
Abstract
While isolating fungal strains from soil samples, two strains, Talaromyces siamensis FKA-61 and Phomopsis sp. FKA-62, were found to produce a black pigment near the boundary with an adjoining colony on agar plates. Moreover, strain FKA-61 inhibited the growth of strain FKA-62. Co-culture of T. siamensis FKA-61 with Phomopsis sp. FKA-62 resulted in the production of an antifungal agent, BE-31405, which proved active against Phomopsis sp. FKA-62. We screened for fungal genera that have the ability to induce production of antifungal BE-31405. Surprisingly, production of BE-31405 was induced only by co-culturing T. siamensis FKA-61 with a variety of BE-31405-sensitive fungal strains but never with BE-31405-insensitive strains. This is the first report of the induced production of a specific antibiotic active against antibiotic-sensitive fungal strains by co-culturing with them.
Similar content being viewed by others

Introduction
Co-culture is a relatively new and promising method to induce the creation of novel microbial metabolites, as well as for increasing the productivity of known metabolites.1, 2, 3 We have already reported the discovery of secopenicillide C, together with four known compounds, penicillide, MC-141, pestalasin A and stromemycin, from a co-culture broth of two fungal strains, Penicillium pinophilum (current name; Talaromyces pinophilus) FKI-5653 and Trichoderma harzianum FKI-5655.4 While searching for useful fungal metabolites using the co-culture method, we found that a combination of Talaromyces siamensis FKA-61 with Phomopsis sp. FKA-62 resulted in the production of the antifungal agent, BE-31405,5 which displays activity against FKA-62. Moreover, we found a wide range of fungal taxa, which could induce the production of BE-31405, when co-cultured with T. siamensis FKA-61. We report here the characterization of fungal metabolites, including BE-31405, isolated from a co-culture broth of two fungal strains, Talaromyces siamensis FKA-61 and Phomopsis sp. FKA-62, as well as the activities of BE-31405 against fungi having the ability to induce production of BE-31405.
Results
Identification of BE-31405, produced by co-culture of T. siamensis with Phomopsis sp. FKA-62, as an antifungal agent
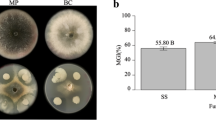
Talaromyces siamensis FKA-61 and Phomopsis sp. FKA-62 were co-cultured on potato dextrose agar (PDA, Difco Laboratories, Detroit, MI, USA), where the colony of FKA-62 could not approach that of FKA-61 (Figure 1). Production of black pigments (probably melanin) was observed at the face of FKA-62 near to the border with FKA-61. Black pigment did not appear in pure culture of either organism.

Co-culture of two fungal strains, Talaromyces siamensis FKA-61 and Phomopsis sp. FKA-62, grown on potato dextrose agar (PDA) at 25 °C for 6, 7 and 10 days. A full color version of this figure is available at The Journal of Antibiotics journal online.
On the liquid culture in a 500-ml Erlenmeyer flask containing potato dextrose broth (Difco), orange pigments were produced by co-culture. Conversely, dark red and yellow pigments were produced during singlar pure culture of FKA-61 and FKA-62, respectively (Figure 2, center).

Pure culture and co-culture of two fungal strains, Talaromyces siamensis FKA-61 and Phomopsis sp. FKA-62, grown on potato dextrose broth (PDB) in a 500-ml Erlenmeyer flask at 27 °C for 1–7 days (center) and on PDB in a dialysis-culture flask at 27 °C for 1–7 days (right) and 1–7-day-old co-culture broth grown on potato dextrose agar (PDA) at 25 °C for 7 days (left). A full color version of this figure is available at The Journal of Antibiotics journal online.
The three culture broths (pure culture of FKA-61, and of FKA-62, plus the co-culture of FKA-61 with FKA-62) added to same volume of EtOH were evaluated for antifungal activity against both FKA-61 and FKA-62 using the paper disc method (Table 1). Only the co-culture broth showed activity against FKA-62. This result strongly indicated that production of antifungal agents by FKA-61 was induced as a result of co-culture with FKA-62. This co-culture broth also had selective antifungal activity against Mucor racemosus.
