
Agrobacterium tumefaciens-Mediated Transformation of the Aquatic Fungus Phialemonium inflatum FBCC-F1546
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains, Growth Media, Culture Conditions, and Hygromycin B Sensitivity
2.2. ATMT of P. inflatum
2.3. Genomic DNA Extraction
2.4. Southern Blot Analysis
2.5. PCR Amplification for Transformant Confirmation
2.6. eGFP Expression Observation in Transformants
2.7. Identification of T-DNA Insertion Sites via Inverse PCR
2.8. Mitotic Transformant Stability
2.9. Data Analysis
3. Results
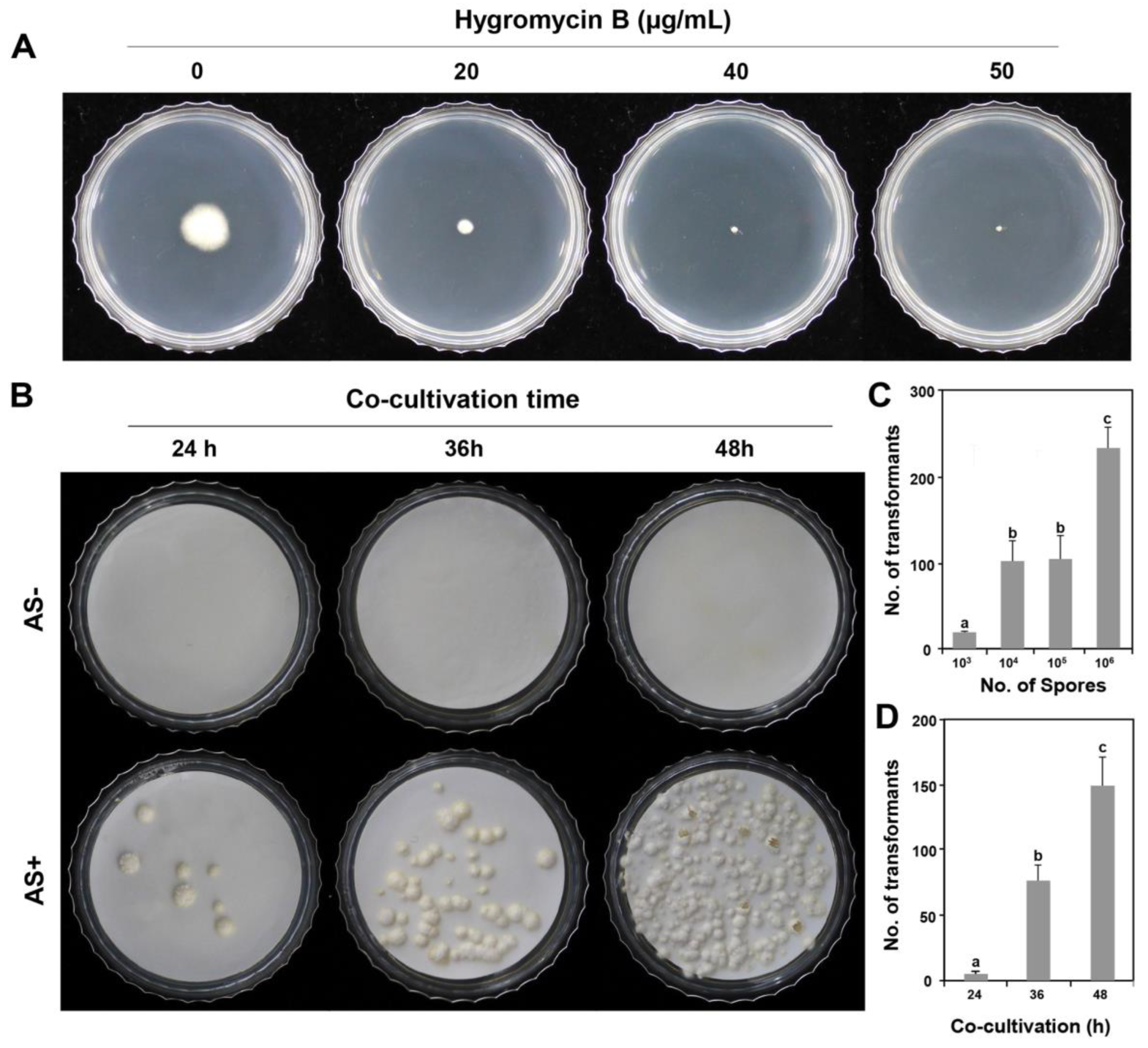
3.1. ATMT Establishment for P. inflatum
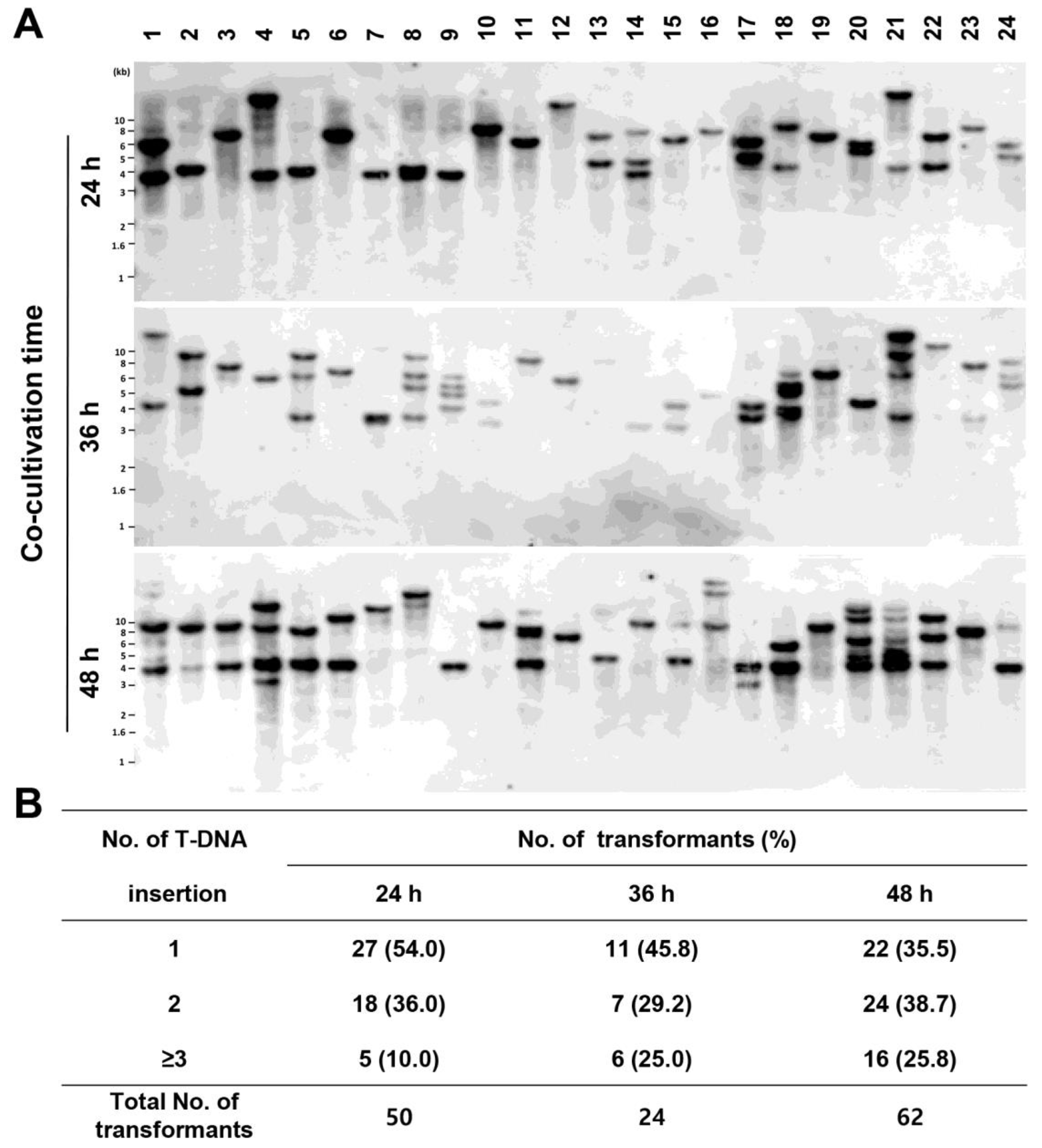
3.2. Analysis of T-DNA Insertion among the Generated Transformants
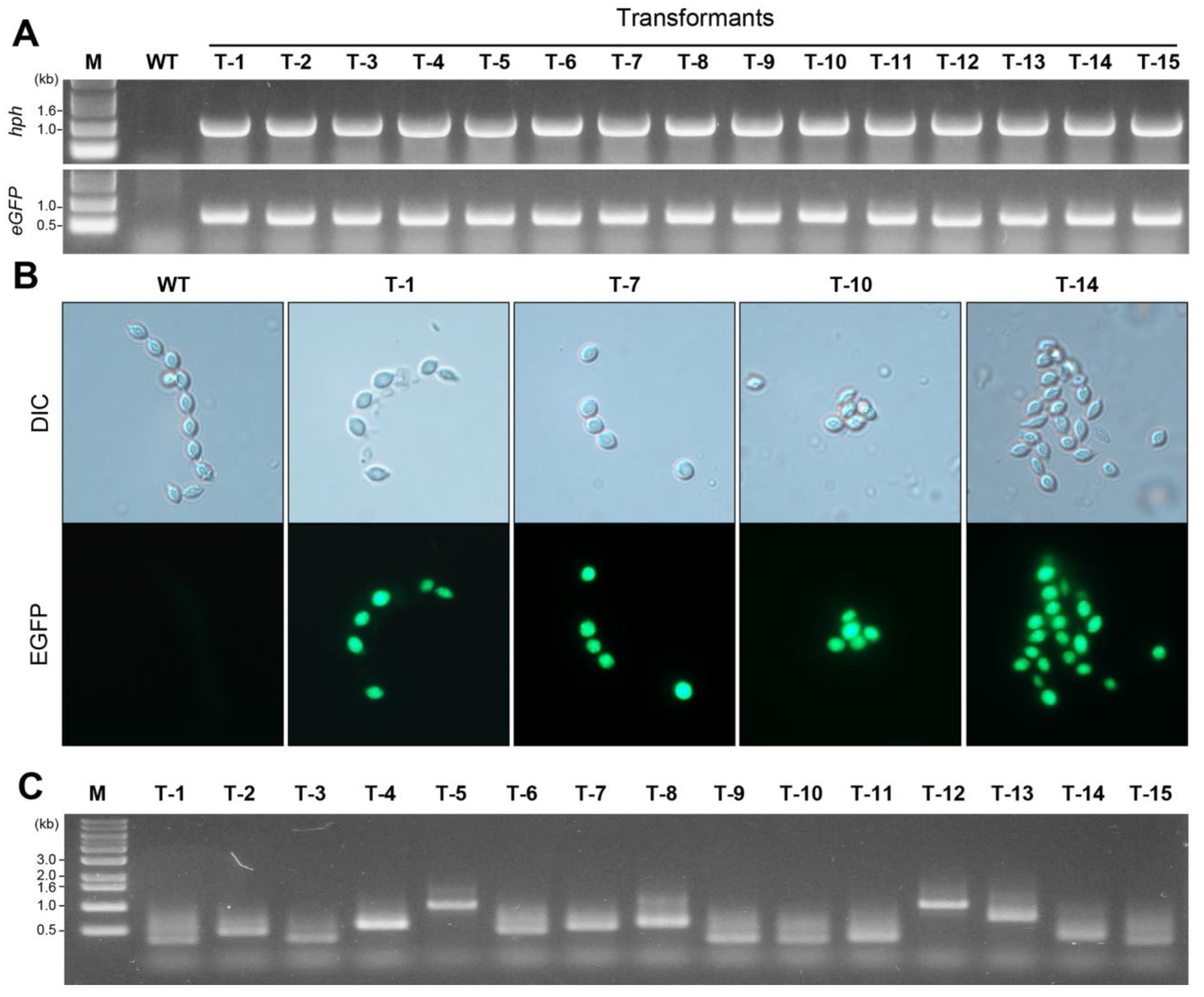
3.3. Confirmation of the Integration of the hph and eGFP Genes in the P. inflatum Genome
3.4. Identification of Genomic Sequences Flanking Inserted T-DNA in the Generated Transformants
3.5. Stability of Transformants in P. inflatum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perdomo, H.; Garcia, D.; Gene, J.; Cano, J.; Sutton, D.A.; Summerbell, R.; Guarro, J. Phialemoniopsi, a new genus of Sordariomycetes, and new species of Phialemonium and Lecythophora. Mycologia 2013, 105, 398–421. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Vega, L.J.; Grunseich, J.M.; Aguirre, N.M.; Valencia, C.U.; Sword, G.A.; Helms, A.M. A beneficial plant-associated fungus shifts the balance toward plant growth over resistance, increasing cucumber tolerance to root herbivory. Plants 2022, 11, 282. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.B.; Simoneau, P.; Barret, M.; Mitter, B.; Compant, S. Editorial special issue: The soil, the seed, the microbes and the plant. Plant Soil 2018, 422, 1–5. [Google Scholar] [CrossRef]
- Zhou, W.; Wheeler, T.A.; Starr, J.L.; Valencia, C.U.; Sword, G.A. A fungal endophyte defensive symbiosis affects plant-nematode interactons in cotton. Plant Soil 2018, 422, 251–266. [Google Scholar] [CrossRef]
