
Transcriptomic Response of Clonostachys rosea Mycoparasitizing Rhizoctonia solani


1
School of Light Industry, Beijing Technology and Business University, Beijing 100048, China
2
Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193, China
*
Author to whom correspondence should be addressed.
J. Fungi 2023, 9(8), 818; https://doi.org/10.3390/jof9080818
Submission received: 18 July 2023
/
Revised: 28 July 2023
/
Accepted: 1 August 2023
/
Published: 2 August 2023
Abstract
:Clonostachys rosea is an important mycoparasitism biocontrol agent that exhibits excellent control efficacy against numerous fungal plant pathogens. Transcriptomic sequencing may be used to preliminarily screen mycoparasitism-related genes of C. rosea against fungal pathogens. The present study sequenced and analyzed the transcriptome of C. rosea mycoparasitizing a Basidiomycota (phylum) fungal pathogen, Rhizoctonia solani, under three touch stages: the pre-touch stage, touch stage and after-touch stage. The results showed that a number of genes were differentially expressed during C. rosea mycoparasitization of R. solani. At the pre-touch stage, 154 and 315 genes were up- and down-regulated, respectively. At the touch stage, the numbers of up- and down-regulated differentially expressed genes (DEGs) were 163 and 188, respectively. The after-touch stage obtained the highest number of DEGs, with 412 and 326 DEGs being up- and down-regulated, respectively. Among these DEGs, ABC transporter-, glucanase- and chitinase-encoding genes were selected as potential mycoparasitic genes according to a phylogenetic analysis. A comparative transcriptomic analysis between C. rosea mycoparasitizing R. solani and Sclerotinia sclerotiorum showed that several DEGs, including the tartrate transporter, SDR family oxidoreductase, metallophosphoesterase, gluconate 5-dehydrogenase and pyruvate carboxylase, were uniquely expressed in C. rosea mycoparasitizing R. solani. These results significantly expand our knowledge of mycoparasitism-related genes in C. rosea and elucidate the mycoparasitism mechanism of C. rosea.
1. Introduction
Rhizoctonia solani (teleomorph: Thanetophorus cucumeris) is a destructive necrotrophic fungal pathogen that belongs to Basidiomycota, Agaricomycetes, Ceratobasidiales, Ceratobasidiaceae [1]. The hyphal color of R. solani changes from whitish to brown as the hyphae develop from young to mature [2]. The hyphae then form into sclerotia, which are the main infection source of Rhizoctonia solani, and exhibit strong resistance to a hostile environment [3,4,5]. Rhizoctonia solani infects plants belonging to 32 families, such as Malvaceae, Fabaceae, Poaceae, Brassicaceae and Amaranthaceae. Rhizoctonia solani infection causes severe plant diseases and losses [6,7,8].
Chemical control is a widely used method to control plant diseases caused by R. solani. However, the following reasons limit the use of pesticides to some extent: environmental pollution and easily generated pesticide resistance to R. solani. Biocontrol is another effective control method that has attracted much attention due to its advantages of being green, safe and sustainable. Microorganisms such as Trichoderma sp., Clonostachys rosea. and Lecanicillium muscarium are important biocontrol agents [9,10,11]. Biocontrol mycoparasites generally target living fungi that parasitize fungal pathogens as a type of biocontrol agent. Several mycoparasites containing Trichoderma strains (T. asperellum, T. koningii, T. virens, T. harzianum, and T. atroviride) and Clonostachys rosea have exhibited desirable control effects against plant diseases caused by R. solani [10,11,12,13,14,15]. In addition to screening more biocontrol mycoparasites, examining the mycoparasitism molecular mechanism is another an important pathway to further improving control efficacy against plant diseases caused by R. solani.
The screening and identification of mycoparasitism-related genes are the basis for investigating the molecular mechanisms of mycoparasitism. Many methods may be used to acquire mycoparasitism-related genes, such as subtractive hybridization, transcriptomic analysis, cDNA microarray, etc. Among these methods, transcriptomic analysis is a crucial and common method. Investigating alterations in the expression levels of genes from mycoparasites during the process of mycoparasitic R. solani using transcriptomic analysis has helped further examine mycoparasitism-related genes. The screening of mycoparasitism-related genes using transcriptomic analysis has primarily focused on Trichoderma species. Numerous mycoparasitism-related genes have been examined from T. virens and T. atroviride using transcriptomic analysis when these Trichoderma species mycoparasitized R. solani [16,17,18].
Clonostachys rosea is an excellent biocontrol mycoparasite that protects against numerous fungal plant pathogens [19]. Numerous mycoparasitism-related genes have been screened from C. rosea against Sclerotinia sclerotiorum, Fusarium graminearum, Botrytis cinerea and Helminthosporium solani according to transcriptomic sequencing and analysis [20,21,22,23]. However, there are no reports of the screening of mycoparasitism-related genes from C. rosea against R. solani. Because taxonomic differences exist between R. solani and the other four pathogens in the phylum, the types of mycoparasitism-related gene may distinguish C. rosea against R. solani against the other reported four pathogens. Therefore, it is important to examine mycoparasitism-related genes from C. rosea against R. solani.
Clonostachys rosea 67-1 is an effective strain that exhibits desirable inhibition of R. solani and control efficacy against rice sheath blight disease caused by R. solani [24]. To screen mycoparasitism-related genes from C. rosea against R. solani, the present study collected the mycelium of C. rosea against R. solani at three stages: pre-touch, touch and after-touch. We investigated differentially expressed genes from C. rosea during the process of mycoparasitizing R. solani. We compared the transcriptomes of C. rosea against other fungal pathogens and analyzed specific mycoparasitism-related genes in C. rosea against R. solani. The results of the present study significantly expand our knowledge of mycoparasitism-related genes in C. rosea and elucidate the mycoparasitism mechanism of C. rosea.
2. Materials and Methods
2.1. Strains
Clonostachys rosea 67-1 was originally isolated from a vegetable yard in Hainan Province, China. Clonostachys rosea 67-1 and the pathogen R. solani were preserved and cultivated on PDA (potato dextrose agar) plates.
2.2. Sample Collection
Clonostachys rosea 67-1 and R. solani were cultivated on PDA plates at 26 °C for 10 d. Sterilized cellophane was placed on the surface of the PDA plates, and equally sized mycelium plugs of C. rosea 67-1 and R. solani were taken and placed at opposite positions on the PDA plates and cultivated at 26 °C. Three stages were selected for sampling to reflect the mycoparasitism process of C. rosea. The pre-touch stage (PT) occurred when the distance between C. rosea 67-1 and R. solani was 0.5 cm. The touch stage (T) occurred when C. rosea 67-1 and R. solani grew together. The after-touch stage (AT) occurred when C. rosea 67-1 and R. solani had crossed growth of 0.5 cm (Figure S1). The mycelium of C. rosea 67-1 from each treatment was collected and used for RNA extraction. Clonostachys rosea 67-1 growth alone at each stage was used as a control. All of the treatments were performed with three replications.
2.3. cDNA Library Construction and Transcriptome Sequencing
The total RNA of the mycelium in each sample was extracted. An Agilent 2100 bioanalyzer was used to detect the quality of the extracted RNAs. Oligo (dT) magnetic beads were used to enrich the mRNAs of each sample. The enriched mRNAs were randomly broken by divalent cations in NEB fragmentation buffer.
The mRNAs were reverse transcribed into the first-strand cDNA using an M-MuLV reverse transcriptase system, and then, the second-stand cDNA was synthesized using a DNA polymerase I system. The purified cDNA was subjected to end repair, the addition of adenine to the 3′ end and adapter connection to generate 250-300 bp cDNA fragments using AMPure XP beads (Beckman Coulter, High Wycombe, UK). cDNA libraries were constructed via the amplification of suitable cDNA fragments. After quality control, the cDNA libraries were sequenced using the Illumina NovaSeq 6000 platform (Illumina Inc., San Diego, CA, USA) at Novogene Corporation Inc.
