
Six Additions to the Genus Periconia (Dothideomycetes: Periconiaceae) from Graminaceous Plants in China



School of Life Science and Technology, Center for Informational Biology, University of Electronic Science and Technology of China, Chengdu 611731, China
*
Author to whom correspondence should be addressed.
J. Fungi 2023, 9(3), 300; https://doi.org/10.3390/jof9030300
Submission received: 5 February 2023
/
Revised: 20 February 2023
/
Accepted: 23 February 2023
/
Published: 25 February 2023
(This article belongs to the Special Issue Recent Advances in Taxonomy, Phylogeny and Evolution of Fungi)
Abstract
:Periconia is a polyphyletic and asexual morphic genus within the family Periconiaceae (Pleosporales). The genus is characterized by a pale to dark brown stipe with an apical conidial head and ellipsoidal to oblong conidia. Species of Periconia are widely distributed throughout the world in various hosts, while most species are isolated from graminaceous plants. During our investigations of microfungal in Sichuan Province, China, 26 Periconia isolates were collected from a wide variety of graminaceous plants. These isolates corresponded to 11 species based on the examination of morphology and multi-locus phylogenetic analysis (SSU, ITS, LSU, TEF1, RPB2). This includes six new species (P. chengduensis, P. cynodontis, P. festucae, P. imperatae, P. penniseti, and P. spodiopogonis) and five new records (P. byssoides, P. chimonanthi, P. cookie, P. pseudobyssoides, and P. verrucosa). A comprehensive description and illustrations of the new species are provided and discussed with comparable taxa. These discoveries expand our knowledge of the species diversity of Periconia taxa in graminaceous plants in China.
1. Introduction
The genus Periconia (Periconiaceae, Pleosporales) was introduced by H.J. Tode, with P. lichenoides as the type species [1]. Most Periconia species are mainly known from their asexual morphs, characterized by conidiophores that are macronematous, mononematous, branched or unbranched, and pale to dark brown [2,3], while conidiogenous cells are discrete on the terminal or intercalary of the stipe and are monoblastic to polyblastic [2,4]. Conidia of Periconia species are globose to ellipsoidal, catenate or solitary, smooth or verruculose, and pale brown to brown [2,4,5,6]. Only five species have been reported to have sexual morphs, viz., P. didymosporum, P. homothallica, P. igniaria, P. prolifica, and P. pseudodigitata [7,8]. The sexual morph is characterized by scattered or grouped, globose ascomata with a central ostiole, hyaline periphyses, 8-spored asci, and broadly fusiform, 1-septate, hyaline and smooth ascospores with an entire sheath [7,8,9,10]. Periconia species are widely distributed and usually found in terrestrial habitats and rarely in aquatic habitats [11,12]. The genus comprises many saprophytes and endophytes, while few are plant pathogens, mainly causing diseases in graminaceous plants [2,4,13,14,15]. For example, P. circinata causes the blackening and rotting of wheat roots and stem bases (milo disease), and P. macrospinosa causes leaf necrosis in the pointed gourd (Cucurbitaceae) [16,17]. Gunasekaran et al. [18] reported that P. atra causes human corneal ulcers. Several species, such as P. atropurpurea and P. siamensis, are known to produce aromatic compounds and macrolide compounds with tremendous pharmacological activities [19,20].
Periconia is a polyphyletic genus in the family Periconiaceae [21]. The members of this genus were previously classified under Massarinaceae. Based on phylogenetic analysis, Tanaka et al. [7] showed that Periconiaceae is a sister clade distinct from Massarinaceae [22]. Phukhamsakda et al. [23] showed that Periconiaceae and Massarinaceae diverged in the late Cretaceous period (around 70 million years ago). The family Periconiaceae includes four genera: Bambusistroma, Flavomyces, Noosia, and Periconia. The genus Periconia has 211 epithets in Index Fungorum (http://www.indexfungorum.org/; accessed on 20 January 2023). Among these, 29 species have been transferred to other genera [24,25,26,27,28,29,30], and 7 species have been synonymized under other Periconia species [2,31,32,33]. In the previous three years, nine Periconia species have been introduced [6,8,34,35,36,37]. There are 124 accepted Periconia species in Species Fungorum, but only 37 species have sequence data.
In this study, 26 Periconia isolates were obtained from several collection sites from July to October 2021. Based on morphological studies and multi-locus phylogenetic analysis, these isolates were assigned to 11 species, including 6 new species and 5 new records. This study aims to describe these new taxa with detailed descriptions and illustrations and to broaden our understanding of the diversity of periconia-like taxa.
2. Materials and Methods
2.1. Sample Collection, Morphological Examination, Isolation, and Preservation
A survey on a fungal diversity of hyphomycetes fungi on graminaceous plants in Sichuan Province, China, was conducted from July to October 2021 at five natural sites in Sichuan Province (Huilonggou, Pengzhou City; Guoxue Park, Chengdu; Longchi National Forest Park, Chengdu; Baiyungou, Chunzhou City; Xiqiang Valley, Wenchuan County, Ngawa Tibetan and Qiang Autonomous Prefecture). These specimens were stored in paper envelopes and returned to the laboratory for examination. The morphological observation was carried out from material on natural substrates using a Motic SMZ 168 series stereomicroscope. The mycelia were placed in sterile water for micromorphological observation, and the fungal microstructures were photographed using the DS-Fi3 camera fitted with a Nikon Eclipse Ni-U microscope. All measurements were conducted using Nikon NIS-Elements D 5.21 software, and the photos were processed using Adobe Photoshop version 21.0. Pure cultures were obtained by single spore isolation according to the method described by Senanayake et al. [38]. The germinated conidia were then transferred individually to potato dextrose agar (PDA) plates and incubated in the dark at 25 °C. Culture characteristics were examined and recorded after one week and later at regular intervals.
The specimens were deposited in the Herbarium of Cryptogams Kunming Institute of Botany Academia Sinica (HKAS), Kunming, China, or the Herbarium of the University of Electronic Science and Technology (HUEST), Chengdu, China. The living cultures were deposited in the China General Microbiological Culture Collection Center (CGMCC), Beijing, China, and the University of Electronic Science and Technology Culture Collection (UESTCC), Chengdu, China. The taxonomic descriptions of the new taxa have been deposited with MycoBank (https://www.mycobank.org/; accessed on 1 February 2023).
2.2. DNA Extraction, PCR Amplification, and Sequencing
Fungal genomic DNA was extracted from mycelia using the TreliefTM Plant Genomic DNA Kit (TSINGKE Biotech, Shanghai, China) according to the manufacturer’s protocol. The DNA was stored at −20 °C for long-term storage. The five loci, which include the nuclear ribosomal internal transcribed spacer (ITS: ITS1-5.8S-ITS2), the partial nuclear ribosomal small subunit rRNA (SSU), the partial nuclear ribosomal large subunit rRNA (LSU), the partial translation elongation factor 1-alpha (TEF1), and the partial second-largest subunit of RNA polymerase II (RPB2), were amplified by polymerase chain reaction (PCR). The corresponding primer pairs and PCR conditions are listed in Table 1. The final PCR reaction system was 25 µL, containing 12.5 μL PCR Master Mix (Sangon Biotech, Shanghai, China), 8.5 µL of double-distilled water (ddH2O), 1 μL each of forward and reverse primers, and 2 μL DNA template. The PCR products were visualized by electrophoresis in 1% agarose gels. Sanger sequencing was conducted by Tsingke Biological Technology (Beijing, China).
2.3. Phylogenetic Analyses
The raw sequencing fragments of corresponding Sanger sequencing chromatograms were manually edited, trimmed, and assembled into consensus sequences using SeqMan Pro version 11.1.0 (DNASTAR, Inc. Madison, WI, USA). Barcode sequences of Periconia species currently available in GenBank and the outgroup taxon Massarina cisti (CBS 266.62) were downloaded from the NCBI nucleotide database using an in-house python script.
The multiple sequence alignment was conducted using MAFFT version 7.310 [46] with options “--maxiterate 1000 --genafpair --adjustdirectionaccurately”, and the alignment results were further trimmed using trimAl version 1.4 [47] with the option “-gapthreshold 0.5”, which only allows 50% of taxa with a gap in each site. The best-fit nucleotide substitution models for each alignment dataset were selected using PartitionFinder version 2.1.1 [48] under the Akaike Information Criterion (AIC).
Maximum likelihood (ML) and Bayesian analysis (BI) were conducted based on the individual and combined datasets. ML phylogenetic trees were obtained using the IQ-TREE version 2.0.3 [49], and the topology was evaluated using 1000 ultrafast bootstrap replicates. The BI was conducted using parallel MrBayes version 3.2.7a [50]. Two different runs with 20 million generations and four chains were executed, and the initial 25% of sample trees were treated as burn-in. Tracer version 1.7.1 [51] was used to confirm that the MCMC runs reached convergence with all ESS values above 200. Then, the ML tree was annotated by TreeAnnotator version 2.6.6 implemented in BEAST version 2.6.6 [52] based on MrBayes MCMC trees with no discard of burn-in and no posterior probability limit. The tree was visualized using ggtree [53] and edited in Adobe Illustrator version 20.0.0.