- Kluczek-Turpeinen, B.; Tuomela, M.; Hatakka, A.; Hofrichter, M. Lignin degradation in a compost environment by the deuteromycete Paecilomyces inflatus. Appl. Microbiol. Biotechnol. 2003, 61, 374–379. [Google Scholar] [CrossRef]
- Bao, R.; Gao, N.; Lv, J.; Ji, C.; Liang, H.; Li, S.; Yu, C.; Wang, Z.; Lin, X. Enhancement of torularhodin production in Rhodosporidium toruloides by Agrobacterium tumefaciens-mediated transformation and culture condition optimization. J. Agric. Food Chem. 2019, 67, 1156–1164. [Google Scholar] [CrossRef]
- Chi, M.H.; Park, S.Y.; Kim, S.; Lee, Y.H. A novel pathogenicity gene is required in the rice blast fungus to suppress the basal defenses of the host. PLoS Pathog. 2009, 5, e1000401. [Google Scholar] [CrossRef]
- Debler, J.W.; Henares, B.M. Targeted disruption of scytalone dehydratase gene using Agrobacterium tumefaciens-mediated transformation leads to altered melanin production in Ascochyta lentis. J. Fungi 2020, 6, 314. [Google Scholar] [CrossRef]
- Jeon, J.; Park, S.Y.; Chi, M.H.; Choi, J.; Park, J.; Rho, H.S.; Kim, S.; Goh, J.; Yoo, S.; Park, J.Y.; et al. Genome-wide functional analysis of pathogenicity genes in the rice blast fungus. Nat. Genet. 2007, 39, 561–565. [Google Scholar] [CrossRef]
- Maruthachalam, K.; Klosterman, S.J.; Kang, S.; Hayes, R.J.; Subbarao, K.V. Identification of pathogenicity-related genes in the vascular wilt fungus Verticillium dahliae by Agrobacterium tumefaciens-mediated T-DNA insertional mutagenesis. Mol. Biotechnol. 2011, 49, 209–221. [Google Scholar] [CrossRef]
- Bundock, P.; den Dulk-Ras, A.; Beijersbergen, A.; Hooykaas, P.J. Trans-kingdom T-DNA transfer from Agrobacterium tumefaciens to Saccharomyces cerevisiae. EMBO J. 1995, 14, 3206–3214. [Google Scholar] [CrossRef] [PubMed]
- Bundock, P.; Hooykaas, P.J. Integration of Agrobacterium tumefaciens T-DNA in the Saccharomyces cerevisiae genome by illegitimate recombination. Proc. Natl. Acad. Sci. USA 1996, 93, 15272–15275. [Google Scholar] [CrossRef]
