
Several Characteristics of Oidiodendron maius G.L. Barron Important for Heather Plants’ Controlled Mycorrhization
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Oidiodendron maius F3860 Strain as an Object of Investigation
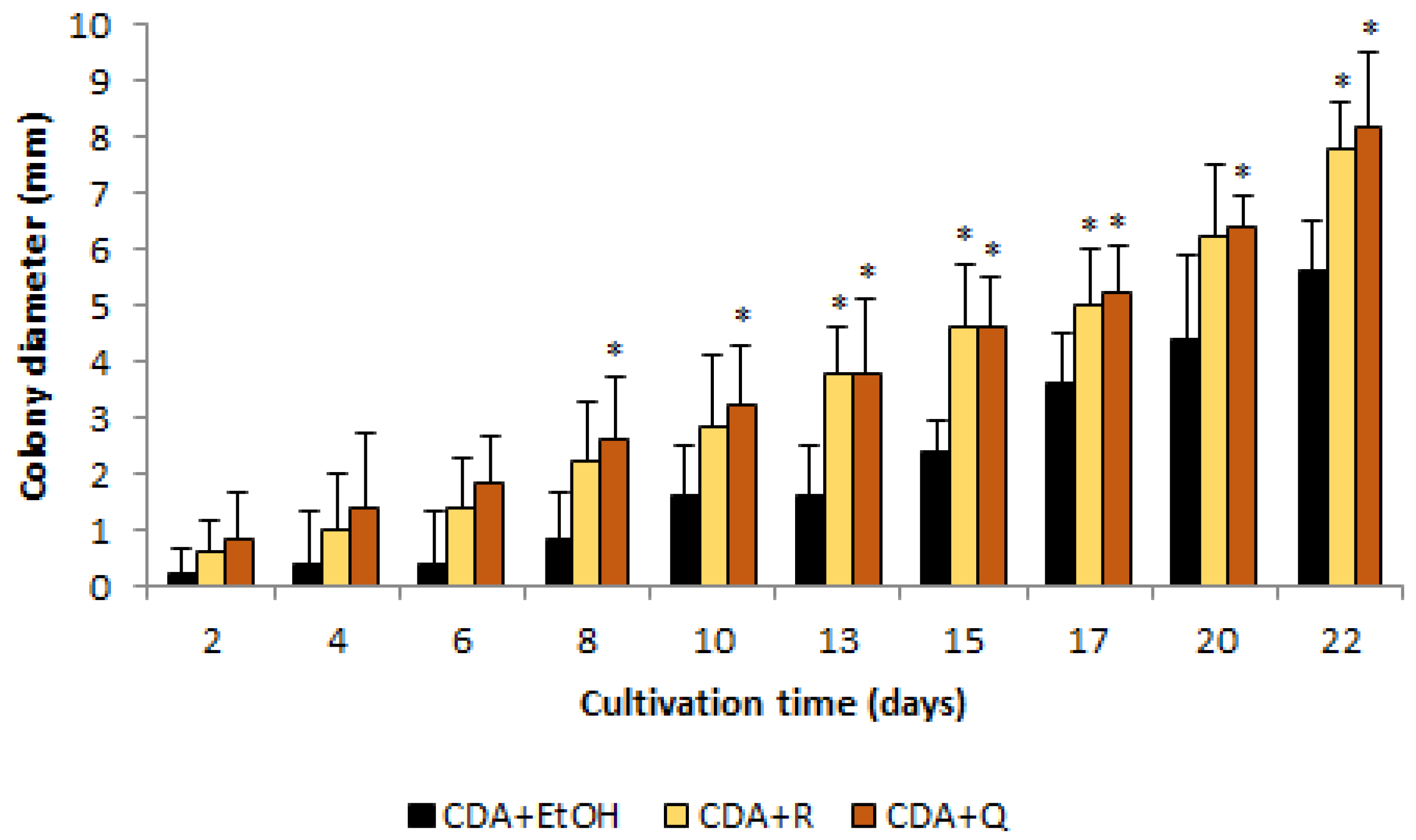
2.2. Influence of Nutrient Media Composition on O. maius F3860 Colonies Growth
2.3. Identification of O. maius F3860 Extracellular Enzyme Activities
2.4. Bioassay for Identification of O. maius F3860 Ability to Synthesize Auxins
2.5. Determination of Indole Acetic Acid or Analogues with Salkowski Reagent
2.6. Statistics
3. Results
3.1. Influence of Nutrient Media Composition on O. maius F3860 Colonies’ Growth
3.2. Identification of O. maius F3860 Extracellular Enzyme Activities
3.3. Bioassay for Identification of O. maius F3860 Ability to Synthesize Auxins
3.4. Determination of Indole Acetic Acid or Analogues with Salkowski Reagent
4. Discussion
4.1. Influence of Nutrient Media Composition on O. maius F3860 Colonies Growth
4.2. Identification of O. maius F3860 Extracellular Enzyme Activities
4.3. Assessment of the O. maius F3860 Strain’s Ability to Synthesize IAA or Analogues
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rice, A.V.; Currah, R.S. Oidiodendron Maius: Saprobe in Sphagnum Peat, Mutualist in Ericaceous Roots? Microbial Roots Endophytes; Schulz, B., Boyle, C., Sieber, T.N., Eds.; Springer-Verlag: Berlin, Germany, 2006; pp. 227–246. [Google Scholar]
- Van der Heijden, M.G.A.; Martin, F.M.; Selosse, M.; Sanders, I.R. Mycorrhizal Ecology and Evolution: The past, the present and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Chen, J.; Zhang, C.; Pan, D. A New Oidiodendron maius Strain Isolated from Rhododendron fortunei and its Effects on Nitrogen Uptake and Plant Growth. Front. Microbiol. 2016, 7, 1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikheev, V.S.; Struchkova, I.V.; Ageyeva, M.N.; Brilkina, A.A.; Berezina, E.V. The role of Phialocephala fortinii in improving plants’ phosphorus nutrition: New puzzle pieces. J. Fungi 2022, 8, 1225. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhang, W.; Zulfiqar, F.; Zhang, C.; Chen, J. Ericoid mycorrhizal fungi as biostimulants for improving propagation and production of ericaceous plants. Front. Plant Sci. 2022, 13, 1027390. [Google Scholar] [CrossRef] [PubMed]
- Martino, E.; Turnau, K.; Girlanda, M.; Bonfante, P.; Perotto, S. Ericoid mycorrhizal fungi from heavy metal polluted soils: Their identification and growth in the presence of zinc ions. Mycol. Res. 2000, 104, 338–344. [Google Scholar] [CrossRef]
- Casarrubia, S.; Daghino, S.; Kohler, A.; Morin, E.; Khouja, H.-R.; Daguerre, Y.; Veneault-Fourrey, C.; Martin, F.M.; Perotto, S.; Martino, E. The Hydrophobin-Like OmSSP1 May Be an Effector in the Ericoid Mycorrhizal Symbiosis. Front. Plant Sci. 2018, 9, 546. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Zhao, X.; Liu, C.; Liu, C.; Bai, L.; Zhao, M.; Li, L. Diversity and characteristics of colonization of root-associated fungi of Vaccinium uliginosum. Sci. Rep. 2018, 8, 15283. [Google Scholar] [CrossRef] [Green Version]