3. Results
3.1. Molecular Phylogeny
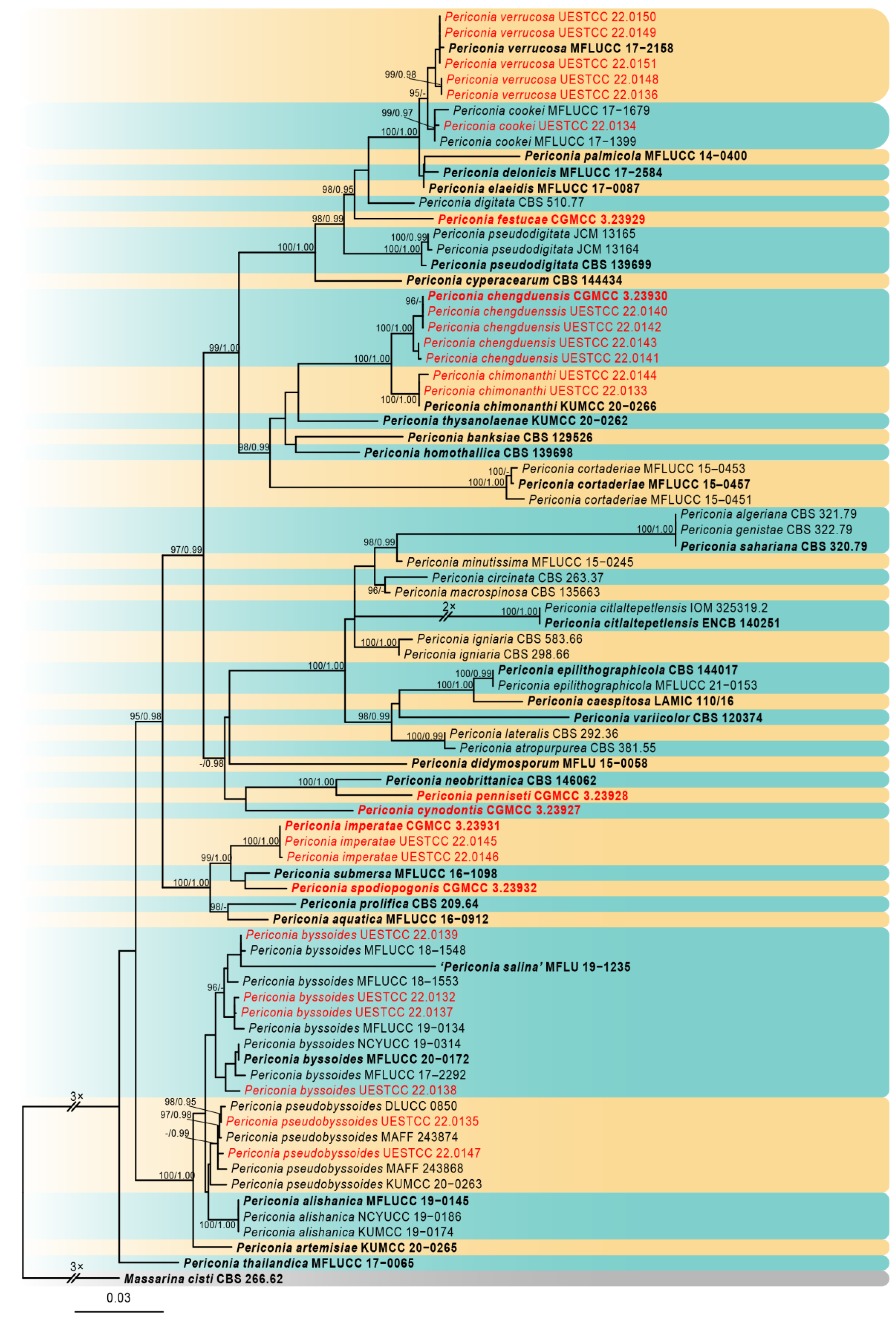
The newly generated sequences were deposited in GenBank, and the accession numbers were listed in Table 2. The combined dataset included five loci, SSU, ITS, LSU, TEF1, and RPB2, from 82 isolates of Periconia with Massarina cisti (CBS 266.62) as the outgroup taxon. The concatenated alignment comprises 4320 characters (SSU: 1–1001; ITS: 1002–1552; LSU: 1553–2411; TEF1: 2412–3309; RPB2: 3310–4320), including gaps, consisting of 1389 distinct patterns, 804 parsimony-informative, 330 singleton sites, and 3186 constant sites. Five single-locus datasets, SSU, ITS, LSU, TEF1, and RPB2, contained 30, 240, 91, 123, and 320 parsimony-informative sites, respectively. The best-fit evolution models were GTR+I+G for the SSU, ITS, LSU, TEF1, and RPB2.
The best-scoring ML consensus tree (lnL = −20,240.300) with ultrafast bootstrap values from ML analyses and posterior probabilities from MrBayes analysis at the node is shown in Figure 1. Phylogenetic analyses showed that our newly collected 26 isolates clustered into 11 clades and can be recognized as 5 known species (P. byssoides, P. chimonanthi, P. cookie, P. pseudobyssoides, and P. verrucosa) and six new species (P. chengduensis, P. cynodontis, P. festucae, P. imperatae, P. penniseti, and P. spodiopogonis).
3.2. Taxonomy
Periconia byssoides Pers., Syn. meth. fung. (Göttingen) 2: 686 (1801) Figure 2.
MycoBank: MB 144538.
Saprobic on dead culms of Poaceae. Asexual morph: Colonies on the natural substrate numerous, effuse, brown to dark brown, hairy. Conidiophores 240–410 μm long ( = 354, n = 15), 12–21 μm wide ( = 17 μm, n = 15), macronematous, mononematous, straight or slightly flexuous, unbranched, solitary, rarely 1–2 together on stroma, brown to dark brown, 2–4-septate (majority 3-septate), smooth to minutely verruculose, thick-walled. Conidiogenous cells polyblastic, pale brown to brown, subglobose, smooth or verruculose. Conidia 11.5–18.5 × 12–18 μm ( = 15 × 14.5 μm, n = 40), solitary or catenate, globose, brown, aseptate, echinulate, or verruculose. Sexual morph: Undetermined.
Materials examined: China, Sichuan Province, Ngawa Tibetan and Qiang Autonomous Prefecture, Wenchuan County, Xiqiang Valley, 31°29′27″ N, 103°37′1″ E, elevation 1500 m, on dead culms of Imperata cylindrica (Poaceae), 20 October 2021, ZH Lu w281_1 (HUEST 22.0133), living culture UESTCC 22.0132; ibid., w282 (HUEST 22.0140), living culture UESTCC 22.0139; Chongzhou City, Baiyungou, 30°47′35″ N, 103°23′49″ E, elevation 990 m, on dead culms of Imperata cylindrica (Poaceae), 27 September 2021, ZH Lu Lu44 (HUEST 22.0138), living culture UESTCC 22.0137; ibid., Lu50 (HUEST 22.0139), living culture UESTCC 22.0138.
Culture characteristics: Colony on PDA reaching 50 mm diam after one week in an incubator under dark conditions at 20 °C, circular, cottony, hairy at the margin, white; pale yellow at the margin and mud yellow at the middle in reverse.
Notes: The phylogenetic tree showed that our four isolates (UESTCC 22.0132, UESTCC 22.0137, UESTCC 22.0138, UESTCC 22.0139) grouped with other P. byssoides isolates, including the type (MFLUCC 20–0172). Our collections (HUEST 22.0133, HUEST 22.0138, HUEST 22.0139, HUEST 22.0140) share similar morphological characteristics in shape and color of conidiophores and conidia with the type of P. byssoides. Therefore, we identified our new isolates as P. byssoides based on the overlapping morphological characteristics and the multi-locus phylogenetic tree, and one collection (HUEST 22.0133) is a new host record from Imperata cylindrica.
Periconia chengduensis Z.H. Lu, P.W. Su, and Maharachch., sp. nov. Figure 3.
MycoBank: MB 847458
Etymology: Name refers to Chengdu, the city where the fungus was collected.
Saprobic on dead culms of Poaceae. Asexual morph: Colonies on the natural substrate numerous, effuse, dark brown to black, hairy. Conidiophores 240–370 μm long ( = 292, n = 15), 10–13 μm wide ( = 12 μm, n = 15), macronematous, mononematous, straight or slightly flexuous, branched, solitary, rarely 1–2 together on stroma, dark brown to black, septate, smooth to minutely verruculose, thick-walled. Conidiogenous cells polyblastic, pale brown to brown, terminal, integrated, subglobose, and smooth to verruculose. Conidia 5–8.5 × 5–8 μm ( = 7.2 × 7 μm, n = 40), solitary or catenate, oval to globose, brown to dark brown, aseptate, echinulate or verruculose. Sexual morph: Undetermined.
Materials examined: China, Sichuan Province, Chengdu City, Longchi National Forest Park, 31°06′15″ N, 103°33′32″ E, elevation 2000 m, on dead culms of Pennisetum purpureum (Poaceae), 19 September 2021, ZH Lu w31_1 (HKAS 126514, holotype), ex-type culture CGMCC 3.23930 = UESTCC 22.0126; ibid., on dead culms of Miscanthus sinensis (Poaceae), ZH Lu Sarah57 (HUEST 22.0141), living culture UESTCC 22.0140; Chongzhou City, Baiyungou, 30°47′35″ N, 103°23′49″ E, elevation 990 m, on dead culms of Imperata cylindrica (Poaceae), 27 September 2021, ZH Lu Lu64 (HUEST 22.0142), living culture UESTCC 22.0141; Ngawa Tibetan and Qiang Autonomous Prefecture, Wenchuan County, Xiqiang Valley, 31°29′27″ N, 103°37′1″ E, elevation 1500 m, on dead culms of Phragmites australis (Poaceae), 20 October 2021, ZH Lu w168_1 (HUEST 22.0143), living culture UESTCC 22.0142; ibid. on dead culms of Neyraudia reynaudiana (Poaceae), ZH Lu w234_3 (HUEST 22.0144), living culture UESTCC 22.0143.
Culture characteristics: Colony on PDA reaching 37 mm diam after 16 days in an incubator under dark conditions at 20 °C, circular, cottony, hairy at the margin, white; reverse: white at the margin and pale yellow at the middle.
Notes: The phylogenetic tree shows the isolates UESTCC 22.0126, UESTCC 22.0140, UESTCC 22.0141, UESTCC 22.0142, and UESTCC 22.0143 form a clade sister to the isolates P. chimonanthi including the ex-type KUMCC 20-0266 (100% ML, 1.00 PP; Figure 1). Periconia chimonanthi was introduced based on the collection from decaying branches of Chimonanthi praecox (Calycanthaceae) in China [8]. Our collections have similar morphological characteristics in the shape of conidiophores and conidia with the P. chimonanthi on natural substrate. However, P. chengduensis differs from P. chimonanthi in having shorter conidiophores (240–370 μm vs. 410–635 μm) [8]. Furthermore, the culture of P. chimonanthi is black on the reverse side of PDA media; however, P. chengduensis (UESTCC 22.0126) is generally pale yellow on the reverse [8]. Thus, considering the difference in morphological characteristics and phylogenetic analysis, we describe the isolates (UESTCC 22.0126, UESTCC 22.0140, UESTCC 22.0141, UESTCC 22.0142, UESTCC 22.0143) as a new species.