- Piers, K.L.; Heath, J.D.; Liang, X.; Stephens, K.M.; Nester, E.W. Agrobacterium tumefaciens-mediated transformation of yeast. Proc. Natl. Acad. Sci. USA 1996, 93, 1613–1618. [Google Scholar] [CrossRef] [PubMed]
- Combier, J.P.; Melayah, D.; Raffier, C.; Gay, G.; Marmeisse, R. Agrobacterium tumefaciens-mediated transformation as a tool for insertional mutagenesis in the symbiotic ectomycorrhizal fungus Hebeloma cylindrosporum. FEMS Microbiol. Lett. 2003, 220, 141–148. [Google Scholar] [CrossRef] [PubMed]
- de Groot, M.J.; Bundock, P.; Hooykaas, P.J.; Beijersbergen, A.G. Agrobacterium tumefaciens-mediated transformation of filamentous fungi. Nat. Biotechnol. 1998, 16, 839–842. [Google Scholar] [CrossRef]
- Gouka, R.J.; Gerk, C.; Hooykaas, P.J.; Bundock, P.; Musters, W.; Verrips, C.T.; de Groot, M.J. Transformation of Aspergillus awamori by Agrobacterium tumefaciens-mediated homologous recombination. Nat. Biotechnol. 1999, 17, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Khang, C.; Park, S.; Lee, Y.; Kang, S. A dual selection based, targeted gene replacement tool for Magnaporthe grisea and Fusarium oxysporum. Fungal Genet. Biol. 2005, 42, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Malonek, S.; Meinhardt, F. Agrobacterium tumefaciens-mediated genetic transformation of the phytopathogenic ascomycete Calonectria morganii. Curr. Genet. 2001, 40, 152–155. [Google Scholar] [CrossRef]
- Michielse, C.B.; Arentshorst, M.; Ram, A.F.; van den Hondel, C.A. Agrobacterium-mediated transformation leads to improved gene replacement efficiency in Aspergillus awamori. Fungal Genet. Biol. 2005, 42, 9–19. [Google Scholar] [CrossRef]
- Michielse, C.B.; Hooykaas, P.J.; van den Hondel, C.A.; Ram, A.F. Agrobacterium-mediated transformation as a tool for functional genomics in fungi. Curr. Genet. 2005, 48, 1–17. [Google Scholar] [CrossRef]
