


Taxonomy and Phylogenetic Relationships of Clavulinopsis (Clavariaceae, Agaricales): Description of Six New Species and One Newly Recorded Species from China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Morphological Studies
2.4. Alignment and Phylogenetic Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
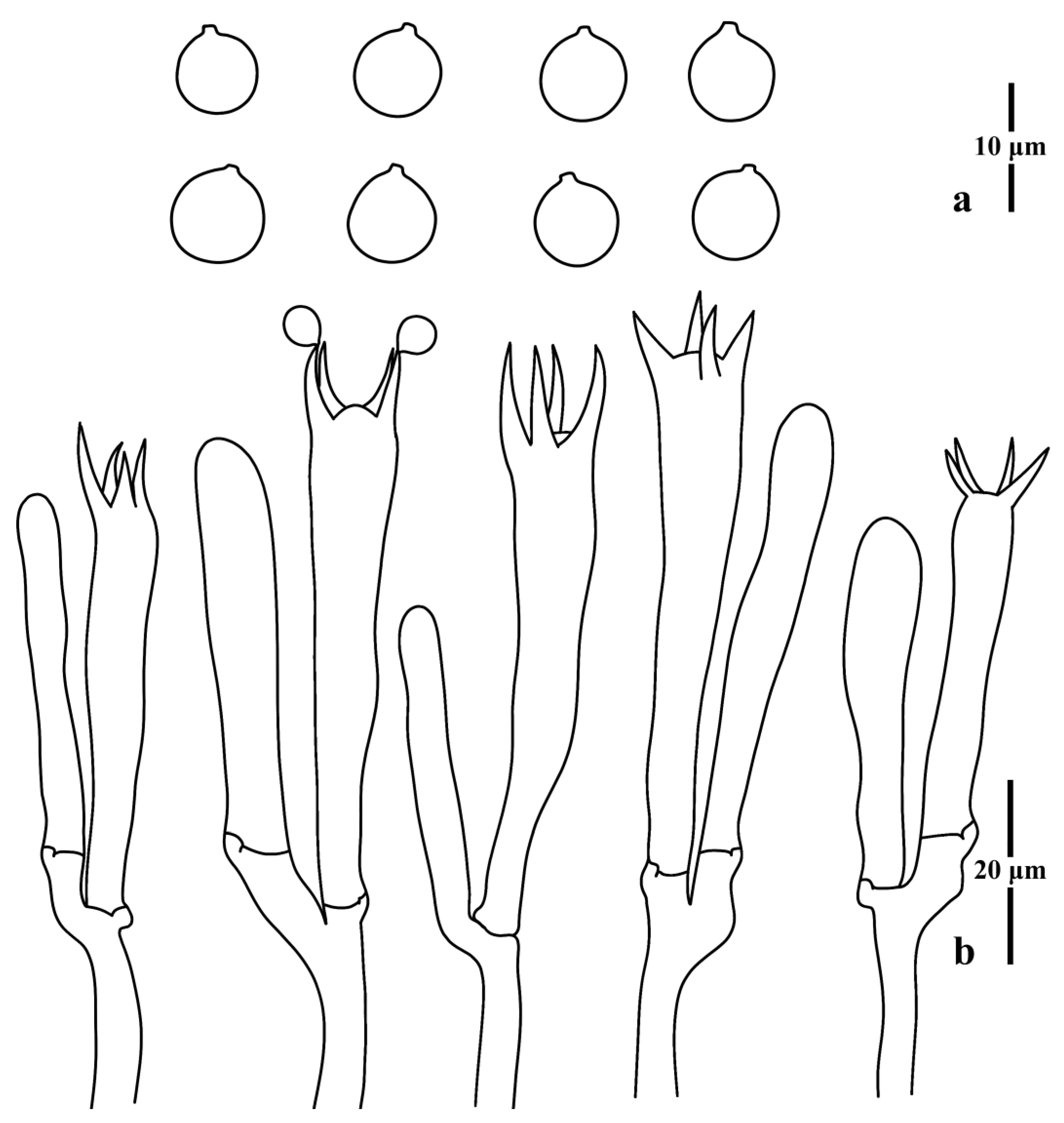{kind=link}
{kind=link}
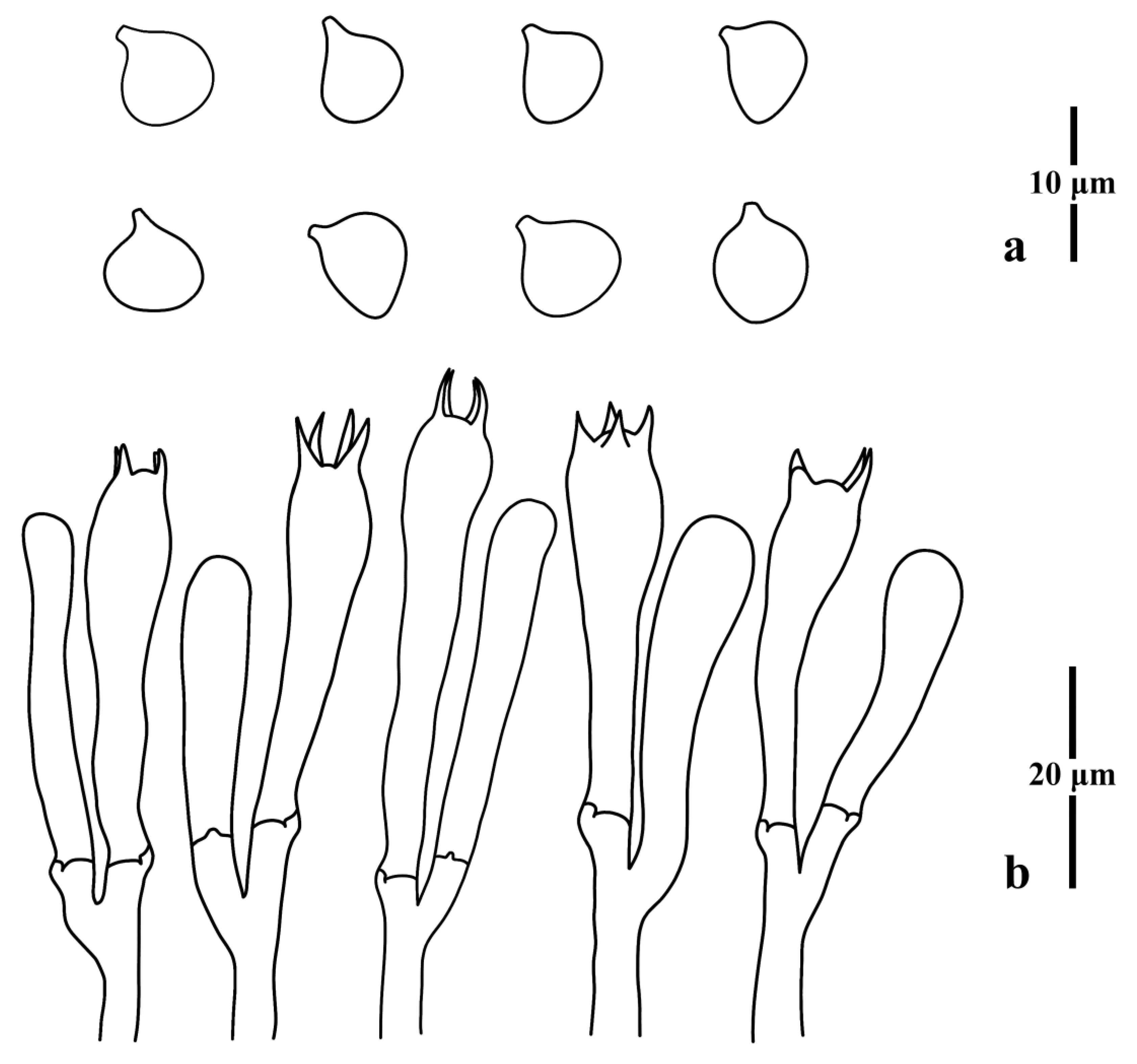{kind=link}
{kind=link}
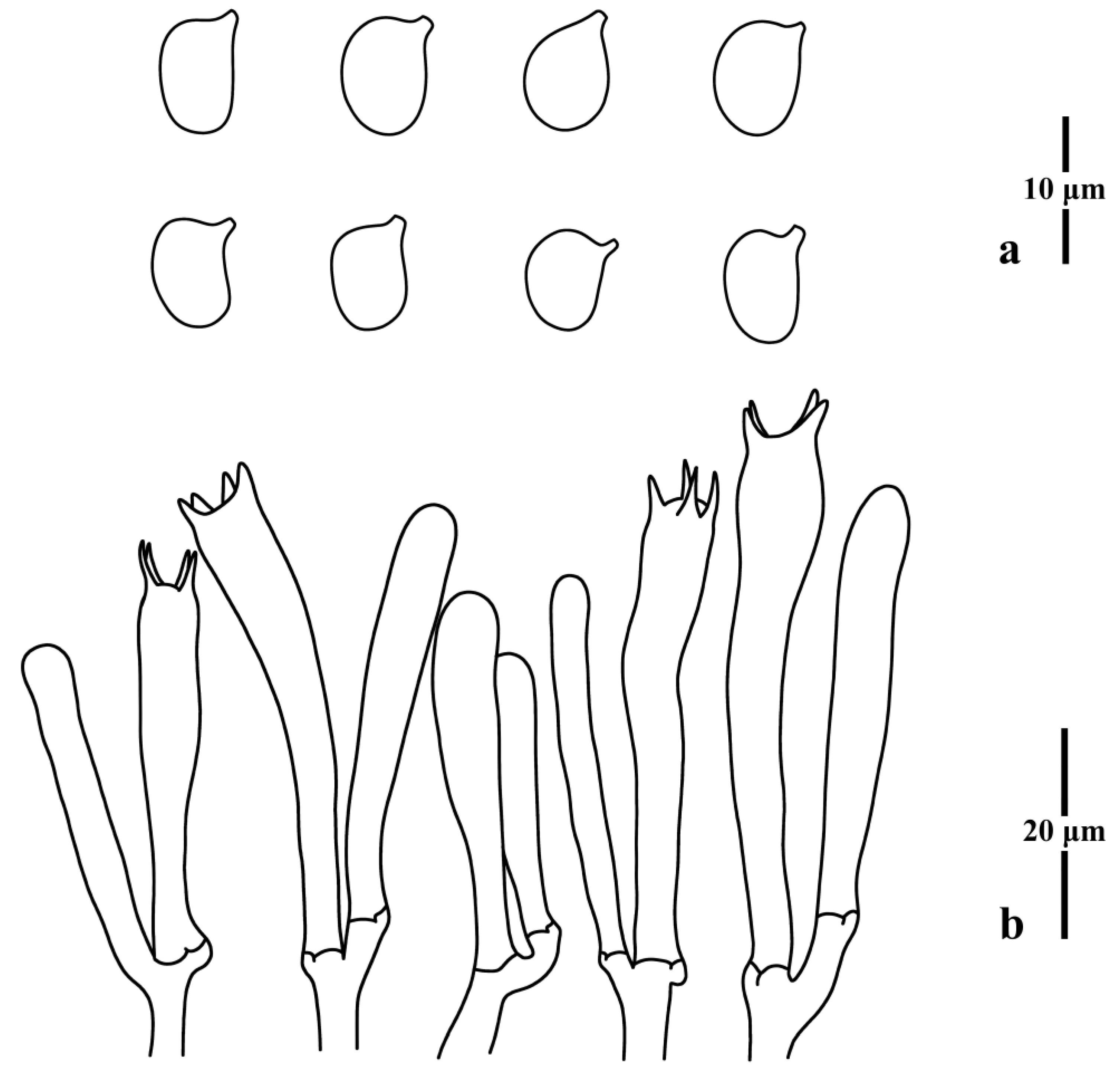{kind=link}
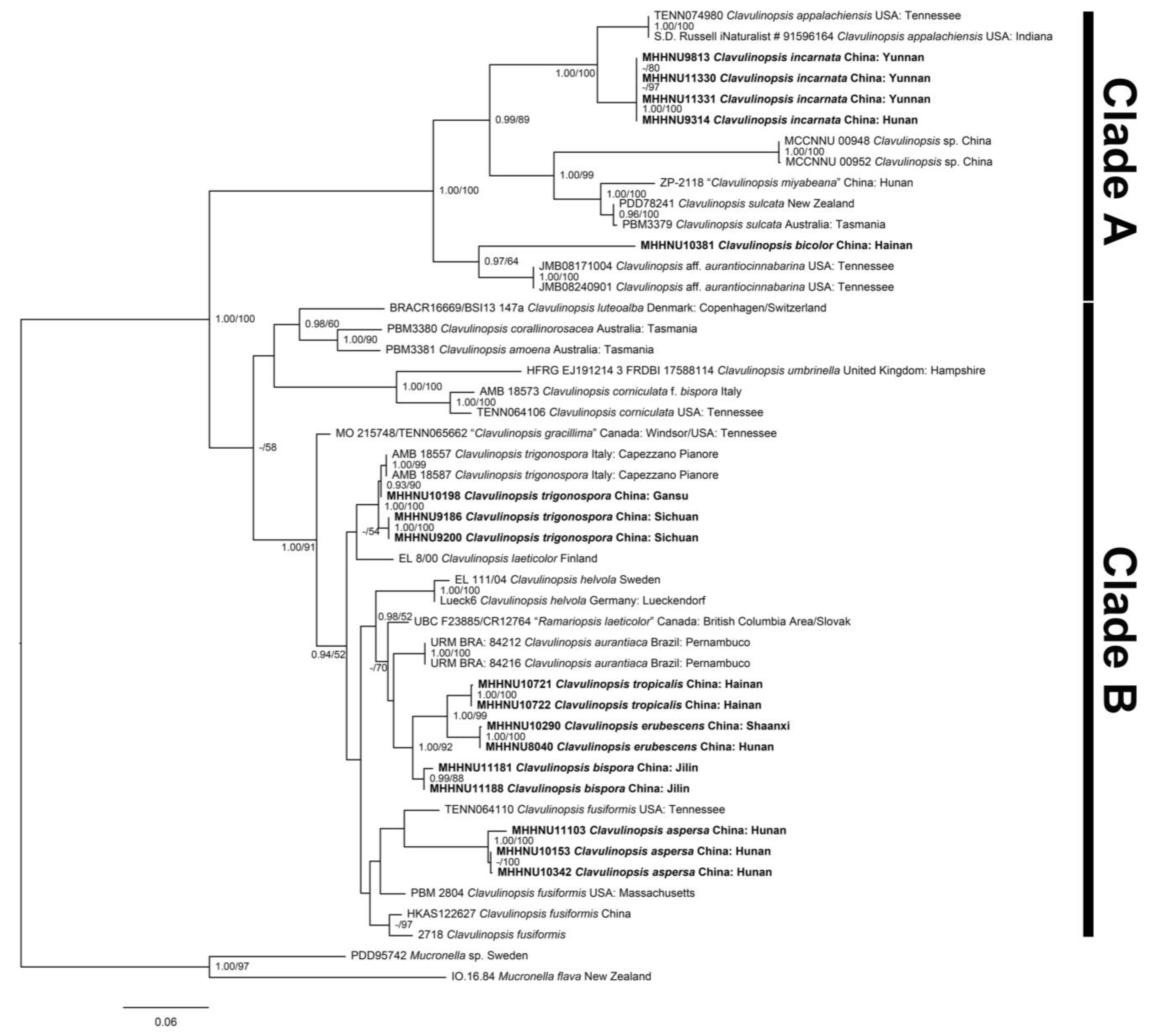{kind=link}
| Identification | Specimen No. | GenBank No. (ITS) | GenBank No. (28S) | Location | References |
|---|---|---|---|---|---|
| Clavulinopsis amoena | PBM3381 | — | HQ877702 | Australia: Tasmania | Birkebak et al. [6]; Hyde et al. [8] |
| C. appalachiensis | S.D. Russell iNaturalist # 91596164 | OM809324 | — | USA: Indiana | Unpublished |
| C. appalachiensis | TENN074980 | MT196965 | — | USA: Tennessee | Unpublished |
| C. aspersa | MHHNU10153 | OQ703777 | OQ703794 | China: Hunan | Present study |
| C. aspersa | MHHNU10342 | OQ703778 | OQ703795 | China: Hunan | Present study |
| C. aspersa | MHHNU11103 | OQ703779 | OQ703796 | China: Hunan | Present study |
| C. aurantiaca | URM <BRA>: 84212 | — | KX227749 | Brazil: Pernambuco | Hyde et al. [8] |
| C. aurantiaca | URM <BRA>: 84216 | KC348464 | NG058946 | Brazil: Pernambuco | Hyde et al. [8] |
| C. aff. aurantiocinnabarina | JMB08171004 | — | HQ877704 | USA: Tennessee | Birkebak et al. [6]; Hyde et al. [8] |
| C. aff. aurantiocinnabarina | JMB08240901 | — | HQ877703 | USA: Tennessee | Birkebak et al. [6]; Hyde et al. [8] |
