
Endophytic Yeasts for the Biocontrol of Phlyctema vagabunda in Apples
by
 , , and
, , and
Ximena Sepúlveda
1, 
Diego Silva
1,
Ricardo Ceballos
2,
Silvana Vero
3,
María Dolores López
1 and
and
Marisol Vargas
1,* 1
Facultad de Agronomía, Universidad de Concepción, Avenida Vicente Méndez 595, Chillán 3780000, Chile
2
Instituto de Investigaciones Agropecuarias, INIA Quilamapu, Laboratorio de Ecología Química, Av. Vicente Méndez 515, Chillán 3780000, Chile
3
Facultad de Química, Universidad de la República, Av. Gral. Flores 2124, Montevideo 11800, Uruguay
*
Author to whom correspondence should be addressed.
Horticulturae 2022, 8(6), 535; https://doi.org/10.3390/horticulturae8060535
Submission received: 6 April 2022
/
Revised: 9 June 2022
/
Accepted: 13 June 2022
/
Published: 16 June 2022
(This article belongs to the Special Issue Biological Control of Pre- and Postharvest Fungal Diseases)
Abstract
:Bull’s-eye rot, produced by Phlyctema vagabunda, is an important postharvest disease in apples. Current measures to control infection include synthetic fungicides, in addition to the application of copper hydroxide and potassium phosphite. However, growing public concern regarding fungicide residues in food has generated interest in developing non-chemical alternative control methods; biological control is one of the most promising alternatives. In this research, native endophytic yeasts were isolated and evaluated for the biocontrol of P. vagabunda in apples. The mechanisms of action involved were also determined. Our research found 2 isolates, Vishniacozyma victoriae EPL4.5 and EPL29.5, which exhibited biocontrol activity against P. vagabunda at 20 °C in apples, the incidence of bull’s-eye rot was reduced by 39% and 61%, respectively, and the severity of the disease was decreased by 67% and 70%, respectively, when apples were inoculated with these yeasts 24 h before applying the pathogen. The main mechanisms that could be involved in the observed biocontrol activity are the ability to form biofilms and the production of volatile organic compounds.
1. Introduction
Phlyctema vagabunda (Guthrie) Verkley (syn. Neofabraea vagabunda (E.J. Guthrie) Verkley), the causal agent of bull’s-eye rot, has become an important and frequent postharvest disease in apples [1]. It has caused great economic losses in Europe [2], the Pacific Northwest of the United States [3], and Chile [1], and is considered a quarantine disease in some fruit destinations, such as China [4]. In Chile, P. vagabunda was first reported in 2005 [5]. Late-harvest cultivars like ‘Cripps Pink’ are the most affected varieties, with disease incidences reaching 60%, depending on the season and locality [1]. In organic production, incidences in these cultivars can be as high as 80–90%. Fruit infection occurs in the orchard [2] throughout the season [3] and is favored by abundant rainfall [6]. Infection can become established between petal fall and harvest, increasing susceptibility gradually during fruit development [7]. However, symptoms appear only a few months after harvest (usually 3–4 months in cold storage), when numerous lesions can develop on a single fruit [8]. This is, therefore, a major limitation for the late harvest variety ‘Cripps Pink’ when apples are held under prolonged cold storage [9].
Current management practices to control P. vagabunda in Chile include pre- and postharvest treatments with fungicides [10]. However, increasing concerns regarding chemical residues in food [11] have generated interest in developing non-chemical control methods, with biological control being one of the most promising and explored alternatives [12].
The use of antagonistic microorganisms has been recognized as one of the most promising alternatives to fungicides [11]. Among these microorganisms, antagonistic yeasts have proven to effectively control numerous postharvest diseases in apples [13,14,15,16,17,18,19,20,21,22,23]. Nonetheless, no studies have yet been carried out on P. vagabunda. Yeasts may be a viable control alternative for this pathogen because they have the ability to colonize the surface of fruits for long periods of time under dry conditions, produce extracellular polysaccharides that enhance their ability to survive, and rapidly use available nutrients. Yeasts are also minimally affected by pesticides [24], do not produce allergenic spores or mycotoxins [25,26], are genetically stable [19], and have simple nutritional requirements [19,26]. In addition, endophytic yeasts grow in fruit tissues under the same growth conditions as the pathogen P. vagabunda and, therefore, are already colonizing these tissues, which gives them an advantage over the pathogen [27].
An effective biocontrol agent requires multiple modes of action to antagonize a pathogen [28]; a combination of different mechanisms thus provides yeast with its antagonistic capacity [29] and reduces the risk of pathogen resistance [30]. The reported modes of action in yeasts include the ability to compete for nutrients and space, oxidative stress tolerance, parasitism, secretion of hydrolytic enzymes, and the ability to produce siderophores, volatile organic compounds, and biofilms, in addition to the induction of resistance [27,31,32,33,34,35,36,37].
In this research, we selected a native endophytic yeast for the biocontrol of P. vagabunda in apples. The mechanisms of action of the yeast strains involved in the biocontrol activity against the pathogen were also determined. This research could generate beneficial knowledge regarding the application of yeasts for food production and security.
2. Materials and Methods
2.1. Endophyte Isolation and Preparation of Inocula
Endophytic yeasts from ‘Cripps Pink’ apples from organic orchards in the Ñuble and Bío Bío Regions of central Chile were isolated according to the methodology described by Glushakova and Kachalkin [38], with some modifications. The fruits were treated according to the following scheme: 70% ethanol for 30 min; 2% sodium hypochlorite for 30 min; 70% ethanol for 30 s, followed by sterile water for 10 min. The exocarp was then removed and macerated in 5 mL of saline solution (0.9% NaCl); 100 µL of the suspension obtained was spread on Yeast Peptone Dextrose Agar (YPD) containing 0.05 g L−1 of streptomycin (Sigma-Aldrich, St. Louis, MO, USA) and chloramphenicol (Sigma-Aldrich, St. Louis, MO, USA). Petri dishes were then incubated at 4 °C to observe the development of the colonies (between 30 and 45 days).
Inocula of the antagonists for all of the experiments were prepared in a flask containing 20 mL of yeast dextrose broth with a loop of yeast inoculum. The liquid culture was incubated on a rotary shaker (150 rpm) for 72 h at 25 °C. Antagonist cells were then collected by centrifugation at 1914× g for 10 min, washed, and re-suspended in sterile distilled water. The concentration of the suspensions was adjusted to 1 × 109 cells mL−1 by means of a Neubauer’s chamber.
2.2. Fruit
‘Cripps Pink’ apples with no visible wounds were harvested from organic orchards in the Maule Region of central Chile. Apples were superficially disinfected with 0.5% sodium hypochlorite for 5 min, rinsed 3 times with distilled water, and air-dried.
