
【Evolution of Tetraodontiformes】Pufferfishes, triggerfishes, boxfishes

Order Tetraodontiformes
Tetraodontiforms belong to Teleostei of actinopterygians, under Subdivision Percomorphaceae, Series Eupercaria, and finally comprise of the order Tetraodontiformes.[1]
Of course, if you were like me (or maybe not, since I’m like trash and ignorant), you might be wondering about where did all those taxonomic terms come from. Well, apparently, the nomenclatural statuses are shown following:
Class
↓
Subdivision (Subdivision Percomorphaceae)
↓
Series (Series Eupercaria)
↓
Order (Order Tetraodontiformes)
(Not to mention all the extra rankings not shown here like sections, cohorts, and the endless permutations by combining with prefixes like “mega” “super” “sub” “infra” uhhh. And now imagine me complaining the existence of tribes in bamboos? Lol.)
Bracketing the controversial taxonomic levels for Actinopterygii and Teleostei (thus why class up there remains empty), which are secondary for the purpose of this article, the classification of the percomorphs can only be said to be, well, messy. All sorts of contradicting phylogenetic analyses, clashes between taxonomy based on morphology and genetics, overlapping taxa, paraphylies, uncertain placements, you name it.[1] And as a member of the Percomorphaceae, the phylogeny of the tetraodontiforms naturally inherits this confusion.
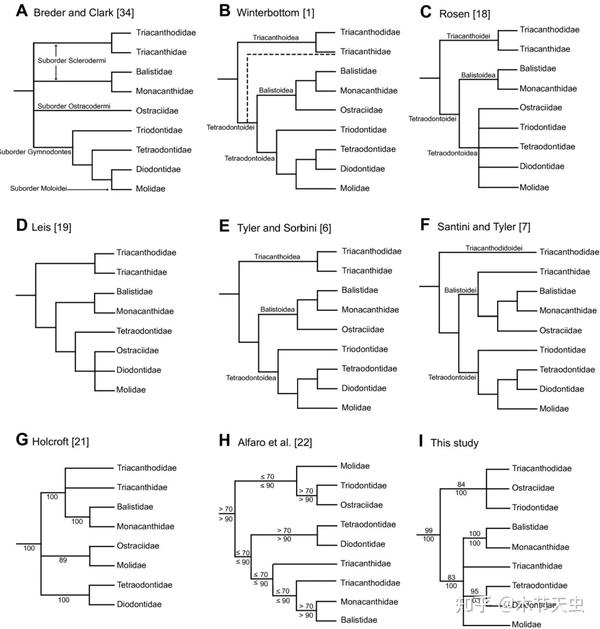

Caption: The pic every tetraodontiform channel uses displaying hypotheses for their evolutionary relationships. Until now, there have already been up to ten credible hypotheses about tetraodontiform phylogeny, and almost every one of them contradicts each other in details. It is easy to grasp just how messy evolution for percomorphs like tetraodontiforms are. Picture from [2].
However, for the purpose of this article, we must choose a phylogeny for our usage. Excuse the author for having no professional knowledge in judging which hypothesis is better than the other, but the only metric I can abide by is recency. And since to the author’s awareness the most recent one is [3], this will be the phylogeny and divergence dates that shall be adopted in this article. (For a summary of the different hypotheses for tetraodontiform phylogeny, see [4])


Caption: Evolutionary tree for the phylogeny chosen above, from appendix document of [3]. Do keep in mind that this is almost certainly not the whole picture since if another research about tetraodontiform evolution came out, it would most likely tell a different story. But at least the original article claims it “generates the most complete tetraodontiform phylogeny to date.” (Troyer et al.) So… [innocent-face-emoji]
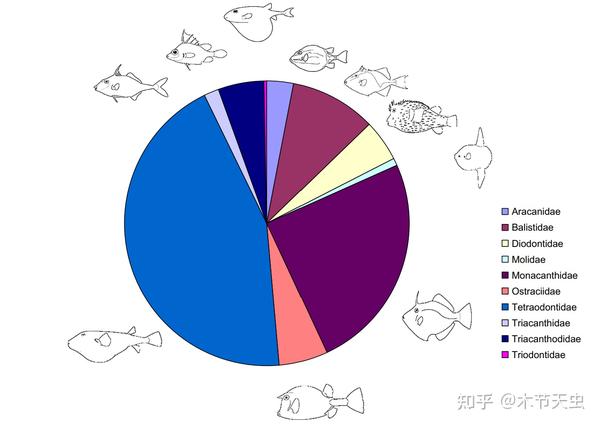

Caption: Tetraodontiformes as an order is extremely diverse in shape and structure. From the gargantuan sunfishes to the exotic threetooth puffers, they may only be one sister family apart. (which actually is true in Alfaro’s hypothesis[5]) Bro, this is just the dream species for evolutionary and comparative studies.[6] And tons of papers can be found on evolution alone for each of the ten different orders. Thus, it would not be surprising that this article fails to be an exhaustive and holistic evolutionary account, and some of the “interesting” information I share might just be trivial hypotheses mentioned in the arbitrary papers the author read. Please be aware of this, and the author is definitively not professional.
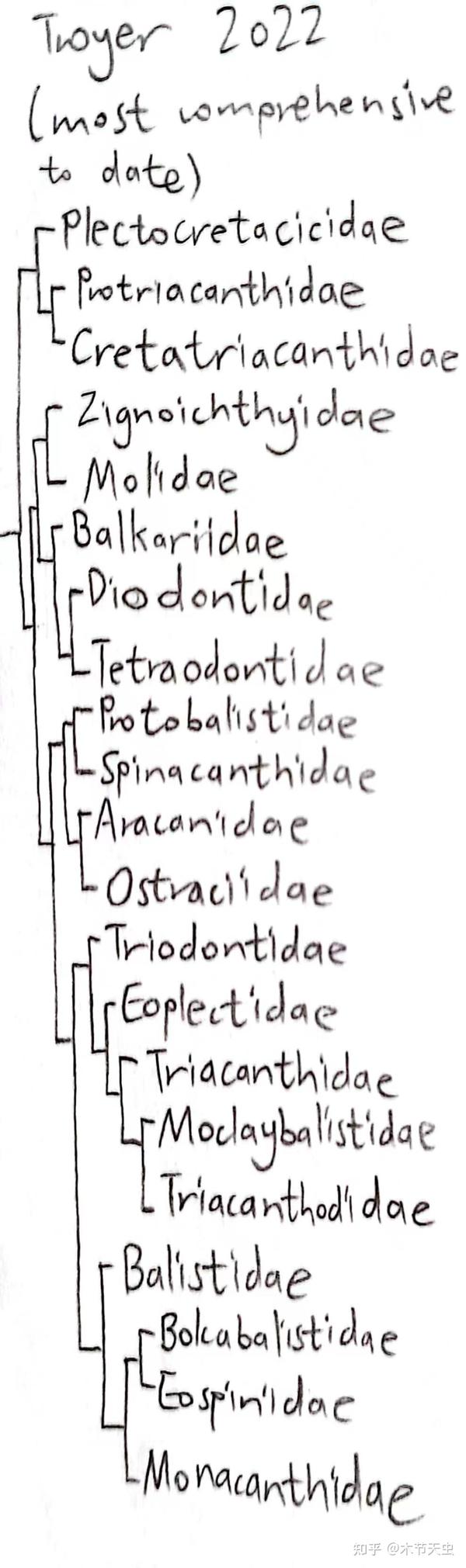