| C. bicolor | MHHNU10381 | OQ703780 | OQ703797 | China: Hainan | Present study |
| C. bispora | MHHNU11181 | OQ703781 | OQ703798 | China: Jilin | Present study |
| C. bispora | MHHNU11188 | OQ703782 | OQ703799 | China: Jilin | Present study |
| C. corallinorosacea | PBM3380 | KP257144 | HQ877707 | Australia: Tasmania | Birkebak et al. [6,23]; Hyde et al. [8] |
| C. corniculata | TENN064106 | KP257145 | HQ877713 | USA: Tennessee | Birkebak et al. [6,23]; Hyde et al. [8] |
| C. corniculata f. bispora | AMB 18573 | MT055953 | — | Italy | Franchi and M. Marchetti [9] |
| C. erubescens | MHHNU8040 | OQ703783 | OQ703800 | China: Hunan | Present study |
| C. erubescens | MHHNU10290 | OQ703784 | OQ703801 | China: Shaanxi | Present study |
| C. fusiformis | HKAS122627 | ON794403 | — | China | Wang et al. [24] |
| C. fusiformis | PBM 2804 | — | EF535273 | USA: Massachusetts | Birkebak et al. [6]; Hyde et al. [8] |
| C. fusiformis | TENN064110 | — | HQ877717 | USA: Tennessee | Birkebak et al. [6]; Hyde et al. [8] |
| C. fusiformis | 2728 | — | KM248914 | — | Unpublished |
| C. gracillima | MO 215748 | KY706170 | — | Canada: Windsor | Hay et al. [25] |
| C. gracillima | TENN065662 | — | HQ877708 | USA: Tennessee | Unpublished |
| C. helvola | EL 111/04 | EU118617 | EU118617 | Sweden | Birkebak et al. [6]; Hyde et al. [8] |
| C. helvola | Lueck6 | KP965770 | KP965788 | Germany: Lueckendorf | Karich et al. [26] |
| C. incarnata | MHHNU9314 | OQ703785 | OQ703802 | China: Hunan | Present study |
| C. incarnata | MHHNU9813 | OQ703786 | OQ703803 | China: Yunnan | Present study |
| C. incarnata | MHHNU11330 | OQ703787 | OQ703804 | China: Yunnan | Present study |