- Mikosch, T.S.; Lavrijssen, B.; Sonnenberg, A.S.; van Griensven, L.J. Transformation of the cultivated mushroom Agaricus bisporus (Lange) using T-DNA from Agrobacterium tumefaciens. Curr. Genet. 2001, 39, 35–39. [Google Scholar] [CrossRef]
- Molina, M.C.; Crespo, A. Comparison of development of axenic cultures of five species of lichen-forming fungi. Mycol. Res. 2000, 104, 595–602. [Google Scholar] [CrossRef]
- Rho, H.S.; Kang, S.; Lee, Y.H. Agrobacterium tumefaciens-mediated transformation of the plant pathogenic fungus, Magnaporthe grisea. Mol. Cells 2001, 12, 407–411. [Google Scholar]
- Mullins, E.; Romaine, C.P.; Chen, X.; Geiser, D.; Raina, R.; Kang, S. Agrobacterium tumefaciens-mediated transformation of Fusarium oxysporum: An efficient tool for insertional mutagenesis and gene transfer. Phytopathology 2001, 91, 173–180. [Google Scholar] [CrossRef]
- Frandsen, R.J.N. A guide to binary vectors and strategies for targeted genome modification in fungi using Agrobacterium tumefaciens-mediated transformation. J. Microbiol. Methods 2011, 87, 247–262. [Google Scholar] [CrossRef] [PubMed]
- Idnurm, A.; Bailey, A.M.; Cairns, T.C.; Elliott, C.E.; Foster, G.D.; Ianiri, G.; Jeon, J. A silver bullet in a golden age of functional genomics: The impact of Agrobacterium-mediated transformation of fungi. Fungal Biol. Biotechnol. 2017, 4, 6. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.H.; Kim, J.A.; Kang, S.; Choi, E.D.; Kim, Y.; Lee, Y.; Jeon, M.J.; Yu, N.H.; Park, A.R.; Kim, J.C.; et al. Optimization of Agrobacterium tumefaciens-mediated transformation of Xylaria grammica EL000614, an endolichenic fungus producing grammicin. Mycobiology 2021, 49, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Jeong, M.H.; Wang, H.Y.; Kim, J.A.; Yu, N.H.; Kim, S.; Cheong, Y.H.; Kang, S.; Lee, Y.H.; Hur, J.S. Agrobacterium tumefaciens-mediated transformation of the lichen fungus, Umbilicaria muehlenbergii. PLoS ONE 2013, 8, e83896. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhu, H.; Chen, Y.; Zheng, L.; Chen, L.; Ma, A. Agrobacterium tumefaciens-mediated transformation of the king tuber medicinal mushroom Lentinus tuber-regium (Agaricomycetes). Int. J. Med. Mushrooms 2018, 20, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.K.; Kuhad, R.C. Genetic transformation of lignin degrading fungi facilitated by Agrobacterium tumefaciens. BMC Biotechnol. 2010, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Daboussi, M.J.; Djeballi, A.; Gerlinger, C.; Blaiseau, P.L.; Bouvier, I.; Cassan, M.; Lebrun, M.H.; Parisot, S.; Brygoo, Y. Transformation of seven species of filamentous fungi using the nitrate reductase gene of Aspergillus nidulans. Curr. Genet. 1989, 15, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Ochman, H.; Gerber, A.S.; Hartl, D.L. Genetic applications of an inverse polymerase chain reaction. Genetics 1988, 120, 621–623. [Google Scholar] [CrossRef] [PubMed]
- Mora-Lugo, R.; Zimmermann, J.; Rizk, A.M.; Fernandez-Lahore, M. Development of a transformation system for Aspergillus sojae based on the Agrobacterium tumefaciens-mediated approach. BMC Microbiol. 2014, 14, 247. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Diaz, B. Strategies for the transformation of filamentous fungi. J. Appl. Microbiol. 2002, 92, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Ha, B.S.; Ro, H.S. Current technologies and related issues for mushroom transformation. Mycobiology 2015, 43, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Guo, A.; Lu, Z.; Tan, S.; Wang, J.; Gao, J.; Zhang, S.; Huang, X.; Zheng, J.; Xi, J.; et al. Agrobacterium tumefaciens-mediated transformation of a hevein-like gene into asparagus leads to stem wilt resistance. PLoS ONE 2019, 14, e0223331. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.H.; Wang, S.T. Agrobacterium tumefaciens-mediated transformation method for Fusarium oxysporum. Methods Mol. Biol. 2022, 2391, 63–73. [Google Scholar] [CrossRef]