2.4. Transcriptome Data Processing
The raw sequencing data contained few reads that had adapters or low sequencing quality. To ensure the reliability of transcriptome data analysis, the raw sequencing data were filtered to acquire clean data as follows. Reads that contained adaptor sequences and that contained N (unknown bases) and low-quality reads were removed. All reads were submitted to the NCBI sequence read archive (SRA) database to obtain accession numbers. The clean sequences were mapped onto the genome of C. rosea 67-1 to obtain their location in the genome using HISAT2 software v2.0.5 [25].
2.5. DEG Analysis
The expression levels of the genes were calculated using FPKM (expected number of fragments per kilobase of transcript sequence per million base pairs sequenced), which corrected the sequencing depth and gene length [26,27]. DESeq2 software (1.20.0) was used to screen differentially expressed genes (DEGs) in C. rosea mycoparasitizing R. solani at three different stages [28]. Genes with significant differential expression were confirmed as having an expression fold change of greater or less than 2 and a p value of less than 0.05. Cluster analysis was performed between all treatments to reflect the relationships of samples or genes. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses of DEGs were performed using clusterProfiler software (3.8.1) based on the hypergeometric distribution principle.
To screen for potential mycoparasitism-related genes, three types of DEG encoding ABC transporters, glucanases and serine proteases from C. rosea were used to perform phylogenetic analysis with corresponding genes with known function from Trichoderma strains. The selected corresponding genes of each type are effective against R. solani (Table 1). Sequences of each type of DEG were aligned using CLUSTAL_X, and phylogenetic analysis was performed using MEGA 6.0 with the maximum likelihood method under the Kimura two-parameter model [29,30].
2.6. Comparative Transcriptome Analysis
Because the taxonomic status of R. solani is quite different from that of other commonly known fungal pathogens, investigating the unique DEGs from C. rosea mycoparasitizing R. solani compared to other fungal pathogens helps further our understanding of the molecular mechanism of C. rosea mycoparasitism. We previously performed transcriptomic analysis of C. rosea mycoparasitizing S. sclerotiorum. Therefore, S. sclerotiorum was used as a typical Ascomycota pathogen for comparative transcriptome analysis in the present study. The unique DEGs and their pathways involved in C. rosea mycoparasitizing R. solani compared to mycoparasitizing S. sclerotiorum were analyzed.
2.7. Quantitative Real-Time PCR Validation
Twelve DEGs were randomly selected and their expression levels were detected through quantitative real-time PCR for validation. The primer pairs of the 12 DEGs were designed using Primer Premier 6.0 software (Table S1).
Quantitative real-time PCR was performed using a CFX96 Real-Time PCR Detection System, using programs of 95 °C for 30 s; 40 cycles of 95 °C for 5 s; and 60 °C for 30 s. The expression levels of the 12 DEGs were calculated using the 2−ΔΔCt method. Elongation factor and actin were used as internal reference genes. Each experiment was performed with three replications.
3. Results
3.1. Transcriptome Sequencing and Data Processing
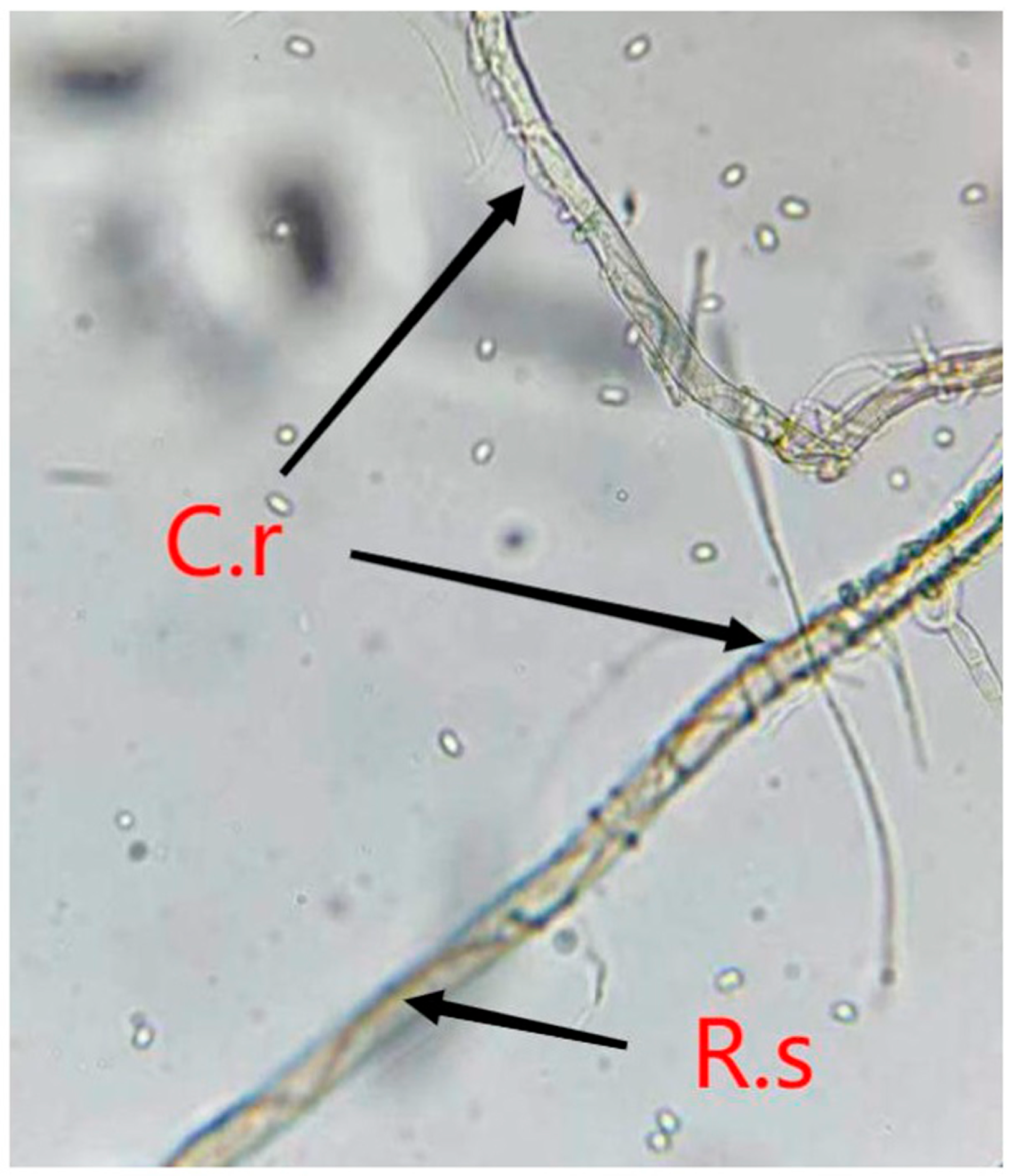
The microscope observation showed that the mycelium of C. rosea was tightly intertwined with R. solani at both the T and AT stages, which indicates that C. rosea was mycoparasitizing R. solani at the time of touch (Figure 1).
After transcriptome sequencing, the Q20 and Q30 of each sample were higher than 96.71% and 91.43%, respectively. The average clean read after data filtration was 45,863,230. The mapping rate to the genome of C. rosea between treatments ranged from 95.01 to 97.33, with an average rate of 96.22%. The sequencing data were submitted to SRA, under the following accession numbers: SRR22822402, SRR23032134, SRR22987521, SRR22981867, SRR22980386, SRR22963970, SRR22953580, SRR22947653, SRR22942454, SRR22937423, SRR22894263, SRR22891490, SRR22891348, SRR22890531, SRR22888634, SRR22876798, SRR22875421 and SRR22859027.