| C. incarnata | MHHNU11331 | OQ703788 | OQ703805 | China: Yunnan | Present study |
| C. laeticolor | EL 8/00 | EU118618 | EU118618 | Finland | Birkebak et al. [6]; Hyde et al. [8] |
| C. luteoalba | BRACR16669 | — | JQ415959 | Denmark: Copenhagen | Hyde et al. [8] |
| C. luteoalba | BSI13 147a | OP538704 | — | Switzerland | Unpublished |
| C. miyabeana | ZP-2118 | MK427059 | — | China: Hunan | Chen and Zhang, P. [27] |
| C. sp. | MCCNNU 00948 | MT587808 | MT587810 | China | Unpublished |
| C. sp. | MCCNNU 00952 | MT587809 | MT587811 | China | Unpublished |
| C. sulcata | PBM3379 | — | HQ877709 | Australia: Tasmania | Birkebak et al. [6]; Hyde et al. [8] |
| C. sulcata | PDD78241 | — | DQ284904 | New Zealand | Dentinger, B.T.M. and McLaughlin, D.J. [28] |
| C. trigonospora | AMB: 18557 | NR176720 | NG088120 | Italy: Capezzano Pianore | Franchi and M. Marchetti [9] |
| C. trigonospora | AMB: 18587 | — | MT055968 | Italy: Capezzano Pianore | Franchi and M. Marchetti [9] |
| C. trigonospora | MHHNU9186 | OQ703789 | OQ703806 | China: Sichuan | Present study |
| C. trigonospora | MHHNU9200 | OQ703790 | OQ703807 | China: Sichuan | Present study |
| C. trigonospora | MHHNU10198 | OQ703791 | OQ703808 | China: Gansu | Present study |
| C. tropicalis | MHHNU10721 | OQ703792 | OQ703809 | China: Hainan | Present study |
| C. tropicalis | MHHNU10722 | OQ703793 | OQ703810 | China: Hainan | Present study |
| C. umbrinella | HFRG_EJ191214_3_FRDBI 17588114 | OQ133539 | OQ133591 | United Kingdom: Hampshire | Unpublished |
| Ramariopsis laeticolor | CR12764 | — | GU299509 | Slovakia | Unpublished |
