
Pyricularia’s Capability of Infecting Different Grasses in Two Regions of Mexico
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolation and General Characterization
2.2. ITS Amplification and Sequencing Analysis
2.3. Haplotype Network Analysis
2.4. Genomic Fingerprinting
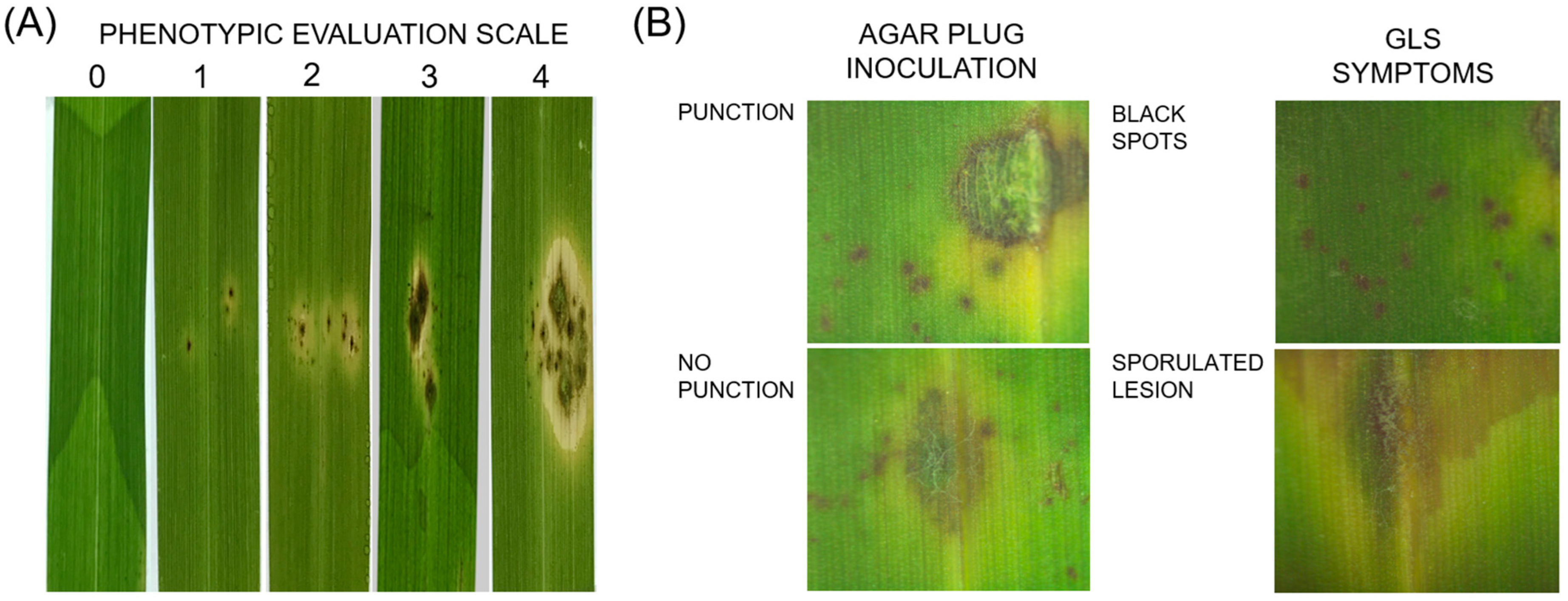
2.5. Infection Assay Using Spot Inoculation
2.6. Cross-Infectivity Assay
2.7. Statistical Analysis
3. Results
3.1. Isolation and General Characterization
3.2. ITS Sequence Analysis
3.3. Genomic Fingerprinting
3.4. Pathogenicity Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chung, H.; Goh, J.; Han, S.S.; Roh, J.H.; Kim, Y.; Heu, S.; Shim, H.K.; Jeong, D.G.; Kang, I.J.; Yang, J.W. Comparative pathogenicity and host ranges of Magnaporthe oryzae and related species. Plant Pathol. J. 2020, 36, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Franco, A.; Méndez-Rodríguez, A.; Garza-Cedillo, R. Tizón foliar del pasto buffel: Su presencia en Tamaulipas; México. Agric. Téc. Méx. 2007, 33, 285–295. [Google Scholar]
- Hernández-Arenas, M.G.; Barrios-Gómez, E.J.; Hernández-Aragón, L.; Berriozabal-Onofre, A. Plagas y Enfermedades del Arroz Cultivado en Morelos, 1st ed.; México, D.F., Ed.; Instituto Nacional de Investigaciones Forestales: Mexico City, Mexico, 2013.