Periconiachimonanthi E.F. Yang, H.B. Jiang, and Phookamsak, in Yang et al. Journal of Fungi 8(3): 243 (2022) Figure 4.
MycoBank: MB 559497
Saprobic on dead leaves of Poaceae. Asexual morph: Colonies on the natural substrate numerous, effuse, dark brown to black, hairy. Conidiophores 200–345 μm long ( = 275, n = 15), 8–13 μm wide ( = 11 μm, n = 15), macronematous, mononematous, straight or slightly flexuous, unbranched, solitary, dark brown to black, septate, smooth to minutely verruculose, thick-walled. Conidiogenous cells polyblastic, pale brown to brown, terminal, integrated, subglobose, smooth to verruculose. Conidia 5.5–9 × 5.5–8.5 μm ( = 7.5 × 7.0 μm, n = 40), solitary or catenate, oval to globose, brown, aseptate, echinulate or verruculose. Sexual morph: Undetermined.
Materials examined: China, Sichuan Province, Chengdu City, Guoxue Park, 30°44′36″ N, 103°55′8″ E, elevation 506 m, on dead leaves of Arundo donax (Poaceae), 16 September 2021, ZH Lu Lu2_2 (HUEST 22.0134), living culture UESTCC 22.0133; ibid., on dead leaves of Imperata cylindrica (Poaceae), ZH Lu Lu8 (HUEST 22.0145), living culture UESTCC 22.0144.
Culture characteristics: Colony on PDA reaching 55 mm diam after 19 days in an incubator under dark conditions at 20 °C, oval almost circular, cottony, hairy at the margin, white at the margin, and gray-green at the middle; reverse: white at the margin and mud yellow to black toward the center.
Notes: The phylogenetic tree showed that our two isolates (UESTCC 22.0133, UESTCC 22.0144) clustered with other P. chimonanthi isolates, including type isolate (KUMCC 20-0266), that were introduced from decaying branches of Chimonanthi praecox (Calycanthaceae) in China [8]. Our two isolates display similar and overlapping morphological characteristics with the holotype of P. chimonanthi [8]. We identified our two collections (UESTCC 22.0133, UESTCC 22.0144) as P. chimonanthi, and this is the first report of P. chimonanthi isolated from Arundo donax and Imperata cylindrica.
Periconia cookei E.W. Mason and M.B. Ellis, Mycol. Pap. 56: 72 (1953) Figure 5.
MycoBank: MB 302477
Saprobic on dead culms of Poaceae. Asexual morph: Colonies on the natural substrate numerous, effuse, brown to black, hairy. Conidiophores 310–515 μm long ( = 365, n = 15), 8.5–14.5 μm wide ( = 11 μm, n = 15), macronematous, mononematous, straight or slightly flexuous, branched, solitary, rarely 1–2 together on stroma, dark brown to black, 3–7-septate, smooth to minutely verruculose, thick-walled. Conidiogenous cells polyblastic, yellowish-brown to brown, terminal, integrated, oval to subglobose, smooth to verruculose. Conidia 5–8 × 5–7 μm ( = 6.5 × 6 μm, n = 40), solitary or catenate, oval to globose, brown to dark brown, aseptate, echinulate or verruculose. Sexual morph: Undetermined.
Material examined: China, Sichuan Province, Chongzhou City, Baiyungou, 30°47′35″ N, 103°23′49″ E, elevation 990 m, on dead culms of Digitaria sanguinalis (Poaceae), 27 September 2021, Z.H. Lu, Lu49_1 (HUEST 22.0135), living culture UESTCC 22.0134.
Culture characteristics: Colony on PDA reaching 39 mm diam after 24 days in an incubator under dark conditions at 20 °C, white, irregular circular, cottony, hairy at the margin; in reverse white at the margin and dark green to black at the middle.
Notes: Periconia cookei was introduced by Mason and Ellis [33] based on the morphology, characterized by conidiophores that are unbranched, septate, pale brown to dark brown, polyblastic and 13–16 μm diameter, catenate, verrucose, brown, mostly spherical conidia. The phylogenetic tree showed that our isolate UESTCC 22.0134 from the dead culms of Digitaria sanguinalis clustered with the isolate of P. cookie MFLUCC 17–1679 [54]. Thus, we identified the isolate UESTCC 22.0134 as a P. cookie. This is the first report of P. cookie occurring on Digitaria sanguinalis in Sichuan Province, China.
Periconia cynodontis Z.H. Lu, P.W. Su, and Maharachch., sp. nov. Figure 6.
MycoBank: MB 847465
Etymology: Name reflects the host genus, Cynodon, from which the fungus was collected.
Saprobic on dead leaves of Poaceae. Asexual morph: Colonies on natural substrate effuse, brown to dark brown, hairy. Mycelium mostly immersed, septate, smooth, brown. Conidiophores 240–335 μm long ( = 310, n = 15), 6.5–10.5 μm wide ( = 8.5 μm, n = 15), macronematous, mononematous, with setiform apices, erect, often curved, solitary, rarely 1–2 together on stroma, unbranched, 6–10-septate, brown to dark brown toward the base, thick-walled, smooth. Conidiogenous cells monoblastic or polyblastic, discrete, directly formed in the middle part of the conidiophores, unilateral, spherical to oval, pale brown to brown, smooth. Conidia 7.5–12.5 × 7–12 μm ( = 10.5 × 10 μm, n = 40), solitary or catenate, globose, brown to dark brown, aseptate, echinulate or verruculose. Sexual morph: Undetermined.
Material examined: China, Sichuan Province, Chengdu City, Guoxue Park, 30°44′36″ N, 103°55′8″ E, elevation 506 m, on dead leaves of Cynodon dactylon (Poaceae), 16 September 2021, ZH Lu Lu4 (HKAS 126515, holotype), ex-type culture CGMCC 3.23927 = UESTCC 22.0127.
Culture characteristics: Colony on PDA reaching 33 mm diam after one week in an incubator under dark conditions at 20 °C, white, circular, cottony, hairy at the margin; in reverse creamy white at the margin and pale yellow at the middle.
Notes: The phylogenetic tree shows that the isolate Periconia cynodontis UESTCC 22.0127 formed a distinct clade within Periconia. Periconia cynodontis resembles P. lateralis in having curved conidiophores, conidiogenous cells directly formed in the middle part of the conidiophores, and solitary or catenate, globose, brown to dark brown conidia [55,56]. However, P. cynodontis differs from P. lateralis by having smaller conidia (7.5–12.5 μm vs. 10.5–15 µm) [55]. Periconia cynodontis differs from P. penniseti and P. neobrittanica by conidiogenous cells that are directly formed in the middle part of the conidiophores [57]. Thus, considering the difference in morphological characteristics and phylogenetic analysis, we describe the isolate UESTCC 22.0127 as Periconia cynodontis sp. nov.
Periconia festucae Z.H. Lu, P.W. Su, and Maharachch., sp. nov. Figure 7.
MycoBank: MB 847466
Etymology: Name after the host genus from which the fungus was collected, Festuca.
Saprobic on dead culms of of Poaceae. Asexual morph: Colonies on the natural substrate numerous, effuse, dark brown to black, hairy. Conidiophores 225–495 μm long ( = 310, n = 15), 10.5–20 μm wide ( = 13 μm, n = 15), macronematous, mononematous, straight or slightly flexuous, solitary or gregarious (1–3 together on substrate), brown to dark brown, 4–8-septate, smooth to minutely verruculose, thick-walled. Conidiogenous cells polyblastic, pale brown to brown, terminal, integrated or discrete, subglobose to globose, smooth to verruculose. Conidia 5–8 × 4.8–8 μm ( = 6.5 × 6.4 μm, n = 40), solitary, oval to globose, light brown to dark brown, aseptate, verruculose. Sexual morph: Undetermined.
Material examined: China, Sichuan Province, Chengdu City, Guoxue Park, 30°44′36″ N, 103°55′8″ E, elevation 506 m, on dead culms of Festuca elata (Poaceae), 16 September 2021, ZH Lu Lu7 (HKAS 126516, holotype), ex-type culture CGMCC 3.23929 = UESTCC 22.0128.
Culture characteristics: Colony on PDA reaching 47 mm diam after 19 days in an incubator under dark conditions at 20 °C, oval to almost circular, cottony, hairy and white at the margin, and pale yellow to white at the middle; in reverse yellow at the margin and mud yellow at the middle.
Notes: The isolate UESTCC 22.0128 is grouped in a well-supported clade and appears to be phylogenetically distinct (98% ML, 0.95 PP; Figure 1) from other sister species. Our collection HKAS 126516 shares similar morphological characteristics in the shape and color of conidiophores and conidia with the P. cookie and P. verrucosa on the natural substrate [54]. However, it differs from P. cookie and P. verrucosa by having significantly wider conidiophores (10.5–20 μm vs. 8.5–14.5 μm and 10.5–15 μm) [34]. The culture of P. cookie is dark green to black on the reverse side of PDA media. However, P. festucae is yellow on the reverse. Periconia festucae and P. verrucosa differ in the number of septa on conidiophores (4–8-septate vs. 2–4-septate). Considering the significant differences in morphology and molecular data, we introduce the isolate UESTCC 22.0128 as a new species.
Periconia imperatae Z.H. Lu, P.W. Su, and Maharachch., sp. nov. Figure 8.
MycoBank: MB 847467
Etymology: Name after the host genus from which the fungus was isolated, Imperata.
Saprobic dead leaves and culms of Poaceae. Asexual morph: Colonies on the natural substrate numerous, effuse, dark brown to black, floccose. Conidiophores 280–520 μm long (= 378, n = 15), 10–16.5 μm wide ( = 12.5 μm, n = 15), macronematous, mononematous, straight or slightly flexuous, branched, solitary, rarely 1–2 together on stroma, brown to dark brown, 3–7-septate, smooth to minutely verruculose, thick-walled. Conidiogenous cells polyblastic, pale brown to brown, terminal, integrated or discrete, oval to subglobose, smooth to verruculose. Conidia 8.5–13.5 × 5.5–8.5 μm ( = 10.5 × 7 μm, n = 40), solitary, ovoid, initially faint yellow or pale brown, becoming brown to dark brown at maturity, aseptate, verruculose. Sexual morph: Undetermined.