2.3. Pathogen Inoculum
P. vagabunda was obtained from apples affected by bull’s-eye rot and identified by sequencing the β-tubulin gene (GenBank ID: OL450471) as described by Cao et al. [39]. Conidia suspensions were attained according to the methodology described by Cameldi et al. [2]. Briefly, a mycelial plug was transferred to Petri dishes with Tomato Agar and incubated in darkness at 5 °C; after 14 days of incubation, pathogen conidia suspensions were prepared by scraping and suspending conidia in sterile distilled water and adjusted to a concentration of 5 × 105 conidia mL−1, using a Neubauer’s chamber.
2.4. Selection and Identification of Yeasts as a Potential Biocontrol of P. vagabunda
Nine isolates of the most frequently isolated yeast were evaluated as biocontrol agents against P. vagabunda. Apples were wounded in the equatorial axis (3 mm diameter and 3 mm deep) using a sterile pipette tip and inoculated with 20 µL of a yeast suspension (1 × 109 cells mL−1). After 24 h, 20 µL of a P. vagabunda suspension (5 × 105 conidia mL−1) was inoculated [8]. In the control treatment, the yeast cell suspension was substituted for sterile distilled water. After 20 days at 20 °C, rot incidence and severity were recorded according to Vero et al. [40].
Yeast strains were identified by phylogenetic analyses of the D1/D2 domain of the 26S LSU of rRNA using the primers NL1 (5′-GCA TAT CAA TAA GCG GAGGAA AAG-3′) and NL4 (5′-GGT CCG TGT TTC AAGACG G-3′) in MEGA version 11. DNA sequences were aligned, together with sequences of homologous regions of closely related species retrieved from the GenBank. Evolutionary distances were computed using the Jukes–Cantor method, and phylogenetic trees were obtained by neighbor-joining. All positions containing alignment gaps and missing data were eliminated only in pairwise sequence comparisons. The stability of clades was assessed with 1000 bootstrap replications.
Strain identification was confirmed by sequencing the 5.8S-ITS rDNA region using the primers ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) in Macrogen Inc. (Seoul, Korea). These sequences were then compared with those published in the GenBank database with the BLAST program [19].
2.5. Antagonistic Activity of Yeast on the Fruit
The isolates that inhibited the fungus during the selection (n = 2) were evaluated for biocontrol activity, apples were wounded as described above, and 20 µL of a yeast suspension (1 × 109 cells mL−1) was inoculated into each wound. After 24 h, the wounds were inoculated with 20 µL of a P. vagabunda suspension (5 × 105 conidia mL−1) [8]. In the control treatment, the yeast cell suspension was substituted for sterile distilled water. After 20 days at 20 °C, rot incidence and severity were recorded according to Vero et al. [40].
Three replicates were established for each treatment in a completely randomized design in which each replicate was made up of eight apples. The entire experiment was also repeated twice.
2.6. Biofilm Formation by Yeast
The crystal violet (CV) methodology [41] was used to quantify the biofilm formation by yeast. Polystyrene tissue culture multi dishes (Nunclon) with 1800 µL of sterile apple juice were inoculated with a yeast suspension (1 × 109 cells mL−1). The yeast suspension in the control treatment was substituted with distilled sterile water. After 2 days of incubation at 25 °C, the wells were emptied and washed with 2 mL of distilled water using a pipette. This step was carried out 3 times. The biofilm layer on the wall of the wells was fixed by air-drying and stained with 2 mL of 1% crystal violet for 20 min; the cells were then washed and dried again, after which 2 mL of ethanol was added. The absorbance of the eluate was determined at 620 nm with a spectrophotometer (Epoch™ Microplate Spectrophotometer, BioTek, Winooski, VT, USA). Biofilm formation was considered to be positive when absorbance was equal to or higher than that of the control, plus three times the standard deviation [41]. Four replicates were performed, and the experiment was repeated twice.
2.7. Production of Volatile Antifungal Compounds
The antifungal effect of the volatile organic compounds (VOCs) produced by the yeast strains was assayed by the double Petri dish assay according to Di Francesco et al. [42]. For this purpose, plates with YPD or an apple juice agar medium (AJA) were inoculated with 100 µL of a yeast suspension (1 × 108 cells mL−1). Then, 48 h later, 100 µL of a P. vagabunda suspension (1 × 104 conidia mL−1) was inoculated in plates with Potato Dextrose Agar (PDA). Subsequently, the plates with the pathogen were individually covered mouth-to-mouth with the plates containing the yeasts, sealed with parafilm, and incubated at 20 °C. The control corresponded to plates containing YPD or AJA without yeast. The inhibition of the colony-forming unit (CFU) and radial growth of the pathogen was calculated after 10 days of incubation using the equation:
where d1 is the number of CFU in the control and d2 is the number of CFU in the treated.
where d1 is the radial growth (mm of the colony diameter) in the control and d2 is the radial growth (mm of the colony diameter) in the treated.
Three replicates were used for each treatment, and the experiment was repeated twice.
2.8. Chemical Characterization of Volatile Organic Compounds
The analysis of the composition of VOCs produced by yeast was carried out as reported by Zhou et al. [37]. Volatile compounds were collected from yeast samples using Headspace Solid Phase Micro Extraction (HS-SPME), which was identified by gas chromatography–mass spectroscopy (GC–MS; QP2010 Ultra, Shimadzu, Kyoto, Japan). HS-SPME was performed with a 2 cm fiber coated with 50/30 μm DVB/CAR/PDMS. Briefly, we cultured the yeast in a 50 mL Erlenmeyer flask sealed with parafilm and sampled the volatile yeast by inserting the SPME fiber into the head-space of the culture in an Erlenmeyer flask for 5 min at 30 °C. The fiber was injected into a gas chromatograph (GC–MS; QP2010 Ultra, Shimadzu, Kyoto, Japan) containing a 30 m × 0.25 mm fused silica Rxi-5ms column. The chromatographic conditions used were inlet 280 °C; column 40 °C for 2 min followed by ramping at 5 °C min–1 to 280 °C. Mass spectral analyses were carried out with gas chromatography–mass spectroscopy (GC–MS; QP2010 Ultra, Shimadzu, Kyoto, Japan). The scan mass range extended from m/z 35 to 500. Mass spectra of VOCs were compared with those obtained from the NIST05 library, and comparison qualities higher than 90% were considered (Standard Reference Data, NIST, Gaithersburg, MD, USA).
2.9. Statistical Analysis
Statistical analyses of incidence and severity of bull’s-eye rot were subjected to one-way analysis of variance (ANOVA), and comparison of means was performed by Tukey’s with a probability level of 5%. Mycelial growth inhibition (%) was subjected to Student’s t-test mean comparison method with a probability level of 5%. All the data were analyzed by statistical software InfoStat (InfoStat® 2011).
3. Results
A total of 29 endophytic yeasts were obtained from ‘Cripps Pink’ apples, and then 9 isolates of the most frequent yeasts, according to their morphological and phenotypical characteristics, were evaluated against bull’s-eye rot in apples at 20 °C in a preliminary assay. Two native yeasts had biocontrol activity against P. vagabunda and were identified as Vishniacozyma victoriae (EPL4.5 GenBank ID: OL453201, EPL29.5 GenBank ID: OL453202) based on the sequencing of the 5.8S-ITS rDNA region and D1/D2 domain of the 26S LSU of rRNA (Figure 1).