Caption: The subordinal classification for this article will follow [1]. As can be seen, many of those surborders contain only one family, and the author strongly suspects that this is because of the volatility of tetraodontiform interfamilial relationships that any attempt to classify multiple families into one suborder (except some of the pretty stable pairs, as will be elucidated after) will just be futile, so putting controversial families each within its own suborder is just a compromise.
The sister group of Tetraodontiformes is Lophiiformes, the anglerfishes, but as with everything, it is disputed.[7] Almost immediately after separating with the Lophiiformes at around the start of Late Cretaceous 100 million years ago, an extinct superfamily known as Plectocretacicoidea diverged from the rest, marking the first known tetraodontiforms. Some scientists have doubted the tetraodontiform identity of plectocretacicoids due to morphological evidence and how when not accepting plectocretacicoids as tetraodontiforms, the divergence for Tetraodontiformes is delayed another 40 million years at around 60 million years ago in the Paleocene.[3] However, here, we will accept plectocretacicoids as tetraodontiforms. (If plectocretacicoids really are tetraodontiforms, this makes me wonder just how inaccurate molecular dating on fossil-lacking lineages must be.)
Superfamily Plectocretacicoidea contains three fossil families, Plectocretacicidae, Protriacanthidae, and Cretatriacanthidae, dotted across the coast of the ancient Tethys.[8] Some might be dubious about how fossil records of Tetraodontiformes are able to appear so quickly after its origin. This is actually not as impossible as one thought since tetraodontiforms, blessed with hardened external anatomies and generally shallow habitats, have excellent fossilization potentials, making oldest fossil ages close to their origination dates.[3][9] Here are some pictures of plectocretacicoids:










From the structures of the species above, some of the evolutionary themes later tetraodontiforms are going to follow are already evident. For example protective armaments like scales and spines, the compromise on swimming speed that comes along, and many skeletal reductions (not directly visible in the reconstructions).[8] Despite only being a stem group, many of their cousins shall pick up their unique traits and end up having their original extinct relatives named after them. For example plectocretacicids after ostraciids and protriacanthids/cretatriacanthids named after triacanthids.
Despite the superficial similarity, tetraodontiforms in the Cretaceous only came in millimeters. This is partly because of a generally higher temperature in which the authors of [3] argue a lower oxygen dissolution limits tetraodontiform body size. But more likely the predominant reason is due to the peripheral niche tetraodontiforms reside in during an era of intense competition and predation, thus specialization visible in their features.[15][16] However, this is all about to change as a giant meteorite happens to find its way toward Earth.


Intriguingly, tetraodontiforms did not appear to suffer a major extinction during the End-Cretaceous Extinction. However, this is most likely because of absence of fossils at the boundary, and the plectocretacicoids most likely went extinct in this event, though it may have disappeared earlier during its last appearance in the Campanian.[18][15] During the entire Paleocene, only one species of tetraodontiforms was unearthed, Moclaybalistes danekrus of family Moclaybalistidae. However, as phylogenetic analysis did resolve moclaybalistids to be sister group with the triacanthodids, this is the official symbol for the appearance of crown tetraodontiforms.[3]


Caption: Fossil for Moclaybalistes of Moclaybalistidae, unearthed in Denmark. The distribution of literally every early tetraodontiform near the Peri-Tethys coast and the flourishing of Monte Bolca later on, also in Tethys, basically confirmed the Tethyan origin of Tetraodontiformes.[19] Picture from [20].
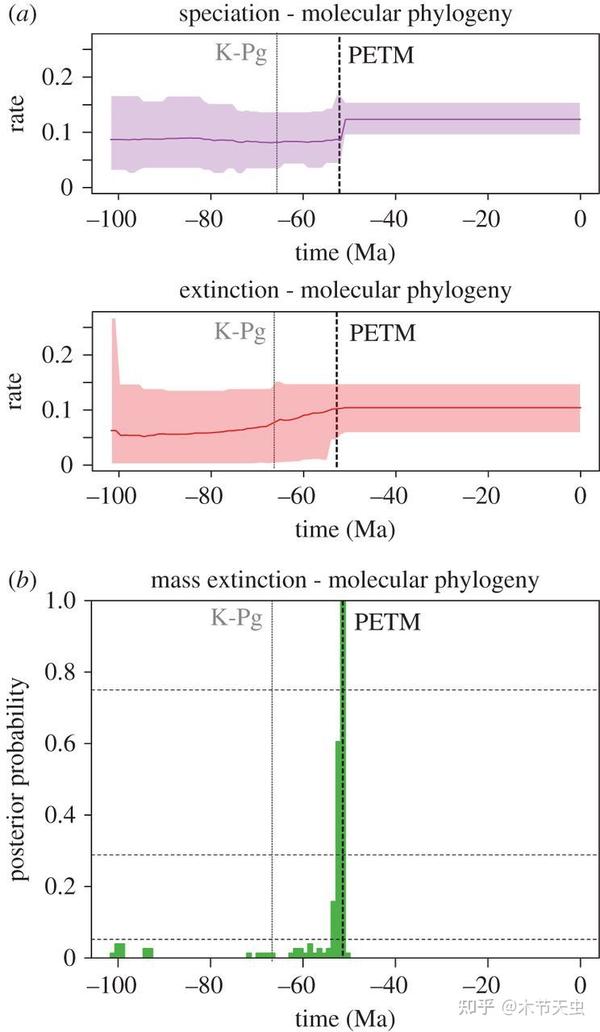

The Paleocene-Eocene Thermal Extreme appeared to have a pretty hard hit on tetraodontiforms, as seen in the figure above. But as the survivors started to thrive in the hot and favorable climate, they started to discover all the new niches and habitats created by the sudden temperature rise; the tetraodontiforms, as well as many other famous lineages all around the globe, began to explode.[18] All of the invisible lineages that have long diverged since the Cretaceous began to take on a plethora of diversities and forms, and leaving their records in the strata for the first time. After a few fossil occurrences at the earliest Eocene, like the sporadic raindrops foreshadowing a giant storm, time was about to come for the first climax of Tetraodontiformes, Italy, Monte Bolca.[6][21]
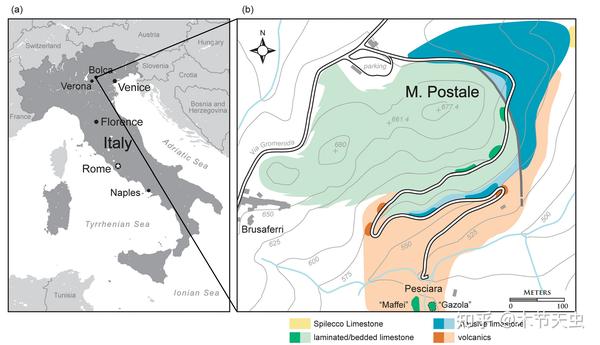

Caption: Location and map of the Monte Bolca Lagerstätten, its two major sites being Monte Postale and Pesciara. Pesciara is the site where the majority of Monte Bolcan tetraodontiform fossils were unearthed from.[22] Picture from [23].
It was the Ypresian of early Eocene, approximately 50 million years ago. The PETM has just occurred, introducing a wave of teleost invasions into reef habitats, filling up the coastal waters. In a tiny corner of the world, at a subtropical lagoon in the ancient Tethys, the anoxia from the thriving microbes and algae enabled a snapshot of this diversity be preserved to awe the eyes of modern ichthyologists.[23] Out of the ten living families of tetraodontiforms today, five were first discovered in Monte Bolca, along with a staggering seven extinct families.[22][3] All those point to a golden age of tetraodontiforms in the Tethys; they were ready to explore abroad.