Materials examined: China, Sichuan Province, Ngawa Tibetan and Qiang Autonomous Prefecture, Wenchuan County, Xiqiang Valley, 31°29′27″ N, 103°37′1″ E, elevation 1500 m, on dead leaves and culms of Imperata cylindrica (Poaceae), 20 October 2021, ZH Lu w236 (HKAS 126517, holotype), ex-type culture CGMCC 3.23931 = UESTCC 22.0129; ibid., w230_1 (HUEST 22.0146), living culture UESTCC 22.0145; ibid., on dead culms of Neyraudia reynaudiana (Poaceae), ZH Lu w234_5 (HUEST 22.0147), living culture UESTCC 22.0146.
Culture characteristics: Colony on PDA reaching 58 mm diam after 11 days in an incubator under dark conditions at 20 °C, circular, cottony, hairy and white at the margin and yellow toward the middle; yellow at the margin and yellow to brown at the middle in reverse; producing yellow pigments on PDA.
Notes: The phylogenetic tree shows that our three collections (UESTCC 22.0129, UESTCC 22.0145, UESTCC 22.01456) formed a distinct lineage within Periconia as the sister to the P. submersa (99% ML, 1.00 PP; Figure 1). Periconia imperatae resembles P. submersa in having ovoid conidia. However, it differs from P. submersa by having significantly larger conidia (8.5–13.5 × 5.5–8.8 μm vs. 6.5–9.5 × 4.5–5.5 μm) [12]. Furthermore, P. imperatae produced yellow pigment on PDA. Thus, considering the difference in morphology and phylogeny, we describe the isolate UESTCC 22.0129 as a new species.
Periconia penniseti Z.H. Lu, P.W. Su, and Maharachch., sp. nov. Figure 9.
MycoBank: MB 847468
Etymology: Name reflects the host genus, Pennisetum, from which the fungus was collected.
Saprobic on dead culms of Poaceae. Asexual morph: Colonies on the natural substrate numerous, effuse, dark brown to black, hairy. Conidiophores 285–540 μm long ( = 370, n = 15), 11–17 μm wide ( = 14.5 μm, n = 15), macronematous, mononematous, straight to flexuous, unbranched, solitary or gregarious (1–3 together on substrate), dark brown to black, 2–5-septate, smooth to minutely verruculose, thick-walled, forming spherical conidial heads at apex. Conidiogenous cells polyblastic, pale brown to brown, occurring in an apical chain on primary or directly on conidiophore, integrated or discrete, smooth to verruculose. Conidia 6–11.5 × 6–11 μm ( = 9 × 8.5 μm, n = 40), catenate, globose, brown to dark brown, aseptate, verruculose. Sexual morph: Undetermined.
Material examined: China, Sichuan Province, Chengdu City, Guoxue Park, 30°44′36″ N, 103°55′8″ E, elevation 506 m, on dead culms of Pennisetum sp. (Poaceae), 16 September 2021, ZH Lu Lu5 (HKAS 126518, holotype), ex-type culture CGMCC 3.23928 = UESTCC 22.0130.
Culture characteristics: Colony on PDA 23 mm diam after 2 weeks in an incubator under dark conditions at 20 °C, irregular circular, cottony, hairy at the margin, colonies from above white grayish to creamy white; in reverse, white at the margin and pale yellow to khaki at the middle.
Notes: The phylogenetic tree shows that the isolate UESTCC 22.0130 clustered with the ex-type strain of P. neobrittanica (CBS 146062), which was isolated from leaves of Melaleuca styphelioides (Myrtaceae) in California, USA [57]. Periconia penniseti can be distinguished from P. neobrittanica in having longer conidiophores (285–540 μm vs. 100–300 μm) and that are more septate (2–5 vs. 0–1) [57]. Periconia penniseti differs from P. cynodontis in the position of conidiogenous cells. In P. penniseti, the conidiogenous cells arise from the apical part of the conidiophores, while in P. cynodontis, the conidiogenous cells arise from the middle part of the conidiophores. Thus, considering the difference in morphological characteristics and phylogenetic analysis, we describe the isolate UESTCC 22.0130 as a new species.
Periconia pseudobyssoides Markovsk. and A. Kačergius, Mycol. Progr. 13(2): 293 (2014) Figure 10.
MycoBank: MB 804763
Saprobic on dead culms of Poaceae. Asexual morph: Colonies on natural substrate numerous, effuse, brown to dark brown, hairy. Conidiophores 300–565 μm long ( = 445, n = 15), 9.5–14.5 μm wide ( = 11.5 μm, n = 15), macronematous, mononematous, straight or slightly flexuous, solitary, pale brown to brown, 4–7-septate, smooth to minutely verruculose, thick-walled. Conidiogenous cells polyblastic, pale brown to brown, terminal, integrated, subglobose, smooth to verruculose. Conidia 10.5–17 × 10.5–16.5 μm ( = 14 × 13.5 μm, n = 40), solitary, globose, yellowish-brown to golden-brown, aseptate, echinulate, or verruculose. Sexual morph: Undetermined.
Materials examined: China, Sichuan Province, Chongzhou City, Baiyungou, 30°47′35″ N, 103°23′49″ E, elevation 990 m, on dead leaves of Digitaria sanguinalis (Poaceae), 27 September 2021, ZH Lu Lu39 (HUEST 22.0136), living culture UESTCC 22.0135; ibid., Lu96 (HUEST 22.0148), living culture UESTCC 22.0147.
Culture characteristics: Colony on PDA reaching 46 mm diam after 10 days in an incubator under dark conditions at 20 °C, irregular circular, cottony, hairy at the margin, white; reverse: pale yellow at the margin and brown to black at the middle.
Notes: The phylogenetic tree showed that our two isolates (UESTCC 22.0135, UESTCC 22.0147) clustered with other P. pseudobyssoides isolates. Morphologically, our two collections are similar to the description of P. pseudobyssoides. Thus, we identified our collections as P. pseudobyssoides. This is the first record of Periconia species occurring on Digitaria sanguinalis.
Periconia spodiopogonis Y.P. Chen, Z.H. Lu, P.W. Su, and Maharachch., sp. nov. Figure 11.
MycoBank: MB 847469
Etymology: Name reflects the host genus, Spodiopogon, from which the fungus was collected.
Saprobic on dead culms of Spodiopogon ludingensis. Asexual morph: Colonies on the natural substrate numerous, effuse, dark brown to black, hairy. Conidiophores 460–720 μm long ( = 595, n = 15), 9–18 μm wide ( = 15 μm, n = 15), macronematous, mononematous, straight or slightly flexuous, branched, solitary, dark brown to black, slightly paler toward the apex, septate, smooth to minutely verruculose, thick-walled. Conidiogenous cells polyblastic, brown, terminal, integrated or discrete, oval to subglobose, smooth to verruculose. Conidia 14.5–24.5 × 9.5–15.5 μm ( = 20 × 12.5 μm, n = 40), solitary, ellipsoidal to cylindrical, brown to dark brown, aseptate, verruculose. Sexual morph: Undetermined.
Material examined: China, Sichuan Province, Pengzhou City, Huilonggou, 31°18′11″ N, 103°45′35″ E, elevation 1400 m, on dead culms of Spodiopogon ludingensis (Poaceae), 28 July 2021, YP Chen 2021072818 (HKAS 126519, holotype), ex-type culture CGMCC 3.23932 = UESTCC 22.0131.
Culture characteristics: Colony on PDA reaching 68 mm diam after 10 days in an incubator under dark conditions at 20 °C, circular, cottony, hairy, and pale gray at the margin and gray to pale brown at the center; gray at the margin and dark brown to black at the middle in reverse.
Notes: The phylogenetic tree shows that the isolate UESTCC 22.0131 clustered with the ex-type of P. submersa (MFLUCC 16-1098), which was introduced from submerged decaying wood in the Nujiang River, Yunnan Province, China [12]. Periconia spodiopogonis shares similar morphological characteristics, such as the shape of conidiophores and conidia, with the holotype of the P. submersa (HKAS 92738) [12]. However, it differs from P. submersa by having significantly longer conidiophores (460–720 μm vs. 485.5–529.5 μm) and larger conidia (14.5–24.5 × 9.5–15.5 μm vs. 6.5–9.5 × 4.5–5.5 μm) [12]. The BLASTn analysis of P. submersa (MFLUCC 16-1098) and P. spodiopogonis (CGMCC 3.23932) shows 96% identity (484/504, 4 gaps) using ITS. Considering the significant differences in morphology and sequence data, we introduce the isolate UESTCC 22.0131 as a Periconia spodiopogonis sp. nov.
Periconia verrucosa Phukhams., Ertz, Gerstmans, and K.D. Hyde, in Phukhamsakda et al. Fungal Diversity, 102(1): 71 (2020) Figure 12.
MycoBank: MB 557143
Saprobic on dead leaves of Poaceae. Asexual morph: Colonies on the natural substrate numerous, effuse, dark brown, hairy. Conidiophores 353–545 μm long (= 410, n = 15), 10.5–15 μm wide ( = 12.5 μm, n = 15), macronematous, mononematous, straight or flexuous, branched, solitary, dark brown to black, 2–4-septate, smooth to minutely verruculose, thick-walled. Conidiogenous cells polyblastic, yellow to pale brown, terminal, integrated, subglobose, smooth to verruculose. Conidia 4.5–8 × 4.5–7.5 μm ( = 6 × 5.5 μm, n = 40), solitary or catenate, oval to globose, pale brown to dark brown, aseptate, echinulate or verruculose. Sexual morph: Undetermined.