3.2. DEG Analysis
Hundreds of genes were differentially expressed during the process of C. rosea mycoparasitizing R. solani. Most of these DEGs belonged to hypothetical proteins. At the pre-touch stage, 154 and 315 genes were up- and down-regulated, respectively, during C. rosea mycoparasitism. The highest expression levels of up-regulated DEGs were annotated to polyketide synthase, MFS transporter and acetylxylan esterase-like protein. The down-regulated DEGs with the highest expression fold changes were annotated to aldehyde dehydrogenase and SGNH hydrolase-type esterase.
The numbers of downregulated DEGs during the touch stage of C. rosea mycoparasitism were lower than those in the pre-touch stage, with 163 and 188 genes being up- and down-regulated, respectively. Up-regulated DEGs such as D-isomer-specific 2-hydroxyacid dehydrogenase, S-layer homology domain-containing protein, chaperonin 10-like protein and NADPH-p450 reductase had the highest fold changes in expression. Among the downregulated DEGs, arabinogalactan endo-1,4-beta-galactosidase, GCN5-N-acetyltransferase and glycosyl hydrolase family 61-domain-containing protein exhibited higher fold changes in expression.
Among the three mycoparasitism stages, the numbers of DEGs in the after-touch stage were higher than those in the other stages. A total of 412 and 326 DEGs were up- and down-regulated, respectively. Genes annotated to aldehyde dehydrogenase, NAD-dependent epimerase/dehydratase and alpha/beta hydrolase protein exhibited the highest expression fold changes of the up-regulated DEGs. For the down-regulated DEGs, the highest expression fold changes of genes were annotated to cytochrome P450, GMC oxidoreductase and L-amino acid oxidase.
3.3. GO Enrichment Analysis
DEGs from all three mycoparasitic stages were assigned to three categories: biological process, cellular components and molecular function. Most DEGs in the pre-touch stage were annotated to carbohydrate metabolic processes and the regulation of gene expression in the biological process category. DEGs annotated to coenzyme binding and transporter activity were the dominant groups in the molecular function category. Most DEGs in the cellular component category were annotated to the extracellular region (Figure 2A).
The dominant groups in the biological process and molecular function categories in the touch stage were similar to those in the pre-touch stage, carbohydrate metabolic processes and transporter activity. The numbers of DEGs in each group of cellular component categories were equal (Figure 2B). The dominant groups among the three categories in the after-touch stage were quite different from those in the other two mycoparasitic stages. The dominant group in the biological process category was the regulation of gene expression, the regulation of macromolecule metabolic processes and small molecule metabolic processes. Organelle part and intracellular organelle part were the main groups in the cellular components category. The dominant groups in the molecular function category were coenzyme binding, ATP binding and adenyl ribonucleotide binding (Figure 2C).
3.4. KEGG Enrichment Analysis
The dominant KEGG pathways among the three mycoparasitic stages were different. The main pathways in the pre-touch stage were the metabolism of xenobiotics by cytochrome P450, the biosynthesis of nucleotide sugars, and amino sugar and nucleotide sugar metabolism (Figure 3A). The dominant pathways in the touch stage were arachidonic acid metabolism and drug metabolism—other enzymes (Figure 3B). The pathways of ribosome biogenesis in eukaryotes and peroxisomes were dominant in the after-touch stage (Figure 3C).
3.5. Screening of Potential Mycoparasitism-Related Genes
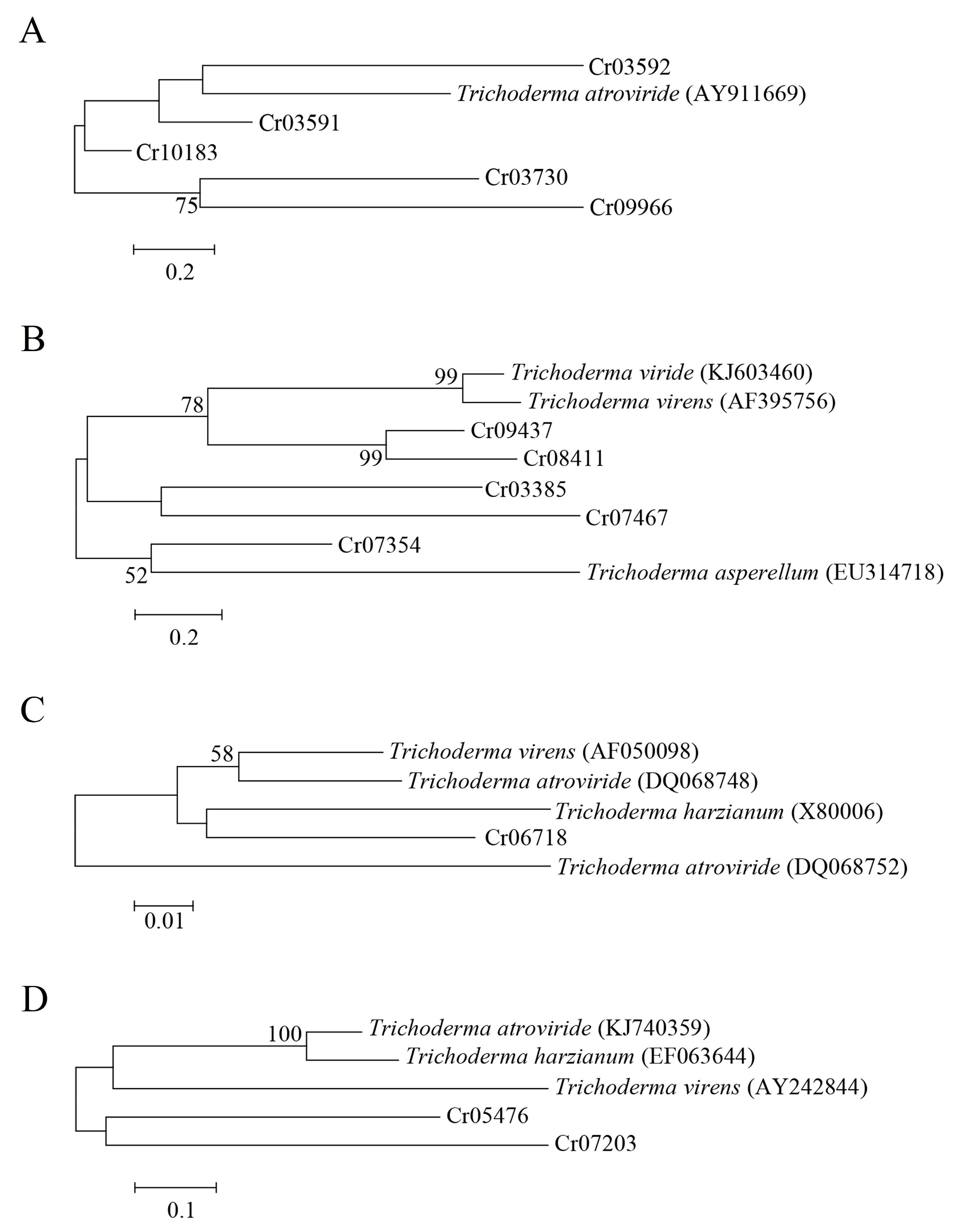
Five ATP-binding cassette (ABC) transporters, five glucanases, two serine proteases and one chitinase were differentially expressed in C. rosea mycoparasitizing R. solani. Phylogenetic analysis showed that the ABC transporter-encoding gene Taabc2 was closely related to Cr03592, which may be a mycoparasitism-related gene compared to other ABC transporter-encoding genes (Figure 4A). For glucanases and chitinase, phylogenetic analysis showed that Cr07354 and Cr06718 were most closely related to exo-β-1,3-glucanase and chitinase, respectively, and may be used as potential mycoparasitism-related genes for further functional verification (Figure 4B,C). The serine protease-encoding genes Cr05476 and Cr07203 from C. rosea were far from all of the serine protease genes from the Trichoderma species according to the phylogenetic analysis (Figure 4D). Therefore, the function of the two serine protease genes in mycoparasitism needs further investigation.