3.1. Antagonistic Activity of Yeast on Fruit
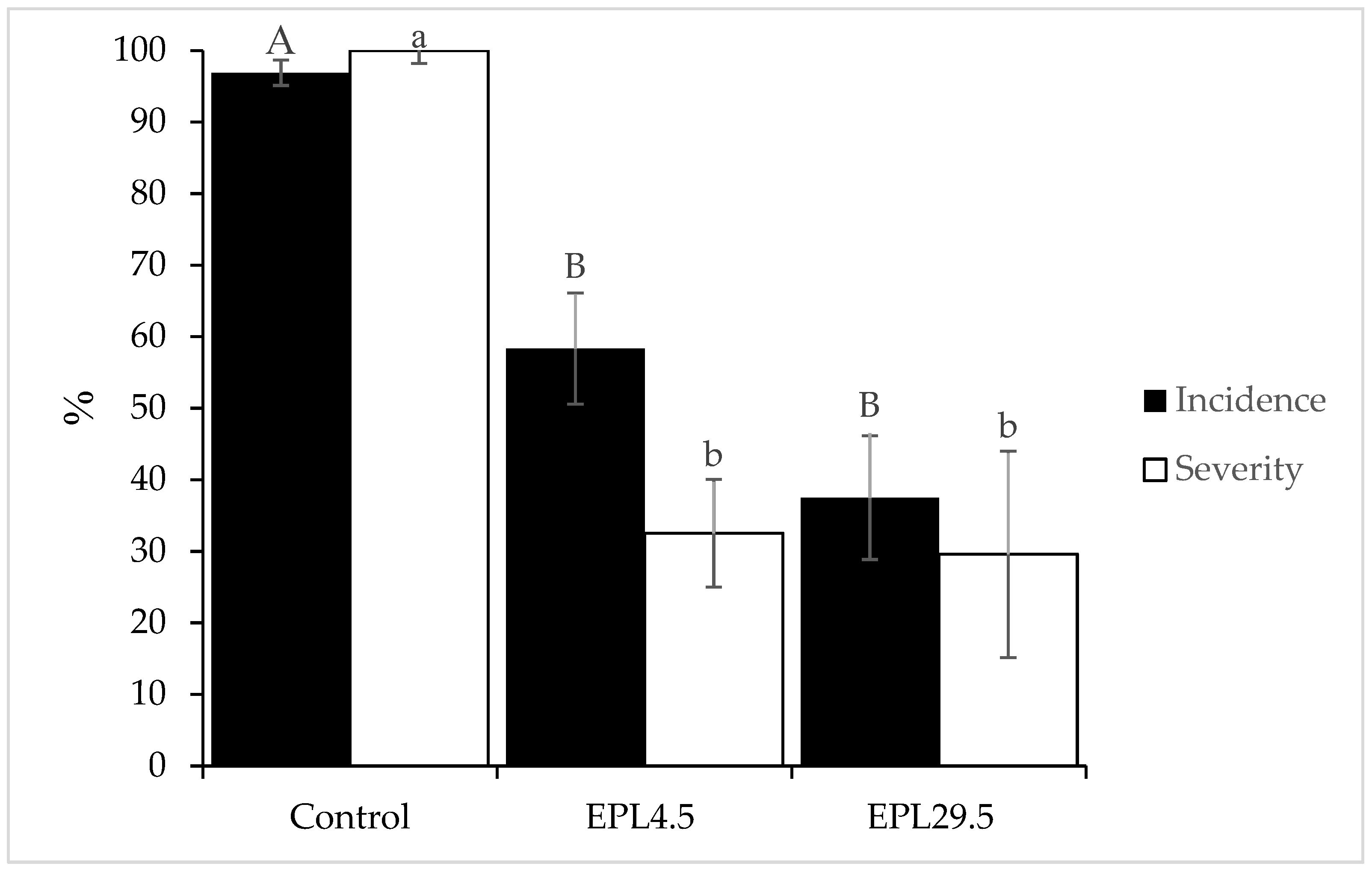
Yeast strains of V. victoriae (EPL4.5 and EPL29.5) inhibited bull’s-eye rot on apples at 20 °C after apple wounds were inoculated with a yeast suspension (1 × 109 cells mL−1), 24 h before inoculation with a P. vagabunda suspension (5 × 105 conidia mL−1) (Figure 2). Yeast strain EPL4.5 reduced the incidence of bull’s-eye disease by 58.3% and the severity by 67.4%, whereas yeast strain EPL29.5 decreased the disease incidence by 37.5% and the severity by 70.3% compared to the control.
3.2. Biofilm Formation
The ability to produce biofilm was evaluated in tissue culture multi dishes (Table 1). One of the two evaluated yeast strains (EPL29.5) was able to produce biofilm, and the optical density value (A620) was found to be higher than the cut-off criterion (0.04).
3.3. Production of Volatile Antifungal Compounds
In the double Petri dish assay system (Figure 3), the VOCs produced by yeast strains EPL4.5 and EPL29.5 on the YPD medium inhibited pathogen colony-forming unit (CFU) by 54.8% and 50.6%, respectively, and mycelial growth was reduced around 69.1% and 66.1%, compared to the control. When the yeast isolates were grown on the AJA medium, strain EPL29.5 reduced CFU by 19.4% and inhibited mycelial growth by a maximum of 43.6%. Strain EPL4.5 inhibited CFU by 10.4% and mycelial growth by 42% (Table 2). These results indicate that the production of volatile organic compounds with biological activity was influenced by the culture medium.
3.4. Chemical Characterization of Volatile Organic Compounds
The VOCs produced by yeast strains EPL4.5 and EPL29.5 on the mediums AJA and YPD were analyzed by GC-MS (Figure 4). The VOCs present in just AJA or YPD medium without any yeast were not considered to be produced by V. victoriae EPL4.5 and EPL29.5. A total of 10 compounds were detected, including 5 alcohols, 2 ketones, and 3 hydrocarbons.
On the YPD medium, the most abundant compound was 1-Butanol; the abundance relative area (RA) of this compound was 46.34% and 38.7% for strains EPL4.5 and EPL29.5, respectively. The rest of the components produced by the strain EPL4.5 on YPD medium mainly corresponded to 1-Butanol, 3-methyl and Disulfide, dimethyl representing an RA of 8.18% and 2.47%. The EPL29.5 strain on YPD medium produced mainly corresponded to 1-Butanol, 3-methyl; 1-Butanol, 2-methyl-, (S)- and 1-Propanol, 2-methyl with an RA between 17.63% and 1.97% (Table 3).