| R. laeticolor | UBC F23885 | KJ146701 | — | Canada: British Columbia Area | Unpublished |
| Mucronella flava | IO.16.84 | MT232354 | MT232307 | Sweden | Olariaga et al. [29]; Yan et al. [30] |
| Mucronella sp. | PDD95742 | HQ533013 | — | New Zealand | Yan et al. [30,31] |
3. Results
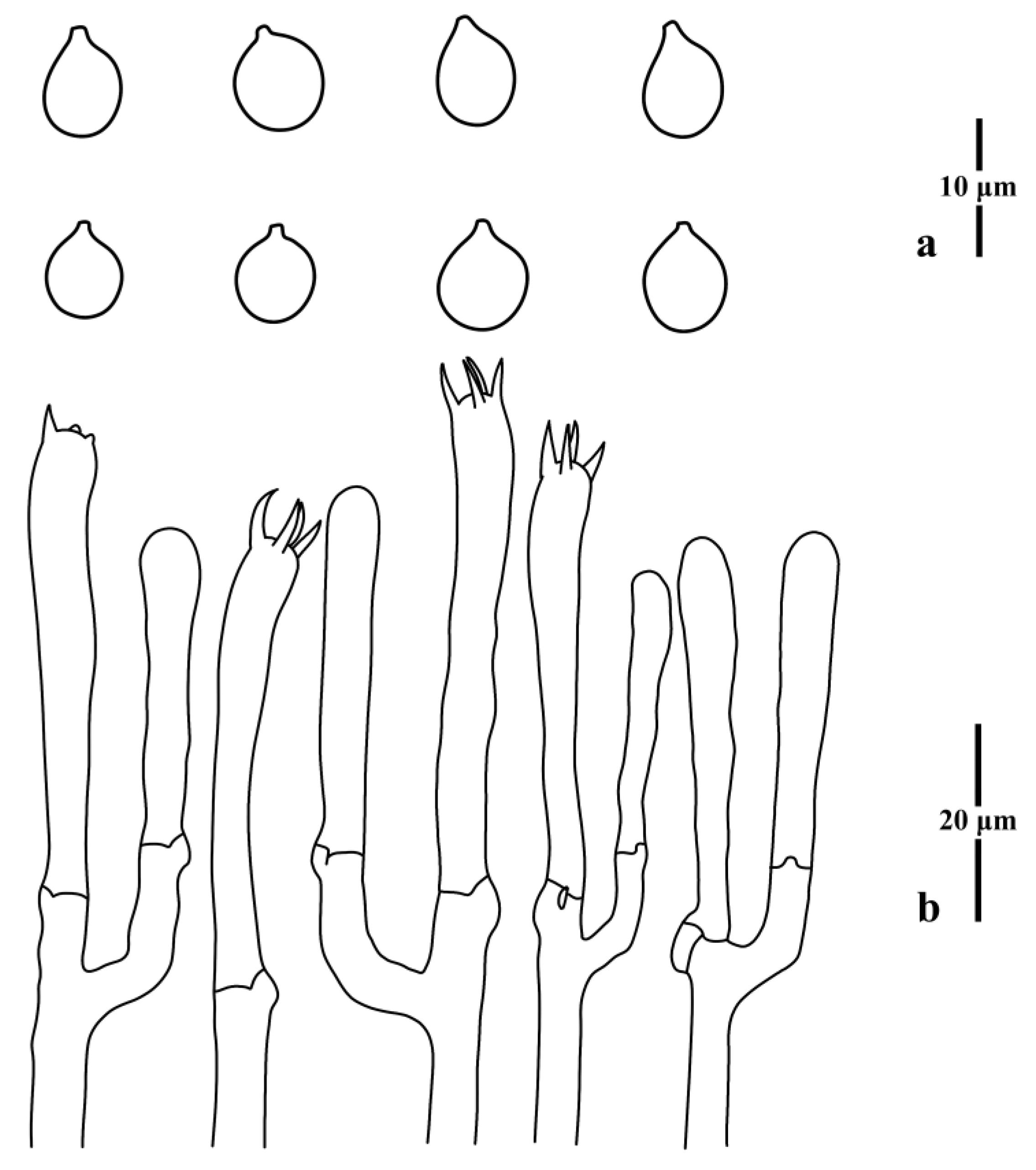
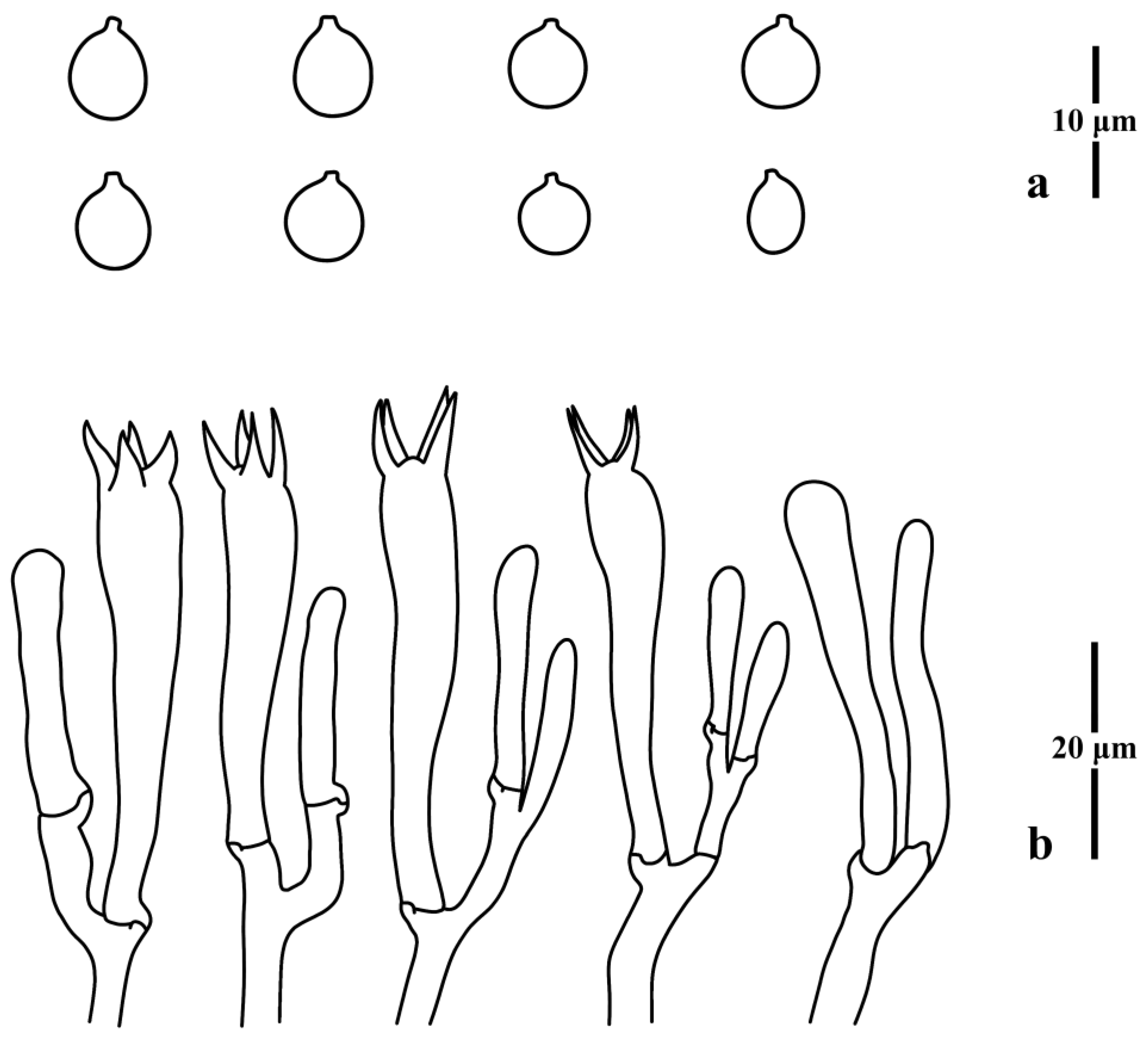
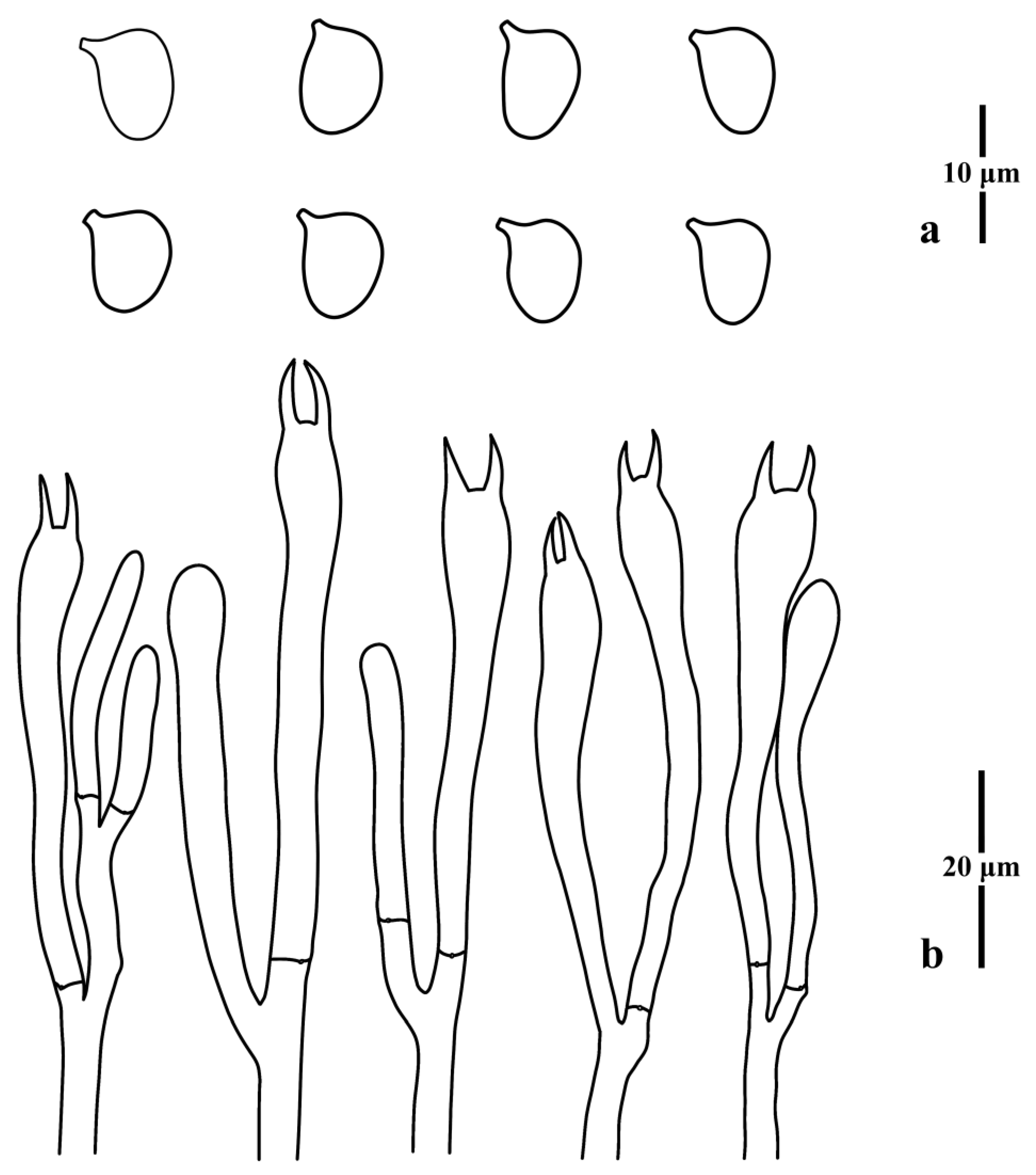
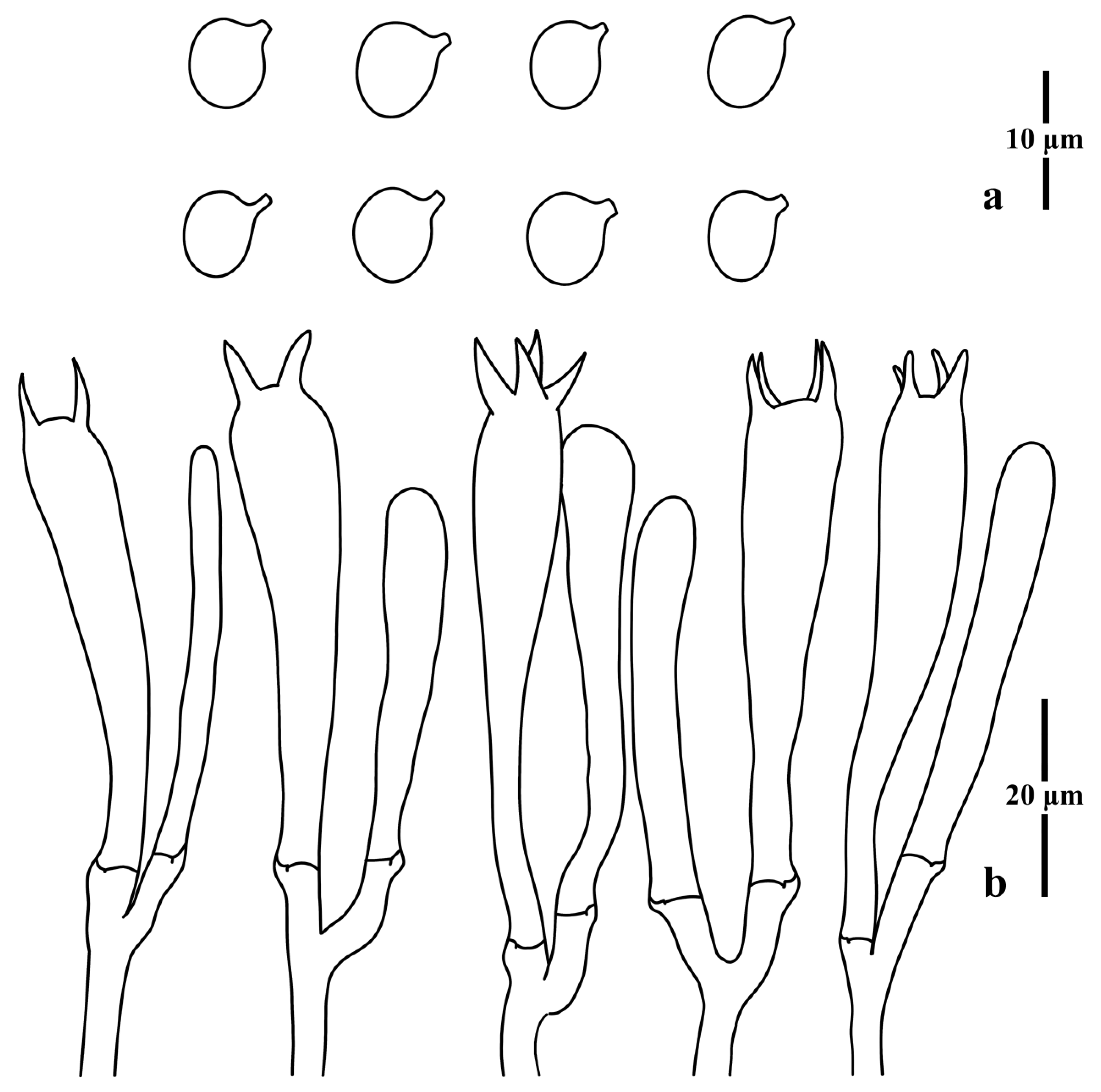
3.1. Taxonomy
3.2. Molecular Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| 1. Basidiomata branched | C. corniculata |
| 1. Basidiomata unbranched | 2 |
| 2. Basidiospores spinous | C. helvola |
| 2. Basidiospores smooth | 3 |
| 3. Basidiospores subtriangular to ellipsoid | C. trigonospora |
| 3. Basidiospores globose, subglobose or ellipsoid | 4 |
| 4. Basidia and hyphae with a very small clamp connection | C. bispora |
| 4. Basidia and hyphae with a normal size clamp connection | 5 |
| 5. Basidiomata bicolored | C. bicolor |
| 5. Basidiomata monochromatic | 6 |
| 6. Basidiomata red or pinkish | 7 |
| 6. Basidiomata white, yellowish or orange | 11 |
| 7. Basidiomata pinkish | C. incarnata |
| 7. Basidiomata red | 8 |
| 8. Basidia two or four tapered sterigmata | C. erubescens |
| 8. Basidia invariably four tapered sterigmata | 9 |
| 9. Basidiospores globose to subglobose | C. sulcata |
| 9. Basidiospores ellipsoid, fusiform or amygdaliform | 10 |
| 10. Basidiomata 15–35 mm tall, basidiospores broadly ellipsoid to ellipsoid | C. tropicalis |
| 10. Basidiomata mostly 60–90 mm tall, basidiospores rather fusiform or amygdaliform | C. corallinorosacea |