- Pescie, M.A.; Montecchia, M.; Lavado, R.S.; Chiocchio, V.M. Inoculation with Oidiodendron maius BP Improves Nitrogen Absorption from Fertilizer and Growth of Vaccinium corymbosum during the Early Nursery Stage. Plants 2023, 12, 792. [Google Scholar] [CrossRef]
- Xiao, G.; Berch, S. Organic nitrogen use by salal ericoidmycorrhizal fungi from northern Vancouver Island and impacts on growth in vitro of Gaultheria shallon. Mycorrhiza 1999, 9, 145–149. [Google Scholar] [CrossRef]
- Vohník, M.; Albrechtová, J.; Vosátka, M. The inoculation with Oidiodendron maius and Phialocephala fortinii alters phosphorus and nitrogen uptake, foliar C: N ratio and root biomass distribution in Rhododendron cv. Azurro. Symbiosis 2005, 40, 87–96. [Google Scholar]
- Daguerre, Y.; Levati, E.; Ruytinx, J.; Tisserant, E.; Morin, E.; Kohler, A.; Montanini, B.; Ottonello, S.; Brun, A.; Veneault-Fourrey, C.; et al. Regulatory networks underlying mycorrhizal development delineated by genome-wide expression profiling and functional analysis of the transcription factor repertoire of the plant symbiotic fungus Laccaria bicolor. BMC Genom. 2017, 18, 737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, E.; Morin, E.; Grelet, G.A.; Kuo, A.; Kohler, A.; Daghino, S.; Barry, K.W.; Cichocki, N.; Clum, A.; Dockter, R.B.; et al. Comparative genomics and transcriptomics depict ericoid mycorrhizal fungi as versatile saprotrophs and plant mutualists. New Phytol. 2018, 217, 1213–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, E.; Murat, C.; Vallino, M.; Bena, A.; Perotto, S.; Spanu, P. Imaging mycorrhizal fungal transformants that express EGFP during ericoid endosymbiosis. Curr. Genet. 2007, 52, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Abbà, S.; Khouja, H.R.; Martino, E.; Archer, D.B.; Perotto, S. SOD1-targeted gene disruption in the ericoid mycorrhizal fungus Oidiodendron maius reduces conidiation and the capacity for mycorrhization. Mol. Plant Microbe Interact. 2009, 22, 1412–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khouja, H.R.; Daghino, S.; Abba, S.; Boutaraa, F.; Chalot, M.; Blaudez, D.; Martino, E.; Perotto, S. OmGOGAT-disruption in the ericoid mycorrhizal fungus Oidiodendron maius induces reorganization of the N pathway and reduces tolerance to heavy-metals. Fungal Genet. Biol. 2014, 71, 1–8. [Google Scholar] [CrossRef]
- Bergero, R.; Perotto, S.; Girlanda, M.; Vidano, G.; Luppi, M.A. Ericoid mycorrhizal fungi are common root associates of a Mediterranean ectomycorrhizal plant (Quercus ilex). Mol. Ecol. 2000, 9, 1639–1649. [Google Scholar] [CrossRef]
- Casarrubia, S.; Sapienza, S.; Fritz, H.; Daghino, S.; Rosenkranz, M.; Schnitzler, J.P.; Martin, F.; Perotto, S.; Martino, E. Ecologically Different Fungi Affect Arabidopsis Development: Contribution of Soluble and Volatile Compounds. PLoS ONE 2016, 14, e0168236. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Griffin, A.; Kernaghan, G. Ericoid mycorrhizal colonization and associated fungal communities along a wetland gradient in the Acadian forest of Eastern Canada. Fungal Ecol. 2022, 56, 101138. [Google Scholar] [CrossRef]