The most abundant compound produced by both yeast strains on the AJA medium was 1-Butanol, 2-methyl-, (S)-; the abundance relative area (RA) of this compound was 19.6% and 13.27% for strains EPL4.5 and EPL29.5, respectively. The rest of the components produced by the strain EPL4.5 on AJA medium mainly corresponded to 1-Butanol; 1-Hexanol and 1-Butanol, 3-methyl with a RA between 3.98% and 1.94%. The EPL29.5 strain on AJA medium produced were 1-Butanol; 1-Hexanol; 1-Propanol, 2-methyl; 2-Nonanone, and 2-Heptanone with a RA between 5.25% and 1.65% (Table 3).
4. Discussion
In this study, we screened endophytic yeasts isolated from organic ‘Cripps Pink’ apples for the control of P. vagabunda. The most effective yeast that significantly decreased the incidence of bull’s-eye rot in apples was the EPL29.5 strain which was identified as Vishniacozyma victoriae. Previously, the yeast V. victoriae had been reported to be an effective biocontrol agent for Botrytis cinerea and Penicillium expansum on pears [47] and Penicillium crustosum and Mucor piriformis on cherries [48]. To the best of our knowledge, no study on P. vagabunda has yet been recorded, making this the first report of a yeast controlling bull’s-eye rot in apples.
P. vagabunda fruit infection occurs in the orchard [2], so the presence of the antagonist prior to or during the initial phases of the disease cycle is of crucial importance, considering the difficulties in controlling previously established infections [48]. We tested that the application of V. victoriae EPL29.5 24 h before the pathogen reduces the incidence of P. vagabunda by 58.3%. The pre-harvest application of antagonistic yeasts could protect fruits against pathogen infection in the orchard, but for the biological control to be successful, the yeast needs to possess effective mechanisms to cope with the abiotic stresses to which they are exposed [49]. In this sense, endophytic yeasts could be a very promising new source of biological control agents because they can grow and develop inside plants, thus avoiding the negative influences of environmental factors such as solar radiation and desiccation [38].
It has been suggested that biological control agents utilize different strategies depending on the pathogen, host, and environment [50]. Understanding these diverse mechanisms is essential to determine how a combination of different yeasts affects pathogen control in order to take advantage of their multiple means of action [51]. This is also an important topic in the development process of bio fungicide formulations because it permits an increase in the performance of biocontrol agents [52].
In this study, we evaluated different mechanisms of action of V. victoriae (EPL4.5 and EPL29.5), including antibiosis, pathogen hyphal adhesion, and siderophore production (Figures S1–S3 Supplementary Material), biofilm-forming capacity and production of volatile organic compounds.
Only V. victoriae EPL29.5 formed biofilms in sterile apple juice at 20 °C. This mechanism has been demonstrated by Lutz et al. [53] at 0 ± 1 °C in pear juice with glucose peptone yeasts extract as a culture medium. Biofilms are a network of cells and extracellular polysaccharides that form a gel that holds microorganisms together [54], creating a mechanical barrier between the wound and the pathogen surface [55], thereby preventing the onset of the infection process.
Several studies have shown that the production of VOCs by yeasts has a significant role in their antagonistic activities [56,57,58]. In this study, VOCs emitted by the V. victoriae strains EPL4.5 and EPL29.5 reduced colony-forming unit (CFU) and mycelial growth of P. vagabunda. Strain EPL4.5 reduced CFU by 54.8% and mycelial growth by 69.1% when grown in a YPD medium, while strain EPL29.5 reduced CFU by 50.63% and mycelial growth by 66.1%.
The analyses of the VOC profiles produced by the evaluated yeast strains were conducted with HS-SMPE coupled with GC-MS, which indicated that the VOCs were mainly alcohols, with the main components being 1-Butanol, 2-methyl-, (S)- in the AJA medium and 1-Butanol in the YPD medium (Table 3). These compounds have previously been observed in the yeasts Aureobasidium pullulans and Meyerozyma caribbica when grown on a Nutrient Broth, Yeast Extract, Dextrose Agar, and PDA [32,43]. Arrate et al. [56] found that the main VOCs produced by C. sake in an AJA medium were 3-Methylbutyl hexanoate, 3-Methylbutyl pentanoate, and 2-Methylpropyl hexanoate; however, none of the compounds found in this research were detected in the current study. This difference may be due to the fact that the production of such volatiles is strongly influenced by the ability of yeast to assimilate and ferment carbohydrates [59].
The production of VOCs characterized as effective biofumigants for disease control in plants [60] may represent an important biological control mechanism for a wide range of postharvest pathogens. In this study, the main components produced by the yeast strains in the media evaluated were alcohols, which damage the plasma membrane and rapidly denature proteins, producing a subsequent interference with metabolism and cell lysis [32] and could explain the observed decreases in colony-forming unit and mycelial growth of P. vagabunda.
Our results suggest that strains EPL4.5 and EPL29.5 of the endophytic yeast V. victoriae are potential biocontrol agents of P. vagabunda in apples and could be used in preharvest applications. Due to the fact that these yeasts are endophytes, they could colonize fruit tissues, including wounds and lenticels, through rapid cell proliferation, thus allowing them to compete with the pathogen. Further research is needed to demonstrate the biocontrol activity of these isolates against bull’s eye rot in orchards.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae8060535/s1, Figure S1: No production of diffusible antifungal compounds of (a) Vishniacozyma victoriae EPL4.5 and (b) Vishniacozyma victoriae EPL29.5 on Potato Dextrose Agar medium at 20 °C. Lawns on plate Phlyctema vagabunda.; Figure S2: Lack of attachment of (a) Vishniacozyma victoriae EPL4.5 and (b) Vishniacozyma victoriae EPL29.5 to the hyphae of Phlyctema vagabunda, after 24 h incubation at 20 °C; Figure S3: No detection of siderophore production by (a) Vishniacozyma victoriae EPL4.5 and (b) Vishniacozyma victoriae EPL29.5 after 15 days of incubation at 20 °C on CAS agar medium.