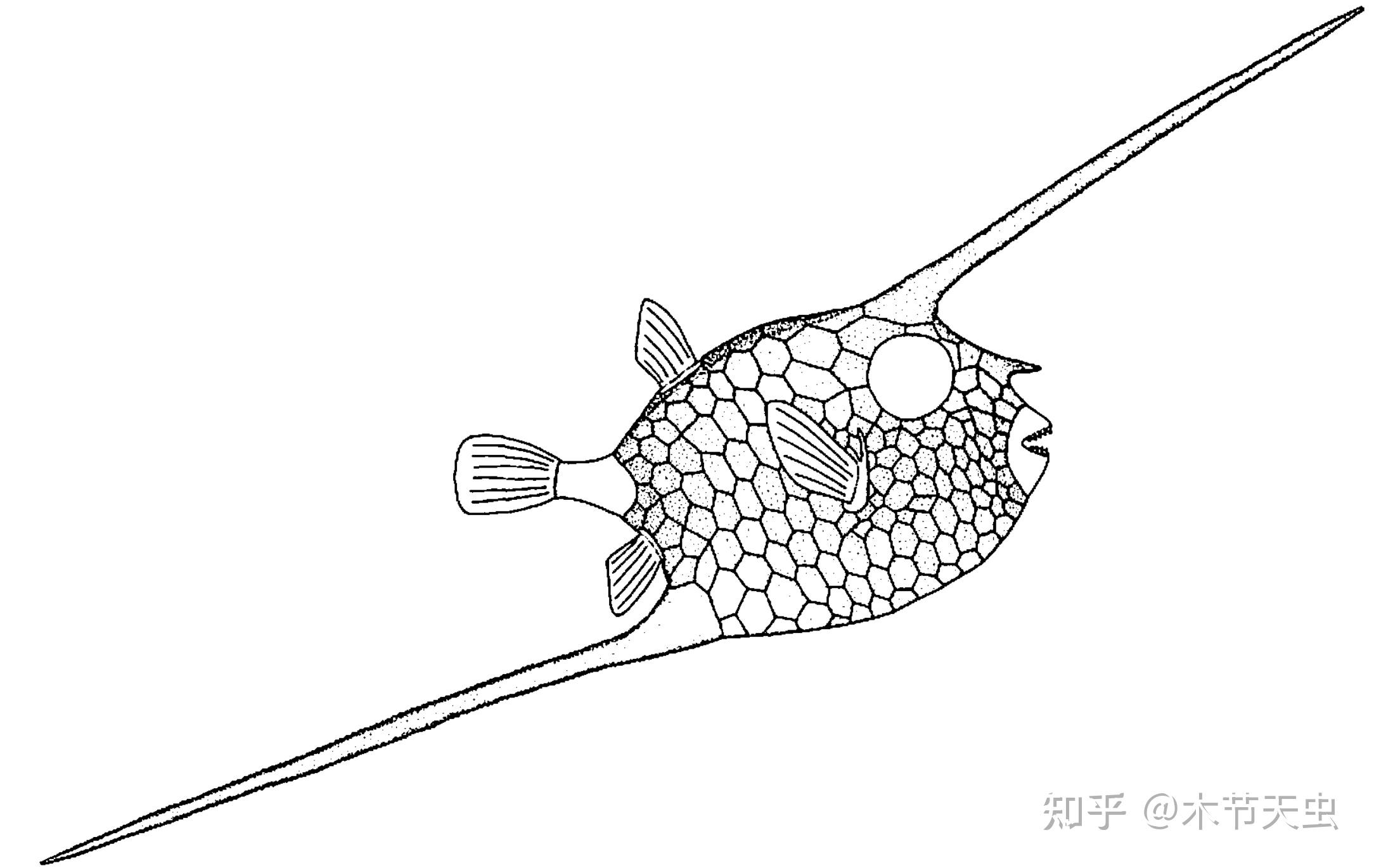
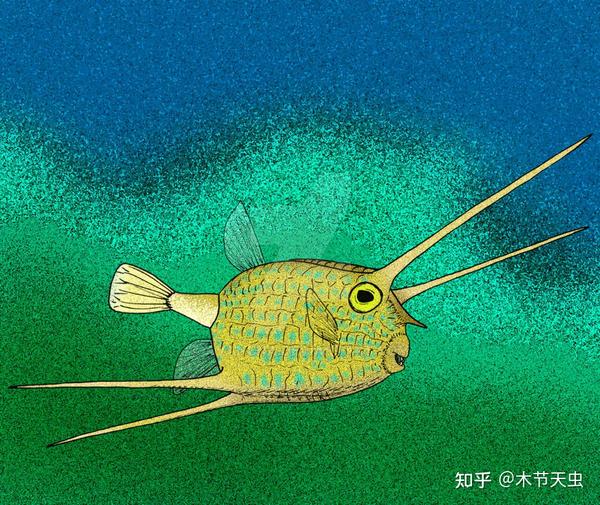



Caption: Another fossil tetraodontiform Zignoichthys of Zignoichthyidae. Unlike many of his coeval pioneers who shined then died out within the Eocene, zignoichthyids survived all the way to the Oligocene. But unfortunately, they failed to find a way of living suitable for themselves, resulting in convergent evolution with the spiny puffers and annihilated by their much more advantageous competitors.[16] Their highly-specialized body at the right seemed to leave its last deformed outcry as the zignoichthyids perished before dawn---every tetraodontiform family found after them shall be the one found today. Pictures from [27].
Do not “underestimate” Eocene tetraodontiform diversity from the fossils found in Monte Bolca though. Many of the families are in fact discriminated from the lagoonal environments in Monte Bolca, for example Molidae being pelagic and Triodontidae and Triacanthodidae living in deep waters.[22] And the orders Balistidae, Triodontidae, and Molidae in fact left their fossils elsewhere during the Eocene as well.[3] And guess who is excluded? Monacanthidae. It diversified too late and it just plain sucks, without any excuses.
As you read all of the above, it is natural to postulate that tetraodontiforms obviously originated in shallow-water habitats. The Monte Bolcans and even the plectocretacicoids are all shallow-dwellers, and the most parsimonious conclusion is just to say tetraodontiforms originated shallow.[19] However, it is worth noting that the agreed sister group of tetraodontiforms, the anglerfishes, have kept the deep-water tradition; sure, you can argue that some anglerfishes are shallow, but because of phylogenetic instabilities, you never know if the shallow ones are primitive or derived, thus unable to deduce whether the common ancestor of anglerfishes and tetraodontiforms are deep as well. [28] As with the fossil record, arguing that deep-water inhabitants are harder to be discovered in fossil record is also compelling. (plus plectocretacicoids not even sure whether to be tetraodontiforms) Uh, as you see, just endless controversies. Another evidence for deep-water origin is how one supposed basal group also lives a deepwater life, the triacanthodids.
Although not shown to be basal in our hypothesis, many hypotheses have shown the Triacanthodidae, most of the time with another family Triacanthidae, as the earliest diverging lineage of tetraodontiforms.[4] Thus, Triacanthodidae and Triacanthidae are traditionally grouped in the superfamily Triacanthoidea.[29] Their early divergence may be hinted by their relative lack of skeletal reductions characteristic of most tetraodontiforms.[28][30] Triacanthids are shallow; some triacanthodids are also shallow, but as with anglerfishes, you never know which life-mode came first.[28] Both families are characterized by their protruding spines (their common names triplespines/spikefish), reminiscent of some fossils we’ve seen and meant to deter predators from swallowing them.[15]