Therefore, we proceeded to purify a co-culture broth guided by antifungal activity against FKA-62. As a result, we identified BE-31405 (Figure 3) 5 as a antifungal agent. BE-31405 showed potent antifungal activity against FKA-62 (inhibition zone: 21 mm per 10 μg), without activity against FKA-61, even at 100 μg (Tables 1 and 2).

Structures of sclerotinin A, griseofulvin, vermistatin, chrodrimanin F, chrodrimanin E, thailandolide B, dihydrogladiolic acid and BE-31405.
Finally, metabolite profile analysis by HPLC confirmed that production of BE-31405 was induced by co-culturing FKA-61 with FKA-62 (Figure 4). BE-31405 was detected only in the co-culture broth, with the productivity of around 170 mg l−1. In comparison, the productivities of five other metaboliotes (sclerotinin A,6 griseofulvin,7 vermistatin,8 chrodrimanin F9 and chrodrimanin E9) produced by FKA-61 in pure culture were not affected by co-culturing with FKA-62. Therefore, we confirmed that the antifungal agent BE-31405 was induced by co-culturing the two strains, FKA-61 and FKA-62.

HPLC chromatograms of 50% EtOH extract of co-culture and pure culture. The right column indicates UV spectra of sclerotinin A, griseofulvin, vermistatin, BE-31405 and thailandolide B produced by Talaromyces siamensis FKA-61, as well as that of dihydrogladiolic acid by Phomopsis sp. FKA-62. A full color version of this figure is available at The Journal of Antibiotics journal online.
To reveal which strain assumes the majority under co-culture conditions, a loopful of each co-culture broth of FKA-61 with FKA-62 from day 1 to day 7 during incubation was spread on a PDA plate, followed by incubation at 27 °C for 7 days (Figure 2, left). As a result, FKA-61 grew dominantly from day 1, despite the growth of FKA-62 being generally faster than that of FKA-61 on PDA (Figure 1). This suggests that, in co-culture, FKA-61 is the dominant strain, probably due to the antifungal effect of BE-31405 produced by FKA-61.
To clarify whether BE-31405 is produced by co-culture in the absence of any direct contact between the two strains, co-culture was performed using a dialysis culture flask (Figure 2, right). As a result, BE-31405 was not detected. This result suggests that direct contact between the two distinct strains is necessary to induce production of BE-31405.
Identification of FKA-61 as a BE-31405-producing strain
In a co-culture, a colony of FKA-62 did not approach one of FKA-61. Additionally BE-31405 was active against FKA-62 but not against FKA-61. Therefore, we predicted that the strain producing an antifungal factor was likely to be FKA-61. To validate this hypothesis, we conducted co-culturing of FKA-61 or FKA-62 with eight other fungal strains, each belonging to quite different families and orders. Surprisingly, we could detect production of BE-31405 in co-cultures of FKA-61 with six of the fungal strains tested (Table 2). However, FKA-62 did not produce BE-31405 with any strains. This confirmed that FKA-61 is the strain that produces BE-31405.
Antifungal activity of BE-31405 against fungal strains with or without the ability to induce production of BE-31405 by FKA-61
We evaluated the antifungal activity of BE-31405 against nine fungal strains, irrespective of whether co-culturing with FKA-61 produced BE-31405. As shown in Table 2, BE-31405 showed antifungal activity against only those six of the strains tested that were able to induce production of BE-31405 by FKA-61. It was not active against the two strains that did not induce FKA-61 to produce BE-31405. This suggests that FKA-61 can produce BE-31405 to attack other fungi that are sensitive to BE-31405 by somehow detecting the presence of such fungi.
Discussion
A co-culture method was effective in stimulating the generation of novel microbial metabolites and increasing the productivity of known metabolites. However, only specific fungal combinations displayed these characteristics. In a previous study, we reported that metabolites production, including novel productivity of Penicillium pinophilum FKI-5653, was enhanced by co-culturing with Trichoderma harzianum FKI-5655.4 At the same time, red pigment was induced, or much increased, by co-culture on an agar plate. In another study, we observed that a novel yellow pigment was produced during co-culture of two different species of the genus Trichoderma on an agar plate as well as in liquid culture.10 Exploiting these phenomena, we screened co-cultured fungal combinations on an agar plate using pigment production as an indicator.