Author Contributions
M.V. conceived this research; M.V. and X.S. designed experiments; X.S. and D.S. performed the experiments and the analyses in labs; X.S. wrote the manuscript, and M.V., D.S., R.C., S.V. and M.D.L. participated in revising it. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by funding from the National Agency for Research and Development (ANID) through the National Ph.D. Scholarship ID 2118096 and CYTED 121RT0110.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to the National Agency for Research and Development (ANID) and CYTED.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Soto-Alvear, S.; Lolas, M.; Rosales, I.M.; Chávez, E.R.; Latorre, B.A. Characterization of the Bull’s Eye Rot of Apple in Chile. Plant Dis. 2013, 97, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameldi, I.; Neri, F.; Menghini, M.; Pirondi, A.; Nanni, I.M.; Collina, M.; Mari, M. Characterization of Neofabraea Vagabunda Isolates Causing Apple Bull’s Eye Rot in Italy (Emilia-Romagna Region). Plant Pathol. 2017, 66, 1432–1444. [Google Scholar] [CrossRef]
- Henriquez, J.L.; Sugar, D.; Spotts, R.A. Etiology of Bull’s Eye Rot of Pear Caused by Neofabraea Spp. in Oregon, Washington, and California. Plant Dis. 2004, 88, 1134–1138. [Google Scholar] [CrossRef] [Green Version]
- Wood, P.N.; Fisher, B.M. The Effect of Fungicides on Spore Germination, Mycelial Growth and Lesion Development of Phlyctema Vagabunda (Syn: Neofabraea Alba) (Bull’s Eye Rot of Apples). N. Z. Plant Prot. 2017, 70, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Henriquez, J.L. First Report of Apple Rot Caused by Neofabraea Alba in Chile. Plant Dis. 2005, 89, 1360. [Google Scholar] [CrossRef] [PubMed]
- Den Breeyen, A.; Rochefort, J.; Russouw, A.; Meitz-Hopkins, J.; Lennox, C.L. Preharvest Detection and Postharvest Incidence of Phlyctema Vagabunda on ‘Cripps Pink’ Apples in South Africa. Plant Dis. 2020, 104, 841–846. [Google Scholar] [CrossRef]
- Pešicová, K.; Kolařík, M.; Hortová, B.; Novotný, D. Diversity and Identification of Neofabraea Species Causing Bull’s Eye Rot in the Czech Republic. Eur. J. Plant Pathol. 2017, 147, 683–693. [Google Scholar] [CrossRef]
- Neri, F.; Mari, M.; Brigati, S.; Bertolini, P. Control of Neofabraea Alba by Plant Volatile Compounds and Hot Water. Postharvest Biol. Technol. 2009, 51, 425–430. [Google Scholar] [CrossRef]
- Lolas, M.; Cáceres, M.; Reyes, J.A.; Díaz, G.A. Bull’s Eye Rot Development in Storage Is Related to the Timing of Apple Fruit Infection by Neofabraea Vagabunda in the Orchard in Chile. Acta Hortic. 2021, 1325, 73–76. [Google Scholar] [CrossRef]
- Lolas, M.; Díaz, G.; Mendez, R.; Cáceres, M.; Neubauer, L. Evaluation of the Efficacy of Fungicide Fludioxonil in the Postharvest Control of Bull’s Eye Rot (Neofabraea Alba) in Chile. Acta Hortic. 2016, 1144, 461–464. [Google Scholar] [CrossRef]
- Spadaro, D.; Droby, S. Development of Biocontrol Products for Postharvest Diseases of Fruit: The Importance of Elucidating the Mechanisms of Action of Yeast Antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Wisniewski, M.; Wilson, C.; Droby, S.; Chalutz, E.; El-Ghaouth, A.; Stevens, C. Postharvest Biocontrol: New Concepts and Applications. In Biological Control: A Global Perspective; Vincent, C., Goettel, M.S., Lazarovits, G., Eds.; CABI: Wallingford, UK, 2007; pp. 262–273. ISBN 978-1-84593-265-7. [Google Scholar]
- Abdelhai, M.H.; Awad, F.N.; Yang, Q.; Mahunu, G.K.; Godana, E.A.; Zhang, H. Enhancement the Biocontrol Efficacy of Sporidiobolus Pararoseus Y16 against Apple Blue Mold Decay by Glycine Betaine and Its Mechanism. Biol. Control 2019, 139, 104079. [Google Scholar] [CrossRef]
- He, F.; Zhao, L.; Zheng, X.; Abdelhai, M.H.; Boateng, N.S.; Zhang, X.; Zhang, H. Investigating the Effect of Methyl Jasmonate on the Biocontrol Activity of Meyerozyma Guilliermondii against Blue Mold Decay of Apples and the Possible Mechanisms Involved. Physiol. Mol. Plant Pathol. 2020, 109, 101454. [Google Scholar] [CrossRef]
- Li, R.; Zhang, H.; Liu, W.; Zheng, X. Biocontrol of Postharvest Gray and Blue Mold Decay of Apples with Rhodotorula Mucilaginosa and Possible Mechanisms of Action. Int. J. Food Microbiol. 2011, 146, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Manso, T.; Nunes, C. Metschnikowia Andauensis as a New Biocontrol Agent of Fruit Postharvest Diseases. Postharvest Biol. Technol. 2011, 61, 64–71. [Google Scholar] [CrossRef]
- Mari, M.; Martini, C.; Spadoni, A.; Rouissi, W.; Bertolini, P. Biocontrol of Apple Postharvest Decay by Aureobasidium Pullulans. Postharvest Biol. Technol. 2012, 73, 56–62. [Google Scholar] [CrossRef]
- Navarta, L.G.; Calvo, J.; Posetto, P.; Cerutti, S.; Raba, J.; Benuzzi, D.; Sanz, M.I. Postharvest Control of Gray Mold in Apples with Lyophilized Formulations of Cryptococcus Laurentii: The Effect of Cold Stress in the Survival and Effectiveness of the Yeast. Food Bioprocess Technol. 2014, 7, 2962–2968. [Google Scholar] [CrossRef]
- Ruiz-Moyano, S.; Martín, A.; Villalobos, M.C.; Calle, A.; Serradilla, M.J.; Córdoba, M.G.; Hernández, A. Yeasts Isolated from Figs (Ficus Carica L.) as Biocontrol Agents of Postharvest Fruit Diseases. Food Microbiol. 2016, 57, 45–53. [Google Scholar] [CrossRef]
- Türkel, S.; Korukluoğlu, M.; Yavuz, M. Biocontrol Activity of the Local Strain of Metschnikowia Pulcherrima on Different Postharvest Pathogens. Biotechnol. Res. Int. 2014, 2014, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Efficacy of the Antagonist Aureobasidium Pullulans PL5 against Postharvest Pathogens of Peach, Apple and Plum and Its Modes of Action. Biol. Control 2010, 54, 172–180. [Google Scholar] [CrossRef]
- Zhang, D.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Potential Biocontrol Activity of a Strain of Pichia Guilliermondii against Grey Mold of Apples and Its Possible Modes of Action. Biol. Control 2011, 57, 193–201. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Ma, L.; Dong, Y.; Jiang, S.; Xu, B.; Zheng, X. Biocontrol of Major Postharvest Pathogens on Apple Using Rhodotorula Glutinis and Its Effects on Postharvest Quality Parameters. Biol. Control 2009, 48, 79–83. [Google Scholar] [CrossRef]