One interesting fact about the distribution of triacanthodids. Triacanthodidae is apparently divided into two subfamilies, Hollardinae and Triacanthodinae. The distribution of all hollardines is in West Atlantic, except one, which is in Hawaii. Meanwhile, the distribution of all triacanthodines is in Indo-Pacific, except for one species, which is in West Atlantic.[19] Traditionally, two Oligocene fossils from Southern Poland were thought to each belong to Hollardinae and Triacanthodinae, justifying each of their previous wide range. Even though our phylogeny now classifies both fossils to be triacanthodines,[3] the author still finds [28]’s hypothesis convincing which is that both subfamilies once had a much wider distribution covering Tethys (tetraodontiform origin after all) and their modern habitats. Hollardinae most likely dispersed to West Atlantic from Tethys in facilitation of the then-narrow Atlantic, and Triacanthodinae have spread throughout Indo-Pacific from Tethys via the then-separate Africa and Eurasia. The disjunct hollardine species in Hawaii probably dispersed from the Caribbean (note that marine species, unlike terrestrial ones, have excellent dispersal potentials, especially with larvae), and the disjunct triacanthodine species in Caribbean probably dispersed there from the Pacific as well, both before the closing of the Panama Isthmus and retaining a close relationship with the other members of each’s subfamily. Temperature fluctuations in the Neogene and Pleistocene, however, killed off most of the triacanthodids living on islands, which are especially sensitive to sea-level changes since they are trapped around island coasts and cannot actively seek suitable habitats elsewhere like those around continents, killing off the intermediate species and forming the distribution pattern today.
Another odd family in the tetraodontiforms are the triodontids. Only one species of this family survives to this day, hiding in the mysterious deeps.[15] However, they had a pretty glorious past, being the first extant tetraodontiform family to be unearthed in the fossil record (early Eocene Ctenoplectus williamsi),[3] and the Triodon genus, the genus the sole surviving triodontid member today belongs to, already had appeared back in the Eocene, signifying an appreciable amount of diversity for a deep-water lineage to appear twice in the fossil record. Well, according to [19], the reason for grouping the Eocene Triodon antiquus in Triodon is mainly a compromise for the lack of coeval triodontids unearthed and the general conservative evolution in triodontids, but still, for a family with only one remaining species, this is pretty impressive.


Triodontids are easily distinguishable by their pelvic bone connected to a pelvic fan, which they extend and flare to intimidate predators when threatened.[31] For some individuals, they would develop an eyespot on their pelvic fans, presumably to also intimidate predators.[15] Amazingly, triodontids appear to also be able to inflate, in the same mechanism as the more famous pufferfishes do by pumping water in their stomachs. However, their ribs haven’t devolved as much as pufferfishes do, meaning they can’t inflate into a ball, but their unique pelvic flaring does have an effect of expanding the abdominal cavity and aiding inflation.[31]


Ostracioidei is genuinely unique even among tetraodontiforms. Suborder Ostracioidei contains the orders Aracanidae and Ostraciidae. Aracanidae is argued to simply belong in Ostraciidae, but here we will accept the commoner view from [7] that they are separate.


The act of applying protective armaments in vertebrates is not uncommon, but to have one as inflexible as ostracioids’ is rare.[34][35] Therefore, the meme of how stacking armor only leads to demise is not necessarily untrue since the inflexible structure reduces evolutionary potentials, but for lineages who have found the ideal purpose as to apply their armor in nature, most notably turtles and ostracioids, it can bring a major success. For at least the past 35 million years, the ostracioids have thrived with their body structure.[35] They have low swimming speeds, and their anti-streamlined shape produces destabilizing vortices which enhance maneuverability but at the cost of directional swimming. Nevertheless, it is an optimal solution for their lifestyle in confined reef spaces.[34] The carapace of ostracioids also proves resistant toward penetrations and bites, enhancing their defense against the numerous reef-predators.[35]




Caption: Those are different species of Lactoria, but they generally symbolize how a Lactoria transform from a juvenile (left, from [36]) to an adult (right, from [37]). Juveniles typically have longer spines while adults’ become less prominent. I don’t know if this relates to you of an ostraciid having disproportionate spines, the Eolactoria we’ve looked at earlier. Well, there’s the possibility that the fossils unearthed are juveniles as well, and in reality, the spine proportion may not have been that dramatic as they grew larger.[27]
Next comes the balistoids, the balistids and the monacanthids. Even though the monacanthids appeared to have diverged pretty early, they were the last tetraodontiform family to appear in the fossil record, making its debut in Miocene.[15] Presumably, they were only a peripheral lineage in the time before Miocene, and they would have gone extinct long before humans will be alive to see just like so many of the other nameless groups that have once inhabited this Earth, if not for a lucky chance. That lucky chance is the diversification of modern reefs in the Late Oligocene and Miocene. Major divisions in reef-associated families like the balistoids and tetraodontoids all took place roughly during this period.[5][38] Monacanthidae was no exception. The next event diversifying tetraodontiform lineages to the species level would have to wait until the Pleistocene glaciation cycles.[39]
Meanwhile, their sister, balistids, in this burst of evolution, emerged with a diversity of skull shape and musculature. One of the most notable ones are the durophagous balistids, who have short and stout skulls to provide crushing force and muscles to forcefully close and stabilize the upper and lower jaws.[40] This brings about one of the highest jaw mechanical advantage among fishes,[40] its reflection on real-life being the notorious reputation of biting people’s fingers off.[41]


Caption: A Miocene balistid, Balistes vegai. This behemoth is almost two meters long, more than twice of the largest balistids today. From [42]. In addition to having powerful jaws, they actually use them actively by possessing one of the most aggressive territoriality among tetraodontiforms.[43] Now imagine swimming with them.
I have been describing the balistids as a brute, but honestly there are some elegancies among them. Tetraodontiformes is an innovative family after all. For example, balistoids are very attentive on their fin elements. Unlike their sister balistids who directionally devolved pelvic fins, many monacanthids retained them, signifying an evolutionary advantage of monacanthids protecting themselves from currents and predators by wedging themselves in reefs and crevices. Fins are also very useful in sexual selection when balistoids manipulated intricately coordinated fin mechanics to produce some elegant mating displays.[44]
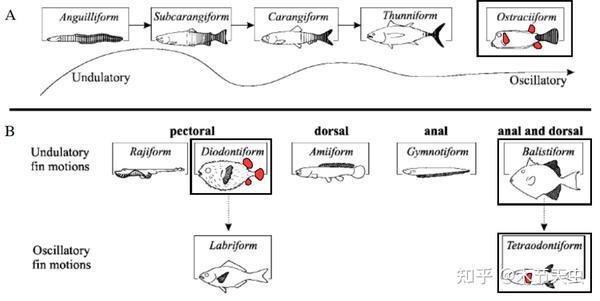

Caption: A diagram showing all the classical swimming modes fishes have taken to swim, with the ones represented by tetraodontiforms boxed in black. From [40]. As can be seen here, the morphological diversity of tetraodontiforms is among the first even in the perspective of the entirety of fishes, taking up four slots among the little swimming modes there are. Incredible.