| 11. Basidiomata white or cream color | 12 |
| 11. Basidiomata yellowish or orange | 13 |
| 12. Basidiospores broadly pip-shaped, the wall slightly thickened | C. brevipes |
| 12. Basidiospores subglobose, thin-walled | C. spiralis |
| 13. Basidiomata orange, orange-red or salmon-orange | 14 |
| 13. Basidiomata yellowish | 15 |
| 14. Basidiomata 15–30 mm tall, basidiospores globose, rarely subglobose | C. aurantiaca |
| 14. Basidiomata 15–70mm tall, basidiospores subglobose | C. aurantiocinnabarina |
| 15. Distributed in the tropics | C. amoena |
| 15. Distributed in the temperate and subtropics | 16 |
| 16. Basidiospore hilar appendix not prominent (<1 μm) | C. luteoalba |
| 16. Basidiospore hilar appendix very prominent (>1 μm) | 17 |
| 17. Tramal hyphae somewhat thick-walled (up to 1 μm) | C. laeticolor |
| 17. Tramal hyphae thin-walled | 18 |
| 18. Basidiomata 50–140 mm tall, densely fasciculate | C. fusiformis |
| 18. Basidiomata 15–50 mm tall, solitary or scattered to gregarious | C. aspersa |
References
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungi-like taxa. Mycosphere 2020, 11, 1160–1456. [Google Scholar] [CrossRef]
- Corner, E.J.H. A Monograph of Clavaria and Allied Genera; Oxford University Press: London, UK, 1950; pp. 1–740. [Google Scholar]
- Corner, E.J.H. Supplement to “A monograph of Clavaria and allied genera”. Beih. Zur Nova Hedwig. 1970, 33, 1–299. [Google Scholar]
- Petersen, R.H. Notes on Clavarioid Fungi. XV. Reorganization of Clavaria, Clavulinopsis and Ramariopsis. Mycologia 1978, 70, 660–671. [Google Scholar] [CrossRef]