- Ibarra, F.A.; Martin, M.; Moreno, S.; Ibarra, F.A.; Retes, R. Impacto económico asociado con los daños del tizón foliar en la producción de semilla del zacate Buffel en el centro de Sonora; México. Rev. Mex. Agronegocios 2019, 45, 313–323. [Google Scholar]
- Tredway, L.P.; Stevenson, K.L.; Burpee, L.L. Genetic structure of Magnaporthe grisea populations associated with St. Augustine grass and tall fescue in Georgia. Phytopathology 2005, 95, 463–471. [Google Scholar] [CrossRef]
- Cazal-Martínez, C.C.; Reyes-Caballero, Y.M.; Chávez, A.; Pérez-Estigarribia, P.E.; Rojas, A.; Arrua, A.; Moura-Mendes, J.; Lopez-Nicora, H.D.; Kohli, M.M. First report of a leaf blight caused by Pyricularia pennisetigena on Cenchrus echinatus in Paraguay. Plant Dis. 2021, 105, 3758. [Google Scholar] [CrossRef] [PubMed]
- Russell, T.R.; Kaminski, J.E.; Dernoeden, P.H. First report of Magnaporthe oryzae causing Gray Leaf Spot on tall fescue in Delaware and Maryland. Plant Dis. 2019, 103, 2966. [Google Scholar] [CrossRef]
- Farinas, C.; Villari, C.; Martin, D.; Taylor, N.J.; Wayne, E.C.; Peduto, H.F. Magnaporthe oryzae perennial ryegrass pathotype causes leaf spots and blight on japanese forest grass in ohio. Plant Dis. 2017, 101, 507. [Google Scholar] [CrossRef]
- Choi, J.; Park, S.Y.; Kim, B.R.; Roh, J.H.; Oh, I.S.; Han, S.S.; Lee, Y.H. Comparative analysis of pathogenicity and phylogenetic relationship in Magnaporthe grisea species complex. PLoS ONE 2013, 8, e57196. [Google Scholar] [CrossRef]
- Zhong, Z.; Norvienyeku, J.; Chen, M.; Bao, J.; Lin, L.; Chen, L.; Lin, Y.; Wu, X.; Cai, Z.; Zhang, Q.; et al. Directional selection from host plants is a major force driving host specificity in Magnaporthe species. Sci. Rep. 2016, 6, 25591. [Google Scholar] [CrossRef]
- Fang, X.; Snell, P.; Barbetti, M.J.; Lanoiselet, V. Rice varieties with resistance to multiple races of Magnaporthe oryzae offer opportunities to manage rice blast in Australia. Ann. Appl. Biol. 2017, 170, 160–169. [Google Scholar] [CrossRef]
- Mutiga, S.K.; Rotich, F.; Ganeshan, V.D.; Mwongera, D.T.; Mgonja, E.M.; Were, V.M.; Harvey, J.W.; Zhou, B.; Wasilwa, L.; Feng, C.; et al. Assessment of the virulence spectrum and its association with genetic diversity in Magnaporthe oryzae populations from sub-saharan Africa. Phytopathology 2017, 107, 852–863. [Google Scholar] [CrossRef]
- Imam, J.; Alam, S.; Mandal, N.P.; Maiti, D.; Variar, M.; Shukla, P. Molecular diversity and mating type distribution of the rice blast pathogen Magnaporthe oryzae in North-East and Eastern India. Indian J. Microbiol. 2014, 55, 108–113. [Google Scholar] [CrossRef]
- Le, L.T.; Adreit, H.; Ha, L.T.; Milazzo, J.; Lebrun, M.; Tharreau, D.; Pham, X.H.; Nguyen, H.T.; Fournier, E.; Hoang, G.T. Population structure of Pyricularia oryzae on rice in Vietnam reveals diversified populations with four pandemic and two endemic clusters. Fungal Genet. Biol. 2023, 166, 103794. [Google Scholar] [CrossRef]
- Consolo, V.F.; Cordo, C.A.; Salerno, G.L. DNA fingerprint and pathotype diversity of Pyricularia oryzae populations from Argentina. Australas. Plant Pathol. 2008, 37, 357–364. [Google Scholar] [CrossRef]