Now, let’s leave our finale to the most populous of all tetraodontiforms, the Tetraodontoidei. Along with Ostracioidei and Balistoidei, those three are the three pairs of families whose sister relations have remained consistent throughout all phylogenetic analyses, which is admittedly pretty rare. Likewise, Tetraodontoidei has two families, the Tetraodontidae, the “smooth puffers,” (even though their common name is smooth, they still have tiny spines in the same way diodontids have spines, only less prominent[15]) and the Diodontidae, the “spiny puffers” which fit our stereotype of spiny pufferfishes.[2]
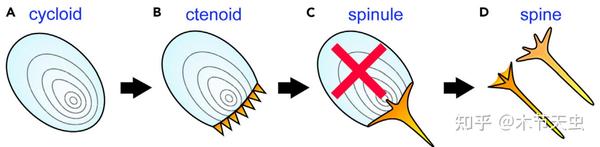

Caption: Scales in fishes have gone through many rounds of modifications, and the spinoid scale of the third picture represents the plesiomorphy of tetraodontiforms, a trait which many groups still retain. In Tetraodontoidei, the base of the scale is removed, leaving only the spine and forming the spines homologous to scales in other fishes you see on tetraodontoids today. Meanwhile, for ostracioids and molids, the spine is removed, and the bases fused to form their protective carapace. Picture from [32].
Naturally judging from each of their names, one of their most iconic abilities is that they puff. In fact, the puffing ability itself is very rare among fishes, but Tetraodontiformes manages to have three independent origins of puffing behavior within one order, namely tetraodontoids, some monacanthids, and triodontids,[45][31] definitively indicating an institutional disposition toward evolving it among tetraodontiforms. Among the three, however, tetraodontoids have gone to extreme measures to make their puffing optimal, such as devolving much of their ribs and very stretchy skin with collagen. The inflation behavior probably arose from practices like coughing water and water-blowing, a prevalent trait in tetraodontiforms. Subtle muscular mutations caused the compression of the cheeks intended to expel water out instead forced the water into the stomach.[45] With an additional feature of strong sphincters in the esophagus preventing pressurized water going into the oral cavity interfering with gill breathing, a completed inflation is done.[46]


The evolution of those traits obviously is meant for deterring predators from swallowing them. But the inedible shape is still not enough to dissuade some predators from trying them out. Many got suffocated by them in the process. Even if the predators do manage to get them eaten, they probably would have died anyways due to the presence of a highly fatal toxin called tetrodotoxin, TTX, in adult tetraodontid liver and ovaries. Although passed down to larvae, in adults, the presence of such toxins is not at all identifiable before eating them, making it truly death on both sides.[47]

Caption: Picture from [15]. How tetrodotoxin poisons a predator to death and how inflation suffocates a predator are all examples of passively (note that it has to be passive, active is another matter) bringing the predator to death along with yourself. Other members of tetraodontiforms, such as ostracioids and triodontids, also don’t lack examples of poisoning or suffocating a predator at the expense of their own death. So this begs the question, what’s the point? Does this behavior of internecinity make sense evolutionarily? Let’s analyze the selective process: First, a bunch of inflating pufferfishes, predator looks at them and doesn’t think it can ingest them, predator leaves and pufferfishes survive, survival rate of those that can inflate increases; even if some predators do try to eat them, they would suffocate and in turn put a selective pressure on the rest of the predators to have a disinclination toward eating the other pufferfishes, which forms a positive feedback cycle and enhancing successful evolution. For tetrodotoxin it’s the same mechanism. Research [47] has already shown that the presence of tetrodotoxin on larval skin enables predators to spit poisonous pufferfishes out before ingesting and killing the pufferfish, analogous to how predators are dissuaded to eat by looking at a pufferfish’s inedible size alone. The act of killing both yourself and predator must have a general advantage on the whole population with this act (like eliminating those who have a tendency to eat pufferfishes) in order for this act to be advantageous, and this means that those having this act must already have a sizeable population. Otherwise, a mutation bringing one poisonous individual who goes on to die alongside a predator eating it will not bring any good. (This is where the dissuasion of a predator from eating them before death of oneself comes useful. This gives individuals with that trait time to build up a sizeable population, where then sporadic deaths would be beneficial) This also means that those with an advantageous trait must have some characteristic that distinguishes them having this trait in order for natural selection of the predator to have something to act on. Ok, we have already discussed three scenarios of killing yourself to kill the predator: predator knowing that a prey is inedible before eating it (most direct natural selection); predator not knowing whether a prey is inedible or not (natural selection is pointless); and how an inedible prey has some characteristic that characterizes their inedibility (this type must already have a sizeable population before being forced to die together with the predator for natural selection to act on the predators.[48] In time, the predators would develop a disinclination toward this trait from the deaths of those who lack this disinclination). The inflation of pufferfishes and poison of pufferfish larvae correspond to the first type, the poison of adult pufferfishes being the second type, and warning coloration being the third type.[49] Because there lacks a specific noun denoting the meaning of someone who kills a predator at the expense of the death of itself, the author is having hard time locating researches dealing this topic. However, the author knows that it is impossible for such a popular evolutionary game theory to have no studies on it, so if any professional knows any resource, the author would be more than grateful if it could be provided. In the end, all the above is just reasonable deduction and tootle from the author alone, and everybody could well take it as my personal studying notes. Thanks.
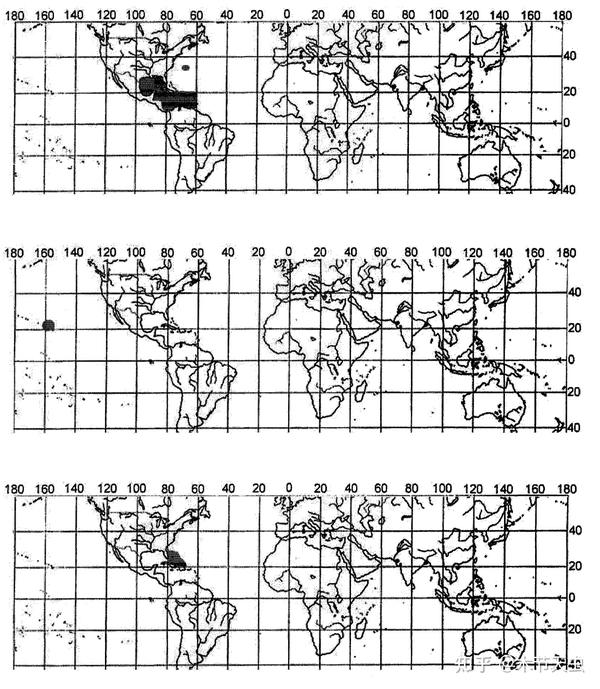
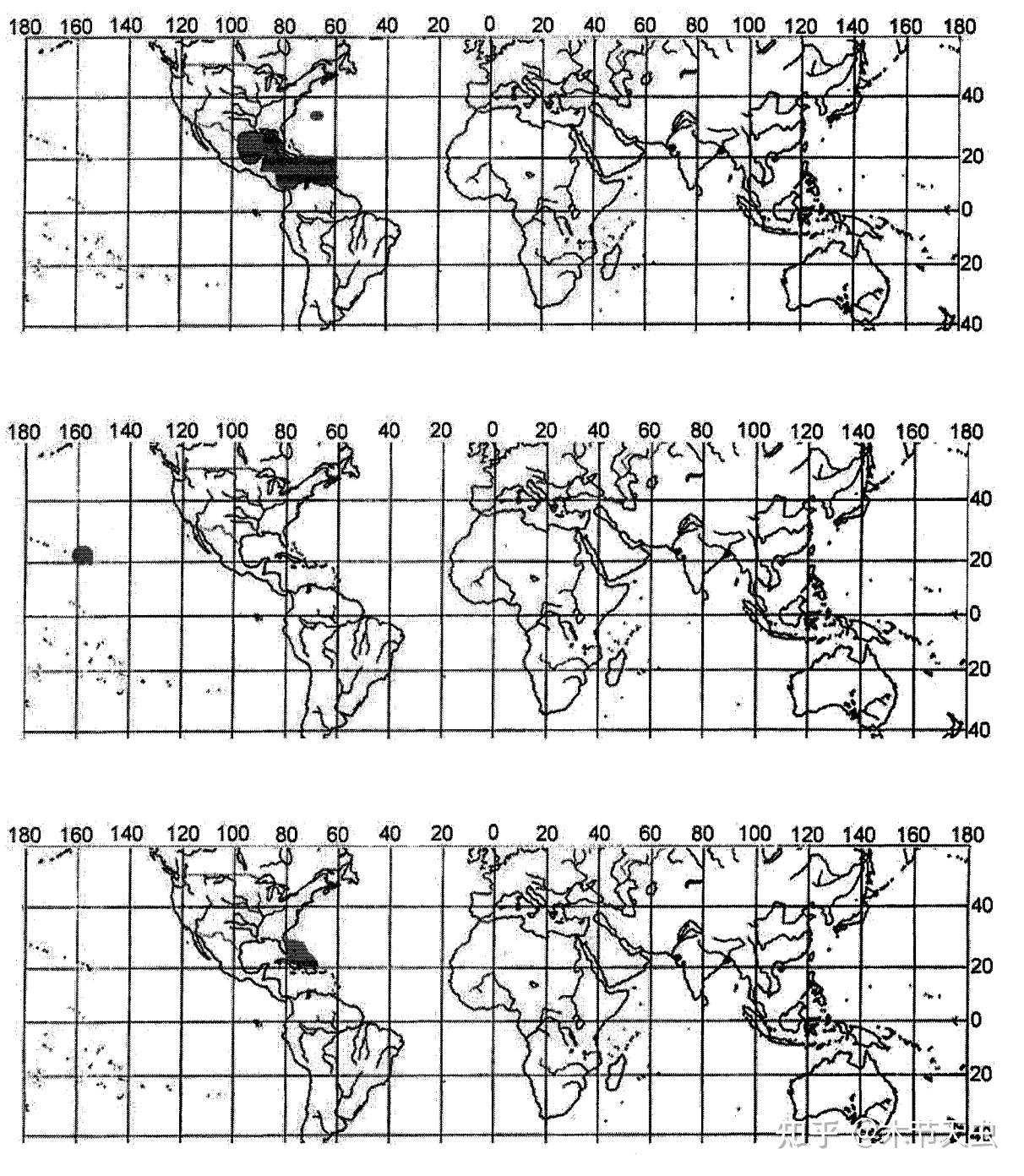
Tetraodontidae is the largest family of Tetraodontiformes, housing a total of 184 species.[7] One reason that adds to its species count is its being the only tetraodontiform family to completely discard its marine lifestyle and colonize freshwater.[15] Through three opportunities, namely the Miocene marine incursion into Northern South America, the end of the marine incursion of what is now Western Sahara, and the collision of Tibet and India. The significance of such incursions is that what once was sea became land, and the native pufferfishes would have been forced into freshwater ecosystems. In fact, many marine and brackish tetraodontids already have the tolerance of surviving in freshwater environments, and even spend some time of their lives, for example, spawning in them (like Takifugu[41]), and the reason freshwater transitions haven’t happened frequently is because of occupied niches in already-existing freshwater ecosystems, which is solved in the aforementioned geological events that created brand-new freshwater ecosystems which pufferfishes can colonize first and settle down. This results in the modern three disjunct tetraodontid freshwater distributions in South America, Central Africa, and Southeast Asia.[50]
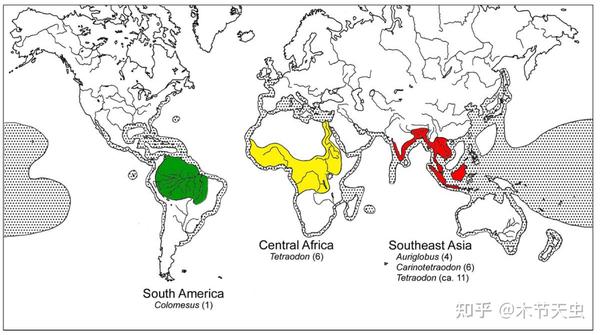