- Wilson, C.L.; Wisniewski, M.E. Biological Control of Postharvest Diseases of Fruits and Vegetables: An Emerging Technology*. Annu. Rev. Phytopathol. 1989, 27, 425–441. [Google Scholar] [CrossRef]
- Ocampo-Suarez, I.B.; López, Z.; Calderón-Santoyo, M.; Ragazzo-Sánchez, J.A.; Knauth, P. Are Biological Control Agents, Isolated from Tropical Fruits, Harmless to Potential Consumers? Food Chem. Toxicol. 2017, 109, 1055–1062. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Biocontrol Ability and Action Mechanism of Food-Isolated Yeast Strains against Botrytis Cinerea Causing Post-Harvest Bunch Rot of Table Grape. Food Microbiol. 2015, 47, 85–92. [Google Scholar] [CrossRef]
- Madbouly, A.K.; Abo Elyousr, K.A.M.; Ismail, I.M. Biocontrol of Monilinia Fructigena, Causal Agent of Brown Rot of Apple Fruit, by Using Endophytic Yeasts. Biol. Control 2020, 144, 104239. [Google Scholar] [CrossRef]
- Chanchaichaovivat, A.; Panijpan, B.; Ruenwongsa, P. Putative Modes of Action of Pichia Guilliermondii Strain R13 in Controlling Chilli Anthracnose after Harvest. Biol. Control 2008, 47, 207–215. [Google Scholar] [CrossRef]
- Bautista-Rosales, P.U.; Calderon-Santoyo, M.; Servín-Villegas, R.; Ochoa-Álvarez, N.A.; Ragazzo-Sánchez, J.A. Action Mechanisms of the Yeast Meyerozyma Caribbica for the Control of the Phytopathogen Colletotrichum Gloeosporioides in Mangoes. Biol. Control 2013, 65, 293–301. [Google Scholar] [CrossRef]
- Nally, M.C.; Pesce, V.M.; Maturano, Y.P.; Rodriguez Assaf, L.A.; Toro, M.E.; Castellanos de Figueroa, L.I.; Vazquez, F. Antifungal Modes of Action of Saccharomyces and Other Biocontrol Yeasts against Fungi Isolated from Sour and Grey Rots. Int. J. Food Microbiol. 2015, 204, 91–100. [Google Scholar] [CrossRef]
- Chan, Z.; Tian, S. Induction of H2O2-Metabolizing Enzymes and Total Protein Synthesis by Antagonistic Yeast and Salicylic Acid in Harvested Sweet Cherry Fruit. Postharvest Biol. Technol. 2006, 39, 314–320. [Google Scholar] [CrossRef]
- Iñiguez-Moreno, M.; Ragazzo-Sánchez, J.A.; Barros-Castillo, J.C.; Sandoval-Contreras, T.; Calderón-Santoyo, M. Sodium Alginate Coatings Added with Meyerozyma Caribbica: Postharvest Biocontrol of Colletotrichum Gloeosporioides in Avocado (Persea Americana Mill. Cv. Hass). Postharvest Biol. Technol. 2020, 163, 111123. [Google Scholar] [CrossRef]
- Liu, Y.; Yao, S.; Deng, L.; Ming, J.; Zeng, K. Different Mechanisms of Action of Isolated Epiphytic Yeasts against Penicillium Digitatum and Penicillium Italicum on Citrus Fruit. Postharvest Biol. Technol. 2019, 152, 100–110. [Google Scholar] [CrossRef]
- Tian, Y.; Li, W.; Jiang, Z.; Jing, M.; Shao, Y. The Preservation Effect of Metschnikowia Pulcherrima Yeast on Anthracnose of Postharvest Mango Fruits and the Possible Mechanism. Food Sci. Biotechnol. 2018, 27, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, L.; Li, S.; Gao, X.; Wu, N.; Zhao, Y.; Sun, W. Control of Postharvest Grey Spot Rot of Loquat Fruit with Metschnikowia Pulcherrima E1 and Potential Mechanisms of Action. Biol. Control 2021, 152, 104406. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.; Xie, J.; Deng, L.; Yao, S.; Zeng, K. Biocontrol Efficacy of Pichia Membranaefaciens and Kloeckera Apiculata against Monilinia Fructicola and Their Ability to Induce Phenylpropanoid Pathway in Plum Fruit. Biol. Control 2019, 129, 83–91. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, W.; Zeng, J.; Shao, Y. Mechanisms of Action of the Yeast Debaryomyces Nepalensis for Control of the Pathogen Colletotrichum Gloeosporioides in Mango Fruit. Biol. Control 2018, 123, 111–119. [Google Scholar] [CrossRef]
- Glushakova, A.M.; Kachalkin, A.V. Endophytic Yeasts in Malus Domestica and Pyrus Communis Fruits under Anthropogenic Impact. Microbiology 2017, 86, 128–135. [Google Scholar] [CrossRef]
- Cao, D.; Li, X.; Cao, J.; Wang, W. PCR Detection of the Three Neofabraea Pathogenic Species Responsible for Apple Bull’s Eye Rot. Adv. Microbiol. 2013, 03, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Vero, S.; Mondino, P.; Burgueño, J.; Soubes, M.; Wisniewski, M. Characterization of Biocontrol Activity of Two Yeast Strains from Uruguay against Blue Mold of Apple. Postharvest Biol. Technol. 2002, 26, 91–98. [Google Scholar] [CrossRef]
- Růžička, F.; Holá, V.; Votava, M.; Tejkalová, R. Importance of Biofilm InCandida Parapsilosis and Evaluation of Its Susceptibility to Antifungal Agents by Colorimetric Method. Folia Microbiol. 2007, 52, 209–214. [Google Scholar] [CrossRef]
- Di Francesco, A.; Di Foggia, M.; Baraldi, E. Aureobasidium Pullulans Volatile Organic Compounds as Alternative Postharvest Method to Control Brown Rot of Stone Fruits. Food Microbiol. 2020, 87, 103395. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of Volatile Organic Compounds by Aureobasidium Pullulans as a Potential Mechanism of Action against Postharvest Fruit Pathogens. Biol. Control 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Jaibangyang, S.; Nasanit, R.; Limtong, S. Biological Control of Aflatoxin-Producing Aspergillus Flavus by Volatile Organic Compound-Producing Antagonistic Yeasts. BioControl 2020, 65, 377–386. [Google Scholar] [CrossRef]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marcello, A.; Oggiano, S.; Balmas, V.; Hassan, Z.U.; Jaoua, S.; Migheli, Q. Effect of Yeast Volatile Organic Compounds on Ochratoxin A-Producing Aspergillus Carbonarius and A. Ochraceus. Int. J. Food Microbiol. 2018, 284, 1–10. [Google Scholar] [CrossRef]
- Qin, X.; Xiao, H.; Cheng, X.; Zhou, H.; Si, L. Hanseniaspora Uvarum Prolongs Shelf Life of Strawberry via Volatile Production. Food Microbiol. 2017, 63, 205–212. [Google Scholar] [CrossRef]
- Lutz, M.C.; Lopes, C.A.; Sosa, M.C.; Sangorrín, M.P. Semi-Commercial Testing of Regional Yeasts Selected from North Patagonia Argentina for the Biocontrol of Pear Postharvest Decays. Biol. Control 2020, 150, 104246. [Google Scholar] [CrossRef]
- Villalba, M.L.; Lutz, M.C.; Lopez, S.; Pildain, M.B.; Sangorrín, M.P. Patagonian Antagonist Yeasts for Food Biopreservation. In Biology and Biotechnology of Patagonian Microorganisms; Olivera, N.L., Libkind, D., Donati, E., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 301–323. ISBN 978-3-319-42799-7. [Google Scholar]