- Malysheva, E.F.; Malysheva, V.F.; Voronina, E.Y.; Kovalenko, A.E. Diversity of fungal communities associated with mixotrophic pyroloids (Pyrola rotundifolia, P. media and Orthilia secunda) in their natural habitats. Bot. Pacifica. J. Plant Sci. Conserv. 2018, 7, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Hambleton, S.; Egger, K.N.; Currah, R.S. The Genus Oidiodendron: Species Delimitation and Phylogenetic Relationships Based on Nuclear Ribosomal DNA Analysis. Mycologia 1998, 90, 854–868. [Google Scholar] [CrossRef]
- Netrusov, A.I.; Egorova, M.A.; Zakharchuk, L.M. Practicum on Microbiology: A Textbook for Students of Higher Educational Institutions; Academia: Moscow, Russia, 2005; 608p. (In Russian) [Google Scholar]
- Schuchovski, C.S.; Biasi, L.A. In vitro establishment of ‘Delite’ rabbiteye blueberry microshoots. Horticulturae 2019, 5, 24. [Google Scholar] [CrossRef] [Green Version]
- Methods of Experimental Mycology; Bilai, V.I. (Ed.) Nauk. Dumka: Kiev, Ukraine, 1982; 552p. (In Russian) [Google Scholar]
- Chernavina, I.A.; Potapov, N.G.; Kosulina, L.G.; Krendeleva, T.E. A High Practicum on Plant Physiology. Mineral Nutrition. Cell Physiology. Growth and Development: Textbook for Students of Biol. Spec. Universities; High School: Moscow, Russia, 1978; 408p. (In Russian) [Google Scholar]
- Cohen, B.A.; Amsellem, Z.; Maor, R.; Sharon, A.; Gressel, J. Transgenically enhanced expression of indole-3-acetic acid confers hypervirulence to plant pathogens. Phytopathology 2002, 92, 590–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumla, J.; Suwannarach, N.; Matsui, K.; Lumyong, S. Biosynthetic pathway of indole-3-acetic acid in ectomycorrhizal fungi collected from northern Thailand. PLoS ONE 2020, 15, e0227478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.C.; Lin, W.R.; Hsu, Y.C.; Pan, H.Y. Influences of Three Oidiodendron maius Isolates and Two Inorganic Nitrogen Sources on the Growth of Rhododendron kanehirae. Hortic. Sci. Technol. 2020, 38, 742–753. [Google Scholar] [CrossRef]
- Zheng, W.; Lehmann, A.; Ryo, M.; Vályi, K.K.; Rilling, M.C. Growth rate trades off with enzymatic investment in soil filamentous fungi. Sci. Rep. 2020, 10, 11013. [Google Scholar] [CrossRef]
- Mehmood, A.; Hussain, A.; Irshad, M.; Khan, N.; Hamayun, M.; Ismail; Afridi, S.G.; Lee, I.J. IAA and flavonoids modulates the association between maize roots and phytostimulant endophytic Aspergillus fumigatus greenish. J. Plant Interact. 2018, 13, 532–542. [Google Scholar] [CrossRef] [Green Version]
- Usmanov, I.Y.; Yumagulova, E.R.; Aleksandrova, V.V.; Ivanov, S.P.; Shcherbakov, A.V.; Gonchar, I.G.; Ivanov, V.B. Fractal analysis of flavonoids in complex chemical compositions in extracts of Chamaedaphne calyculata (L.) Moench (ericaceae) in oligotrophic swamps of Western Siberia. Mod. Phytomorphol. 2019, 13, 35–40. [Google Scholar] [CrossRef]