- Shirke, M.D.; Mahesh, H.B.; Gowda, M. Genome-wide comparison of Magnaporthe species reveals a host-specific pattern of secretory proteins and transposable elements. PLoS ONE 2016, 11, e0162458. [Google Scholar] [CrossRef]
- Jagadeesh, D.; Prasanna Kumar, M.K.; Chandrakanth, R.; Devaki, N.S. Molecular diversity of internal transcribed spacer among the monoconidial isolates of Magnaporthe oryzae isolated from rice in Southern Karnataka; India. J. Genet. Eng. Biotechnol. 2018, 16, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Mehta, Y.R.; Rosato, Y.B. ERIC- and REP-PCR amplify non-repetitive fragments from the genome of Drechslera avenae and Stemphylium solani. FEMS Microbiol. Lett. 2002, 211, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Abdollahzadeh, J.; Zolfaghari, S. Efficiency of rep-PCR fingerprinting as a useful technique for molecular typing of plant pathogenic fungal species: Botryosphaeriaceae species as a case study. FEMS Microbiol. Lett. 2014, 361, 144–157. [Google Scholar] [CrossRef]
- Sirisathaworn, T.; Srirat, T.; Longya, A.; Jantasuriyarat, C. Evaluation of mating type distribution and genetic diversity of three Magnaporthe oryzae avirulence genes; PWL-2; AVR-Pii and Avr-Piz-t; in Thailand rice blast isolates. Agric. Nat. Resour. 2017, 51, 7–14. [Google Scholar] [CrossRef]
- Zhang, N.; Zhao, S.; Shen, Q. A six-gene phylogeny reveals the evolution of mode of infection in the rice blast fungus and allied species. Mycologia 2011, 103, 1267–1276. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Suzuki, F.; Arai, M.; Yamaguchi, J. DNA fingerprinting of Pyricularia grisea by rep-PCR using a single primer based on the terminal inverted repeat from either of the transposable elements Pot2 and MGR586. J. Gen. Plant Pathol. 2006, 72, 314–317. [Google Scholar] [CrossRef]
- Pavel, A.B.; Vasile, C.I. PyElph—A software tool for gel images analysis and phylogenetics. BMC Bioinformatics 2012, 13, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Valent, B.; Lee, F.N. Determination of host responses to Magnaporthe grisea on detached rice leaves using a spot inoculation method. Plant Dis. 2003, 87, 129–133. [Google Scholar] [CrossRef]
- Takahashi, W.; Miura, Y.; Sasaki, T. A novel inoculation method for evaluation of grey leaf spot resistance in italian ryegrass. J. Plant Pathol. 2009, 91, 171–176. [Google Scholar] [CrossRef]
- Harp, T.L.; Correll, J.C. Recovery and characterization of spontaneous; selenate-resistant mutants of Magnaporthe grisea; the rice blast pathogen. Mycologia 1998, 90, 954–963. [Google Scholar] [CrossRef]
- Tagle, A.G.; Chuma, I.; Hisano, H.; Sato, K.; Tosa, Y. Genetic analysis of the resistance of barley to cryptic species of Pyricularia. J. Gen. Plant Pathol. 2016, 82, 302–306. [Google Scholar] [CrossRef]
- Genovesi, A.D.; Jessup, R.W.; Burson, B.L.; Engelke, M.C.; Chandra, A. Interspecific Hybrids between Pembagrass and St. Augustine grass. Crop Sci. 2017, 57, 2819–2827. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, X.; Garcia, S.; Leitch, A.R.; Kovařík, A. Intragenomic rDNA variation—The product of concerted evolution; mutation; or something in between? Heredity 2022, 131, 179–188. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Zhang, S.; Liu, X.Z.; Wen, H.A.; Wang, M. A simple method of genomic DNA extraction suitable for analysis of bulk fungal strains. Lett. Appl. Microbiol. 2010, 51, 114–118. [Google Scholar] [CrossRef]