- Sui, Y.; Wisniewski, M.; Droby, S.; Liu, J. Responses of Yeast Biocontrol Agents to Environmental Stress. Appl. Environ. Microbiol. 2015, 81, 2968–2975. [Google Scholar] [CrossRef] [Green Version]
- Di Francesco, A.; Ugolini, L.; D’Aquino, S.; Pagnotta, E.; Mari, M. Biocontrol of Monilinia Laxa by Aureobasidium Pullulans Strains: Insights on Competition for Nutrients and Space. Int. J. Food Microbiol. 2017, 248, 32–38. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, W.; Zhou, Y.; Yao, S.; Deng, L.; Zeng, K. Isolation, Identification and in Vitro Screening of Chongqing Orangery Yeasts for the Biocontrol of Penicillium Digitatum on Citrus Fruit. Biol. Control 2017, 110, 18–24. [Google Scholar] [CrossRef]
- Pesce, V.M.; Nally, M.C.; Carrizo, G.P.; Rojo, C.; Pérez, B.A.; Toro, M.E.; Castellanos de Figueroa, L.I.; Vazquez, F. Antifungal Activity of Native Yeasts from Different Microenvironments against Colletotrichum Gloeosporioides on Ripe Olive Fruits. Biol. Control 2018, 120, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Lutz, M.C.; Lopes, C.A.; Rodriguez, M.E.; Sosa, M.C.; Sangorrín, M.P. Efficacy and Putative Mode of Action of Native and Commercial Antagonistic Yeasts against Postharvest Pathogens of Pear. Int. J. Food Microbiol. 2013, 164, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.N.; Kupper, K.C. Biofilm Production by Aureobasidium Pullulans Improves Biocontrol against Sour Rot in Citrus. Food Microbiol. 2018, 69, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Francesco, A.; Martini, C.; Mari, M. Biological Control of Postharvest Diseases by Microbial Antagonists: How Many Mechanisms of Action? Eur. J. Plant Pathol. 2016, 145, 711–717. [Google Scholar] [CrossRef]
- Arrarte, E.; Garmendia, G.; Rossini, C.; Wisniewski, M.; Vero, S. Volatile Organic Compounds Produced by Antarctic Strains of Candida Sake Play a Role in the Control of Postharvest Pathogens of Apples. Biol. Control 2017, 109, 14–20. [Google Scholar] [CrossRef]
- Konsue, W.; Dethoup, T.; Limtong, S. Biological Control of Fruit Rot and Anthracnose of Postharvest Mango by Antagonistic Yeasts from Economic Crops Leaves. Microorganisms 2020, 8, 317. [Google Scholar] [CrossRef] [Green Version]
- Medina-Córdova, N.; López-Aguilar, R.; Ascencio, F.; Castellanos, T.; Campa-Córdova, A.I.; Angulo, C. Biocontrol Activity of the Marine Yeast Debaryomyces Hansenii against Phytopathogenic Fungi and Its Ability to Inhibit Mycotoxins Production in Maize Grain (Zea Mays L.). Biol. Control 2016, 97, 70–79. [Google Scholar] [CrossRef]
- Fialho, M.B.; Toffano, L.; Pedroso, M.P.; Augusto, F.; Pascholati, S.F. Volatile Organic Compounds Produced by Saccharomyces Cerevisiae Inhibit the in Vitro Development of Guignardia Citricarpa, the Causal Agent of Citrus Black Spot. World J. Microbiol. Biotechnol. 2010, 26, 925–932. [Google Scholar] [CrossRef]
- Huang, R.; Li, G.Q.; Zhang, J.; Yang, L.; Che, H.J.; Jiang, D.H.; Huang, H.C. Control of Postharvest Botrytis Fruit Rot of Strawberry by Volatile Organic Compounds of Candida Intermedia. Phytopathology 2011, 101, 859–869. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Phylogenetic analysis of yeast Vishniacozyma victoriae strains EPL4.5 and Vishniacozyma victoriae EPL29.5 with large subunit ribosomal gene nucleotide sequences. This analysis was inferred using the neighbor-joining method. The percentages of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches. The scale bar (0.02) indicates 0.2% divergence. This analysis involved 26 nucleotide sequences.
Figure 1.
Phylogenetic analysis of yeast Vishniacozyma victoriae strains EPL4.5 and Vishniacozyma victoriae EPL29.5 with large subunit ribosomal gene nucleotide sequences. This analysis was inferred using the neighbor-joining method. The percentages of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches. The scale bar (0.02) indicates 0.2% divergence. This analysis involved 26 nucleotide sequences.

Figure 2.
Incidence and severity of bull’s-eye rot in ‘Cripps Pink’ apples treated with endophytic yeasts. Fruits were wounded and treated with a yeast suspension (1 × 109 cells mL−1). After 24 h, a spore suspension of pathogen fungus (5 × 105 spores mL−1) was applied, and the apples were then stored at 20 °C for 20 days. Mean values of incidence or severity linked by the same letter (upper or lower case, respectively) are not significantly different according to the Tukey’s test (Incidence dF = 2; F= 10.16; p = 0.0119; Severity dF = 2; F = 32.25; p = 0.0006).
Figure 2.
Incidence and severity of bull’s-eye rot in ‘Cripps Pink’ apples treated with endophytic yeasts. Fruits were wounded and treated with a yeast suspension (1 × 109 cells mL−1). After 24 h, a spore suspension of pathogen fungus (5 × 105 spores mL−1) was applied, and the apples were then stored at 20 °C for 20 days. Mean values of incidence or severity linked by the same letter (upper or lower case, respectively) are not significantly different according to the Tukey’s test (Incidence dF = 2; F= 10.16; p = 0.0119; Severity dF = 2; F = 32.25; p = 0.0006).

Figure 3.
Inhibition of Phlyctema vagabunda growth by Vishniacozyma victoriae strains EPL4.5 and EPL29.5 antifungal volatile compounds activity on double Petri dish. Pathogen colony-forming unit and mycelial growth were inhibited in the presence of the yeast compared to the control.
Figure 3.
Inhibition of Phlyctema vagabunda growth by Vishniacozyma victoriae strains EPL4.5 and EPL29.5 antifungal volatile compounds activity on double Petri dish. Pathogen colony-forming unit and mycelial growth were inhibited in the presence of the yeast compared to the control.

Figure 4.
Gas Chromatography–Mass Spectrometry traces (total ion chromatograms) of VOCs collected in SPME. (a) Vishniacozyma victoriae EPL.4.5 on AJA medium, (b) Vishniacozyma victoriae EPL.4.5 on YPD medium, (c) Vishniacozyma victoriae EPL.29.5 on AJA medium, (d) Vishniacozyma victoriae EPL.29.5 on YPD medium. Peak top numbers refer to compounds listed in Table 3.