3.6. Comparative Transcriptomic Analysis
Compared to C. rosea mycoparasitizing S. sclerotiorum [20], several DEGs were unique during the process of C. rosea mycoparasitizing R. solani at three stages. At the pre-touch stage, genes encoding tartrate transporter protein, synaptic vesicle transporter and phosphoacetylglucosamine mutase were uniquely expressed. Among these unique DEGs, the gene encoding phosphoacetylglucosamine mutase was involved in the biosynthesis pathways of nucleotide sugars, amino sugar and nucleotide sugar metabolism.
The DEGs encoding formylmethionine deformylase, pyruvate carboxylase and victoriocin were unique at the after-touch stage. The pyruvate carboxylase gene participated in multiple pathways, including the citrate cycle, pyruvate metabolism, the biosynthesis of amino acids and carbon metabolism. Several DEGs, including gluconate 5-dehydrogenase, SDR family oxidoreductase and metallophosphoesterase, were unique at the touch stage. Genes encoding SDR family oxidoreductases were involved in the peroxisome pathway.
3.7. Quantitative Real-Time PCR Validation
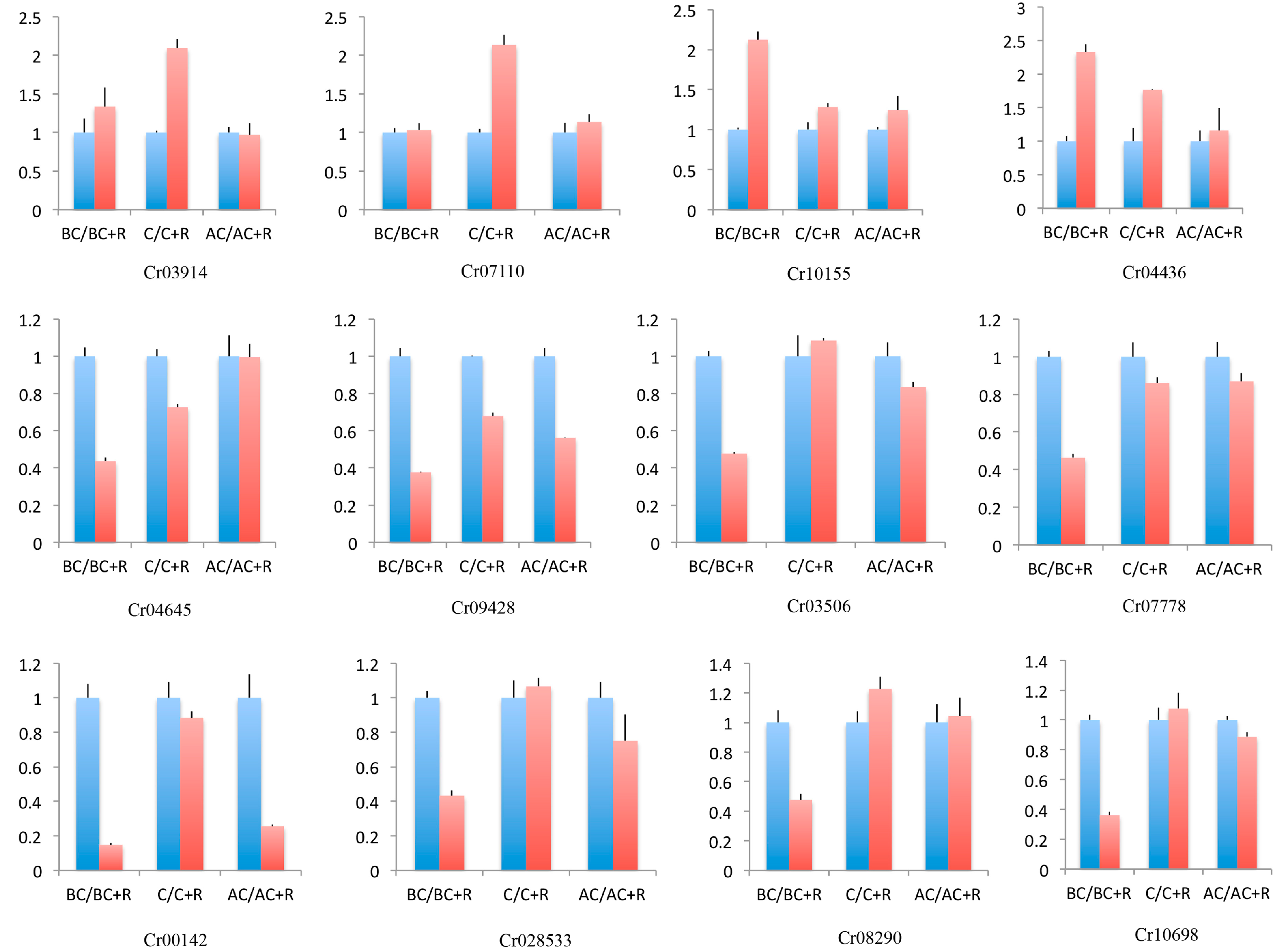
The expression levels of the 12 selected DEGs were consistent with the transcriptomic analysis, which indicates that the expression levels of the DEGs from the transcriptome were reliable (Figure 5).
4. Discussion
The screening of DEGs is the basis for exploring the molecular mechanism of mycoparasitism. Transcriptome sequencing and analysis are commonly used to screen DEGs. Although several fungal plant pathogens, such as S. sclerotiorum, F. graminearum, B. cinerea and H. solani, have been used as target pathogens during C. rosea mycoparasitism to screen for DEGs using transcriptome sequencing, all of these pathogens belong to the phylum Ascomycota. Therefore, screening DEGs from C. rosea mycoparasitizing other taxonomic pathogens is helpful for investigating the molecular mechanism of mycoparasitism to provide new insights. The present study selected R. solani because its taxonomic status is quite different from that of the other reported pathogens. Three stages, pre-touch, touch and after-touch, were used to represent the early, middle and final stages of C. rosea mycoparasitism, respectively. DEGs were screened in three mycoparasitic stages using transcriptomic sequencing and analyzed. The unique DEGs were also analyzed in C. rosea mycoparasitizing R. solani compared to S. sclerotiorum.
Transcriptomic sequencing and analysis revealed that numerous genes were differentially expressed during C. rosea mycoparasitization of R. solani. Among these DEGs, several genes contained ABC transporters, heat shock protein 70, transcription factors, polyketide synthase, cell wall-degrading enzymes such as chitinase, β-glucanase and serine proteases, which were previously reported to have mycoparasitic ability against fungal plant pathogens using gene function analysis [31,32,33,34,35,36]. The present study used ABC transporters, β-glucanase, serine proteases and chitinase as potential mycoparasitic-related genes to perform phylogenetic analysis for the following reasons: there are differences between R. solani and other Ascomycetes fungal plant pathogens, and only a few genes had verified functions as biocontrol agents against R. solani. The above four types of gene were previously reported to be related to R. solani in Trichoderma species [37,38,39,40]. Several genes of each type were differentially expressed during C. rosea mycoparasitization of R. solani in the present study. Therefore, we analyzed which DEGs were closely related to the function-known mycoparasitism genes using phylogenetic analysis, and the potential mycoparasitism genes in C. rosea against R. solani were also screened.