- Hirata, K.; Kusaba, M.; Chuma, I.; Osue, J.; Nakayashiki, H.; Mayama, S.; Tosa, Y. Speciation in Pyricularia inferred from multilocus phylogenetic analysis. Mycol. Res. 2007, 111, 799–808. [Google Scholar] [CrossRef]
- Klaubauf, S.; Tharreau, D.; Fournier, E.; Groenewald, J.Z.; Crous, P.W.; de Vries, R.P.; Lebrun, M.H. Resolving the polyphyletic nature of Pyricularia (Pyriculariaceae). Stud. Mycol. 2014, 79, 85–120. [Google Scholar] [CrossRef]
- Murata, N.; Aoki, T.; Kusaba, M.; Tosa, Y.; Chuma, I. Various species of Pyricularia constitute a robust clade distinct from Magnaporthe salvinii and its relatives in Magnaporthaceae. J. Gen. Plant Pathol. 2014, 80, 66–72. [Google Scholar] [CrossRef]
- Chadha, S.; Sharma, M. Transposable elements as stress adaptive capacitors induce genomic instability in fungal pathogen Magnaporthe oryzae. PLoS ONE 2014, 9, e94415. [Google Scholar] [CrossRef]
- George, M.L.C.; Nelson, R.J.; Zeigler, R.S.; Leung, H. Rapid population analysis of Magnaporthe grisea by using rep-PCR and endogenous repetitive DNA sequences. Phytopathology 1998, 88, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Gladieux, P.; Ravel, S.; Rieux, A.; Cros-Arteil, S.; Adreit, H.; Milazzo, J.; Thierry, M.; Fournier, E.; Terauchi, R.; Tharreau, D. Coexistence of multiple endemic and pandemic lineages of the rice blast pathogen. mBio 2018, 9, e01806-17. [Google Scholar] [CrossRef]
- Gomez, L.L.B.; Tsai, I.J.; Chuma, I.; Tosa, Y.; Chen, Y.H.; Li, J.Y.; Li, M.Y.; Lu, M.Y.J.; Nakayashiki, H.; Li, W.H. Blast fungal genomes show frequent chromosomal changes; gene gains and losses; and effector gene turnover. Mol. Biol. Evol. 2019, 36, 1148–1161. [Google Scholar] [CrossRef] [PubMed]
- Motallebi, P.; Javan-Nikkhah, M.; Okhovvat, S.M. Characterization of Magnaporthe grisea populations associated with rice and weeds in Iran. Australas. Plant Pathol. 2013, 42, 693–700. [Google Scholar] [CrossRef]
- Viji, G.; Uddin, W. Distribution of mating type alleles and fertility status of Magnaporthe grisea causing gray leaf spot of perennial ryegrass and St. Augustine grass turf. Plant Dis. 2002, 86, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Asoufi, H.; Hameed, K.M.; Mahasneh, A. The cellulase and pectinase activities associated with the virulence of indigenous Sclerotinia sclerotiorum isolates in Jordan valley. Plant. Pathol. J. 2007, 23, 233–238. [Google Scholar] [CrossRef]
- Challagulla, V.; Bhattarai, S.; Midmore, D.J. In-vitro vs in-vivo inoculation: Screening for resistance of Australian rice genotypes against blast fungus. Rice Sci. 2015, 22, 132–137. [Google Scholar] [CrossRef]
- Ploetz, R.C. Diseases of Tropical Fruit Crops, 1st ed.; Ploetz Randy, C., Ed.; CABI Publishing: Brooker, FL, USA, 2003. [Google Scholar]
- Campos-Rivero, G.; Sánchez-Teyer, L.F.; De la Cruz-Arguijo, E.A.; Ramírez-González, M.S.; Larralde-Corona, C.P.; Narváez-Zapata, J.A. Bioprospecting for fungi with potential pathogenic activity on leaves of Agave tequilana Weber var. Azul. J. Phytopathol. 2019, 167, 283–294. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Isolate | Blast Identity (99%) | MAT1 Allele | Location | Host |
|---|---|---|---|---|---|