Materials examined: China, Sichuan Province, Chongzhou City, Baiyungou, 30°47′35″ N, 103°23′49″ E, elevation 990 m, on dead leaves of Stipa tenuissima (Poaceae), 27 September 2021, ZH Lu Lu40_1 (HUEST 22.0137), living culture UESTCC 22.0136; ibid., on dead culms of Phyllostachys nigra (Poaceae), Lu53_1 (HUEST 22.0149), living culture UESTCC 22.0148; ibid., Lu98_2 (HUEST 22.0150), living culture UESTCC 22.0149; Ngawa Tibetan and Qiang Autonomous Prefecture, Wenchuan County, Xiqiang Valley, 31°29′27″ N, 103°37′1″ E, elevation 1500 m, on dead culms of Pennisetum polystachion (Poaceae), 20 October 2021, ZH Lu w232_2 (HUEST 22.0151), living culture UESTCC 22.0150; ibid., on dead culms of Neyraudia reynaudiana (Poaceae), w234_4 (HUEST 22.0152), living culture UESTCC 22.0151.
Culture characteristics: Colony on PDA reaching 37 mm diam after 19 days in an incubator under dark conditions at 20 °C, white, circular, cottony, hairy at the margin; in reverse pale yellow at the margin and yellow toward the center.
Notes: The phylogenetic tree showed our five isolates (UESTCC 22.0136, UESTCC 22.0148, UESTCC 22.0149, UESTCC 22.0150, UESTCC 22.0151) clustered with the existing P. verrucosa collections including the type (MFLUCC 17–2158). The type species of P. verrucosa was isolated from decaying stems of Clematis viticella in Belgium [34]. Morphologically, there are no significant differences between our collections and the type species of P. verrucosa. Therefore, our five collections are identified as P. verrucosa, and this is the first report of P. verrucosa from terrestrial habitats in China.
4. Discussion
Species of Periconia have been reported from a large number of host plants, including graminaceous plants [4,8,55]. For example, previously, P. hispidula and P. thysanolaenae were recorded from graminaceous plants in China [8,58]. It is known that this genus produces a number of bioactive secondary metabolites, including terpenes, polyketides, aromatic compounds, and carbohydrate derivatives [59]. Periconia sp. isolated from Torreya grandifolia produced taxol, known as an anticancer compound [60]. Similarly, compounds 2,4-dihydroxy-6-[(1′E,3′E)-penta-1′,3′-dienyl]-benzaldehyde isolated from P. atropurpurea had vigorous antifungal activity against Cladosporium sphaerospermum and C. cladosporioides [19]. Therefore, the genus Periconia has excellent potential for natural product exploration and the development of pharmacological agents. Thus, future genetic and secondary metabolites studies on this chemically highly diverse genus may lead to the discovery of novel biochemical properties unique to the Periconia group.
Sichuan Province along the Yangtze River is a biodiversity hot spot [61,62]. In Sichuan Province, we regularly conduct fungal diversity surveys. This survey collected samples from five sites; 26 Periconia strains were morphologically and phylogenetically characterized, which resulted in 11 species identified and six new taxa described. In the phylogenetic tree, P. salina was clustered within P. byssoides, but Yang et al. [8] showed that P. byssoides and P. salina were not conspecific by the pairwise homogeneity index (PHI) test. Periconia is an ancient and species-rich genus and among 130 morphologically accepted Periconia species [8]. However, only 43 species (including six new species in this study) have molecular data in GenBank. Even though many Periconia species only have ITS and LSU loci. As a highly diverse group, it is also difficult to distinguish species within Periconia using only ITS and LSU gene regions. Therefore, in future studies, more phylogenetic markers with high resolving power are essential to understand the species boundaries in this highly diverse genus. In addition, due to a lack of information and sequence data on many of the species, including the type of Periconia, epitype with living cultures are essential for further comprehensive studies to clarify the taxonomic status of the genus.
Author Contributions
Conceptualization, P.S., Z.L. and S.S.N.M.; methodology, P.S.; writing—original draft preparation, P.S.; writing—review and editing, S.S.N.M.; visualization, P.S., Z.L., W.T. and Y.C.; supervision, S.S.N.M.; funding acquisition, S.S.N.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Talent Introduction and Cultivation Project, University of Electronic Science and Technology of China, grant number A1098531023601245.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequence data are available in NCBI GenBank following the accession numbers in the manuscript.
Acknowledgments
We thank the University of Electronic Science and Technology of China for funding this research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tode, H.J. Fungi Mecklenburgenses Selecti. Fasc. II; Generum Novorum Appendicem; Kessinger Publishing, LLC.: Lüneburg, Germany, 1791; Volume 2, pp. 1–67. [Google Scholar]
- Ellis, M.B. Dematiaceous Hyphomycetes; Commonwealth Mycological Institute: Surrey, UK, 1971; pp. 344–353. [Google Scholar]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Lücking, R.; Boonmee, S.; Bhat, J.D.; Liu, N.-G.; et al. Refined families of Dothideomycetes: Orders and families incertae sedis in Dothideomycetes. Fungal Divers. 2020, 105, 17–318. [Google Scholar] [CrossRef]
- Markovskaja, S.; Kačergius, A. Morphological and molecular characterisation of Periconia pseudobyssoides sp. nov. and closely related P. byssoides. Mycol. Prog. 2014, 13, 291–302. [Google Scholar] [CrossRef]
- Liu, N.; Hongsanan, S.; Yang, J.; Bhat, D.J.; Liu, J.; Jumpathong, J.; Liu, Z. Periconia thailandica (Periconiaceae), a new species from Thailand. Phytotaxa 2017, 323, 253–263. [Google Scholar] [CrossRef]
- Calvillo-Medina, R.P.; Cobos-Villagran, A.; Raymundo, T. Periconia citlaltepetlensis sp. nov. (Periconiaceae, Pleosporales): A psychrotolerant fungus from high elevation volcanic glacier (Mexico). Phytotaxa 2020, 459, 235–247. [Google Scholar] [CrossRef]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Kudo, H.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y.; et al. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, E.-F.; Phookamsak, R.; Jiang, H.-B.; Tibpromma, S.; Bhat, D.J.; Karunarathna, S.C.; Dai, D.-Q.; Xu, J.-C.; Promputtha, I. Taxonomic reappraisal of Periconiaceae with the description of three new Periconia species from China. J. Fungi 2022, 8, 243. [Google Scholar] [CrossRef] [PubMed]
- Booth, C. Didymosphaeria igniaria sp. nov., the perfect state of Periconia igniaria. Trans. Br. Mycol. Soc. 1968, 51, 803–805. [Google Scholar] [CrossRef]
- Kohlmeyer, J. Marine fungi of Hawaii including the new genus Helicascus. Can. J. Bot. 1969, 47, 1469–1487. [Google Scholar] [CrossRef]
- Cantrell, S.; Hanlin, R.; Emiliano, A. Periconia variicolor sp. nov., a new species from Puerto Rico. Mycologia 2007, 99, 482–487. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Thilini Chethana, K.W.; Clericuzio, M.; Dayarathne, M.C.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.-Q.; et al. Fungal diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Matsushima, T. Microfungi of the Solomon Islands and Papua-New Guinea; Published by the Author: Kobe, Japan, 1971. [Google Scholar]
- Matsushima, T. Icones Microfungorum a Matsushima Lectorum; Published by the Author: Kobe, Japan, 1975. [Google Scholar]
- Ellis, M.B. More Dematiaceous Hyphomycetes; Commonwealth Mycological Institute: Kew, Surrey, England, 1976; pp. 352–359. [Google Scholar]
- Goga, N. Periconia circinata—A new pathogen of roots and stem base of wheat in northwestern Romania. An. Inst. De Cercet. Pentru Cereale Plante Teh. Fundulea 2000, 67, 205–214. [Google Scholar]
- Sarkar, T.; Chakraborty, P.; Karmakar, A.; Saha, A.; Saha, D. First report of Periconia macrospinosa causing leaf necrosis of pointed gourd in India. J. Plant Pathol. 2019, 101, 1281. [Google Scholar] [CrossRef] [Green Version]
- Gunasekaran, R.; Janakiraman, D.; Rajapandian, S.G.K.; Appavu, S.P.; Namperumalsamy, V.P.; Prajna, L. Periconia species—An unusual fungal pathogen causing mycotic keratitis. Indian J. Med. Microbiol. 2021, 39, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Teles, H.L.; Sordi, R.; Silva, G.H.; Castro-Gamboa, I.; Bolzani, V.d.S.; Pfenning, L.H.; Magalhães de Abreu, L.; Costa-Neto, C.M.; Young, M.C.M.; Araújo, Â.R. Aromatic compounds produced by Periconia atropurpurea, an endophytic fungus associated with Xylopia aromatica. Phytochemistry 2006, 67, 2686–2690. [Google Scholar] [CrossRef]
- Bhilabutra, W.; Techowisan, T.; Peberdy, J.F.; Lumyong, S. Antimicrobial activity of bioactive compounds from Periconia siamensis CMUGE015. J. Microbiol. 2007, 2, 749–755. [Google Scholar] [CrossRef]
- Nannizzi, A. Repertorio Sistematico dei Miceti dell’uomo e degli Animali; SA Poligrafica Meini Siena: Siena, Italy, 1934; pp. 1–557. [Google Scholar]
- Hyde, K.D.; Jones, E.B.G.; Liu, J.-K.; Ariyawansa, H.; Boehm, E.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.-Q.; et al. Families of Dothideomycetes. Fungal Divers. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; Hongsanan, S.; Ryberg, M.; Ariyawansa, H.; Chomnunti, P.; Bahkali, A.; Hyde, K. The evolution of Massarineae with Longipedicellataceae fam. nov. Mycosphere 2016, 7, 1713–1731. [Google Scholar] [CrossRef]