Cell wall-degrading enzymes commonly have biocontrol ability against fungal plant pathogens because these enzymes can degrade the cell walls of fungal pathogens. There are three different types of cell wall-degrading enzyme: chitinase, glucanase and protease. Clonostachys rosea has been reported to be able to produce the above three types of cell wall-degrading enzyme [41]. Pasqualetti found that C. rosea IG119 could produce chitinolytic enzymes [42,43]. The activities of chitinase produced by C. rosea 67-1 were enhanced after overexpression of the chitinase-encoding gene [34]. Moreover, the activities of β-1,3-glucanase produced by C. rosea were induced under the presence of fungal pathogen cell walls [44]. Serine protease was purified from C. rosea and exhibited the ability to degrade the nematode cuticle [45]. The deletion or overexpression of genes encoding cell wall-degrading enzymes reduces or increases the biocontrol ability of biocontrol agents. Five glucanase genes, two serine protease genes and one chitinase gene were differentially expressed in C. rosea 67-1 mycoparasitizing R. solani. Genes encoding chitinases, glucanases and serine proteases are involved in Trichoderma against R. solani. Co-expression of the β-1,3- and β-1,6-glucanase genes in T. virens improved the bioprotection of cotton seedlings against R. solani [38]. The expression of the β-1,3-glucanase gene from Trichoderma spp. in indica and japonica rice cultivars reduced the disease severity of R. solani [46]. The disruption and overexpression of chitinase-encoding gene cht42 could significantly decrease and enhance the biocontrol ability of T. virens against cotton seedling diseases caused by R. solani compared with the wild strain [47]. Phylogenetic analysis revealed that one β-glucanase gene, Cr07354, and one chitinase gene, Cr06718, were most closely related to the reported genes, respectively, which indicates that Cr07354 and Cr06718 in C. rosea 67-1 have potential against R. solani. However, both serine protease genes were far from the reported genes in the phylogenetic analysis, and their function in mycoparasitism needs further exploration.
ABC transporters function in detoxication [48]. The deletion and complementation of the ABC transporter-encoding gene abcG5 in C. rosea IK726 influenced xenobiotic tolerance, which may be due to abcG5’s involvement in detoxifying the Fusarium mycotoxin zearalenone and other fungicides [49]. ABC transporters also play crucial roles in biocontrol. Deletion of the ABC transporter-encoding gene Taabc2 in T. atroviride strongly reduced the ability to overgrow, inhibit and degrade the mycelia of R. solani. The mutant significantly reduced the protective effect of tomato plants infected with R. solani [37]. Five ABC transporter genes were used to perform phylogenetic analysis, and only one ABC transporter gene, Cr03592, was most closely related to the reported gene, which suggests that Cr03592 was a mycoparasitism-related gene against R. solani.
Additionally, genes associated with up-stream signal transduction, such as as G protein; transcribing, such as transcription factor; and down-stream mycoparasitism-related metabolite synthesis, such as polyketide synthase, were all differentially expressed during C. rosea mycoparasitizing R. solani. As an important signal transduction protein, G protein could regulate the transduction of mycoparasitism-related signals through signaling pathways, and then, influence the control ability of biocontrol agents. G protein was involved in fungal growth, development and secondary metabolism, as well as virulence to the host. Several studies have reported that G protein plays important roles in biocontrol among different biocontrol agents. The disruption of the G protein α subunit-encoding gene thga3 in T. harzianum could dramatically affect its mycoparasitic ability against R. solani [50]. In T. atroviride, the deletion of the G protein α subunit-encoding gene tga1 could lose its mycoparasitic ability against R. solani, B. cinerea, and S. sclerotiorum, as well as reducing chitinase activity in the mutant. Moreover, overexpressed tga1 in T. atroviride could improve the mycoparasitic ability against R. solani [51,52]. The disruption of another G protein α subunit-encoding gene, tga3, in T. atroviride could influence its mycoparasitic ability against R. solani and B. cinerea, infection structures and chitinase activity [53]. In other Trichoderma species, G protein α subunit had similar effect on mycoparasitic ability. The deletion of the G protein α subunit-encoding gene tgaA in T. virens could influence the mycoparasitic ability to S. rolfsii [54]. Besides Trichoderma, the deletion of the G protein α subunit-encoding gene MrGPA1 in Metarhizium robertsii could reduce the virulence in Galleria mellonella larvae [55].
Transcription factors are trans-acting proteins that can be activated by signaling pathways, thereby regulating the expression levels of target genes. Transcription factors are highly conserved in fungus and associated with numerous biological processes such as fungal growth, development and tolerance to environmental stress. Transcription factors have been reported to be involved in biocontrol in both mycoparasitism and entomopathogenic fungal agents. In biocontrol mycoparasites, deletion of transcription factor-encoding genes in C. rosea could reduce the virulence and mycoparasitic ability to nematode and S. sclerotiorum, respectively [33,56]. In T. virens, the disruption of the transcription factor-encoding gene pacC could influence the mycoparasitic capacity against R. solani and S. rolfsii [57]. Similarly, the roles of transcription factors in biocontrol have been found in Coniothyrium minitans, and disruption of the transcription factor-encoding gene could affect the mycoparasitic ability against S. sclerotiorum, as well as the activities of chitinase and β-1,3-glucanase [58]. In entomopathogenic fungal agents, the absence of the transcription factor-encoding genes MaPacC and Mrap1 could influence the virulence of M. acridum and M. rileyi in Locusta migratoria manilensis and Spodoptera litura larvae [59,60]. The disruption of the transcription factor-encoding genes Bbmcm1 and BbHapX in Beauveria bassiana could affect the virulence against G. mellonella larvae [61,62].
Polyketide synthases were able to synthesize different types of polyketide, among which antibiotics and toxins had the potential to act against fungal plant pathogens. Polyketide synthase-encoding genes have been reported to be involved in biocontrol. The deletion of the polyketide synthase-encoding gene pks29 in C. rosea could influence the antagonistic ability against B. cinerea and biocontrol capacity against fusarium foot rot in barley. The absence of another polyketide synthase-encoding gene, pks22, in C. rosea could influence the production of Clonorosein A and B, which exhibit antifungal activity against F. graminearum and B. cinerea [35]. The deletion of the polyketide synthase-encoding gene pks4 in T. reesei could influence the antagonistic abilities against Rhizoctonia solani, Sclerotinia sclerotiorum, and Alternaria alternate [63]. Similarly, the disruption of the polyketide synthase-encoding gene could affect the virulence of B. bassiana against beet armyworms [64].
Comparative transcriptomic analysis revealed that several DEGs were unique at three stages during C. rosea mycoparasitizing R. solani compared to S. sclerotiorum. Although studies of these DEGs that participate in the biocontrol process are lacking, these genes were differentially and uniquely expressed during C. rosea mycoparasitization of R. solani. Therefore, exploring the function of these unique DEGs in C. rosea mycoparasitism is important for extending the mycoparasitism-related gene pool of C. rosea. Future work should focus on investigating the function of these DEGs during C. rosea mycoparasitization of R. solani using gene knockout, complementation or overexpression.
5. Conclusions
Transcriptomic sequencing and analysis were performed on C. rosea mycoparasitizing R. solani under three touch stages. A total of 154 and 315 genes, 163 and 188 genes, and 412 and 326 genes were up- and down-regulated at the pre-touch stage, touch stage, and after-touch stage, respectively. ABC transporter-, glucanase- and chitinase-encoding genes were screened as potential mycoparasitism genes during C. rosea against R. solani using phylogenetic analysis. DEGs such as SDR family oxidoreductase, metallophosphoesterase and tartrate transporter were unique in C. rosea mycoparasitizing R. solani compared to that mycoparasitizing S. sclerotiorum according to our comparative transcriptomic analysis. These results expand the types of mycoparasitism-related gene in C. rosea and elucidate the mycoparasitism mechanism of C. rosea.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof9080818/s1, Figure S1: Photos of Clonostachys rosea mycoparasitizing Rhizoctonia solani on plates; Table S1: Primers used for quantitative real-time PCR.