| MT785889 | SY1 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| MT785890 | SY2 a | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| MT785891 | SY4 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| MT785892 | SZ1 | P. oryzae | Mat1-1 | Zacatepec, Morelos | S. secundatum |
| MT785893 | SZ3 | P. oryzae | Mat1-1 | Zacatepec, Morelos | S. secundatum |
| OK185290 | SO1-1 | P. oryzae | Mat1-1 | Oacalco, Morelos | S. secundatum |
| OK185291 | SO1-2 | P. oryzae | Mat1-1 | Oacalco, Morelos | S. secundatum |
| OK185292 | SO1-3 | P. oryzae | Mat1-1 | Oacalco, Morelos | S. secundatum |
| OK185293 | SO2-1 | P. oryzae | Mat1-1 | Oacalco, Morelos | S. secundatum |
| OK185294 | SO2-2 | P. oryzae | Mat1-1 | Oacalco, Morelos | S. secundatum |
| OK185295 | SO2-3 | P. oryzae | Mat1-1 | Oacalco, Morelos | S. secundatum |
| OK185296 | SO3-2 | P. oryzae | Mat1-1 | Oacalco, Morelos | S. secundatum |
| OK185297 | SO3-3 | P. oryzae | Mat1-1 | Oacalco, Morelos | S. secundatum |
| OK185298 | SO3-4 | P. oryzae | Mat1-1 | Oacalco, Morelos | S. secundatum |
| OK185299 | SO3-5 | P. oryzae | Mat1-1 | Oacalco, Morelos | S. secundatum |
| OK185300 | SRc1-1 | P. oryzae | Mat1-1 | Reynosa, Tamaulipas | S. secundatum |
| OK185301 | SRc1-2 | P. oryzae | Mat1-1 | Reynosa, Tamaulipas | S. secundatum |
| OK185302 | SRc1-3 | P. oryzae | Mat1-1 | Reynosa, Tamaulipas | S. secundatum |
| OK185303 | SRc2-1 | P. oryzae | Mat1-1 | Reynosa, Tamaulipas | S. secundatum |
| OK185304 | SRc2-3 | P. oryzae | Mat1-1 | Reynosa, Tamaulipas | S. secundatum |
| OK185305 | SRc2-4 | P. oryzae | Mat1-1 | Reynosa, Tamaulipas | S. secundatum |
| MT785894 | SR1 a | P. oryzae | Mat1-2 | Reynosa, Tamaulipas | S. secundatum |
| OK185306 | SRf1 | P. oryzae | Mat1-2 | Reynosa, Tamaulipas | S. secundatum |
| OK185307 | SRf4 | P. oryzae | Mat1-2 | Reynosa, Tamaulipas | S. secundatum |
| OK185308 | SRf6 | P. oryzae | Mat1-2 | Reynosa, Tamaulipas | S. secundatum |
| OK185309 | SRf10 | P. oryzae | Mat1-2 | Reynosa, Tamaulipas | S. secundatum |
| OK185310 | SRf11 | P. oryzae | Mat1-2 | Reynosa, Tamaulipas | S. secundatum |
| OK185311 | SRf12 | P. oryzae | Mat1-2 | Reynosa, Tamaulipas | S. secundatum |
| OK185312 | SRf8 | P. oryzae | Mat1-1 | Reynosa, Tamaulipas | S. secundatum |
| OK185313 | SRf7 | P. oryzae | Mat1-1 | Reynosa, Tamaulipas | S. secundatum |
| OK185314 | SRf13 | P. oryzae | Mat1-1 | Reynosa, Tamaulipas | S. secundatum |
| OK185315 | SRf15 | P. oryzae | Mat1-1 | Reynosa, Tamaulipas | S. secundatum |
| OK185316 | SYb1-1 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185317 | SYb1-2 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185318 | SYb2-1 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185319 | SYc2-1 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185320 | SYc3-1 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185321 | SYc3-3 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185322 | SYc3-4 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185323 | SYc3-5 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185324 | SYe1-1 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185325 | SYe2-1 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185326 | SY1-1 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185327 | SY1-2 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185328 | SY2-2 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185329 | SY3-1 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185330 | SY3-2 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185331 | SYi1-1 | P. oryzae | Mat1-1 | Yautepec, Morelos | S. secundatum |