Caption: Distribution for Tetraodontidae. The dotted areas are the marine distributions. The three freshwater distributions are also indicated. Southeast Asian freshwater has the most diverse pufferfishes, with Central Africa being second and South America being least, having the shortest diversifying history. Picture from [50].
Despite this, freshwater counts do not have that much of a contribution on tetraodontid species-richness. Freshwater lineages still inevitably have lower rates of speciation than their marine counterparts. For explosive radiation of Tetraodontidae, one shall still look at the Takifugu. Potentially driven by the Pleistocene ice cycles, Takifugu distribution covered an area from Southern Australia to Japan and being one of the youngest and most speciose genera in Tetraodontidae.[39] Besides this, tetraodontids are accompanied by another amazing feat, possessing the smallest vertebrate genome.[51]


Caption: Although not aligned with our adopted phylogeny, this shows the known genome size for each tetraodontiform family at the time. As can be seen, not only do tetraodontids have exceptionally small genomes, the balistoids also possess small genomes relative to the rest of Vertebrata. The genomes of two tetraodontids have already been completely sequenced.[44] I mean, why not, it’s so convenient, and just ironic for another reason of the tetraodontiforms being the perfect model group. Figure from [52].
Through studies and comparisons, the decreased tetraodontid genome size is mostly through deletion of small non-coding sequences and repetitive elements.[53] Tetraodontid sister, diodontids, have up to two times the genome size of tetraodontids. Therefore, it can be safely assumed that, whether through actively deleting those genes or losing the ability to accumulate those genes, the small genome is a result of a derived characteristic of tetraodontids.[51] Unfortunately, however, no research, as far as I found, has been dedicated to the evolutionary selective mechanism for the small genome. Therefore, I formulated a hypothesis myself. Tetraodontiforms with both small and large genomes share and overlap similar niches, so it is unlikely that the small genome is a result of selection to provide advantages like faster regeneration rates.[52] And since the relatively rare trait of having small genomes is found independently in two lineages within Tetraodontiformes, the author is more convinced of certain genetic tendency that predisposes tetraodontiform genomes to become small without external selection having to do with it. The ancestors of tetraodontids and balistoids had common mutations that triggered this mechanism, resulting in the small genome as genome sizes between different families began to drift apart with the millions of years of independent evolution.