Two fungal strains, Talaromyces siamensis FKA-61 and Phomopsis sp. FKA-62, produced a black pigment in the vicinity of the boundary of the adjoining colony when cultured on an agar plate. Moreover, strain FKA-61 inhibited the growth of strain FKA-62. In an earlier report of co-culturing of fungal strains, enhancement of pigment productivity of Monascus occurred as a result of co-culture with either Saccharomyces cerevisiae or Aspergillus oryzae.11 Further experiments revealed that hydrolytic enzymes (for example, amylase, chitinase and protease) from a variety of fungi were effective in enhancing Monascus pigment production.11 The authors suggested that hydrolysis of Monascus cell walls stimulates a defense mechanism resulting in overproduction of hydrophobic substances, such as pigments, in order to block enzyme attack.11 Melanins also protect fungi from hydrolytic enzymes, UV, solar or gamma radiation, extreme temperatures, heavy metals and several other toxic compounds.12 Similarly, Phomopsis sp. FKA-62 also had the potential to produce black pigments (probably melanin), apparently in order to protect against the antifungal compound BE-31405.
We discovered that a co-culture broth of Talaromyces siamensis FKA-61 and Phomopsis sp. FKA-62 was potently inhibitive of FKA-62, but pure culture broths produced no such activity. We also identified BE-31405 as an antifungal agent. When strains FKA-61 and FKA-62 were co-cultured on a PDA plate, FKA-62 was not seen to approach FKA-61. Furthermore, BE-31405 was active against FKA-62 but not active against FKA-61. We therefore predicted that FKA-61 was capable of producing BE-31405. For validation of this hypothesis, FKA-61 and FKA-62 were co-cultured with eight different fungal strains. Only strain FKA-61 produced BE-31405 when co-cultured with six of the eight strains tested. In contrast, FKA-62 did not produce this compound, on its own or in co-culture with any fungal strains.
When strains FKA-61 and FKA-62 were co-cultured on a PDA plate, FKA-62 produced a black pigment. However, when these strains were co-cultured in liquid medium, an orange pigment was produced. When a co-culture broth was grown on a PDA plate, strain FKA-61 grew relatively better from day 1, even though FKA-62 generally grows faster than FKA-61. Moreover, strain FKA-62 produced a visible black pigment on day 6. In liquid medium, FKA-62 does not produce black pigment. In addition, BE-31405 was detected from day 1 of a co-culture broth by HPLC (data not shown). This clearly suggests that strain FKA-61 was capable of detecting and suppressing FKA-62 by producing the antifungal BE-31405 at a very early stage.
BE-31405 was active against six fungal strains that possessed the ability to induce BE-31405 production when co-cultured with strain FKA-61. It would therefore appear that strain FKA-61 is capable of detecting BE-31405-sensitive fungi and producing the compound to minimize competition. It has been reported that arbuscular mycorrhizal fungi recognize their host plant using signalling molecules (strigolactones and 5-deoxy-strigol) from the root exudates of legume plant Lotus japonicus.13 It seems reasonable to assume that strain FKA-61 has the ability to recognize fungi as potential competitors perhaps though similar mechanisms, such as signal compounds. However, co-culture in a dialysis-culture flask did not result in the appearance of BE-31405. Consequently, another mechanism must be responsible, possibly the necessity of direct contact between hyphae of each fungus. Recently, Schroeckh et al.14 discovered that intimate physical interaction of Aspergillus nidulans with the soil-dwelling bacterium Streptomyces rapamycinicus specifically activated silent fungal secondary metabolism genes, resulting in the production of the archetypal polyketide orsellinic acid and its derivatives. The Saga/Ada complex containing the histone acetyltransferase is required for induction of the orsellinic acid gene cluster by the bacterium, and the Saga/Ada-dependent increase of histone 3 acetylation at lysine 9 and 14 occurs during interaction of fungus and bacterium.15 In our research, there is a possibility that a similar phenomenon occurred during interaction of the two fungi. The entomopathogenic fungus, Beauveria bassiana, is also capable of recognizing insects. The lectin, a kind of carbohydrate-binding glycoprotein, on the conidial surface of B. bassiana interacts with the insect curticle.16 Thus strain FKA-61 may recognize a fungal competitor by a physical difference in hyphal surface structure or hyphal component.