- D’Spain, S.; Andrade, P.I.; Brockman, N.E.; Fu, J.; Wickes, B.L. Agrobacterium tumefaciens-mediated transformation of Candida glabrata. J. Fungi 2022, 8, 596. [Google Scholar] [CrossRef]
- Li, D.; Wei, X.; Liu, T.; Liu, C.; Chen, W.; Xuan, Y.H.; Gao, L. Establishment of an Agrobacterium tumefaciens-mediated transformation system for Tilletia foetida. J. Microbiol. Methods 2020, 169, 105810. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, X.; Zhao, Y.; Yang, Y.; Liu, Z. An effective method for Agrobacterium tumefaciens-mediated transformation of Jatropha curcas L. using cotyledon explants. Bioengineered 2020, 11, 1146–1158. [Google Scholar] [CrossRef]
- Choi, J.; Park, J.; Jeon, J.; Chi, M.H.; Goh, J.; Yoo, S.Y.; Jung, K.; Kim, H.; Park, S.Y.; Rho, H.S.; et al. Genome-wide analysis of T-DNA integration into the chromosomes of Magnaporthe oryzae. Mol. Microbiol. 2007, 66, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Montoya, M.R.A.; Massa, G.A.; Colabelli, M.N.; Ridao, A.D.C. Efficient Agrobacterium tumefaciens-mediated transformation system of Diaporthe caulivora. J. Microbiol. Methods 2021, 184, 106197. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Hou, H.; Huang, T.; Zhou, Z.; Tu, H.; Wang, L. Agrobacterium tumefaciens-mediated transformation of Coniella granati. J. Microbiol. Methods 2021, 182, 106149. [Google Scholar] [CrossRef] [PubMed]
- Khang, C.H.; Berruyer, R.; Giraldo, M.C.; Kankanala, P.; Park, S.Y.; Czymmek, K.; Kang, S.; Valent, B. Translocation of Magnaporthe oryzae effectors into rice cells and their subsequent cell-to-cell movement. Plant Cell 2010, 22, 1388–1403. [Google Scholar] [CrossRef]
- Yi, M.; Chi, M.H.; Khang, C.H.; Park, S.Y.; Kang, S.; Valent, B.; Lee, Y.H. The ER chaperone LHS1 is involved in asexual development and rice infection by the blast fungus Magnaporthe oryzae. Plant Cell 2009, 21, 681–695. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Transformants | T-DNA Copy Number | Sequences Flanking the Inserted T-DNA (5′-3′) a |