Author Contributions
Conceptualization, Z.-B.S.; methodology, S.-F.Y., M.-H.S. and S.-D.L.; formal analysis, Y.-F.H. and H.-J.S.; writing—original draft preparation, Z.-B.S.; writing—review and editing, Z.-B.S.; funding acquisition, Z.-B.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Science and Technology general project of the Beijing Municipal Education Commission (KM202110011002) and the National Key Research and Development Program of China (2019YFD1002000).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gonzalez, M.; Pujol, M.; Metraux, J.P.; Gonzalez-Garcia, V.; Bolton, M.D.; Borrás-Hidalgo, O. Tobacco leaf spot and root rot caused by Rhizoctonia solani Kühn. Mol. Plant Pathol. 2011, 12, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Sarkar, M.; Chowdhury, A.; Rakwal, R.; Agrawal, G.K.; Sarkar, A. A comprehensive insight into the biology of Rhizoctonia solani AG1-IA Kühn, the causal organism of the sheath blight disease of rice. J. Plant Pathol. 2022, 104, 79–98. [Google Scholar] [CrossRef]
- Abbas, A.; Khan, S.U.; Khan, W.U.; Saleh, T.A.; Khan, M.H.U.; Ullah, S.; Ali, A.; Ikram, M. Antagonist effects of strains of Bacillus spp. against Rhizoctonia solani for their protection against several plant diseases: Alternatives to chemical pesticides. C R Biol. 2019, 342, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Senapati, M.; Tiwari, A.; Sharma, N.; Chandra, P.; Bashyal, B.M.; Ellur, R.K.; Bhowmick, P.K.; Bollinedi, H.; Vinod, K.K.; Singh, A.K.; et al. Rhizoctonia solani Kühn pathophysiology: Status and prospects of sheath blight disease management in rice. Front Plant Sci. 2022, 13, 881116. [Google Scholar] [CrossRef] [PubMed]
- Sumner, D.R. Sclerotia formation by Rhizoctonia species and their survival. In Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology and Disease Control; Sneh, B., Jabaji-Hare, S., Neate, S., Dijst, G., Eds.; Kluwer Academic Publications: Dordrecht, NL, USA, 1996; pp. 207–215. [Google Scholar]
- Chahal, K.S.; Sokhi, S.S.; Rattan, G.S. Investigations on sheath blight of rice in Punjab. Indian Phytopathol. 2003, 56, 22–26. [Google Scholar]
- Kozaka, T. Ecological studies on sheath blight of rice plant caused by Pellicularia sasakii and its chemical control. Chugoku Agric. Res. 1961, 20, 1–13. [Google Scholar]
- Zhang, C.Q.; Liu, Y.H.; Ma, X.Y.; Feng, Z.; Ma, Z.H. Characterization of sensitivity of Rhizoctonia solani, causing rice sheath blight, to mepronil and boscalid. Crop Prot. 2009, 28, 381–386. [Google Scholar] [CrossRef]
- Hussain, M.; Zouhar, M.; Ryšánek, P. Suppression of meloidogyne incognita by the entomopathogenic fungus Lecanicillium muscarium. Plant Dis. 2018, 102, 977–982. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Ji, S.; Wang, Z.; Zhang, H.; Wang, Y.; Liu, Z. Trichoderma asperellum xylanases promote growth and induce resistance in poplar. Microbiol. Res. 2021, 248, 126767. [Google Scholar] [CrossRef]
- Xue, A.G. Biological control of pathogens causing root rot complex in field Pea using Clonostachys rosea strain ACM941. Phytopathology 2003, 93, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Safari Motlagh, M.R.; Jahangiri, B.; Kulus, D.; Tymoszuk, A.; Kaviani, B. Endophytic fungi as potential biocontrol agents against Rhizoctonia solani J.G. Kühn, the causal agent of rice sheath blight disease. Biology 2022, 11, 1282. [Google Scholar] [CrossRef]
- Huang, X.; Chen, L.; Ran, W.; Shen, Q.; Yang, X. Trichoderma harzianum strain SQR-T37 and its bio-organic fertilizer could control Rhizoctonia solani damping-off disease in cucumber seedlings mainly by the mycoparasitism. Appl. Microbiol. Biotechnol. 2011, 91, 741–755. [Google Scholar] [CrossRef]
- Nawrocka, J.; Gromek, A.; Małolepsza, U. Nitric oxide as a beneficial signaling molecule in Trichoderma atroviride TRS25-induced systemic defense responses of cucumber plants against Rhizoctonia solani. Front. Plant Sci. 2019, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Harris, A.R. Biocontrol of Rhizoctonia solani and Pythium ultimum on Capsicum by Trichoderma koningii in potting medium. Microbiol. Res. 1999, 154, 131–135. [Google Scholar] [CrossRef]
- Halifu, S.; Deng, X.; Song, X.; Song, R.; Liang, X. Inhibitory Mechanism of Trichoderma virens ZT05 on Rhizoctonia solani. Plants 2020, 9, 912. [Google Scholar] [CrossRef]
- Reithner, B.; Ibarra-Laclette, E.; Mach, R.L.; Herrera-Estrella, A. Identification of mycoparasitism-related genes in Trichoderma atroviride. Appl. Environ. Microbiol. 2011, 77, 4361–4370. [Google Scholar] [CrossRef] [Green Version]
- Atanasova, L.; Le Crom, S.; Gruber, S.; Coulpier, F.; Seidl-Seiboth, V.; Kubicek, C.P.; Druzhinina, I.S. Comparative transcriptomics reveals different strategies of Trichoderma mycoparasitism. BMC Genom. 2013, 14, 121. [Google Scholar] [CrossRef] [Green Version]
- Keyser, C.A.; Jensen, B.; Meyling, N.V. Dual effects of Metarhizium spp. and Clonostachys rosea against an insect and a seed-borne pathogen in wheat. Pest Manag. Sci. 2016, 72, 517–526. [Google Scholar] [CrossRef]
- Sun, Z.B.; Sun, M.H.; Li, S.D. Identification of mycoparasitism-related genes in Clonostachys rosea 67-1 active against Sclerotinia sclerotiorum. Sci. Rep. 2015, 5, 18169. [Google Scholar] [CrossRef] [Green Version]
- Demissie, Z.A.; Witte, T.; Robinson, K.A.; Sproule, A.; Foote, S.J.; Johnston, A.; Harris, L.J.; Overy, D.P.; Loewen, M.C. Transcriptomic and exometabolomic profiling reveals antagonistic and defensive modes of Clonostachys rosea action against Fusarium graminearum. Mol. Plant Microbe. Interact. 2020, 33, 842–858. [Google Scholar] [CrossRef]
- Nygren, K.; Dubey, M.; Zapparata, A.; Iqbal, M.; Tzelepis, G.D.; Durling, M.B.; Jensen, D.F.; Karlsson, M. The mycoparasitic fungus Clonostachys rosea responds with both common and specific gene expression during interspecific interactions with fungal prey. Evol. Appl. 2018, 11, 931–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lysøe, E.; Dees, M.W.; Brurberg, M.B. A three-way transcriptomic interaction study of a biocontrol agent (Clonostachys rosea), a fungal pathogen (Helminthosporium solani), and a potato host (Solanum tuberosum). Mol. Plant Microbe. Interact. 2017, 30, 646–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.F.; Ma, G.Z.; Bao, Z.H.; Li, S.D. Field test report about biocontrol fungus Gliocladium roseum WP control rice sheath blight. J. Anhui Agri. Sci. 2013, 41, 13242–13255. [Google Scholar]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Audic, S.; Claverie, J.M. The significance of digital gene expression profiles. Genome Res. 1997, 7, 986–995. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Dubey, M.; Jensen, D.F.; Karlsson, M. The ABC transporter ABCG29 is involved in H2O2 tolerance and biocontrol traits in the fungus Clonostachys rosea. Mol. Genet. Genom. 2016, 291, 677–686. [Google Scholar] [CrossRef]