- Link, H.F. Observationes in ordines plantarum naturales. Dissertatio. Mag Ges Naturf Freunde Berlin 1809, 3, 3–42. [Google Scholar]
- Saccardo, P.A. Conspectus generum fungorum Italiae inferorium. Michelia 1880, 2, 1–38. [Google Scholar]
- Saccardo, P.A.; Traverso, G.B.; Trotter, A. Sylloge Fungorum Omnium Hucusque Cognitorum; Sumptibus Auctoris: Patavii, Italy, 1886. [Google Scholar]
- Okada, G.; Jacobs, K.; Kirisits, T.; Louis-Seize, G.; Seifert, K.; Sugita, T.; Takematsu, A.; Wingfield, M. Epitypification of Graphium penicillioides Corda, with comments on the phylogeny and taxonomy of graphium-like synnematous fungi. Stud. Mycol. 2000, 45, 169–186. [Google Scholar]
- Partridge, E.C.; Morgan-Jones, G. Notes on Hyphomycetes. LXXXVIII. New genera in which to classify Alysidium resinae and Pycnostysanus azaleae, with a consideration of Sorocybe. Mycotaxon 2002, 83, 335–352. [Google Scholar]
- Schubert, K.; Groenewald, J.Z.; Braun, U.; Dijksterhuis, J.; Starink, M.; Hill, C.F.; Zalar, P.; de Hoog, G.S.; Crous, P.W. Biodiversity in the Cladosporium herbarum complex (Davidiellaceae, Capnodiales), with standardisation of methods for Cladosporium taxonomy and diagnostics. Stud. Mycol. 2007, 58, 105–156. [Google Scholar] [CrossRef]
- Linnakoski, R.; de Beer, Z.W.; Duong, T.A.; Niemelä, P.; Pappinen, A.; Wingfield, M.J. Grosmannia and Leptographium spp. associated with conifer-infesting bark beetles in Finland and Russia, including Leptographium taigense sp. nov. Antonie Van Leeuwenhoek 2012, 102, 375–399. [Google Scholar] [CrossRef] [PubMed]
- Persoon, C.H. Synopsis Methodica Fungorum; Apvd Henricvm Dieterich: Gottinga, Germany, 1801. [Google Scholar]
- Saccardo, P.A.; Trotter, A. Supplementum Universale, Pars IX. Ascomycetae—Deuteromycetae; Sylloge Fungorum: Padova, Italy, 1913; Volume 22. [Google Scholar]
- Mason, E.W.; Ellis, M.B. British species of Periconia. Mycol. Papers 1953, 56, 1–127. [Google Scholar]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Gareth Jones, E.B.; Bhat, D.J.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.-G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Dayarathne, M.; Jones, E.G.B.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.; Khongphinitbunjong, K.; Chomnunti, P.; Hyde, K.D. Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Tennakoon, D.S.; Kuo, C.-H.; Maharachchikumbura, S.S.N.; Thambugala, K.M.; Gentekaki, E.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; de Silva, N.I.; Promputtha, I.; et al. Taxonomic and phylogenetic contributions to Celtis formosana, Ficus ampelas, F. septica, Macaranga tanarius and Morus australis leaf litter inhabiting microfungi. Fungal Divers. 2021, 108, 1–215. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-coverage ITS primers for the DNA-based identification of ascomycetes and basidiomycetes in environmental samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [Green Version]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibbett, D.S. Phylogenetic evidence for horizontal transmission of group I introns in the nuclear ribosomal DNA of mushroom-forming fungi. Mol. Biol. Evol. 1996, 13, 903–917. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Jaklitsch, W.M.; Komon, M.; Kubicek, C.P.; Druzhinina, I.S. Hypocrea voglmayrii sp. nov. from the Austrian Alps represents a new phylogenetic clade in Hypocrea/Trichoderma. Mycologia 2005, 97, 1365–1378. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Sung, G.-H.; Sung, J.-M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Yu, G. Using ggtree to visualize data on tree-like structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Chaiwan, N.; Norphanphoun, C.; Boonmee, S.; Camporesi, E.; Chethana, K.W.T.; Dayarathne, M.C.; De Silva, N.I.; Dissanayake, A.J.; Ekanayaka, A.H.; et al. Mycosphere notes 169–224. Mycosphere 2018, 9, 271–430. [Google Scholar] [CrossRef]
- Chuaseeharonnachai, C.; Somrithipol, S.; Boonyuen, N. Periconia notabilis sp. nov. and a new record and notes on the genus in Thailand. Mycotax 2016, 131, 491–502. [Google Scholar] [CrossRef]
- Prasher, I.; Verma, R. Periconia species new to North-Western Himalayas. J. New Biol. Rep. 2012, 1, 01–02. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Lombard, L.; Roets, F.; Swart, W.J.; Alvarado, P.; Carnegie, A.J.; Moreno, G.; Luangsaard, J.; Thangavel, R.; et al. Fungal Planet description sheets: 951–1041. Persoonia 2019, 43, 223. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, T. Flora Fungorum Sinicorum, Vol. 51, Dematiaceous Hyphomycetes from Soil; Science Press: Beijing, China, 2019; pp. 322–328. [Google Scholar]
- Azhari, A.; Supratman, U. The chemistry and pharmacology of fungal genus Periconia: A Review. Sci. Pharm. 2021, 89, 34. [Google Scholar] [CrossRef]
- Li, J.Y.; Sidhu, R.S.; Ford, E.J.; Long, D.M.; Hess, W.M.; Strobel, G.A. The induction of taxol production in the endophytic fungus—Periconia sp. from Torreya grandifolia. J. Ind. Microbiol. Biotechnol. 1998, 20, 259–264. [Google Scholar] [CrossRef]
- Yan, K.; Abbas, M.; Meng, L.; Cai, H.; Peng, Z.; Li, Q.; El-Sappah, A.H.; Yan, L.; Zhao, X. Analysis of the fungal diversity and community structure in sichuan dark tea during pile-fermentation. Front. Microbiol. 2021, 12, 706714. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Wei, Q.; Feng, R.; Liu, Y.; Liang, H.; Li, J.; Yan, K. Diversity and spatial distribution of endophytic fungi in Cinnamomum longepaniculatum of Yibin, China. Arch. Microbiol. 2021, 203, 3361–3372. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogram of the best-scoring ML consensus tree based on a combined dataset (SSU, ITS, LSU, TEF1, and RPB2) of Periconia. Novel isolates are indicated in red. Isolates from type materials are in bold. The ML ultrafast bootstrap values/Bayesian PP greater than 95%/0.95 are shown at the respective nodes. The tree is rooted with Massarina cisti CBS 266.62 (Massarinaceae, Pleosporales).
Figure 1.
Phylogram of the best-scoring ML consensus tree based on a combined dataset (SSU, ITS, LSU, TEF1, and RPB2) of Periconia. Novel isolates are indicated in red. Isolates from type materials are in bold. The ML ultrafast bootstrap values/Bayesian PP greater than 95%/0.95 are shown at the respective nodes. The tree is rooted with Massarina cisti CBS 266.62 (Massarinaceae, Pleosporales).

Figure 2.
Periconia byssoides (HUEST 22.0133). (a,b) Colonies on the natural substrate; (c,d) conidiophores with spherical conidial heads; (e,f) conidial heads bearing conidiogenous cells and conidia; (g–j) conidia; (k) germinating conidium; (l,m) colony on PDA from above and below. Scale bars: (c) = 100 μm, (e,g,k) = 10 μm. Scale bar of (c) applies to (d). Scale bar of (e) applies to (f). Scale bar of (g) applies to (h). Scale bar of (i) applies to (j).
Figure 2.
Periconia byssoides (HUEST 22.0133). (a,b) Colonies on the natural substrate; (c,d) conidiophores with spherical conidial heads; (e,f) conidial heads bearing conidiogenous cells and conidia; (g–j) conidia; (k) germinating conidium; (l,m) colony on PDA from above and below. Scale bars: (c) = 100 μm, (e,g,k) = 10 μm. Scale bar of (c) applies to (d). Scale bar of (e) applies to (f). Scale bar of (g) applies to (h). Scale bar of (i) applies to (j).

Figure 3.
Periconia chengduensis (HKAS 126514, holotype). (a,b) Colonies on the natural substrate; (c,d) conidiophores with spherical conidial heads; (e,f) apically branch conidiophores with conidial head; (g–l) conidia; (m) germinating conidium; (n,o) colony on PDA from above and below. Scale bars: (c) = 100 μm, (e,g,m) = 10 μm. Scale bar of (c) applies to (d). Scale bar of (e) applies to (f). Scale bar of (g) applies to (h–l).
Figure 3.
Periconia chengduensis (HKAS 126514, holotype). (a,b) Colonies on the natural substrate; (c,d) conidiophores with spherical conidial heads; (e,f) apically branch conidiophores with conidial head; (g–l) conidia; (m) germinating conidium; (n,o) colony on PDA from above and below. Scale bars: (c) = 100 μm, (e,g,m) = 10 μm. Scale bar of (c) applies to (d). Scale bar of (e) applies to (f). Scale bar of (g) applies to (h–l).

Figure 4.
Periconia chimonanthi (HUEST 22.0134). (a,b) Colonies on the natural substrate; (c,d) conidiophores with spherical conidial heads; (e,f) conidial heads bearing conidiogenous cells and conidia; (g–k) conidia; (l) germinating conidium; (m,n) colony on PDA from above and below. Scale bars: (c) = 100 μm, (e,g,i) = 10 μm. Scale bar of (c) applies to (d). Scale bar of (e) applies to (f). Scale bar of (g) applies to (h–k).
Figure 4.
Periconia chimonanthi (HUEST 22.0134). (a,b) Colonies on the natural substrate; (c,d) conidiophores with spherical conidial heads; (e,f) conidial heads bearing conidiogenous cells and conidia; (g–k) conidia; (l) germinating conidium; (m,n) colony on PDA from above and below. Scale bars: (c) = 100 μm, (e,g,i) = 10 μm. Scale bar of (c) applies to (d). Scale bar of (e) applies to (f). Scale bar of (g) applies to (h–k).

Figure 5.
Periconia cookei (HUEST 22.0135). (a,b) Colonies on the natural substrate; (c,d) conidiophores with spherical conidial heads; (e,f) apically branch conidiophores with conidial head; (g–j) conidia; (k) germinating conidium; (l,m) colony on PDA from above and below. Scale bars: (c) = 100 μm, (e,g,k) = 10 μm. Scale bar of (c) applies to (d) Scale bar of (e) applies to (f). Scale bar of (g) applies to (h–j).
Figure 5.