- Berezina, E.V.; Brilkina, A.A.; Veselov, A.P. Content of phenolic compounds, ascorbic acid, and photosynthetic pigments in Vaccinium macrocarpon Ait. dependent on seasonal plant development stages and age (the example of introduction in Russia). Sci. Hortic. 2017, 218, 139–146. [Google Scholar] [CrossRef]
- Ştefănescu, B.E.; Szabo, K.; Mocan, A.; Crişan, G. Phenolic compounds from five Ericaceae species leaves and their related bioavailability and health benefits. Molecules 2019, 24, 2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shishodia, S.K.; Tiwari, S.; Hoda, S.; Vijayaraghavan, P.; Shankar, J. SEM and qRT-PCR revealed quercetin inhibits morphogenesis of Aspergillus flavus conidia via modulating calcineurin-Crz1 signalling pathway. Mycology 2020, 11, 118–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Peng, X.; Lin, J.; Zhang, Y.; Zhang, J.; Gao, H.; Tian, X.; Zhang, R.; Zhao, G. Quercetin ameliorates Aspergillus fumigatus keratitis by inhibiting fungal growth, toll-like receptors and inflammatory cytokines. Int. Immunopharmacol. 2021, 93, 107435. [Google Scholar] [CrossRef] [PubMed]
- Tempesti, T.C.; Alvarez, M.G.; de Araújo, M.F.; Catunda Júnior, F.E.A.; de Carvalho, M.G.; Durantini, E.N. Antifungal activity of a novel quercetin derivative bearing a trifluoromethyl group on Candida albicans. Med. Chem. Res. 2012, 21, 2217–2222. [Google Scholar] [CrossRef]
- Negahdari, R.; Bohlouli, S.; Sharifi, S.; Maleki Dizaj, S.; Rahbar Saadat, Y.; Khezri, K.; Raeesi, S. Therapeutic benefits of rutin and its nanoformulations. Phytother. Res. 2021, 35, 1719–1738. [Google Scholar] [CrossRef]
- Roy, S.; Nuckles, E.; Archbold, D.D. Effects of phenolic compounds on growth of Colletotrichum spp. in vitro. Curr. Microbiol. 2018, 75, 550–556. [Google Scholar] [CrossRef]
- Gomes, B.; Castro, F.; Santos, R.; Figueiredo, P.; Silva, M.; Vidal, M.; Ferreira, I.; Nunes, J.; Machado, H.; Gomes, F. Effect of quercetin on mycorrhizal synthesis between Tuberborchii and Arbutusunedo L. in vitro plants. Microbiol. Res. 2021, 12, 69–81. [Google Scholar] [CrossRef]
- Ngonkeu, E.L.M.; Nwaga, D.; Tsetchoua, R.; Mogo, A.; Mongoue, A.F.; Mballa, T.A.N.; Boyomo, O. Effect of Quercetin and Pectinase on Spore Germination and Hyphal Growth of Arbuscular Mycorrhizal Fungi and Biomass Production of Nicotiana tabacum L. and Calopogonium mucunoides desv. Annu. Res. Rev. Biol. 2017, 12, 1–11. [Google Scholar] [CrossRef]
- Scervino, J.M.; Ponce, M.A.; Erra-Bassells, R.; Vierheilig, H.; Ocampo, J.A.; Godeas, A. Arbuscular mycorrhizal colonization of tomato by Gigaspora and Glomus species in the presence of root flavonoids. J. Plant Physiol. 2005, 162, 625–633. [Google Scholar] [CrossRef]
