation and hyaline | Colletotrichum sublineola |
| Isolate ID Code | County | ITS Accession | Closest Match in Blast | Size (bp) | Similarity (%) |
|---|---|---|---|---|---|
| KAT015-A | Makueni | ON764330 | Colletotrichum sublineola | 567 | 100 |
| KT001-A | Kitui | ON764342 | Colletotrichum sublineola | 538 | 100 |
| W008-L | Makueni | ON764382 | Colletotrichum sublineola | 549 | 100 |
| MUT006-B | Kitui | ON764366 | Colletotrichum sublineola | 564 | 100 |
| MAK017-D | Makueni | ON764351 | Colletotrichum sublineola | 563 | 100 |
| IKA009-G | Kitui | ON764322 | Colletotrichum sublineola | 564 | 100 |
| MTH022-A | Machakos | ON764362 | Colletotrichum sublineola | 516 | 100 |
| Genotype | AUDPC | Disease Severity Score | DRC |
|---|---|---|---|
| Dark Red | 5.57 bc | 2.78 bc | MR |
| Gadam | 5.94 ab | 2.97 bc | MR |
| IESV 24,029 SH | 5.17 c | 2.58 c | MR |
| Kaguru | 7 a | 3.5 b | MS |
| KARI Mtama-1 | 2 e | 1 e | R |
| Kateng’u | 6.7 ab | 3.35 b | MS |
| Kauwi | 3.8 d | 1.9 d | R |
| Kiboko Local-2 | 5.1 c | 2.55 c | MR |
| Makueni Local-2 | 6.52 ab | 3.27 bc | MS |
| Marcia | 2 e | 1 e | R |
| Mugeta | 2 e | 1 e | R |
| Rasta | 7.67 a | 3.83 ab | MS |
| Seredo | 5.77 b | 2.88 bc | MR |
| Serena | 5.47 bc | 2.73 c | MR |
| Kiboko Season 1 | Kiboko Season 2 | |||||
|---|---|---|---|---|---|---|
| AUDPC | Severity | DRC | AUDPC | Severity | DRC | |
| Dark Red | 35.16 e | 2.38 c | MR | 51.03 d | 3.31 bc | MS |
| Gadam | 50.46 de | 3.07 b | MS | 59.71 c | 3.49 bc | MS |
| IESV 24,029 SH | 47.33 de | 3.09 b | MS | 54.75 cd | 3.3 c | MS |
| Kaguru | 57.8 b | 3.64 a | S | 76.83 a | 4.34 a | S |
| KARI Mtama-1 | 20 ef | 1 d | R | 20 f | 1 f | R |
| Kateng’u | 54.7 d | 3.42 a | MS | 75.29 a | 4.29 a | S |
| Kauwi | 12.94 f | 1.14 d | R | 25.75 e | 1.48 d | R |
| Kiboko Local-2 | 54.41 d | 3.26 ab | MS | 63.97 b | 3.74 b | S |
| Makueni Local-2 | 55.66 d | 3.5 a | S | 64.42 b | 3.83 ab | S |
| Marcia | 20 ef | 1 d | R | 20 f | 1 f | R |
| Mugeta | 20.79 ef | 1.09 d | R | 22.42 f | 1.2 e | R |
| Rasta | 64.27 a | 3.81 a | S | 78.46 a | 4.39 a | S |
| Seredo | 33.5 e | 2.33 c | MR | 47 de | 3.12 c | MS |
| Serena | 51.19 de | 3.08 b | MS | 57.92 c | 3.44 bc | MS |
| Ithookwe Season 1 | Ithookwe Season 2 | |||||
|---|---|---|---|---|---|---|
| AUDPC | Severity | DRC | AUDPC | Severity | DRC | |
| Dark Red | 34.89 d | 2.7 c | MS | 44.98 b | 3.01 b | MS |
| Gadam | 74.11 a | 4.1 a | S | 76.01 a | 4.3 a | S |
| IESV 24,029 SH | 52.21 b | 3.49 b | MS | 46.5 b | 3.11 b | MS |
| Kaguru | 43.18 bc | 3.1 bc | MS | 70 a | 4.37 a | S |
| KARI Mtama 1 | 20 e | 1 d | R | 20 d | 1 d | R |
| Kateng’u | 53.83 b | 3.61 b | S | 75.64 a | 4.4 a | MS |
| Kauwi | 22.33 e | 1.27 d | R | 26.63 c | 1.55 c | R |
| Kiboko Local-2 | 40.42 c | 2.97 bc | MS | 46.83 b | 3.06 b | MS |