Strain FKA-61 does not take any action against Talaromyces pinophilus FKI-5653 and Trichoderma harzianum FKI-5655. FKI-5653 is of the same genus (Talaromyces) as FKA-61 but is a different species. In a previous study, we found an enhancement of metabolite productivity of Talaromyces pinophilus FKI-5653 when it was co-cultured with Trichoderma harzianum FKI-5655.4 Furthermore, all metabolites of FKI-5653 were not active against FKI-5655.4 This suggest that the genera Trichoderma and Talaromyces can exist alongside each other. Now we are trying to identify the mechanism by which strain FKA-61 is able to recognize other fungal species and how its chemical defenses are triggered. We are also proceeding with co-culture experiments with a view to discover yet more novel antifungal agents.
Methods
General
NMR spectra were measured on a Varian XL-400 spectrometer with 1H NMR at 400 MHz and 13C NMR at 100 MHz (Agilent Technologies, Santa Clara, CA, USA). The chemical shifts are expressed in p.p.m. and are referenced to the solvent CD3OD (3.30 p.p.m.) in the 1H NMR spectra and to the solvent CD3OD (49.0 p.p.m.) in the 13C NMR spectra. FAB-MS and ESI-MS spectra were measured on a JEOL JMS AX-505 HA mass spectrometer and a JEOL AccuTOF apparatus (JEOL, Akishima, Tokyo, Japan).
Antimicrobial activities
Antimicrobial activities of culture broths against representative microbes
Antimicrobial activity was measured using the paper disc method (8 mm, ADVANTEC, Tokyo, Japan). Samples contained 50 μl of 50% EtOH extract of pure culture broths of strains FKA-61 and FKA-62, as well as the co-culture broth. Culture conditions were as follows: Bacillus subtilis ATCC 6633 (Davis synthetic medium (0.7% K2HPO4, 0.2% KH2PO4, 0.5% sodium citrate, 0.1% ammonium sulfate, 0.2% glucose, 0.01% MgSO4· 7H2O and 0.8% agar), 1.0% inoculation, 37 °C, 24 h), Kocuria rhizophila ATCC 9341 (Nutrient agar (0.5% peptone, 0.5% meat extract and 0.8% agar), 0.2% inoculation, 37 °C, 24 h), Escherichia coli NIHJ (Nutrient agar, 0.5% inoculation, 37 °C, 24 h), Xanthomonas camestris pv. oryzae KB 88 (Nutrient agar, 1.0% inoculation, 37 °C, 24 h), Mucor racemosus IFO 4581 (GY agar (1.0% glucose, 0.5% yeast extract and 0.8% agar adjusted in pH 6.0), 0.3% inoculation, 27 °C, 48 h), Talaromyces siamensis FKA-61 (PDA (Difco), 0.5% inoculation, 27 °C, 48 h), and Phomopsis sp. FKA-62 (PDA, 0.5% inoculation, 27 °C, 72 h).
Antifungal activity of BE-31405 against co-cultured fungal strains
Antifungal activity of BE-31405 against each fungal strain using a co-cultured broth was also measured by the paper disc method (8 mm, ADVANTEC). Paper discs contained varying amounts of BE-31405. Culture conditions were: Talaromyces siamensis FKA-61, Phomopsis sp. FKA-62, Phomopsis grabrae FKI-6685, Pestalotiopsis foedans FKI-6651, Trichoderma harzianum FKI-5655, Mariannaea elegans var. punicea FKI-6644, Perpreocillium lilacinum FKI-4665, Talaromyces pinophilus FKI-5653, Staphylotrichum boninense FKI-4751, and Gymnascella aurantiaca FKI-6588 (PDA (Difco) adjusted in pH 4.3, 0.5% inoculation, 27 °C, 48–72 h).
References
Pettit, R. K. Mixed fermentation for natural product drug discovery. Appl. Microbiol. Biotechnol. 83, 19–25 (2009).