- Leonard, W.; Zhang, P.; Ying, D.; Adhikari, B.; Fang, Z. Fermentation transforms the phenolic profiles and bioactivities of plant-based foods. Biotechnol. Adv. 2021, 49, 107763. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Thangavelu, B.; Chun, S.C.; Sathiyabama, M. Proteases from phytopathogenic fungi and their importance in phytopathogenicity. J. General. Plant Pathol. 2016, 82, 233–239. [Google Scholar] [CrossRef]
- Westrick, N.M.; Smith, D.L.; Kabbage, M. Disarming the host: Detoxification of plant defense compounds during fungal necrotrophy. Front. Plant Sci. 2021, 12, 651716. [Google Scholar] [CrossRef] [PubMed]
- Bizabani, C.; Dames, J. Effects of inoculating Lachnum and Cadophora isolates on the growth of Vaccinium corymbosum. Microbiol. Res. 2015, 181, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Rice, A.V.; Currah, R.S. Oidiodendron: A survey of the named species and related anamorphs of Myxotrichum. Stud. Mycol. 2005, 53, 83–120. [Google Scholar] [CrossRef] [Green Version]
- Wurzburger, N.; Higgins, B.P.; Hendrick, R.L. Ericoid mycorrhizal root fungi and their multicopper oxidases from a temperate forest shrub. Ecol. Evol. 2012, 2, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Prigione, V.; Spina, F.; Tigini, V.; Giovando, S.; Varese, G.C. Biotransformation of industrial tannins by filamentous fungi. Appl. Microbiol. Biotechnol. 2018, 102, 10361–10375. [Google Scholar] [CrossRef]
- Martino, E.; Perotto, S.; Parsons, R.; Gadd, G.M. Solubilization of insoluble inorganic zinc compounds by ericoid mycorrhizal fungi derived from heavy metal polluted sites. Soil. Biol. Biochem. 2003, 35, 133–141. [Google Scholar] [CrossRef]
- Neetu, N.; Sharma, M.; Mahto, J.K.; Kumar, P. Biophysical and in-silico studies of phytochemicals targeting chorismate synthase from drug-resistant Moraxella catarrhalis. Protein J. 2020, 39, 449–460. [Google Scholar] [CrossRef]
- Liu, X.; Han, R.; Cao, Y.; Turner, B.L.; Ma, L.Q. Enhancing Phytate Availability in Soils and Phytate-P Acquisition by Plants: A Review. Environ. Sci. Technol. 2022, 56, 9196–9219. [Google Scholar] [CrossRef]
- Berezina, E.V.; Brilkina, A.A.; Veselov, A.P. The content of phenolic compounds in Vaccinium vitis-idaea and Oxycoccus palustris (Ericaceae) during different vegetation periods. Rastit. Resur. 2015, 51, 88–100. (In Russian) [Google Scholar]
- Perotto, S.; Daghino, S.; Martino, E. Ericoid mycorrhizal fungi and their genomes: Another side to the mycorrhizal symbiosis? New Phytol. 2018, 220, 1141–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, B.R.; Mitchell, D.T. Phosphatases of ericoid mycorrhizal fungi: Kinetic properties and the effect of copper on activity. Mycol. Res. 2005, 109, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Della Monica, I.F.; Mario Saparrat, M.; Godeas, A.M.; Scervino, J.M. The co-existence between DSE and AMF symbionts affects plant P pools through P mineralization and solubilization processes. Fungal Ecol. 2015, 17, 10–17. [Google Scholar] [CrossRef]