Periconia cookei (HUEST 22.0135). (a,b) Colonies on the natural substrate; (c,d) conidiophores with spherical conidial heads; (e,f) apically branch conidiophores with conidial head; (g–j) conidia; (k) germinating conidium; (l,m) colony on PDA from above and below. Scale bars: (c) = 100 μm, (e,g,k) = 10 μm. Scale bar of (c) applies to (d) Scale bar of (e) applies to (f). Scale bar of (g) applies to (h–j).

Figure 6.
Periconia cynodontis (HKAS 126515, holotype). (a–c) Colonies on the natural substrate; (d,e) conidiophores; (f,g) conidiophores bearing conidia laterally; (h–j) conidia; (k) germinating conidium; (l,m) colony on PDA from above and below. Scale bars: (d) = 20 μm, (f,h,k) = 10 μm. Scale bar of (d) applies to (e). Scale bar of (f) applies to (g). Scale bar of (h) applies to (i,j).
Figure 6.
Periconia cynodontis (HKAS 126515, holotype). (a–c) Colonies on the natural substrate; (d,e) conidiophores; (f,g) conidiophores bearing conidia laterally; (h–j) conidia; (k) germinating conidium; (l,m) colony on PDA from above and below. Scale bars: (d) = 20 μm, (f,h,k) = 10 μm. Scale bar of (d) applies to (e). Scale bar of (f) applies to (g). Scale bar of (h) applies to (i,j).

Figure 7.
Periconia festucae (HKAS 126516, holotype). (a–c) Colonies on the natural substrate; (d,e) conidiophores with spherical conidial heads; (f,g) apical branch conidiophores with conidial head; (h–l) conidia; (m) germinating conidium; (n,o) colony on PDA from above and below. Scale bars: (d) = 100 μm, (f,h,m) = 10 μm. Scale bar of (d) applies to (e). Scale bar of (f) applies to (g). Scale bar of (h) applies to (i–l).
Figure 7.
Periconia festucae (HKAS 126516, holotype). (a–c) Colonies on the natural substrate; (d,e) conidiophores with spherical conidial heads; (f,g) apical branch conidiophores with conidial head; (h–l) conidia; (m) germinating conidium; (n,o) colony on PDA from above and below. Scale bars: (d) = 100 μm, (f,h,m) = 10 μm. Scale bar of (d) applies to (e). Scale bar of (f) applies to (g). Scale bar of (h) applies to (i–l).

Figure 8.
Periconia imperatae (HKAS 126517, holotype). (a–c) Colonies on the natural substrate; (d,e) conidiophores with spherical conidial heads; (f,g) apically branch conidiophores with conidial head; (h–k) conidia; (l) germinating conidium; (m,n) colony on PDA from above and below. Scale bars: (d) = 100 μm, (f,h,l) = 10 μm. Scale bar of (d) applies to (e). Scale bar of (f) applies to (g). Scale bar of (h) applies to (i–k).
Figure 8.
Periconia imperatae (HKAS 126517, holotype). (a–c) Colonies on the natural substrate; (d,e) conidiophores with spherical conidial heads; (f,g) apically branch conidiophores with conidial head; (h–k) conidia; (l) germinating conidium; (m,n) colony on PDA from above and below. Scale bars: (d) = 100 μm, (f,h,l) = 10 μm. Scale bar of (d) applies to (e). Scale bar of (f) applies to (g). Scale bar of (h) applies to (i–k).

Figure 9.
Periconia penniseti (HKAS 126518, holotype). (a,b) Colonies on the natural substrate; (c,d) conidiophores with spherical conidial heads; (e,f) conidial heads bearing conidiogenous cells and conidia; (g–i) conidiogenous cells bearing conidia in short chains; (j–l) conidia; (m) germinating conidium; (n,o) colony on PDA from above and below. Scale bars: (c,d) = 100 μm, (e,g,j,m) = 10 μm. Scale bar of (e) applies to (f). Scale bar of (g) applies to (h,i). Scale bar of (j) applies to (k,l).
Figure 9.
Periconia penniseti (HKAS 126518, holotype). (a,b) Colonies on the natural substrate; (c,d) conidiophores with spherical conidial heads; (e,f) conidial heads bearing conidiogenous cells and conidia; (g–i) conidiogenous cells bearing conidia in short chains; (j–l) conidia; (m) germinating conidium; (n,o) colony on PDA from above and below. Scale bars: (c,d) = 100 μm, (e,g,j,m) = 10 μm. Scale bar of (e) applies to (f). Scale bar of (g) applies to (h,i). Scale bar of (j) applies to (k,l).

Figure 10.
Periconia pseudobyssoides (HUEST 22.0136). (a–c) Colonies on the natural substrate; (d,e) conidiophores with spherical conidial heads; (f,g) conidial heads bearing conidiogenous cells and conidia; (h–j) conidia; (k) germinating conidium; (l,m) colony on PDA from above and below. Scale bars: (d) = 100 μm, (f,h,k) = 10 μm. Scale bar of (d) applies to (e). Scale bar of (f) applies to (g). Scale bar of (h) applies to (i,j).
Figure 10.
Periconia pseudobyssoides (HUEST 22.0136). (a–c) Colonies on the natural substrate; (d,e) conidiophores with spherical conidial heads; (f,g) conidial heads bearing conidiogenous cells and conidia; (h–j) conidia; (k) germinating conidium; (l,m) colony on PDA from above and below. Scale bars: (d) = 100 μm, (f,h,k) = 10 μm. Scale bar of (d) applies to (e). Scale bar of (f) applies to (g). Scale bar of (h) applies to (i,j).

Figure 11.
Periconia spodiopogonis (HKAS 126519, holotype). (a–c) Colonies on the natural substrate; (d,e) conidiophores; (f,g) apically branch conidiophores with conidial head; (h–k) conidia; (l) germinating conidium; (m,n) colony on PDA from above and below. Scale bars: (d) = 100 μm, (f,g,h,j,l) = 20 μm. Scale bar of (d) applies to (e). Scale bar of (h) applies to (i). Scale bar of (j) applies to (k).
Figure 11.
Periconia spodiopogonis (HKAS 126519, holotype). (a–c) Colonies on the natural substrate; (d,e) conidiophores; (f,g) apically branch conidiophores with conidial head; (h–k) conidia; (l) germinating conidium; (m,n) colony on PDA from above and below. Scale bars: (d) = 100 μm, (f,g,h,j,l) = 20 μm. Scale bar of (d) applies to (e). Scale bar of (h) applies to (i). Scale bar of (j) applies to (k).

Figure 12.
Periconia verrucosa (HUEST 22.0137). (a,b) Colonies on the natural substrate; (c,d) conidiophores with spherical conidial heads; (e,f) apically branch conidiophores with conidial head; (g–i) conidia; (j) germinating conidium; (k,l) colony on PDA from above and below. Scale bars: (c) = 100 μm, (e,g,j) = 10 μm. Scale bar of (c) applies to (d). Scale bar of (e) applies to (f). Scale bar of (g) applies to (h–i).
Figure 12.
Periconia verrucosa (HUEST 22.0137). (a,b) Colonies on the natural substrate; (c,d) conidiophores with spherical conidial heads; (e,f) apically branch conidiophores with conidial head; (g–i) conidia; (j) germinating conidium; (k,l) colony on PDA from above and below. Scale bars: (c) = 100 μm, (e,g,j) = 10 μm. Scale bar of (c) applies to (d). Scale bar of (e) applies to (f). Scale bar of (g) applies to (h–i).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Loci used in this study with the corresponding PCR primers and conditions.
| Locus | PCR Primers | PCR Thermal Cycles | References |
|---|---|---|---|
| ITS | ITS9mun/ITS4_KYO1 | (94 °C: 3 min) × 1 cycles (94 °C: 30 s, 56 °C: 30 s, 72 °C: 30 s) × 35 cycles (72 °C: 5 min) × 1 cycles | [39] |
| LSU | LR0R/LR5 | (94 °C: 3 min) × 1 cycles (94 °C: 30 s, 56 °C: 30 s, 72 °C: 1 min) × 35 cycles (72 °C: 5 min) × 1 cycles | [40] |
| SSU | PNS1/NS41 | (94 °C: 3 min) × 1 cycles (94 °C: 30 s, 56 °C: 30 s, 72 °C: 1 min) × 35 cycles (72 °C: 5 min) × 1 cycles | [41] |
| TEF1 | EF1-983/EF1-2218R or TEF1LLErev | (94 °C: 3 min) × 1 cycles (94 °C: 30 s, 52 °C: 30 s, 72 °C: 1 min) × 35 cycles (72 °C: 5 min) × 1 cycles | [42,43] |
| RPB2 | dRPB2-5f or RPB2-5F2/fRPB2-7cR | (94 °C: 3 min) × 1 cycles (94 °C: 30 s, 52 °C: 30 s, 72 °C: 1 min) × 35 cycles (72 °C: 5 min) × 1 cycles | [44,45] |
Table 2.