Wang, J.-P., Lin, W., Wray, V., Lai, D. & Proksch, P. Induced production of depsipeptides by co-culturing Fusarium tricinctum and Fusarium begoniae. Tetrahedron Lett. 54, 2492–2496 (2013).
Bertrand, S. et al. Detection of metabolite induction in fungal co-cultures on solid media by high-throughput differential ultra-high pressure liquid chromatography-time-of-flight mass spectrometry fingerprinting. J. Chromatogr. A. 1292, 219–228 (2013).
Nonaka, K. et al. Enhancement of metabolites productivity of Penicillium pinophilum FKI-5653, by co-culture with Trichoderma harzianum FKI-5655. J. Antibiot. 64, 769–774 (2011).
Okada, H. et al. BE-31405, a new antifungal antibiotic produced by Penicillium minioluteum. I. Description of producing organism, fermentation, isolation, physico-chemical and biological properties. J. Antibiot. 51, 1081–1086 (1998).
Sassa, T., Aoki, H., Namiki, M. & Munakata, K. Plant growth promoting metabolites of Sclerotinia sclerotiorum. Part I. Isolation and structures of sclerotinin A and B. Agrc. Biol. Chem. 32, 1432–1439 (1968).
Grove, J. F., MacMillan, J., Mulholland, T. P. C. & Rogers, M. A. T. 759. Griseofulvin. Part I. J. Chem. Soc. 3949–3958 (1952).
Fuska, J., Uhrín, D., Proksa, B., Votický, Z. & Ruppeldt, J. The structure of vermistatin, a new metabolite from Penicillium vermiculatum. J. Antibiot. 39, 1605–1608 (1986).
Hayashi, H., Oka, Y., Kai, K. & Akiyama, K. New chrodrimanin congeners, chrodrimanins D-H, from YO-2 of Talaromyces sp. Biosci. Biotechnol. Biochem. 76, 1765–1768 (2012).
Kaifuchi, S. et al. Inducing pigmentation using Hypocrea/Trichoderma co-cultures. Jpn. J. Mycol. 53, 71–81 (2012).
Shin, C. S., Kim, H. J., Kim, M. J. & Ju, J. Y. Morphological change and enhanced pigment production of Monascus when cocultured with Saccharomyces cerevisiae or Aspergillus oryzae. Biotechnol. Bioeng. 5, 576–581 (1998).
Nosanchuk, J. D. & Casadevall, A. Impact of melanin on microbial virulence and clinical resistance to antimicrobial compounds. Antimicrob. Agents Chemother. 50, 3519–3528 (2006).
Akiyama, K., Matsuzaki, K. & Hayashi, H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 435, 824–827 (2005).
Schroeckh, V. et al. Intimate bacterial-fungal interaction triggers biosynthesis of archetypal polyketides in Aspergillus nidulans. Proc. Natl Acad. Sci. USA 106, 14558–14563 (2009).
Nützmann, H. W. et al. Bacteria-induced natural product formation in the fungus Aspergillus nidulans requires Saga/Ada-mediated histone acetylation. Proc. Natl Acad. Sci. USA 108, 14282–148287 (2011).
Sandhu, S. S. et al. Myco-biocontrol of insect pests: factors involved, mechanism, and regulation. J. Pathog. 2012, 126819 (2012).
Acknowledgements
This study was supported, in part, by funds from the Quality Assurance Framework of Higher Education from the Japan’s Ministry of Education, Culture, Sports, Science and Technology (MEXT) and a Kitasato University Research Grant for Young Researchers. We thank Dr. Yukihiro Asami (Kitasato University) for helpful comments on the manuscript and Dr. Kenichiro Nagai (Kitasato University) for measurements of MS.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Supplementary Information accompanies the paper on The Journal of Antibiotics website
Supplementary information
Rights and permissions
About this article

Cite this article
Nonaka, K., Iwatsuki, M., Horiuchi, S. et al. Induced production of BE-31405 by co-culturing of Talaromyces siamensis FKA-61 with a variety of fungal strains. J Antibiot 68, 573–578 (2015). https://doi.org/10.1038/ja.2015.28
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ja.2015.28