- Fu, S.-F.; Wei, J.-Y.; Chen, H.-W.; Liu, Y.-Y.; Lu, H.-Y.; Chou, J.-Y. Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant Signal Behav. 2015, 10, 17–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.Y.; Zhang, F.; Zhang, D.J.; Srivastava, A.K.; Wu, Q.S.; Zou, Y.N. Mycorrhiza stimulates root-hair growth and IAA synthesis and transport in trifoliate orange under drought stress. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Johnston-Monje, D.; Raizada, M.N. Plant and Endophyte relationships: Nutrient management. Compr. Biotechnol. 2011, 713–727. [Google Scholar]
- Morffy, N.; Strader, L.C. Old Town Roads: Routes of auxin biosynthesis across kingdoms. Curr. Opin. Plant Biol. 2020, 55, 21–27. [Google Scholar] [CrossRef]
- Wei, X.; Chen, J.; Zhang, C.; Liu, H.; Zheng, X.; Mu, J. Ericoid mycorrhizal fungus enhances microcutting rooting of Rhododendron fortunei and subsequent growth. Hortic. Res. 2020, 7, 140. [Google Scholar] [CrossRef]
- Junaidi, A.R.; Bolhassan, M.H. Screening of Indole-3-Acetic Acid (IAA) Productions by Endophytic Fusarium oxysporum Isolated from Phyllanthus niruri. Borneo J. Resour. Sci. Technol. 2017, 7, 56–59. [Google Scholar] [CrossRef] [Green Version]
- Mehmood, A.; Irshad, M.; Husna; Ahmad, A.; Hussain, A. In Vitro Maize Growth Promotion by Endophytic Fusarium oxysporum WLW. J. Agricult. Biol. Environ. Stat. 2018, 8, 30–35. [Google Scholar]
- Numponsak, T.; Kumla, J.; Suwannarach, N.; Matsui, K.; Lumyong, S. Biosynthetic pathway and optimal conditions for the production of indole-3-acetic acid by an endophytic fungus, Colletotrichum fructicola CMU-A109. PLoS ONE 2018, 13, e0205070. [Google Scholar] [CrossRef] [PubMed]
- Brilkina, A.A.; Melnikova, N.E.; Berezina, E.V.; Veselov, A.P. Exogenous auxins usage for rhizogenesis activation during adaptation of highbush blueberry microplants. Vest. Nizhegorodskaya Selskokhozyaistvennaya Acad. 2019, 1, 4–8. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient Media | KD | Reliability Parameter for Linear Approximation R² |
|---|---|---|
| CDA | 0.4661 | 0.9741 |
| And | 0.7230 | 0.9900 |
| WPM | 0.6612 | 0.9942 |
| MEA | 0.9624 | 0.9942 |
| PDA | 1.0878 | 0.9943 |
| CDA + EtOH | 0.2638 | 0.9249 |
| CDA + R | 0.3445 | 0.9811 |
| CDA + Q | 0.3382 | 0.9704 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikheev, V.S.; Struchkova, I.V.; Churkina, L.M.; Brilkina, A.A.; Berezina, E.V. Several Characteristics of Oidiodendron maius G.L. Barron Important for Heather Plants’ Controlled Mycorrhization. J. Fungi 2023, 9, 728. https://doi.org/10.3390/jof9070728
Mikheev VS, Struchkova IV, Churkina LM, Brilkina AA, Berezina EV. Several Characteristics of Oidiodendron maius G.L. Barron Important for Heather Plants’ Controlled Mycorrhization. Journal of Fungi. 2023; 9(7):728. https://doi.org/10.3390/jof9070728
Chicago/Turabian StyleMikheev, Vyacheslav S., Irina V. Struchkova, Ludmila M. Churkina, Anna A. Brilkina, and Ekaterina V. Berezina. 2023. "Several Characteristics of Oidiodendron maius G.L. Barron Important for Heather Plants’ Controlled Mycorrhization" Journal of Fungi 9, no. 7: 728. https://doi.org/10.3390/jof9070728