|---|---|---|
| T-01 | 1 | GTTTAAACTATCAGTGTTTGA b cggaacggatcatcaggctcgattt c |
| T-02 | 1 | GTTTAAACTATCAGTGTTTGAgtccttcatcctggactttcttcgatttta |
| T-03 | 1 | GTTTAAACTATCAGTGTTTGAtaaacaaattgacgcttaaaccacttaa |
| T-04 | 1 | GTTTAAACTATCAGTGTTTGAaggttctgcgcccaccatgggcaggc |
| T-05 | 1 | GTTTAAACTATCAGTGTTTGAcaatttgctggtagtcctatcccgtccc |
| T-06 | 1 | GTTTAAACTATCAGTGTTTGAcccagccaggcttttccccaggatacc |
| T-07 | 1 | GTTTAAACTATCAGTGTTTGAtgggtggacctcgattttacgcacatat |
| T-08 | 1 | GTTTAAACTATCAGTGTTTGAgagatcgattttacgcacatatgcgcat |
| T-09 | 1 | GTTTAAACTATCAGTGTTTGAcgaggatgaggaagatgaaggcgac |
| T-10 | 1 | GTTTAAACTATCAGTGTTTGAcccatcctctccagtaccaaactacctc |
| T-11 | 1 | GTTTAAACTATCAGTGTTTGAggtgtgtgtgtgtgagtgtgtctgggc |
| T-12 | 1 | GTTTAAACTATCAGTGTTTGAcctacctgcctgtgtaccggagctcac |
| T-13 | 1 | GTTTAAACTATCAGTGTTTGAcctcctgtctaattatccttgcttcgtttc |
| T-14 | 1 | GTTTAAACTATCAGTGTTTGAtttttgttggtcgattttacgcacatatg |
| T-15 | 1 | GTTTAAACTATCAGTGTTTGAcgttccgattcggaaaggaagaggg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, J.; Kim, Y.; Kim, S.; Jeong, H.; Park, J.; Jeong, M.-H.; Park, S.; Jo, M.; An, S.; Park, J.; et al. Agrobacterium tumefaciens-Mediated Transformation of the Aquatic Fungus Phialemonium inflatum FBCC-F1546. J. Fungi 2023, 9, 1158. https://doi.org/10.3390/jof9121158
Yoon J, Kim Y, Kim S, Jeong H, Park J, Jeong M-H, Park S, Jo M, An S, Park J, et al. Agrobacterium tumefaciens-Mediated Transformation of the Aquatic Fungus Phialemonium inflatum FBCC-F1546. Journal of Fungi. 2023; 9(12):1158. https://doi.org/10.3390/jof9121158
Chicago/Turabian StyleYoon, Jonghan, Youngjun Kim, Seoyeon Kim, Haejun Jeong, Jiyoon Park, Min-Hye Jeong, Sangkyu Park, Miju Jo, Sunmin An, Jiwon Park, and et al. 2023. "Agrobacterium tumefaciens-Mediated Transformation of the Aquatic Fungus Phialemonium inflatum FBCC-F1546" Journal of Fungi 9, no. 12: 1158. https://doi.org/10.3390/jof9121158