- Sun, Z.B.; Wang, Q.; Sun, M.H.; Li, S.D. The heat shock protein 70 gene is involved for colony morphology, sporulation and mycoparasitism of Clonostachys rosea. FEMS Microbiol. Lett. 2019, 366, fnz188. [Google Scholar] [CrossRef]
- Sun, Z.B.; Wang, Q.; Zhang, J.; Jiang, W.Z.; Wang, Q.; Li, S.D.; Ma, G.Z.; Sun, M.H. The transcription factor-encoding gene crtf is involved in Clonostachys chloroleuca mycoparasitism on Sclerotinia sclerotiorum. Microbiol. Res. 2018, 210, 6–11. [Google Scholar] [CrossRef]
- Sun, Z.B.; Sun, M.H.; Zhou, M.; Li, S.D. Transformation of the endochitinase gene Chi67-1 in Clonostachys rosea 67-1 increases its biocontrol activity against Sclerotinia sclerotiorum. AMB Express 2017, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Fatema, U.; Broberg, A.; Jensen, D.F.; Karlsson, M.; Dubey, M. Functional analysis of polyketide synthase genes in the biocontrol fungus Clonostachys rosea. Sci. Rep. 2018, 8, 15009. [Google Scholar] [CrossRef] [Green Version]
- Zou, C.G.; Tao, N.; Liu, W.J.; Yang, J.K.; Huang, X.W.; Liu, X.Y.; Tu, H.H.; Gan, Z.W.; Zhang, K.Q. Regulation of subtilisin-like protease prC expression by nematode cuticle in the nematophagous fungus Clonostachys rosea. Environ. Microbiol. 2010, 12, 3243–3252. [Google Scholar] [CrossRef]
- Ruocco, M.; Lanzuise, S.; Vinale, F.; Marra, R.; Turrà, D.; Woo, S.L.; Lorito, M. Identification of a new biocontrol gene in Trichoderma atroviride: The role of an ABC transporter membrane pump in the interaction with different plant-pathogenic fungi. Mol. Plant Microbe. Interact. 2009, 22, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Djonović, S.; Vittone, G.; Mendoza-Herrera, A.; Kenerley, C.M. Enhanced biocontrol activity of Trichoderma virens transformants constitutively coexpressing beta-1,3- and beta-1,6-glucanase genes. Mol. Plant Pathol. 2007, 8, 469–480. [Google Scholar] [CrossRef]
- Pozo, M.J.; Baek, J.M.; García, J.M.; Kenerley, C.M. Functional analysis of tvsp1, a serine protease-encoding gene in the biocontrol agent Trichoderma virens. Fungal Genet. Biol. 2004, 41, 336–348. [Google Scholar] [CrossRef]
- Limón, M.C.; Pintor-Toro, J.A.; Benítez, T. Increased antifungal activity of Trichoderma harzianum transformants that overexpress a 33-kDa chitinase. Phytopathology 1999, 89, 254–261. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.B.; Li, S.D.; Ren, Q.; Xu, J.L.; Lu, X.; Sun, M.H. Biology and applications of Clonostachys rosea. J. Appl. Microbiol. 2020, 129, 486–495. [Google Scholar] [CrossRef] [Green Version]
- Pasqualetti, M.; Barghini, P.; Giovannini, V.; Fenice, M. High production of chitinolytic activity in halophilic conditions by a new marine strain of Clonostachys rosea. Molecules 2019, 24, 1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasqualetti, M.; Gorrasi, S.; Giovannini, V.; Braconcini, M.; Fenice, M. Polyextremophilic chitinolytic activity by a marine strain (IG119) of Clonostachys rosea. Molecules 2022, 27, 688. [Google Scholar] [CrossRef] [PubMed]
- Chatterton, S.; Punja, Z.K. Chitinase and beta-1,3-glucanase enzyme production by the mycoparasite Clonostachys rosea f. catenulata against fungal plant pathogens. Can. J. Microbiol. 2009, 55, 356–367. [Google Scholar] [PubMed]
- Li, J.; Yang, J.K.; Huang, X.W.; Zhang, K.Q. Purification and characterization of an extracellular serine protease from Clonostachys rosea and its potential as a pathogenic factor. Process Biochem. 2006, 41, 925–929. [Google Scholar] [CrossRef]
- Pathania, S.; Lore, J.S.; Kalia, A.; Kaur, A.; Sharma, M.; Mangat, G.S.; Sandhu, J.S. Conversion of sheath blight susceptible indica and japonica rice cultivars into moderately resistant through expression of antifungal β-1,3-glucanase transgene from Trichoderma spp. Transgenic. Res. 2022, 31, 537–551. [Google Scholar] [CrossRef]
- Baek, J.M.; Howell, C.R.; Kenerley, C.M. The role of an extracellular chitinase from Trichoderma virens Gv29-8 in the biocontrol of Rhizoctonia solani. Curr. Genet. 1999, 35, 41–50. [Google Scholar] [CrossRef]
- Ferrari, M.; Negri, A.; Romeo, C.; Boccazzi, I.V.; Nodari, R.; Habluetzel, A.; Molteni, G.; Corbett, Y. Adenosine triphosphate-binding cassette transporters are not involved in the detoxification of azadirachta indica extracts In anopheles stephensi Larvae. J. Am. Mosq. Control Assoc. 2018, 34, 311–314. [Google Scholar] [CrossRef]
- Dubey, M.K.; Jensen, D.F.; Karlsson, M. An ATP-binding cassette pleiotropic drug transporter protein is required for xenobiotic tolerance and antagonism in the fungal biocontrol agent Clonostachys rosea. Mol. Plant Microbe. Interact. 2014, 27, 725–732. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Mei, J.; Huang, P.; Tian, Y.; Liang, Y.; Jiang, X.L.; Li, M. Gα3 subunit Thga3 positively regulates conidiation, mycoparasitism, chitinase activity, and hydrophobicity of Trichoderma harzianum. AMB Express 2020, 10, 221. [Google Scholar] [CrossRef]
- Reithner, B.; Brunner, K.; Schuhmacher, R.; Peissl, I.; Seidl, V.; Krska, R.; Zeilinger, S. The G protein alpha subunit Tga1 of Trichoderma atroviride is involved in chitinase formation and differential production of antifungal metabolites. Fungal Genet. Biol. 2005, 42, 749–760. [Google Scholar] [CrossRef]
- Rocha-Ramirez, V.; Omero, C.; Chet, I.; Horwitz, B.A.; Herrera-Estrella, A. Trichoderma atroviride G-protein alpha-subunit gene tga1 is involved in mycoparasitic coiling and conidiation. Eukaryot. Cell 2002, 1, 594–605. [Google Scholar] [CrossRef] [Green Version]
- Zeilinger, S.; Reithner, B.; Scala, V.; Peissl, I.; Lorito, M.; Mach, R.L. Signal transduction by Tga3, a novel g protein subunit alpha of Trichoderma atroviride. Appl. Environ. Microbiol. 2005, 71, 1591–1597. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, P.K.; Latha, J.; Hadar, R.; Horwitz, B.A. Role of two G-protein alpha subunits, TgaA and TgaB, in the antagonism of plant pathogens by Trichoderma virens. Appl. Environ. Microbiol. 2004, 70, 542–549. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Wu, H.; Liu, Z.; Wang, Z.; Huang, B. G-Protein Subunit Gαi in mitochondria, MrGPA1, affects conidiation, stress resistance, and virulence of entomopathogenic fungus Metarhizium robertsii. Front. Microbiol. 2020, 11, 1251. [Google Scholar] [CrossRef]