- Pegler, D.N.; Young, T.W.K. Basidiospore structure in Ramariopsis (Clavariaceae). Trans. Br. Mycol. Soc. 1985, 84, 207–436. [Google Scholar] [CrossRef]
- Birkebak, J.M.; Mayor, J.R.; Ryberg, K.M.; Matheny, P.B. A systematic, morphological and ecological overview of the Clavariaceae (Agaricales). Mycologia 2013, 105, 896–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furtado, A.N.M.; Daniëls, P.P.; Neves, M.A. New species and new records of Clavariaceae (Agaricales) from Brazil. Phytotaxa 2016, 253, 1–26. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.; Jones, E.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Franchi, P.; Marchetti, M. I Funghi Clavarioidi in Italia; A. M. B. Fondazione Centro studi Micologici: Vicenza, Italy, 2021; Volume 1, pp. 1–664. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Wu, C.L.; He, Y.; Yan, J.; Zhang, P. Two new species of Clavulina (Cantharellales) from southwestern China based on morphological and molecular evidence. Mycol. Prog. 2019, 18, 1071–1078. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Fyre Methuen: London, UK, 1978; pp. 1–252. [Google Scholar]
- Ridgway, R. Color Standards and Color Nomenclature; Published by the author: Washington, DC, USA, 1912; pp. 1–252. [Google Scholar]
- Katoh, K.; Standley, D.M. A simple method to control over-alignment in the MAFFT multiple sequence alignment program. Bioinformatics 2016, 32, 1933–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Vaidya, G.; Lohman, D.J.; Meier, R. Sequencematrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihoodbased phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre: Uppsala, Sweden, 2004. [Google Scholar]