However, as the tetraodontiforms radiated and dispersed out throughout the world, their position at Tethys was in fast decline. The vicissitude of time leaves only four endemic families to the Mediterranean, the remnant of the Tethys, while its other emigrants set journey to other parts of the world and abandoned their homeland forever.[15] But exploration means chance. After all, tetraodontiforms do represent diversity and distribution range not fortunate enough to be present in most other fish lineages. Even though I personally don’t agree with some’s view of tetraodontiforms being one of the “highest” and “advanced” lineages among ray-finned fishes since all extant organisms have been through the same time of evolution, but if the final result is for the affection of tetraodontiforms, what does it matter really?
Finally, apologies for not including molids in this article. Actually, the author first had the idea to write about tetraodontiforms because of molids, but due of that, I want to devote a pretty large section toward them, but currently the space limit doesn’t allow. So I will probably send a second part of tetraodontiforms, on the evolution of molid. Hopefully soon. Thanks for reading here, if any, bye.
Works Cited
[1] Betancur-R, Ricardo, Wiley, Edward, et al. “Phylogenetic Classification of Bony Fishes.” BMC Evolutionary Biology, vol. 17, no. 162, 2017, pp. 1-40. Accessed 28 May 2023.
[2] Yamanoue, Yusuke, et al. “A New Perspective on Phylogeny and Evolution of Tetraodontiform Fishes (Pisces: Acanthopterygii) Based on Whole Mitochondrial Genome Sequences: Basal Ecological Diversification?” BMC Evolutionary Biology, vol. 8, no. 212, 19 Jul 2008, pp. 1-14. Accessed 4 Jun 2023.
[3] Troyer, Emily, et al. “The Impact of Paleoclimatic Changes on Body Size Evolution in Marine Fishes.” PNAS, vol. 119, no. 29, Jul 2022, pp. 1-9. Accessed 4 Jun 2023.
[4] 木节天虫. “自己发个动态来当参考资料[doge],要不然真的太杂了[笑哭].” Bilibili, 4 Jun 2023, http://www.bilibili.com/opus/803288214416654370?spm_id_from=444.41.0.0. Accessed 4 Jun 2023.
[5] Alfaro, Michael, et al. “Do Reefs Drive Diversification in Marine Teleosts? Evidence from the Pufferfishes and Their Allies (Order Tetraodontiformes).” Evolution, vol. 61, no. 9, 15 Aug 2007, pp. 2104-2126. Accessed 4 Jun 2023.
[6] Close, Roger, et al. “Mosaicism in a New Eocene Pufferfish Highlights Rapid Morphological Innovation near the Origin of Crown Tetraodontiforms.” Palaeontology, vol. 59, no. 4, 10 Jun 2016, pp. 499-514. Accessed 4 Jun 2023.
[7] Matsuura, Keiichi. “Taxonomy and Systematics of Tetraodontiform Fishes: A Review Focusing Primarily on Progress in the Period from 1980 to 2014.” Ichthyological Research, vol. 62, 11 Nov 2014, pp. 72-113. Accessed 4 Jun 2023.
[8] Tyler, James and Sorbini, Lorenzo. “New Superfamily and Three New Families of Tetraodontiform Fishes from the Upper Cretaceous: The Earliest and Most Morphologically Primitive Plectognaths.” Smithsonian Contributions to Paleobiology, vol. 82, no. 82, Jan 1996, pp. 1-69. Accessed 4 Jun 2023.
[9] Arcila, Dahiana, et al. “An Evaluation of Fossil Tip-dating versus Node-age Calibrations in Tetraodontiform Fishes (Teleostei: Percomorphaceae).” Molecular Phylogenetics and Evolution, vol. 82, 24 Oct 2014, pp. 131-145. Accessed 4 Jun 2023.
[10] “Plectocretacicus-31.” Expo Hakel, http://expo-hakel.com/product/plectocretacicus-31/. Accessed 4 Jun 2023.
[11] Oilshale. “Plectocretacicus clarae.” The Fossil Forum, 26 Dec 2009, http://www.thefossilforum.com/index.php?/gallery/image/8168-plectocretacicus-clarae/. Accessed 4 Jun 2023.
[12] Avancna. “Plectocretacicus clarae.” Deviant Art, 13 Aug 2008, http://www.deviantart.com/avancna/art/Plectocretacicus-clarae-94724521. Accessed 4 Jun 2023.
[13] Avancna. “Protriacanthus gortanii.” Deviant Art, 13 Aug 2008, http://www.deviantart.com/avancna/art/Protriacanthus-gortanii-94698828. Accessed 4 Jun 2023.
[14] Avancna. “Cretatriacanthus guidotti.” Deviant Art, 27 Mar 2013, http://www.deviantart.com/avancna/art/Cretatriacanthus-guidotti-360497835. Accessed 4 Jun 2023.
[15] 闪光鱵. “从棘刺到剧毒:鲀形目的演化之路.” 微博, 15 Jul 2018, http://weibo.com/ttarticle/p/show?id=2309404262014041073811. Accessed 4 Jun 2023.
[16] PAI派崔克and跑跑散人Dominnic兰. “走叠甲一定短命?叠甲小鱼的逆袭之路———鲀形目的演化史.” Bilibili, 15 Oct 2021, http://www.bilibili.com/video/BV1dv411g7LV. Accessed 4 Jun 2023.
[17] Avancna. “Nardorex zorzini.” Deviant Art, 13 Sep 2016, http://www.deviantart.com/avancna/art/Nardorex-zorzini-629962216. Accessed 4 Jun 2023.
[18] Arcila, Dahiana and Tyler, James. “Mass Extinction in Tetraodontiform Fishes Linked to the Palaeocene-Eocene Thermal Maximum.” Proc Biol Sci, vol. 284, no. 1866, 15 Nov 2017, pp. 1-8. Accessed 4 Jun 2023.
[19] Santini, Francesco and Tyler, James. “A Phylogeny of the Families of Fossil and Extant Tetraodontiform Fishes (Acanthomorpha, Tetraodontiformes), Upper Cretaceous to Recent.” Zoological Journal of the Linnean Society, vol. 139, July 2003, pp. 565-617. Accessed 4 Jun 2023.
[20] Sylvestersten. “Pufferfish - Moclaybalistes danekrus.” The Fossil Forum, 10 May 2013, http://www.thefossilforum.com/index.php?/gallery/image/26072-pufferfish-moclaybalistes-danekrus/. Accessed 4 Jun 2023.
[21] Bannikov, A. et al. “A New Family of Gymnodont Fish (Tetraodontiformes) from the Earliest Eocene of the Peri-Tethys (Kabardino-Balkaria, Northern Caucasus, Russia).” Journal of Systematic Palaeontology, vol. 15, no. 2, Jan 2017, pp. 129-146. Accessed 4 Jun 2023.
[22] Tyler, James and Santini, Francesco. “Reviews and Reconstructions of the Tetraodontiform Fishes from the Eocene of Monte Bolca, Italy, with Comments on Related Tertiary Data.” Studi e Ricerche sui Giacimenti Terziari di Bolca, vol. 9, Jan 2002, pp. 47-119. Accessed 4 Jun 2023.