- Zou, C.G.; Tu, H.H.; Liu, X.Y.; Tao, N.; Zhang, K.Q. PacC in the nematophagous fungus Clonostachys rosea controls virulence to nematodes. Environ. Microbiol. 2010, 12, 1868–1877. [Google Scholar] [CrossRef]
- Trushina, N.; Levin, M.; Mukherjee, P.K.; Horwitz, B.A. PacC and pH-dependent transcriptome of the mycotrophic fungus Trichoderma virens. BMC Genom. 2013, 14, 138. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Han, Y.C.; Yang, L.; Wu, M.D.; Zhang, J.; Cheng, J.S.; Wang, M.Y.; Jiang, D.H.; Chen, W.D.; Li, G.Q. CmpacC regulates mycoparasitism, oxalate degradation and antifungal activity in the mycoparasitic fungus Coniothyrium minitans. Environ. Microbiol. 2015, 17, 4711–4729. [Google Scholar]
- Zhang, M.G.; Wei, Q.L.; Xia, Y.X.; Jin, K. MaPacC, a pH-responsive transcription factor, negatively regulates thermotolerance and contributes to conidiation and virulence in Metarhizium acridum. Curr. Genet. 2020, 66, 397–408. [Google Scholar] [CrossRef]
- Song, Z.Y.; Yin, Y.P.; Lin, Y.L.; Du, F.; Ren, G.W.; Wang, Z.K. The bZIP transcriptional factor activator protein-1 regulates Metarhizium rileyi morphology and mediates microsclerotia formation. Appl. Microbiol. Biotechnol. 2018, 102, 4577–4588. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, X.; Lu, Z.; Wang, H.; He, Z.; Zhou, G.; Luo, Z.; Zhang, Y. MADS-box transcription factor Mcm1 controls cell cycle, fungal development, cell integrity and virulence in the filamentous insect pathogenic fungus Beauveria bassiana. Environ. Microbiol. 2019, 21, 3392–3416. [Google Scholar] [CrossRef]
- Peng, Y.J.; Wang, J.J.; Lin, H.Y.; Ding, J.L.; Feng, M.G.; Ying, S.H. HapX, an indispensable bZIP transcription factor for iron acquisition, regulates infection initiation by orchestrating conidial oleic acid homeostasis and cytomembrane functionality in mycopathogen Beauveria bassiana. mSystems 2020, 5, e00695-20. [Google Scholar] [CrossRef] [PubMed]
- Atanasova, L.; Knox, B.P.; Kubicek, C.P.; Druzhinina, I.S.; Baker, S.E. The polyketide synthase gene pks4 of Trichoderma reesei provides pigmentation and stress resistance. Eukarot. Cell 2013, 12, 1499–1508. [Google Scholar]
- Toopaang, W.; Phonghanpot, S.; Punya, J.; Panyasiri, C.; Klamchao, K.; Wasuwan, R.; Srisuksam, C.; Sangsrakru, D.; Sonthirod, C.; Tangphatsornruang, S.; et al. Targeted disruption of the polyketide synthase gene pks15 affects virulence against insects and phagocytic survival in the fungus Beauveria bassiana. Fungal Biol. 2017, 121, 664–675. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mycoparasitism of Clonostachys rosea on Rhizoctonia solani under microscope observation. C.r. means Clonostachys rosea; R.s. represents Rhizoctonia solani.
Figure 1.
Mycoparasitism of Clonostachys rosea on Rhizoctonia solani under microscope observation. C.r. means Clonostachys rosea; R.s. represents Rhizoctonia solani.

Figure 2.
Gene ontology (GO) categories of the differentially expressed genes. (A) Pre-touch stage; (B) touch stage; (C) after-touch stage.
Figure 2.
Gene ontology (GO) categories of the differentially expressed genes. (A) Pre-touch stage; (B) touch stage; (C) after-touch stage.

Figure 3.
KEGG analysis of the differentially expressed genes. (A) Pre-touch stage; (B) touch stage; (C) after-touch stage.
Figure 3.
KEGG analysis of the differentially expressed genes. (A) Pre-touch stage; (B) touch stage; (C) after-touch stage.

Figure 4.
Phylogenetic analysis of genes from Clonostachys rosea using the maximum likelihood method. Numbers at the nodes indicate the bootstrap values of 1000 bootstraps. Bars represent sequence divergence. (A) ABC transporter; (B) glucanases; (C) chitinase; (D) serine protease.
Figure 4.
Phylogenetic analysis of genes from Clonostachys rosea using the maximum likelihood method. Numbers at the nodes indicate the bootstrap values of 1000 bootstraps. Bars represent sequence divergence. (A) ABC transporter; (B) glucanases; (C) chitinase; (D) serine protease.

Figure 5.
Quantitative real-time PCR to validate the expression levels of DEGs. The X-axis represents different sampling stages, and the Y-axis represents relative expression levels of DEGs. The blue column represents Clonostachys rosea only, and the red column represents added Rhizoctonia solani. Error bars indicate the standard deviation (SD) of three replications.
Figure 5.
Quantitative real-time PCR to validate the expression levels of DEGs. The X-axis represents different sampling stages, and the Y-axis represents relative expression levels of DEGs. The blue column represents Clonostachys rosea only, and the red column represents added Rhizoctonia solani. Error bars indicate the standard deviation (SD) of three replications.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Genes used for phylogenetic analysis.
| Gene Annotation | Gene Name | Isolate | Accession No. |
|---|---|---|---|
| ABC transporter | Taabc2 | T. atroviride | AY911669 |
| β-1,3-glucanase | TvBng2 | T. virens | AF395756 |
| Exo-β-1,3-glucanase | tag83 | T. asperellum | EU314718 |
| β-1,3-glucanase | - | Trichoderma spp. | KJ603460 |
| Subtilisin-like serine protease | TghSS42 | T. ghanense | KJ740359 |
| Subtilisin-like protease | SS10 | T. harzianum | EF063644 |
| Serine protease | tvsp1 | T. virens | AY242844 |
| Chitinase | chit42 | T. virens | AF050098 |
| Chitinase | chit33 | T. harzianum | X80006 |
| Chitinase | chi18-10 | T. atroviride | DQ068748 |
| Chitinase | chi18-13 | T. atroviride | DQ068752 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sun, Z.-B.; Yu, S.-F.; Sun, M.-H.; Li, S.-D.; Hu, Y.-F.; Song, H.-J. Transcriptomic Response of Clonostachys rosea Mycoparasitizing Rhizoctonia solani. J. Fungi 2023, 9, 818. https://doi.org/10.3390/jof9080818
AMA Style
Sun Z-B, Yu S-F, Sun M-H, Li S-D, Hu Y-F, Song H-J. Transcriptomic Response of Clonostachys rosea Mycoparasitizing Rhizoctonia solani. Journal of Fungi. 2023; 9(8):818. https://doi.org/10.3390/jof9080818
Chicago/Turabian StyleSun, Zhan-Bin, Shu-Fan Yu, Man-Hong Sun, Shi-Dong Li, Ya-Feng Hu, and Han-Jian Song. 2023. "Transcriptomic Response of Clonostachys rosea Mycoparasitizing Rhizoctonia solani" Journal of Fungi 9, no. 8: 818. https://doi.org/10.3390/jof9080818
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.