- Rambaut, A. FigTree v1. 4; University of Edinburgh: Edinburgh, UK, 2012; Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 5 April 2023).
- Birkebak, J.M.; Adamčík, S.; Looney, B.P.; Matheny, P.B. Multilocus phylogenetic reconstruction of the Clavariaceae (Agaricales) reveals polyphyly of agaricoid members. Mycologia 2016, 108, 860–868. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Herrera, M.; Xu, W.J.; Moreno, J.P.; Colinas, C. Ethnomycological study on wild mushrooms in Pu’er Prefecture, Southwest Yunnan, China. J. Ethnobiol. Ethnomed. 2022, 18, 55. [Google Scholar] [CrossRef]
- Hay, C.R.J.; Thorn, R.G.; Jocobs, C.R. Taxonomic survey of Agaricomycetes (Fungi: Basidiomycota) in Ontario tallgrass prairies determined by fruiting body and soil rDNA sampling. Can. Field-Nat. 2018, 132, 4. [Google Scholar] [CrossRef]
- Karich, A.; Kellner, H.; Schmidt, M.; Ullrich, R. Ein bemerkenswertes Mykotop im Zittauer Gebirge mit Microglossum rufescens als Erstnachweis für Deutschland. Boletus 2015, 36, 151–163. [Google Scholar]
- Chen, Z.H.; Zhang, P. Atlas of Macrofungi in Hunan; Hunan Normal University Press: Changsha, China, 2019; pp. 1–425. [Google Scholar]
- Dentinger, B.T.M.; McLaughlin, D.J. Reconstructing the Clavariaceae using nuclear large subunit rDNA sequences and a new genus segregated from Clavaria. Mycologia 2006, 98, 746–762. [Google Scholar] [CrossRef] [PubMed]
- Olariaga, I.; Huhtinen, S.; Læssøe, T.; Petersen, J.H.; Hansen, K. Phylogenetic origins and family classification of typhuloid fungi, with emphasis on Ceratellopsis, Macrotyphula and Typhula (Basidiomycota). Stud. Mycol. 2020, 96, 155–184. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Li, G.W.; Liu, W.H.; Chen, Z.H.; Zhang, P. Updated taxonomy of Chinese Clavaria subg. Syncoryne (Clavariaceae, Agaricales): Description of two new species and one newly recorded species. Mycol. Prog. 2022, 21, 67. [Google Scholar] [CrossRef]
- Yan, J.; Wang, X.Y.; Wang, X.H.; Chen, Z.H.; Zhang, P. Two new species of Clavaria (Agaricales, Basidiomycota) from Central China. Phytotaxa 2020, 477, 71–80. [Google Scholar] [CrossRef]
- Petersen, R.H. The Clavarioid Fungi of New Zealand; DSIR Science Information Publishing: Wellington, New Zealand, 1988; pp. 1–170. [Google Scholar]
- Oberwinkler, F. Fungus-alga interactions in basidiolichens. Beih. Zur Nova Hedwig. 1984, 79, 739–774. [Google Scholar]















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, J.; Wen, J.; Li, G.-W.; Wu, S.-W.; Zhang, P. Taxonomy and Phylogenetic Relationships of Clavulinopsis (Clavariaceae, Agaricales): Description of Six New Species and One Newly Recorded Species from China. J. Fungi 2023, 9, 656. https://doi.org/10.3390/jof9060656
Yan J, Wen J, Li G-W, Wu S-W, Zhang P. Taxonomy and Phylogenetic Relationships of Clavulinopsis (Clavariaceae, Agaricales): Description of Six New Species and One Newly Recorded Species from China. Journal of Fungi. 2023; 9(6):656. https://doi.org/10.3390/jof9060656
Chicago/Turabian StyleYan, Jun, Jing Wen, Gui-Wu Li, Shao-Wu Wu, and Ping Zhang. 2023. "Taxonomy and Phylogenetic Relationships of Clavulinopsis (Clavariaceae, Agaricales): Description of Six New Species and One Newly Recorded Species from China" Journal of Fungi 9, no. 6: 656. https://doi.org/10.3390/jof9060656