| OK185332 | CR1 a | P. pennisetigena | N.A. | Reynosa, Tamaulipas | C. ciliaris |
| OK185333 | CRc1 | P. pennisetigena | N.A. | Reynosa, Tamaulipas | C. ciliaris |
| OK185334 | CRc3 | P. pennisetigena | N.A. | Reynosa, Tamaulipas | C. ciliaris |
| OK185335 | CRc4 | P. pennisetigena | N.A. | Reynosa, Tamaulipas | C. ciliaris |
| OK185336 | CRf1 | P. pennisetigena | N.A. | Reynosa, Tamaulipas | C. ciliaris |
| OK185337 | CY1 a | P. pennisetigena | N.A. | Yautepec, Morelos | C. ciliaris |
| OK185338 | CY3 | P. pennisetigena | N.A. | Yautepec, Morelos | C. ciliaris |
| OK185339 | DY1 a | Pyricularia sp. | N.A. | Yautepec, Morelos | D. ciliaris |
| OK185340 | DY2 | Pyricularia sp. | N.A. | Yautepec, Morelos | D. ciliaris |
| Isolate | Location | Reaction Type Scale | Damaged Area (mm2) | ||
|---|---|---|---|---|---|
| Plug | Plug + Punction | Plug | Plug + Punction | ||
| SY1 | Yautepec, Morelos | 3 | 4 | 23.0 ± 3.6 b | 75.1 ± 9.5 a |
| SY2 | Yautepec, Morelos | 4 | 4 | 76.7 ± 6.2 a | 73.6 ± 13.0 a |
| SY4 | Yautepec, Morelos | 2 | 4 | 5.1 ± 2.3 c | 73.7 ± 6.2 a |
| SZ1 | Zacatepec, Morelos | 1 | 4 | 1.8 ± 1.8 c | 15.4 ±7.0 c |
| SZ3 | Zacatepec, Morelos | 1 | 4 | 1.6 ± 1.1 c | 17.6 ± 8.3 c |
| SR1 | Reynosa, Tamaulipas | 3 | 4 | 24.07 ± 4.8 b | 42.1 ± 5.5 b |
| Host | Representative Isolate | Leaf Lesions (Number) a | Damaged Area (mm2) | Damaged Area (%) | SES Scale b |
|---|---|---|---|---|---|
| Stenotaphrum secundatum | SY2 | 4.35 ± 1.1 | 57.57 ± 12.61 | 12.64 ± 2.58 | 6 |
| SR1 | 2 ± 0.79 | 9.82 ± 3.11 | 2.44 ± 0.87 | 4 | |
| CY1 | – | – | – | – | |
| CR1 | 2.1 ± 0.91 | 13.05 ± 5.04 | 2.92 ± 0.98 | 4 | |
| DY1 | – | – | – | – | |
| Cenchrus ciliaris | SY2 | – | – | – | – |
| SR1 | 1.3 ± 0.57 | 3.52 ± 1.58 | 0.43 ± 0.31 | 3 | |
| CY1 | 5.85 ± 2.21 | 80.45 ± 28.26 | 15.30 ± 3.21 | 6 | |
| CR1 | 2 ± 0.65 | 16.41 ± 4.75 | 2.79 ± 0.78 | 4 | |
| DY1 | – | – | – | 0 | |
| Digitaria ciliaris | SY2 | – | – | – | – |
| SR1 | – | – | – | – | |
| CY1 | – | – | – | – | |
| CR1 | – | – | – | – | |
| DY1 | 2.4 ± 0.9 | 20.59 ± 3.56 | 3.17 ± 0.57 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sequera-Grappin, I.; Ventura-Zapata, E.; De la Cruz-Arguijo, E.A.; Larralde-Corona, C.P.; Narváez-Zapata, J.A. Pyricularia’s Capability of Infecting Different Grasses in Two Regions of Mexico. J. Fungi 2023, 9, 1055. https://doi.org/10.3390/jof9111055
Sequera-Grappin I, Ventura-Zapata E, De la Cruz-Arguijo EA, Larralde-Corona CP, Narváez-Zapata JA. Pyricularia’s Capability of Infecting Different Grasses in Two Regions of Mexico. Journal of Fungi. 2023; 9(11):1055. https://doi.org/10.3390/jof9111055
Chicago/Turabian StyleSequera-Grappin, Ivan, Elsa Ventura-Zapata, Erika Alicia De la Cruz-Arguijo, Claudia Patricia Larralde-Corona, and Jose Alberto Narváez-Zapata. 2023. "Pyricularia’s Capability of Infecting Different Grasses in Two Regions of Mexico" Journal of Fungi 9, no. 11: 1055. https://doi.org/10.3390/jof9111055