Figure 4.
Gas Chromatography–Mass Spectrometry traces (total ion chromatograms) of VOCs collected in SPME. (a) Vishniacozyma victoriae EPL.4.5 on AJA medium, (b) Vishniacozyma victoriae EPL.4.5 on YPD medium, (c) Vishniacozyma victoriae EPL.29.5 on AJA medium, (d) Vishniacozyma victoriae EPL.29.5 on YPD medium. Peak top numbers refer to compounds listed in Table 3.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Biofilm formation in polystyrene tissue culture multi dishes by Vishniacozyma victoriae yeast strains EPL4.5 and EPL29.5 at 20 °C. Biofilm formation was considered to be positive when absorbance was equal to or higher than that of the control, plus three times the standard deviation.
Table 1.
Biofilm formation in polystyrene tissue culture multi dishes by Vishniacozyma victoriae yeast strains EPL4.5 and EPL29.5 at 20 °C. Biofilm formation was considered to be positive when absorbance was equal to or higher than that of the control, plus three times the standard deviation.
| Yeast Strain | Biofilm Formation | |
|---|---|---|
| Absorbance (A620) | ||
| EPL4.5 | 0.023 ± 0.003 | − |
| EPL29.5 | 0.061 ± 0.006 | + |
+ Presence of biofilm. − Absence of biofilm. Cut-off value of biofilm formation = 0.04. Data on biofilm formation is expressed as mean ± standard error.
Table 2.
Effect of VOCs produced by Vishniacozyma victoriae yeast strains EPL4.5 and EPL29.5 in Yeast Peptone Dextrose Agar (YPD) and apple juice agar (AJA) mediums on colony-forming unit and mycelial growth of Phlyctema vagabunda at 20 °C.
Table 2.
Effect of VOCs produced by Vishniacozyma victoriae yeast strains EPL4.5 and EPL29.5 in Yeast Peptone Dextrose Agar (YPD) and apple juice agar (AJA) mediums on colony-forming unit and mycelial growth of Phlyctema vagabunda at 20 °C.
| Yeast Strain | Colony Forming Unit Inhibition (%) | Mycelial Growth Inhibition (%) | ||
|---|---|---|---|---|
| YPD | AJA | YPD | AJA | |
| EPL4.5 | 54.84 ± 27.4 | 10.38 ± 21.9 | 69.14 ± 3.6 * | 42.1 ± 7.4 |
| EPL29.5 | 50.63 ± 7.6 | 19.44 ± 7.3 | 66.10 ± 12.1 | 43.62 ± 2.4 |
Data expressed as mean ± standard error. Asterisks denote a significant difference among treatments for the same yeast strain (EPL4.5 dF = 3, p = 0.046; EPL29.5 dF = 3, p = 0.0997), according to Student’s t-test.
Table 3.
Chemical characterization and relative abundance (%) of volatile fraction of Vishniacozyma victoriae yeast strains EPL4.5 and EPL29.5 in YPD and AJA media.
Table 3.
Chemical characterization and relative abundance (%) of volatile fraction of Vishniacozyma victoriae yeast strains EPL4.5 and EPL29.5 in YPD and AJA media.
| Peak Number | RT(min) | Possible Compound | Molecular Formula | m/z | Relative Abundance (%) | Reference | |||
|---|---|---|---|---|---|---|---|---|---|
| EPL4.5 | EPL29.5 | ||||||||
| AJA | YPD | AJA | YPD | ||||||
| 1 | 2.14 | Hexane # | C6H14 | 41 | 0.49 | N.D. | 2.15 | N.D. | |
| 2 | 2.32 | Isobutyl chloride | C4H9Cl | 43 | 0.65 | 1.83 | N.D. | N.D. | |
| 3 | 2.41 | 1-Propanol, 2-methyl | C4H10O | 43 | 0.46 | 0.53 | 2.19 | 1.97 | [43,44] |
| 4 | 2.77 | 1-Butanol | C4H10O | 56 | 3.98 | 46.34 | 5.25 | 38.7 | |
| 5 | 3.18 | Silanediol, dimethy l # | C2H8O2Si | 77 | 3.97 | 2.35 | 7.82 | 2.69 | |
| 6 | 3.43 | 2,5-Dimethylfuran | C6H8O | 45 | 1.08 | N.D. | N.D. | N.D. | |
| 7 | 3.87 | 1-Butanol, 3-methyl | C5H12O | 56 | 1.94 | 8.18 | N.D. | 17.63 | [32,35,43] |
| 8 | 3.98 | 1-Butanol, 2-methyl-, (S)- | C5H12O | 41 | 19.06 | 1.33 | 13.27 | 2.97 | [32,43,44] |
| 9 | 4.13 | Disulfide, dimethyl | C2H6S2 | 94 | N.D. | 2.47 | N.D. | N.D. | [45] |
| 10 | 5.95 | Cyclotrisiloxane, hexamethyl- # | C6H18O3Si3 | 207 | 5.55 | 3.25 | N.D. | N.D. | |
| 11 | 7.32 | 1-Hexanol | C6H14O | 56 | 3.39 | N.D. | 3.27 | N.D. | [35,46] |
| 12 | 7.99 | 2-Heptanone | C7H14O | 43 | N.D. | N.D. | 1.65 | N.D. | [45] |
| 13 | 14.51 | 2-Nonanone | C9H18O | 43 | N.D. | N.D. | 2.04 | 0.49 | [35] |
| 14 | 16.45 | Cyclopentasiloxane, decamethyl # | C10H30O5Si5 | 73 | N.D. | N.D. | 3.99 | 1.34 | |
| 15 | 20.35 | Cyclohexasiloxane, dodecamethyl # | C12H36O6Si6 | 88 | N.D. | N.D. | 2.94 | N.D. | |
Not detected is referred to as N.D. # Putative compounds from the column and fiber.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sepúlveda, X.; Silva, D.; Ceballos, R.; Vero, S.; López, M.D.; Vargas, M. Endophytic Yeasts for the Biocontrol of Phlyctema vagabunda in Apples. Horticulturae 2022, 8, 535. https://doi.org/10.3390/horticulturae8060535
AMA Style
Sepúlveda X, Silva D, Ceballos R, Vero S, López MD, Vargas M. Endophytic Yeasts for the Biocontrol of Phlyctema vagabunda in Apples. Horticulturae. 2022; 8(6):535. https://doi.org/10.3390/horticulturae8060535
Chicago/Turabian StyleSepúlveda, Ximena, Diego Silva, Ricardo Ceballos, Silvana Vero, María Dolores López, and Marisol Vargas. 2022. "Endophytic Yeasts for the Biocontrol of Phlyctema vagabunda in Apples" Horticulturae 8, no. 6: 535. https://doi.org/10.3390/horticulturae8060535
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.