| Makueni Local-2 | 36.61 d | 2.79 c | MS | 48.67 b | 3.21 b | MS |
| Marcia | 20 e | 1 d | R | 20 d | 1 d | R |
| Mugeta | 21.08 e | 1.14 d | R | 22.02 d | 1.17 d | R |
| Rasta | 43.02 bc | 3.11 bb | MS | 66.75 a | 4.17 a | S |
| Seredo | 35.52 d | 2.73 c | MS | 46.63 b | 3.07 b | MS |
| Serena | 34.9 d | 2.7 c | MS | 47.58 b | 3.1 b | MS |
| KALRO-Ithookwe-2021 (Season 1 & 2) | KALRO-Kiboko-2021 (Season 1 & 2) | |||||||
|---|---|---|---|---|---|---|---|---|
| * Grain Weight (g) | # Grain Yield (t/ha) | 100 Seeds Weight (g) | $ Harvest Index | * Grain Weight (g) | # Grain Yield (t/ha) | 100 Seeds Weight (g) | $ Harvest Index | |
| Gadam | 35.7 d | 1.8 h | 2.2 c | 0.31 b | 32.2 ef | 1.6 f | 2.2 d | 0.26 c |
| Kateng’u | 20.6 ef | 1.32 hi | 1.5 d | 0.26 d | 14.4 h | 1.5 f | 1.7 e | 0.28 c |
| Marcia | 46.9 c | 3.2 c | 2.4 b | 0.4 a | 54.5 b | 3.4 b | 2.7 ab | 0.41 a |
| IESU24029 SH | 43.5 c | 2.9 d | 2.4 b | 0.26 d | 42.6 c | 2.3 e | 2.4 c | 0.26 c |
| Kauwi | 43.2 c | 2.6 e | 2.4 b | 0.31 b | 35.2 e | 2.1 ef | 2.5 b | 0.3 b |
| KARI Mtama-1 | 51.4 b | 3.7 a | 2.6 a | 0.41 a | 55.4 b | 3.6 a | 2.9 a | 0.4 a |
| Kiboko Local-2 | 53.9 a | 3.3 c | 2.7 a | 0.26 d | 44.34 c | 3.4 b | 2.9 a | 0.29 b |
| Rasta | 16.3 f | 0.8 i | 1.8 cd | 0.15 e | 17.7 g | 1.2 g | 1.9 e | 0.21 d |
| Makueni Local-2 | 47.7 c | 2.9 d | 2.5 a | 0.28 c | 61 a | 3.1 c | 3.1 a | 0.29 b |
| Serena | 48.6 c | 2.3 f | 2.5 a | 0.32 b | 40.5 d | 2 ef | 2.3 d | 0.28 b |
| Mugeta | 15.7 f | 0.5 j | 2.2 c | 0.08 f | 17.8 g | 0.2 h | 2.2 d | 0.03 e |
| Seredo | 36.9 d | 2 g | 1.9 cd | 0.33 b | 37.9 e | 2 ef | 2.1 de | 0.3 b |
| Kaguru | 28.4 e | 0.9 i | 1.8 cd | 0.2 de | 25.1 f | 0.75 gh | 2 de | 0.15 de |
| Dark red | 29 e | 1.5 hi | 1.9 cd | 0.2 de | 30.4 ef | 1.5 f | 2 de | 0.24 c |
| Mean | 36.99 | 1.89 | 2.2 | 0.27 | 36 | 2.1 | 2.4 | 0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koima, I.N.; Kilalo, D.C.; Orek, C.O.; Wagacha, J.M.; Nyaboga, E.N. Identification and Characterization of Colletotrichum Species Causing Sorghum Anthracnose in Kenya and Screening of Sorghum Germplasm for Resistance to Anthracnose. J. Fungi 2023, 9, 100. https://doi.org/10.3390/jof9010100
Koima IN, Kilalo DC, Orek CO, Wagacha JM, Nyaboga EN. Identification and Characterization of Colletotrichum Species Causing Sorghum Anthracnose in Kenya and Screening of Sorghum Germplasm for Resistance to Anthracnose. Journal of Fungi. 2023; 9(1):100. https://doi.org/10.3390/jof9010100
Chicago/Turabian StyleKoima, Irene Njeri, Dora Chao Kilalo, Charles O. Orek, John Maina Wagacha, and Evans N. Nyaboga. 2023. "Identification and Characterization of Colletotrichum Species Causing Sorghum Anthracnose in Kenya and Screening of Sorghum Germplasm for Resistance to Anthracnose" Journal of Fungi 9, no. 1: 100. https://doi.org/10.3390/jof9010100