[23] Friedman, Matt and Carnevale, Giorgio. “The Bolca Lagerstätten: Shallow Marine Life in the Eocene.” Journal of the Geological Society, vol. 175, no. 4, 8 May 2018, pp. 569-579. Accessed 4 Jun 2023.
[24] Avancna. “Protobalistum imperiale.” Deviant Art, 10 Oct 2010, http://www.deviantart.com/avancna/art/Protobalistum-imperiale-182133059. Accessed 4 Jun 2023.
[25] Avancna. “Eospinus daniltshenkoi v1.” Deviant Art, 21 Jul 2008, http://www.deviantart.com/avancna/art/Eospinus-daniltshenkoi-v1-92341522. Accessed 4 Jun 2023.
[26] Avancna. “Eolactoria sorbinii.” Deviant Art, 17 Mar 2010, http://www.deviantart.com/avancna/art/Eolactoria-sorbinii-157521304. Accessed 4 Jun 2023.
[27] 海洋欢乐谷. “蒙特波卡:始新世的海洋之窗(4)——真鲈形系鲀形目(作者:达达).” 知乎, 17 Jul 2019, http://zhuanlan.zhihu.com/p/73974774. Accessed 4 Jun 2023.
[28] Santini, Francesco. Phylogeny and Biogeography of the Triacanthodidae. University of Toronto, 2003. Accessed 4 Jun 2023.
[29] Santini, Francesco, et al. “A New Phylogeny of Tetraodontiform Fishes (Tetraodontiformes, Acanthomorpha) Based on 22 Loci.” Molecular Phylogenetics and Evolution, vol. 69, no. 1, Oct 2013, pp. 177-187. Accessed 4 Jun 2023.
[30] Santini, Francesco and Tyler, James. “Phylogeny and Biogeography of the Extant Species of Triplespine Fishes (Triacanthidae, Tetraodontiformes).” Zoologica Scripta, vol. 31, no. 4, Jul 2002, pp. 321-330. Accessed 4 Jun 2023.
[31] Bemis, Katherine, et al. “Pelvic-fan Flaring and Inflation in the Three-Tooth Puffer, Triodon macropterus (Tetraodontiformes: Triodontidae), with Additional Observations on Their Behavior in Captivity.” Ichthyology & Herpetology, vol. 111, no. 2, 18 Apr 2023, pp. 222-240. Accessed 4 Jun 2023.
[32] Shono, Takanori, et al. “Evolution and Developmental Diversity of Skin Spines in Pufferfishes.” iScience, vol. 19, 27 Sep 2019, pp. 1248-1259. Accessed 4 Jun 2023.
[33] Dunson, Bill. “Coastal Marvels of Nature in SW Florida.” Lemon Bay Conservancy, 24 Nov 2017, http://lemonbayconservancy.org/coastal-marvels-nature-sw-florida/. Accessed 4 Jun 2023.
[34] Van Wassenbergh, S., et al. “Boxfish Swimming Paradox Resolved: Forces by the Flow of Water around the Body Promote Manoeuvrability.” Journal of the Royal Society Interface, vol. 12, no. 103, 6 Feb 2015, pp. 1-11. Accessed 4 Jun 2023.
[35] Yang, Wen, et al. “The Armored Carapace of the Boxfish.” Acta Biomaterialia, vol. 23, 1 Sep 2015, pp. 1-10. Accessed 4 Jun 2023.
[36] Berthomier, Jack. “Lactoria cornuta – (Linnaeus, 1758).” Image du Monde, http://www.picture-worl.org/actinopterygien-nouvelle-caledonie-9d-lactoria-cornuta-linnaeus-1758.html. Accessed 4 Jun 2023.
[37] Shaw, Ian. “Lactoria fornasini.” Reef Life Survey, http://reeflifesurvey.com/species/lactoria-fornasini/. Accessed 4 Jun 2023.
[38] Santini, Francesco, et al. “New Multi-locus Timescale Reveals the Evolutionary Basis of Diversity Patterns in Triggerfishes and Filefishes (Balistidae, Monacanthidae; Tetraodontiformes).” Molecular Phylogenetics and Evolution, vol. 69, no. 1, Oct 2013, pp. 165-176. Accessed 4 Jun 2023.
[39] Santini, Francesco, et al. “Do Habitat Shifts Drive Diversification in Teleost Fishes? An Example from the Pufferfishes (Tetraodontidae).” Journal of Evolutionary Biology, vol. 26, no. 5, 15 Mar 2013, pp. 1003-1018. Accessed 4 Jun 2023.
[40] George, Andrew, and Westneat, Mark. Functional Morphology and Evolution of the Balistiform Swimming Mode. University of Chicago, 2020. Accessed 4 Jun 2023.
[41] 龙女之声and果酱白鱀不是白框. “体重三吨速不如人 压力一大毒死自己 长着人牙却比鲨鱼还要危险,专注叠甲的鱼有多强【鲀形目】.” Bilibili, 18 Feb 2023, http://www.bilibili.com/video/BV1Fe4y1c7vk. Accessed 4 Jun 2023.
[42] HodariNundu. “Diving with Balistes vegai.” Deviant Art, 28 Feb 2019, http://www.deviantart.com/hodarinundu/art/Diving-with-Balistes-vegai-787555982. Accessed 4 Jun 2023.
[43] Deo9. “鳞鲀对其他鱼类的竞争优势.” Bilibili, 26 Jul 2021, http://www.bilibili.com/read/cv12330743. Accessed 4 Jun 2023.
[44] Yamanoue, Yusuke, et al. “Unique Patterns of Pelvic Fin Evolution: A Case Study of Balistoid Fishes (Pisces: Tetraodontiformes) based on Whole Mitochondrial Genome Sequences.” Molecular Phylogenetics and Evolution, vol. 50, no. 1, Jan 2009, pp. 179-189. Accessed 4 Jun 2023.
[45] Wainwright, Peter and Turingan, Ralph. “Evolution of Pufferfish Inflation Behavior.” Evolution, vol. 51, no. 2, Apr 1997, pp. 506-518. Accessed 4 Jun 2023.
[46] McGee, Georgia and Clark, Timothy “All Puffed Out: Do Pufferfishes Hold Their Breath while Inflated?” Biology Letters, vol. 10, no. 12, Dec 2014, pp. 1-4. Accessed 4 Jun 2023.
[47] Itoi, Shiro, et al. “Role of Maternal Tetrodotoxin in Survival of Larval Pufferfish.” Toxicon, vol. 148, 15 Jun 2018, pp. 95-100. Accessed 4 Jun 2023.
[48] 龙女之声. “伪装答辩 肛门加速 极限反击 保住性命需要多少技巧?” Bilibili, 13 May 2023, http://www.bilibili.com/video/BV1Z24y1N7TN. Accessed 4 Jun 2023.
[49] 赵泠. “有哪些进化出同归于尽能力的生物?如果没有这种生物,为什么不会进化出同归于尽这样的能力?” 知乎, 20 Mar 2023, http://www.zhihu.com/question/27036464/answer/76064118. Accessed 4 Jun 2023.
[50] Yamanoue, Yusuku, et al. “Multiple Invasions into Freshwater by Pufferfishes (Teleostei: Tetraodontidae): A Mitogenomic Perspective.” Plos One, vol. 6, no. 2, 25 Feb 2011, pp. 1-13. Accessed 4 Jun 2023.
[51] Guo, Baocheng, et al. “Genome Size Evolution in Pufferfish: An Insight from BAC Clone-based Diodon holocanthus Genome Sequencing.” BMC Genomics, vol. 11, no. 396, 23 Jun 2010, pp. 1-15. Accessed 4 Jun 2023.
[52] Brainerd, Elizabeth, et al. “Patterns of Genome-size Evolution in Tetraodontiform Fishes.” Evolution, vol. 55, no. 11, Nov 2001, pp. 2363-2368. Accessed 4 Jun 2023.
[53] Imai, Shuichiro, et al. “The Genome Size Evolution of Medaka (Oryzia latipes) and Fugu (Takifugu rubripes).” Genes Genet. Syst., vol. 82, 29 Jan 2007, pp. 135-144. Accessed 4 Jun 2023.
Title: Meeesh. “Just in Case You’ve Never Seen a Baby Pufferfish Before.” 9GAG, 21 Jan 2015, http://9gag.com/gag/anXpNZV. Accessed 4 Jun 2023.