Species details and their GenBank accession numbers used in phylogenetic analyses.
| Organism | Culture/Specimen No. | SSU | ITS | LSU | TEF1 | RPB2 |
|---|---|---|---|---|---|---|
| Massarina cisti | CBS 266.62 | FJ795490 | LC014568 | AB807539 | AB808514 | FJ795464 |
| Periconia algeriana | CBS 321.79 | - | MH861212 | MH872979 | - | - |
| P. alishanica | KUMCC 19–0174 = NCYU19–0347 | - | MW063167 | MW063231 | MW183792 | - |
| P. alishanica | MFLUCC 19–0145 | - | MW063165 | MW063229 | MW183790 | - |
| P. alishanica | NCYUCC 19–0186 | - | MW063166 | MW063230 | MW183791 | - |
| P. aquatica | MFLUCC 16–0912 | - | KY794701 | KY794705 | KY814760 | - |
| P. artemisiae | KUMCC 20–0265 | MW448658 | MW448657 | MW448571 | MW460898 | - |
| P. atropurpurea | CBS 381.55 | - | MH857524 | MH869061 | - | - |
| P. banksiae | CBS 129526 = CPC 17282 | - | JF951147 | NG_064279 | - | - |
| P. byssoides | MFLUCC 17–2292 = KUMCC 18–0272 = C292 | MK347858 | MK347751 | MK347968 | MK360069 | MK434886 |
| P. byssoides | MFLUCC 18–1548 | MK347902 | MK347794 | MK348013 | - | MK434863 |
| P. byssoides | MFLUCC 18–1553 = C457 | MK347914 | MK347806 | MK348025 | MK360068 | MK434858 |
| P. byssoides | MFLUCC 19–0134 = NCYUCC 19–0166 = MFLU 18–2545 | - | MW063164 | MW063228 | MW183789 | - |
| P. byssoides | MFLUCC 20–0172 = NCYUCC 19–0313 | - | MW063162 | MW063226 | - | - |
| P. byssoides | NCYUCC 19–0314 = NYCU 19–0040 | - | MW063163 | MW063227 | - | - |
| P. byssoides | UESTCC 22.0132 | OP956054 | OP955985 | OP956010 | OP961451 | OP961468 |
| P. byssoides | UESTCC 22.0137 | OP956036 | OP955967 | OP955992 | OP961433 | OP961458 |
| P. byssoides | UESTCC 22.0138 | OP956038 | OP955969 | OP955994 | OP961435 | OP961461 |
| P. byssoides | UESTCC 22.0139 | OP956055 | OP955986 | OP956011 | OP961452 | - |
| P. caespitosa | LAMIC 110/16 | - | MH051906 | MH051907 | - | - |
| P. chengduensis | CGMCC 3.23930 = UESTCC 22.0126 | OP956056 | OP955987 | OP956012 | OP961453 | OP961469 |
| P. chengduensis | UESTCC 22.0140 | OP956046 | OP955977 | OP956002 | OP961443 | OP961465 |
| P. chengduensis | UESTCC 22.0141 | OP956041 | OP955972 | OP955997 | OP961438 | - |
| P. chengduensis | UESTCC 22.0142 | OP956047 | OP955978 | OP956003 | OP961444 | - |
| P. chengduensis | UESTCC 22.0143 | OP956050 | OP955981 | OP956006 | OP961447 | OP961466 |
| P. chimonanthi | KUMCC 20–0266 | MW448656 | NR_176752 | MW448572 | MW460897 | - |
| P. chimonanthi | UESTCC 22.0133 | OP956033 | OP955964 | OP955989 | OP961430 | OP961455 |
| P. chimonanthi | UESTCC 22.0144 | OP956043 | OP955974 | OP955999 | OP961440 | - |
| P. circinata | CBS 263.37 | - | MW810265 | MH867413 | MW735660 | - |
| P. citlaltepetlensis | ENCB 140251 = IOM 325319.1 | - | MH890645 | MT625978 | - | - |
| P. citlaltepetlensis | IOM 325319.2 | - | MT649221 | MT649216 | - | - |
| P. cookei | MFLUCC 17–1399 | - | MG333490 | MG333493 | MG438279 | - |
| P. cookei | MFLUCC 17–1679 | - | - | MG333492 | MG438278 | - |
| P. cookei | UESTCC 22.0134 | OP956037 | OP955968 | OP955993 | - | OP961459 |
| P. cortaderiae | MFLUCC 15–0451 | KX986346 | KX965734 | KX954403 | KY429208 | - |
| P. cortaderiae | MFLUCC 15–0453 = ICMP 21429 | - | KX965733 | KX954402 | KY320574 | - |
| P. cortaderiae | MFLUCC 15–0457 = ICMP 21414 | KX986345 | KX965732 | KX954401 | KY310703 | - |
| P. cynodontis | CGMCC 3.23927 = UESTCC 22.0127 | OP909920 | OP909925 | OP909921 | OP961434 | OP961460 |
| P. cyperacearum | CPC 32138 = CBS 144434 | - | MH327815 | MH327851 | - | - |
| P. delonicis | MFLUCC 17–2584 = KUMCC 18–0275 | MK347832 | - | MK347941 | MK360071 | MK434901 |
| P. didymosporum | MFLU 15–0058 | KP761738 | KP761734 | KP761731 | KP761728 | KP761721 |
| P. digitata | CBS 510.77 | AB797271 | LC014584 | AB807561 | AB808537 | - |
| P. elaeidis | MFLUCC 17–0087 | MH108551 | MG742713 | MH108552 | - | - |
| P. epilithographicola | CBS 144017 | - | NR_157477 | - | - | - |
| P. epilithographicola | MFLUCC 21–0153 | OL606144 | OL753687 | OL606155 | OL912948 | - |
| P. festucae | CGMCC 3.23929 = UESTCC 22.0128 | OP956042 | OP955973 | OP955998 | OP961439 | OP961463 |
| P. genistae | CBS 322.79 | - | MH861213 | MH872980 | - | - |
| P. homothallica | CBS 139698 = JCM 13100 = MAFF 239610 = KT916 = HHUF 29105 | AB797275 | AB809645 | NG_059397 | AB808541 | - |
| P. igniaria | CBS 298.66 | - | MH858798 | MH870438 | - | - |
| P. igniaria | CBS 583.66 | - | MH858888 | MH870553 | - | - |
| P. imperatae | CGMCC 3.23931 = UESTCC 22.0129 | OP956053 | OP955984 | OP956009 | OP961450 | OP961467 |
| P. imperatae | UESTCC 22.0145 | OP956048 | OP955979 | OP956004 | OP961445 | - |
| P. imperatae | UESTCC 22.0146 | OP956052 | OP955983 | OP956008 | OP961449 | - |
| P. lateralis | CBS 292.36 | - | MH855804 | MH867311 | - | - |
| P. macrospinosa | CBS 135663 = CPC 22898 | KP184080 | KP183999 | KP184038 | - | - |
| P. minutissima | MFLUCC 15–0245 | - | KY794703 | KY794707 | - | - |
| P. neobrittanica | CPC 37903 = CBS 146062 | - | MN562149 | MN567656 | - | - |
| P. palmicola | MFLUCC14–0400 | MN648319 | - | MN648327 | MN821070 | - |
| P. penniseti | CGMCC 3.23928 = UESTCC 22.0130 | OP956040 | OP955971 | OP955996 | OP961437 | OP961462 |
| P. prolifica | CBS 209.64 | - | NR_160097 | MH870050 | - | - |
| P. pseudobyssoides | DLUCC 0850 | - | MG333491 | MG333494 | MG438280 | - |
| P. pseudobyssoides | H4151 = MAFF 243868 | AB797278 | LC014587 | AB807568 | AB808544 | - |
| P. pseudobyssoides | KUMCC 20–0263 | MW444853 | MW444851 | MW444852 | MW460894 | - |
| P. pseudobyssoides | MAFF 243874 = TS102 = HHUF 28257 = H 4790 | AB797270 | LC014588 | AB807560 | AB808536 | - |
| P. pseudobyssoides | UESTCC 22.0135 | OP956034 | OP955965 | OP955990 | OP961431 | OP961456 |
| P. pseudobyssoides | UESTCC 22.0147 | OP956044 | OP955975 | OP956000 | OP961441 | - |
| P. pseudodigitata | CBS 139699 = JCM 13166 = MAFF 239676 = KT 1395 = HHUF 29730 | AB797274 | LC014591 | AB807564 | AB808540 | - |
| P. pseudodigitata | JCM 13164 = MAFF 239674 = KT644 | AB797272 | LC014589 | AB807562 | AB808538 | - |
| P. pseudodigitata | JCM 13165 = MAFF 239675 = KT 1195A | AB797273 | LC014590 | AB807563 | AB808539 | - |
| P. sahariana | CBS 320.79 | – | MH861211 | MH872978 | - | - |
| P. salina | GJ374 = MFLU 19–1235 | MN017912 | MN047086 | MN017846 | - | - |
| P. spodiopogonis | CGMCC 3.23932 = UESTCC 22.0131 | OP956032 | OP955963 | OP955988 | OP961429 | OP961454 |
| P. submersa | MFLUCC 16–1098 | - | KY794702 | KY794706 | KY814761 | - |
| P. thailandica | MFLUCC 17–0065 | KY753889 | KY753887 | KY753888 | – | - |
| P. thysanolaenae | KUMCC 20–0262 | NG_081407 | MW442967 | MW444850 | MW460896 | - |
| P. variicolor | CBS 120374 = SACCR 64 | - | DQ336713 | - | - | - |
| P. verrucosa | MFLUCC 17–2158 | MT226686 | MT310617 | MT214572 | MT394631 | - |
| P. verrucosa | UESTCC 22.0136 | OP956035 | OP955966 | OP955991 | OP961432 | OP961457 |
| P. verrucosa | UESTCC 22.0148 | OP956039 | OP955970 | OP955995 | OP961436 | - |
| P. verrucosa | UESTCC 22.0149 | OP956045 | OP955976 | OP956001 | OP961442 | OP961464 |
| P. verrucosa | UESTCC 22.0150 | OP956049 | OP955980 | OP956005 | OP961446 | - |
| P. verrucosa | UESTCC 22.0151 | OP956051 | OP955982 | OP956007 | OP961448 | - |
The newly generated sequences are indicated in red, and the ex-type strains are in bold. Missing sequences are indicated by “-”.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Su, P.; Lu, Z.; Tian, W.; Chen, Y.; Maharachchikumbura, S.S.N. Six Additions to the Genus Periconia (Dothideomycetes: Periconiaceae) from Graminaceous Plants in China. J. Fungi 2023, 9, 300. https://doi.org/10.3390/jof9030300
AMA Style
Su P, Lu Z, Tian W, Chen Y, Maharachchikumbura SSN. Six Additions to the Genus Periconia (Dothideomycetes: Periconiaceae) from Graminaceous Plants in China. Journal of Fungi. 2023; 9(3):300. https://doi.org/10.3390/jof9030300
Chicago/Turabian StyleSu, Pengwei, Zhenghua Lu, Whenhui Tian, Yanpeng Chen, and Sajeewa S. N. Maharachchikumbura. 2023. "Six Additions to the Genus Periconia (Dothideomycetes: Periconiaceae) from Graminaceous Plants in China" Journal of Fungi 9, no. 3: 300. https://doi.org/10.3390/jof9030300
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.